
Abstract
Although upper extremity functional recovery is a high priority for spinal cord injured patients with cervical injuries, few injury models have been developed in mice with sustained deficits in forelimb motor function. Here, we characterize a dorsolateral funiculus (DLF) crush model in mice, which ablates the rubrospinal tract (RST) unilaterally and thus allows correlation of motor recovery to axonal regeneration in the assessment of molecular regeneration targets. We conducted unilateral DLF crush injuries at cervical levels C4 and C6 and assessed motor recovery in a battery of tests: the rearing test of forelimb asymmetry, the grooming test, staircase pellet reaching, a horizontal ladder task, and CatWalk gait analysis. All tasks revealed lesion effects on forepaw function when DLF crush was instigated at level C4, but deficits were generally only transient in mice with DLF crush performed at level C6. Anterograde tracing of the RST with biotinylated dextran amine revealed the tract's complete ablation. The characterization of a C4 DLF model in mice provides an important tool for assessing molecular regeneration targets to promote functional recovery after spinal cord injury.
Introduction
R
Here, we performed a cervical dorsolateral funiculus crush to sever the RST in adult mice and assess functional impairments in five behavioral paradigms: the cylinder rearing test of forelimb preference and usage, the grooming test, which assesses forelimb range of motion during spontaneous grooming behavior, 15,16 CatWalk gait analysis, which assesses gait and motor parameters during continuous locomotion along a walkway, 17 staircase pellet reaching, which assesses forelimb skilled reaching, 18,19 and horizontal ladder walking. 20 We sought to determine the extent to which DLF lesions result in long-term deficits when implemented at cervical levels C4 or C6, and, therefore, the appropriateness of the injury at these levels in permitting the assessment of the extent to which RST regeneration correlates with motor recovery. Animals sustained a DLF crush lesion at C4 or C6, and behavior was evaluated over a 4 week recovery period. C4 DLF crush caused sustained deficits in ipsilateral forelimb skilled movement, reaching, and preference, whereas C6 DLF crush led to only transient deficits. This C4 DLF crush model should facilitate the assessment of molecular targets to promote axonal regeneration in promoting upper extremity recovery following SCI.
Methods
All procedures were approved by the Animal Care Committee of the University of British Columbia in accordance with the guidelines of the Canadian Council for Animal Care.
DLF crush
A total of 28 adult female C57Bl6 mice (Charles River Laboratories, Wilmington, MA) were used in this study. Animals were anesthetized with isofluorane. The skin in the neck region was shaved and disinfected, and animals were given a subcutaneous injection of lactated Ringer's solution (Hospira Inc., Lake Forest, IL) and buprenorphine (0.03 mg/kg). Once a surgical plane of anaesthesia was reached, a hemilaminectomy was performed to expose the left half of either the fourth cervical spinal cord segment (C4; n=20; including an n=5 of sham operated mice which received no crush injury) or the sixth cervical spinal cord segment (C6; n=8), and the dura mater overlying the gray matter was pierced with a 26 gauge needle. Fine tipped Dumont #5 forceps were modified for this purpose by grinding their blades to a width of ∼200 μm and placing a mark at 1 mm distance from the tip. One prong of these fine-tipped forceps was inserted into the dorsal horn gray matter to a depth of ∼1 mm (using the mark), whereas the other prong remained outside the spinal cord, that is, lateral to the DLF. The forceps were closed and held for 15 sec to crush the DLF, including the RST; the forceps was removed and inserted again in order to repeat this crush once. The muscles and skin were then closed with sutures, and the animals were permitted to recover in a heated incubator (35°C) for 2–3 h before returning to their home cage. Animal weights were recorded daily for 1 week post-surgery, and twice a week thereafter. Bladder function was also assessed for 7 days post-surgery, but every animal retained bladder function postoperatively and no expressions were necessary.
Behavioral assessments
Animals were kept on a 12 h:12 h light:dark cycle (lights off at 9:00 a.m.). On days 2, 7, 14, 21, and 28 postoperatively, mice were assessed in the pellet reaching and rearing tasks between the hours of 10:00 a.m. and 1:00 p.m. inclusive, and in the CatWalk analysis between 4:00 p.m. and 7:00 p.m. inclusive. On days 3, 8, 15, 22, and 29 postoperatively, mice were assessed in the grooming task between the hours of 10:00 a.m. and 12:00 noon inclusive, and in the horizontal ladder task between 4:00 p.m. and 6:00 p.m. inclusive. All behavioural analyses were conducted by personnel blind to group inclusion.
Rearing test
The rearing test assesses forelimb preference during spontaneous vertical exploration and was previously developed for use in rats 21 –25 but has been modified for use in mice. 26 –28 Like rats, mice, when placed in a cylinder, spontaneously rear using either one or both forepaws for weight support against its wall. Forelimb preference for weight support against the wall is thought to be indicative of forelimb functionality. Mice were placed in a clear plexiglass cylinder (20 cm in height, 15 cm in diameter) for 15 min. Two mirrors were placed at an angle such that forepaws and rearing forelimb preferences could be viewed irrespective of mouse orientation. The testing session was videotaped, and a blinded experimenter scored initial and subsequent forepaw use for a total of 10 rearing sessions (characterized by the initial placement and any subsequent placements the animal did until placing a paw back down on the ground). Initial placements were scored as either “left” or “right” when the left or right forepaw was used for weight support for>0.25 sec independently of the other forepaw, respectively. Otherwise, in cases where both forepaws were used within 0.25 sec of each other for weight support, a score of both was given. After scoring a “both,” every subsequent combination of two limb movements was described as a subsequent “both” unless both paws were removed from the cylinder and only one paw was re-placed, which would be described as a left or right forepaw subsequent placement. Prior to surgery, mice were placed in the cylinder for three sessions for habituation and to obtain pre-injury scores. Mice were then tested on days 2, 7, 14, 21, and 28 post-injury. Percent use of the ipsilateral limb, contralateral limb, and both limbs together for initial rears was calculated as a fraction of the total number of rears (=10) for each animal at each time point. Percent ipsilateral limb use was calculated based on the number of times the ipsilateral limb was used for both initial and subsequent rearing events over the course of the first 20 initial and subsequent events for each animal at each time point.
Grooming test
Forelimb grooming function was assessed using a scoring system adapted from Gensel and colleagues for use in rats. 21 Cool saline was applied to the mouse's head and back with gauze, and the animal was placed in a cylinder with mirrors at angles, as described, for 15 min. The gauze was only damp with saline and not completely saturated, such that the mouse's fur was only moderately wet to induce a grooming response. The testing session was videotaped with a high definition camera (Sony, HDR-Hc1). Mice were tested preoperatively to obtain baseline scores, and on days 3, 8, 15, 22, and 29 post-surgery. At a later date, videos were watched frame by frame by an experimenter blinded to the treatment group of each mouse, who scored the maximum grooming score achieved by each paw independently according to the following scoring system: 0, the animal was unable to contact any part of the face or head; 1, the animal's forepaw touched the underside of the chin and/or the mouth area; 2, the animal's forepaw contacted the area between the nose and the eyes, but not the eyes; 3, the animal's forepaw contacted the eyes and the area up to, but not including, the front of the ears; 4, the animal's forepaw contacted the front but not the back of the ears; 5, the animal's forepaw contacted the area of the head behind the ears.
Horizontal ladder
Error (slip, miss, or drag) frequency was assessed for each limb of each mouse using a horizontal ladder modified as previously described. 20.29 Briefly, mice were videotaped with a high-definition camera (Sony, HDR-Hc1) as they crossed a 5.2 cm wide horizontal ladder with 31 rungs each spaced 1.3 cm apart. Each mouse repeated the task six times, and one repetition was excluded from analysis for each mouse based on failure to complete the task appropriately (by turning around midway through the sequence, rearing against the wall of the apparatus). In cases where all six runs were completed adequately, the sixth run was automatically excluded. Frame-by-frame video analysis provided scores for number of errors for each limb over each run, and summed for the total over the five runs for each animal at each time point to generate a limb-specific cumulative error score. Mice were tested preoperatively to obtain pre-injury scores, and on days 3, 8, 15, 22, and 29 postoperatively.
Pellet reaching
The staircase test of pellet reaching assesses skilled forepaw reaching and was initially developed for rats, 18,30 but a scaled-down assessment is in use for mice. 19,28 Mice were placed in a holding box with access to a plinth with a graded staircase on each side (eight steps per side). Two 20 mg chocolate food pellets (NOYES Precision Pellets, Research Diets Inc., USA) were placed in each well of each staircase such that mice could retrieve them by reaching with their forepaws down either side of the plinth. In this manner, pellets in the left staircase were only reachable by the left forelimb, and the right staircase by the right forelimb. Mice were familiarized with food pellets for 1 week by placing ∼100 into each home cage each day, and then pre-trained to complete the task itself by having each well baited with two pellets and placing each mouse within the apparatus for 15 min/day for 1 week. Mice were fasted for 18–20 h prior to each pre-training session, and prior to each testing session throughout the experiment. During assessment of preoperative scores and at each experimental time point, pellets were marked with food coloring to allow for the assessment of how far each mouse could reach by placing pellets of different colors at different well heights. Each mouse was granted access to the apparatus individually for 30 min for each experimental testing session. The total number of pellets eaten from each side was calculated at the end of each testing session for each animal. Pellets eaten from the top two wells on each side were discounted from analysis, as mice could retrieve these with the use of their tongues.
CatWalk gait analysis
The CatWalk apparatus and CatWalk XT 8.1 software (Noldus, The Netherlands) were used. Each animal was allowed to cross a transparent platform (5.2 cm wide and 60 cm long) with opaque and black walls in a darkened room. Animals were recorded from beneath with a high-definition camera (Sony, HDR-HC1) illuminated with a light source below the platform. The light internally reflects in the glass floor and illuminates the mouse's paws at the contact area, producing a footprint. Animals were trained to run across the runway consistently for 5 days prior to testing, with each training session consisting of three runs across the platform. Mice were then tested on days 2, 7, 14, 21, and 28 postoperatively. To ensure consistent performance of the task, mice were encouraged to run across the platform quickly. Additionally, mice were trained to run from a foreign cage back to their home cage, and inside their home cage chocolate pellets (NOYES Precision Pellets, Research Diets Inc., USA) were provided as a reward. At each experimental time point, three runs for each animal were recorded from below the glass by a digital video camera. Runs were then analyzed to measure stride length, swing duration, paw area, and paw intensity for each limb, in addition to base of support and speed in crossing the platform.
Anterograde tracing of rubrospinal axons
Two weeks before the end of the survival period and 30 days postoperatively, animals were anesthetized with isofluorane, the skin overlying the skull was shaved and disinfected, and animals were given a subcutaneous injection of lactated Ringer's solution (Hospira Inc., Lake Forest, IL) and buprenorphine (0.003 mg/kg). Upon the animal's reaching a surgical plane of anesthesia, a burr hole was drilled into the skull, and 1.0 μL of 10% biotinylated dextran amine (BDA; 10,000 kDa molecular mass, Molecular Probes, Eugene, OR) was stereotaxically injected into the vicinity of the right red nucleus at the following coordinates: 3.6 mm caudal to bregma, 0.7 mm lateral to the midline, and 3.6 mm depth with respect to the surface of the brain. The injection occurred at a rate of 100 nL/min, and the syringe remained in place for 5 min after the injection to prevent efflux. Mice were permitted to recover in a heated incubator (35°C) for 2–3 hours postoperatively before being returned to their home cages.
Immunohistochemistry
Two weeks after BDA injection, animals were killed with an overdose of chloral hydrate (100 mg/kg, i.p.) and perfused transcardially with phosphate-buffered saline (PBS) followed by phosphate-buffered 4% paraformaldehyde (pH 7.4). The cervical spinal cords were dissected, post-fixed in 4% paraformaldehyde overnight, and cryoprotected in 12%, 18%, and 24% sucrose in 0.1 M phosphate buffer over 3 days before being snap frozen in isopentane over dry ice. Cervical segments from C2 to C8 were cut into 20 μm longitudinal sections in the horizontal plane on a cryostat and stored at −80°C. Frozen sections were thawed for 1 h, rehydrated in 10 mM PBS for 10 min, and incubated with 10% normal donkey serum (in 0.1% Triton X-100) for 30 min to prevent nonspecific binding. Chicken anti-glial fibrillary acidic protein (GFAP) (1:1000; Chemicon) was applied overnight at room temperature. Secondary antibody (1:200, Jackson) raised in donkey and conjugated to DyLight 405 was applied for 2 h at room temperature. BDA was visualized by using Cy3-conjugated streptavidin (1:200, Jackson Laboratories) applied for 2 h at room temperature. Sections were cover-slipped in Fluoromount-G (Southern Biotech).
Image analysis and assessment of rubrospinal axon sparing
Digital images were captured with an Axioplan 2 microscope (Zeiss, Jena, Germany) equipped with Northern Eclipse software (Empix Imaging, Inc., Mississauga, ON, Canada) at 10×. GFAP+ staining was used to identify the center of the lesion, and images of the lesion center were taken in every section containing BDA+ labelling of the RST on every other slide, encompassing the entirety of the tract as well as it could be observed by the tracer. Images were then assessed for sparing by drawing a line through the middle of the lesion and noting the propensity of BDA+ axons to make it into or through the middle of the lesion site on every image captured. All histological analyses were conducted by personnel blinded to the experimental grouping.
Eriochrome Cyanine R (ECR) staining
Assessment of white and gray matter sparing was conducted by staining coronal sections through the injury site with ECR to visualize myelinated white matter. Sections were rehydrated through two toluene solutions followed by graded ethanol solutions: twice in 100%, once in 95%, once in 70%, and one in 50% ethanol. After rehydration in distilled water, slides were stained in ECR solution (0.16% ECR, 0.4% sulphuric acid, and 0.4% ferric chloride) at room temperature for 10 min, and gently rinsed in distilled water. Sections were then differentiated in 0.5% ferric ammonium sulphate at room temperature for ∼30 sec, rinsed in distilled water, and then stained in 1% neutral red solution for 2 min before being dehydrated in graded ethanol solutions: once in 50%, once in 70%, once in 95%, and twice in 100%. Sections were then cleared in xylene two times and cover-slipped using Entallan mounting medium (EM Science, Gibbstown, NJ). Each section was digitally photographed (Zeiss AxioPlan2 imaging microscope; Zeiss, Inc., Thronwood, NY) and manually traced using imaging analysis software (Sigma Scan Pro 5, Systat Software Inc., San Jose, CA). White matter sparing was based on positive (blue) staining of myelin via ECR, whereas gray matter sparing was based on normal cytoarchitecture of gray matter relying on ECR staining and neutral red counterstaining.
Statistical analysis
All statistical tests were conducted with Sigma Stat 3.0.1 (SPSS, Inc.). All data are presented as mean±standard error of the mean (SEM). The grooming test, rearing test of forelimb asymmetry, horizontal ladder, and pellet reaching were analyzed using Mann–Whitney U tests to compare groups at individual time points and the Wilcoxon test for paired samples to compare the same group at different time points. CatWalk Gait Analysis was analyzed using a two way repeated measures (RM) analysis of variance (ANOVA) (groups vs. days) and follow-up Student's t tests. Gray and white matter sparing histological data were analyzed using one way ANOVA, and group differences were ascertained using Tukey's post-hoc comparisons. The significance level for all tests was set at p<0.05.
Results
Basic characterization of adult mouse cervical DLF crush SCI
We endeavored to develop a unilateral RST injury model that would lead to sustained deficits in forelimb function and to ascertaining the level of the cervical spinal cord at which such an injury would lead to functional deficits in forelimb motor function. To do so, DLF crush was employed (Fig 1A). Following hemilaminectomy, the dura was pierced using a 26 gauge needle, and one prong of the forceps was inserted ∼1 mm into the gray matter of the spinal cord and the other lateral to the cord as shown. Modified #5 Dumont forceps that were thinned to a prong width of ∼200 μm were used for this purpose (Fig 1B).

Basic characterization of dorsolateral funiculus (DLF) crush lesion.
Following either C4 or C6 DLF crush, mice displayed a closed forepaw on the injured side and uncoordinated overground locomotion. The mice were able to freely locomote around their cages and no observable deficit was observed in the forelimb on the contralateral side. No change in body weight was observed following C4 or C6 DLF crush at any time point postoperatively compared with preoperatively or between C4 DLF crush, C6 DLF crush, and sham injured mice (Fig 1C). Additionally, although typical animal monitoring within our laboratory requires that we check at least daily for the 1st week postoperatively, no manual expression of the bladder was required.
Assessment of gray and white matter sparing in and around the lesion site
To visualize the rostral/caudal and dorsal/ventral extent of damage following DLF crush, coronal spinal cord sections of C4 DLF crushed (n=5) and control sham operated (n=5) mice were stained for ECR/neutral red to visualize myelin 6 weeks postoperatively. Tissue damage in the white and gray matter following C4 DLF crush was predominantly confined to a 200 μm area surrounding the lesion epicenter (Fig. 2). Importantly, no damage to the dorsal columns (and, therefore, the main contingent of corticospinal axons) was observed, nor was damage observed on the side of the spinal cord contralateral to the injury.

Eriochrome cyanine (EC)/neutral red-stained spinal cord sections following C4 DLF crush. From top left (caudal) to bottom right (rostral), photomicrographs of representative spinal cord cross sections are stained with myelin dye eriochrome cyanine at the epicenter and at 100 μm increments rostral-caudal to the epicenter to a total distance of 800 μm. At 4 weeks postoperatively, EC-stained sections demonstrate ipsilateral white matter and gray matter damage with extensive damage of the dorsolateral funiculus, but complete sparing of the ipsilateral dorsal column, which contains the main contingent of corticospinal axons. No damage was observed on the side contralateral to the injury. Color image is available online at
We quantified spared white and gray matter area at 100 μm intervals rostral and caudal to the lesion epicenter for a total distance of 800 μm on both the contralateral and ipsilateral side of the spinal cord with respect to the injury site (Fig. 3). At 100 μm rostral and caudal to the lesion epicenter and at the epicenter itself, there was a significant difference in the percentage of gray matter spared and white matter spared following C4 DLF crush compared with sham operated mice (Tukey's post-hoc test; p<0.05). At the lesion epicenter, there was 43.98±3.18% and 35.26±4.77% sparing of white and gray matter, respectively.
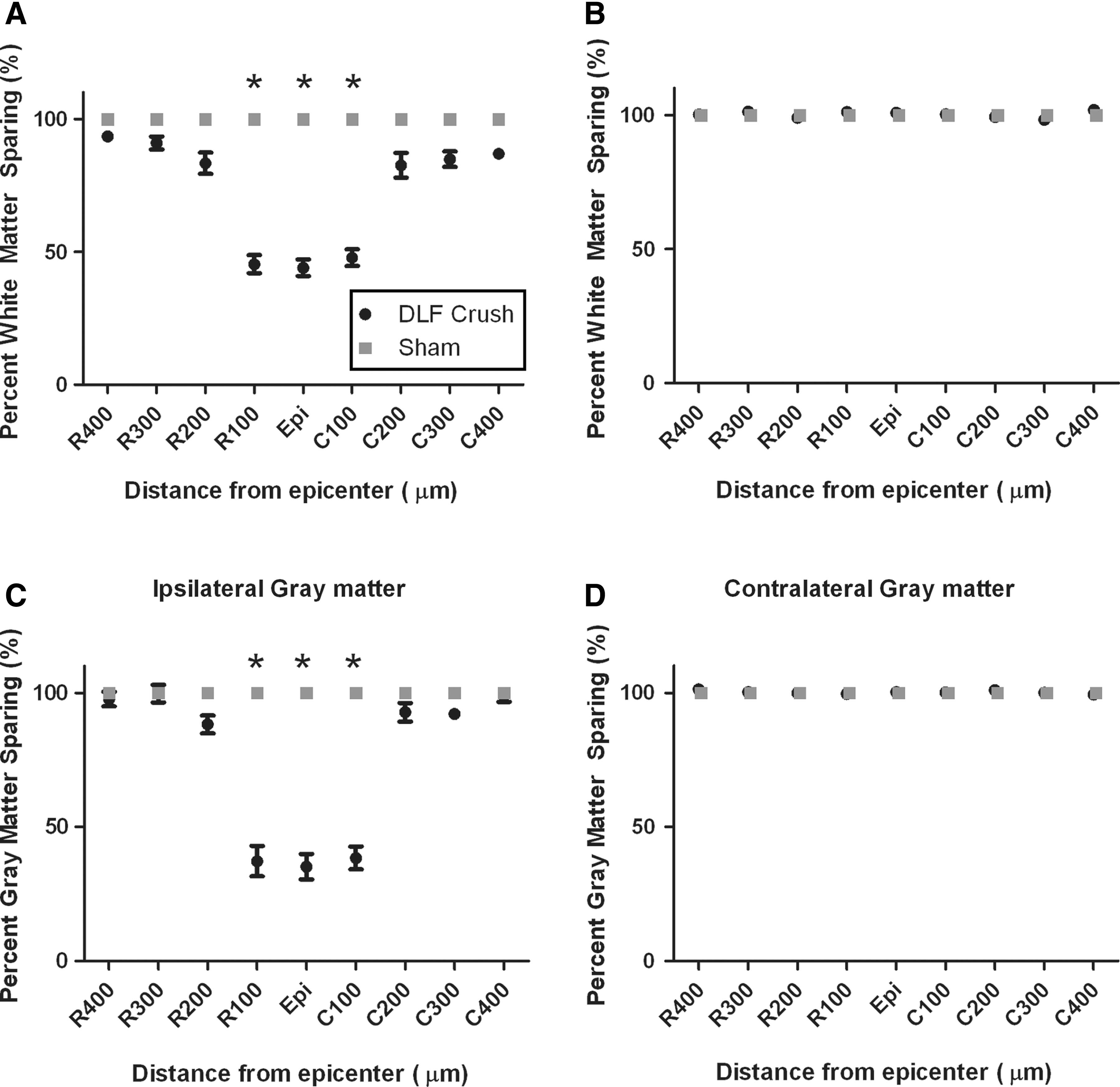
Tissue sparing quantification of spinal cord tissue following C4 dorsolateral funiculus (DLF) crush or sham operation. Measurement of the rostral-caudal extent of ipsilateral
Assessment of rubrospinal axotomy following DLF crush
To verify complete ablation of the RST, we injected BDA into the vicinity of the right red nucleus following the conclusion of behavioral assessments, and 2 weeks prior to euthanizing a cohort of n=18 mice. The extent of RST sparing was determined using immunostaining for GFAP to identify the lesion site, and BDA to visualize RST fibers (Fig. 4). We captured images on every section on which BDA+RST fibers could be identified on every other slide, encompassing the entirety of the tract as well as it could be observed based on anterograde tracing. In 346 images taken, RST fibers could be identified within the lesion site itself, but no RST fibers with morphology of spared axons 32 were observed within white matter caudal to the lesion site in any of the animals.
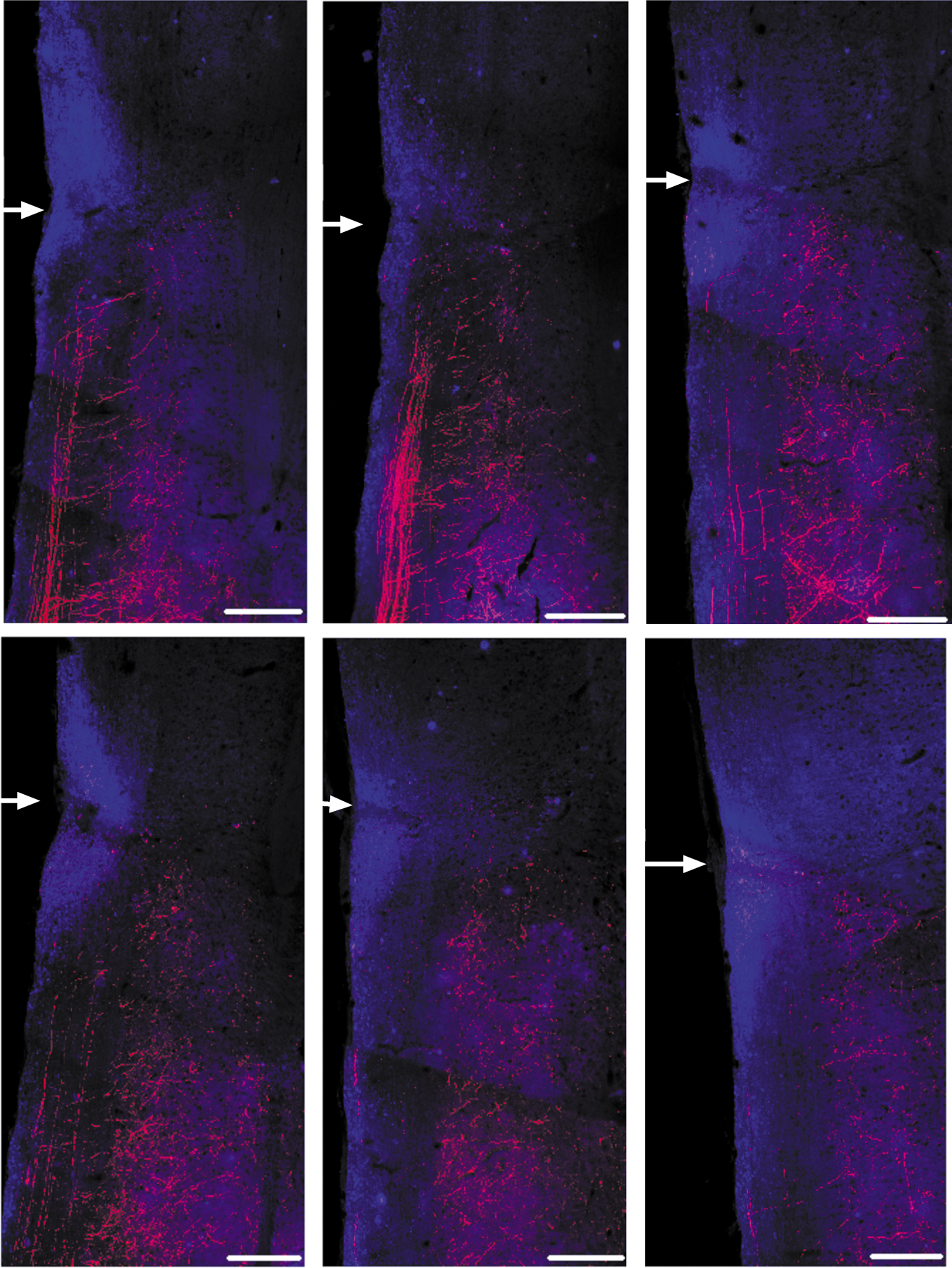
Assessment of rubrospinal axotomy following dorsolateral funiculus (DLF) crush via immunostaining for biotinylated dextran amine (BDA) (red)/ glial fibrillary acidic protein (GFAP) (blue) in longitudinal sections through the cervical spinal cord. Dorsal to ventral from top left to bottom right, images are spaced 80 μm apart, encompassing the main bundle of rubrospinal tract (RST) nerve fibers and their projections into gray matter in 420 μm total distance; 80 μm dorsal to the top left image or ventral to the bottom right image, only sparse labelling of BDA+axons in lateral funicular white matter is observed. In 346 images taken of every other slide onto which sections were collected and every section on each slide labelling positive for BDA within the lateral funiculus (thus encompassing the entirety of the RST as well as it could be observed by BDA+labelling), no BDA+fibers were observed making it through the lesion site or in caudal white matter. White arrows denote the middle of the lesion site. Scale bars are 100 μm. Color image is available online at
Rearing test of forelimb asymmetry
During preoperative baseline testing, our mice used their left forepaws alone for initial weight support against the wall in 37% (±7.89%) of the rears in the C4 injury group, and in 36.25% (±9.99%) of the rears in the C6 injury group. Somewhat surprisingly, the usage of their right forepaws for initial weight support was less frequent (25±5.82% in the C4 group and 23.75±8.% in the C6 group). Both limbs together were used in 38.0% (±7.27%) and 40.00% (±10.00%) of the C4 and C6 group respectively. After DCL crush at C4, a sustained change in the pattern of forelimb use for initial weight support was observed (Fig. 5 A–C). The percentage of initial placements made with the contralateral forepaw significantly increased (Fig. 5B), whereas the percentage of initial placements made with either the ipsilateral forelimb or both forelimbs together significantly decreased at all time points post-injury compared with preoperatively (p<0.05) (Fig. 5 A,C). At 28 days post-injury, mice with a C4 DLF crush lesion used their left/ipsilateral forelimbs alone in 15% (±4.77%), their right/contralateral forelimb alone in 72% (±6.63%), and both forelimbs together in 14.0% (±3.40%) of the initial paw placements against the cylinder wall.

Effect of C4 and C6 dorsolateral funiculus (DLF) crush on the rearing test.
Following C6 DLF crush, the percentage of initial placements made with the contralateral forepaw and both forepaws together were significantly lower than postoperatively at all time points, except at 28 days post-injury. The percentage of initial placements with the ipsilateral forepaw was significantly decreased at 2 days post-injury compared with the preoperative usage following C6 DLF crush, but not at any other time point (Fig 5 A). At 28 days post-injury, mice with a C6 DLF crush lesion used their left/ipsilateral forelimb alone 25% (±9.64%), the right/contralateral forelimb alone 47.5% (±11.30%), and both forelimbs together 27.5% (±6.20%) to make initial placements. There was a trend toward a difference in initial contralateral forepaw use between C4 and C6 at 28 days post-injury (p=0.098).
We also calculated percent ipsilateral limb use based on frequency of use during the first 20 initial and subsequent rearing events for each animal, and found a sustained decrease at each time point in comparison with baseline values in mice with a C4 DLF crush (p<0.05), but this difference in mice with a C6 DLF crush was only seen 2 days post-injury. Thus, C4 DLF crush lesions led to a decreased usage on the ipsilateral forelimb during rearing, whereas C6 DLF crush lesions led to only a transient decline in usage of the ipsilateral forelimb.
Grooming test
We adapted a grooming assessment used in rats for use in mice (Fig. 6). The mouse was gently scrubbed with gauze coated in saline until the fur was displaced, and recorded for 15 min in a cylinder with mirrors placed at angles to allow visualization of the mouse from every angle and assessment of grooming responses of the left and right forelimbs (Fig. 6A). Preoperatively, mice displayed normal grooming patterns, repeatedly licking their forepaws and grooming from their mouth and nose up to the eye region and to the back of the head. Some mice only reached to the front of the head over the 15 min testing period, obtaining a score of 4 in the absence of injury. A score of either 4 or 5 was also achieved by the contralateral limb of every mouse preoperatively and at every time point post-injury, and there was no significant difference in grooming scores between the contralateral limb of C4 crushed and C6 crushed mice at any time point. Following DLF crush at C4, there was a permanent ipsilateral deficit for the duration of the experimental period (p<0.05, Fig. 6B). None of the animals injured at C4 could touch the front of the ears, such that they could not achieve a grooming score>3. In contrast, the mice with C6 dorsolateral funiculus crush showed a deficit 2 days post-injury (p<0.05, Fig. 6B); however, by 7 days, there was no significant difference in grooming score compared with preoperative scores. Importantly, at 7 and 28 days post- injury, there was a significant difference in grooming score between C4 and C6 crushed mice (p<0.05). There was no difference in grooming score in the contralateral forelimb in either group at any time point (Fig. 6C). There was no difference in grooming score following sham operation at 2 days and 7 days postoperatively compared with preoperatively.
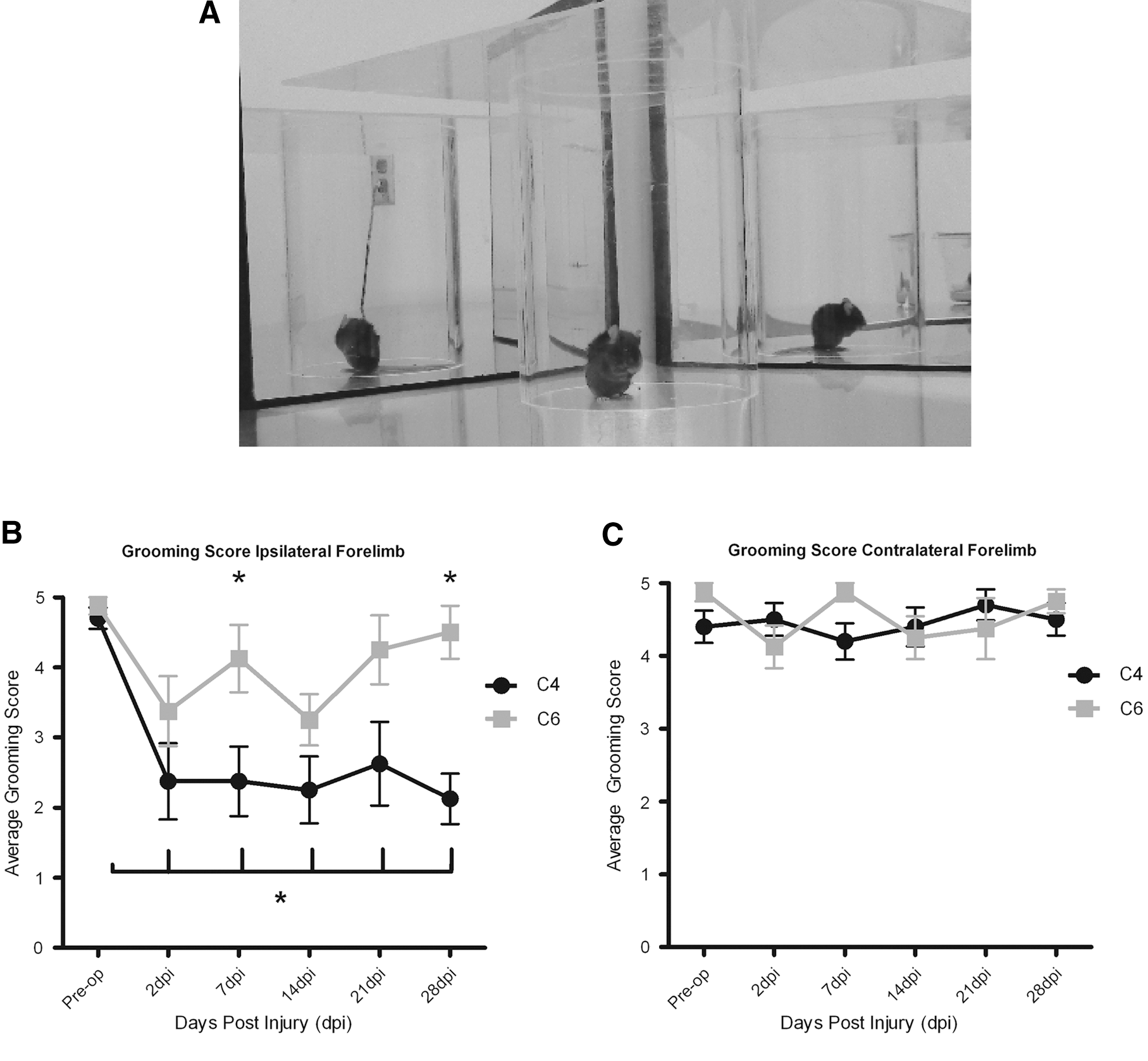
Grooming.
Pellet reaching
Preoperatively, mice were able to successfully reach, grasp, and eat four or five pellets with each of their forelimbs (Fig. 7 A,B). After C4 DLF crush, there was a sustained deficit in the ability to successfully reach, grasp, and eat pellets with the ipsilateral forepaw at all time points post-injury (p<0.05). Preoperatively, mice ate 5.1±0.43 pellets, whereas 28 days following C4 DLF crush, mice ate 2.0±0.20 pellets with the ipsilateral/left forelimb. In contrast, the apparent decline in the number of pellets eaten with the ipsilateral/left forelimb following C6 DLF crush did not reach significance (vs. preoperative values; p=0.148). At 28 days post-injury, fewer pellets were eaten by mice with a C4 DLF crush and those with a C6 DLF crush (2.0±0.20 pellets for C4 vs. 3.14±0.51 pellets for C6; p<0.05). The number of pellets eaten with the contralateral/right forelimb did not change at any time point post-injury and was similar in both groups. There was no difference in pellets eaten following sham operation at 2 days and 7 days postoperatively compared with preoperatively.

Staircase pellet reaching test.
Horizontal ladder
Preoperatively, the mice were able to cross the ladder with few errors, although with significantly more errors made by the forelimbs than with the hindlimbs across both groups (p<0.05). After C4 or C6 DLF crush, the cumulative error (CE) score (i.e., the number of errors committed by each limb over five trials runs) significantly increased for both the ipsilateral forelimb and hindlimb compared with preoperative values, and a sustained increase was observed throughout the testing period in both ipsilateral limbs (p<0.05; Fig. 8 A, B). No differences were observed for the CE score for the contralateral paws (Fig. 8 C, D), nor were differences observed in the CE scores of sham operated mice at 2 or 7 days postoperatively compared with preoperatively.
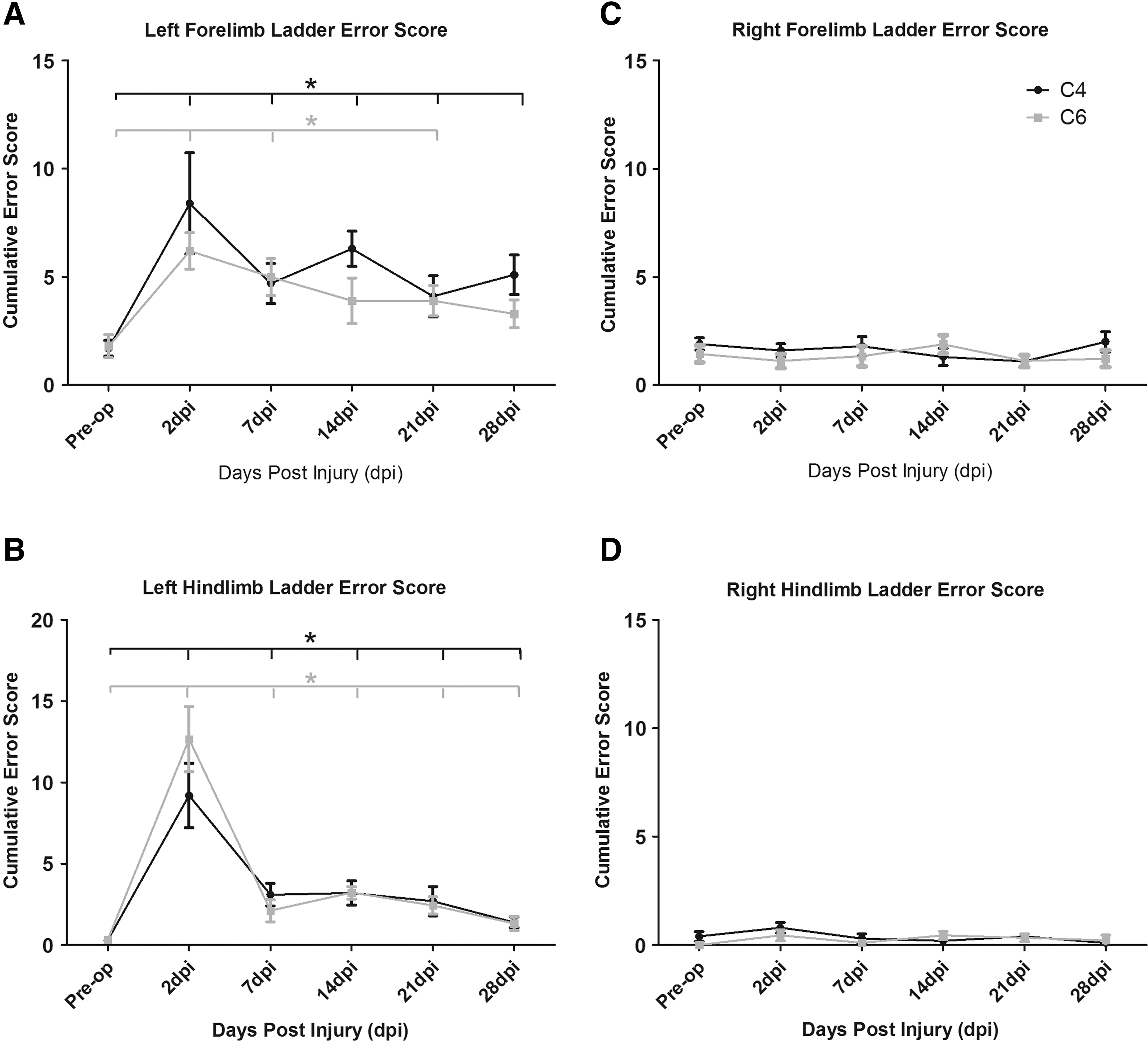
Horizontal ladder.
We also compared the ipsilateral versus contralateral CE scores to determine asymmetry between the limbs. After both C4 and C6 DLF crush, significant differences were observed between the CE scores on the left (ipsilateral) side versus the right (contralateral; Fig 8 C,D) side for both forelimbs and hindlimbs (p<0.05, data not shown).
CatWalk gait analysis
Footprint analysis revealed a robust reduction of stride length during locomotion along a walkway following DLF crush at C4, but not at C6, compared with preoperative stride lengths (Fig. 9). The length of strides taken by mice injured at C4 was significantly shorter than that by mice with C6 DLF crush. These differences in the stride lengths between C4 and C6 lesioned mice were observed in all four limbs, the ipslateral/left forelimb at 2, 7, 21, and 28 days post-injury (Fig. 9 A), the contralateral/right forelimb at 7, 14, 21, and 28 days post-injury (Fig. 9 C), the ipsilateral/left hindlimb at 2, 7, 21, and 28 days post injury (Fig. 9 B), and the contralateral/right hindlimb at 2, 7, 21, and 28 days post injury (Fig. 9 D (p<0.05 all two way RM ANOVA). Although mice with a C4 DLF crush and C6 DLF crush showed a significant difference compared with preoperatively in base of support, paw contact area, and paw intensity at 2 days post-injury (all p<0.05, data not shown), these deficits were transient, with no significant differences observed 7 days post-injury or later. There were no differences in swing speed at any time point (data not shown). In sham operated mice, there was no difference in any parameter 2 and 7 days postoperatively compared with preoperatively.
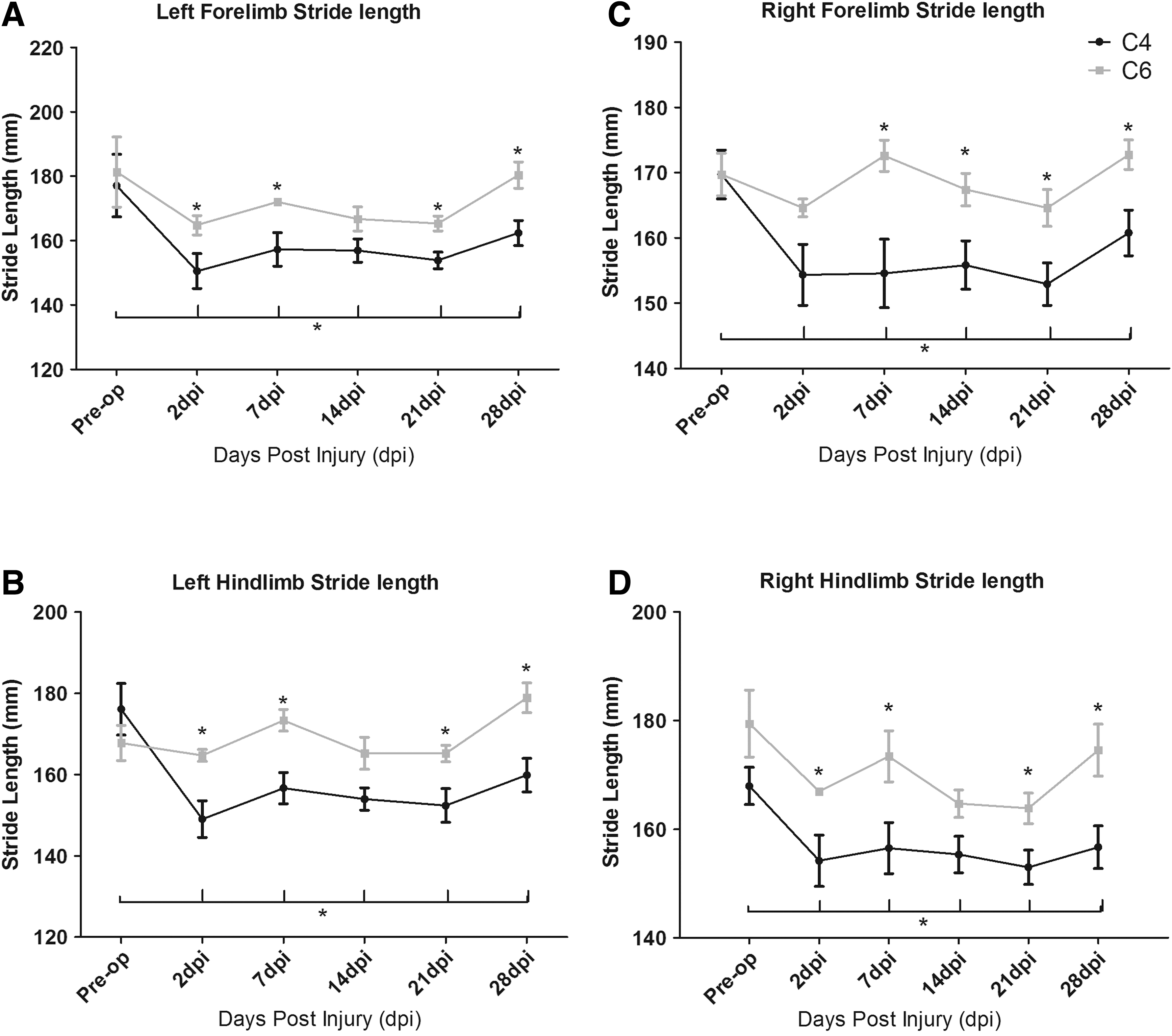
CatWalk automated gait analysis.
We also measured the speed at which the mice crossed the platform, as speed affects stride length. Although mice with a C4 DLF crush traveled more slowly at 2 days and 21 days post-injury compared with their preoperative speed, and mice with a C6 DLF crush traveled more slowly at 14 days post-injury compared with preoperatively (both p<0.05), there was no significant difference in speed between mice with C4 DLF crush and those with C6 DLF crush at any time point (data not shown). Therefore, the differences in stride length observed in mice with C4 DLF and those with C6 DLF cannot be explained by a difference in speed.
Discussion
In this study, we characterize a consistent and reliable DLF crush model in adult mice to axotomize the RST and compare the effects of this lesion when instigated at cervical level C4 or cervical level C6 in a number of tests that evaluate motor behavior, particularly forelimb function. We sought to determine the cervical level at which DLF lesioning would result in sustained deficits up to 4 weeks post-injury. This timeline would permit the assessment of the extent to which RST regeneration correlates with motor function improvement. Ablation of the RST is a useful model for SCI research, particularly studies focused on degeneration and regeneration, because of its anatomical convenience. The mouse and rat RST cross the midline in the ventral tegmental decussation and travel in the ventrolateral hindbrain before descending in the spinal cord within the DLF. 6,9 Because the vast majority of the tract is crossed, it can be reliably transected by DLF lesions, allowing assessment of regenerative outcome following SCI in the absence of sparing. 31 Additionally, because the RST arises from the red nucleus, a relatively dense neuronal cluster found within the midbrain, assessment of the cell body response to axotomy is achievable with comparisons to the relatively uninjured red nucleus of the contralateral tract. 11,23
Our results in the pellet reaching and horizontal ladder tasks indicate that the DLF of the mouse is particularly important in skilled forelimb movement. Much of this deficit is likely caused by disruption of the RST. Double staining analysis has previously indicated that rubrospinal axons make direct synaptic contact with motor neurons innervating distal muscles of the forelimb, which is consistent with the RST's role in skilled forelimb movement in the rat. 33 More recent evidence suggests that similar connections exist in the mouse. 9 This importance of the RST in skilled reaching has further been documented in the rat using ibotenic acid lesions to the red nucleus, which interfere with aiming, pronation, arpeggio, and supination of the limb. 34,35 In particular, DLF lesioning indicates the RST's importance for arpeggio movement; that is, pronation of the paw and positioning of the digits with wrist movement when grasping for food. 36 We found a significant reduction in the number of pellets eaten in the staircase pellet test with the ipsilesional forepaw after C4 dorsolateral crush. In the cat, Alstermark and colleagues have shown that ablation of the DLF at C5/C6 resulted in complete loss of food-taking ability. 37 Indeed, if the C5 DLF lesion was incomplete and as little as 20% of the RST was left intact, food taking ability recovery was faster and more complete. Interestingly, unilateral ablation of the corticospinal tract (CST) via pyramidotomy had no significant effect on the ability of mice to grasp and eat food pellets from the wells of a graded staircase, 28 suggesting that the circuitry encompassed within the DLF of the mouse is more important in this regard or at the very least able to compensate for the loss of the CST.
Our results also indicate that the DLF is important in spontaneous limb use. DLF ablation resulted in significant changes in patterns of weight-bearing support during spontaneous vertical exploration and grooming. In the rat, C3/C4 DLF lesioning leads to a sustained decline in use of the ipsilateral limb in spontaneous vertical exploration, whereas transplantation of brain derived neurotrophic factor (BDNF) expressing fibroblasts aiming at the promotion of RST regeneration leads to a significant increase in ipsilateral forelimb use. 23 Consistent with these results, Webb and Muir found that 88% of rubrospinal injured rats had decreased usage of the ipsilateral forelimb in the same task. 38 In the mouse, it appears that the DLF and the CST share a role in forepaw use in weight-bearing support, as unilateral pyramidotomy results in a similar sustained impact on the forelimb up to 28 days post-injury in the same task. 28 Additional pathways and cells involved in forelimb and hindlimb locomotion would be damaged by lateral funicular lesions, and the behavioral results observed here cannot be construed as arising from ablation of the RST alone. Dorsolateral corticospinal and lateral reticulospinal fibers contribute to motor function in the mouse, and would be damaged in this model. Because our lesions included a significant extent of gray matter, the lesion area included premotor interneurons, propriospinal neurons, and, possibly, motor neurons that form the segmental circuitry underlying forelimb movement. 39 In rats, retrograde tracing has shown that motor neurons supplying digital flexors are located in pools extending from C5 through T1. 40 Destruction of such segmental circuitry would be characterized by the symptoms of lower motor neuron injuries, including flaccid paralysis and loss of segmental reflexes. However, it is unlikely that motor neuron loss was a significant factor in the sustained deficits observed, as we were careful to avoid crushing the ventral horn and the rostrocaudal extent of our lesion was relatively small, as the forceps were only ∼200 μm in width; compared with the extent of the motor neuron pools over several segments.
DLF crush lesioning at either C4 or C6 had only transient effects on several parameters of overground locomotion determined via CatWalk gait analysis, including base of support, paw contact area, and paw intensity, whereas swing duration was not significantly different at any time point. Instead, the only difference observed was a decrease in stride length of both hindlimbs and forelimbs on both the ipsilateral and contralateral sides of the lesion following C4 DLF crush. These findings suggest that the role of DLF circuitry in overground locomotion is well compensated for following injury. Previous work in the rat has found only minor deficits in overground locomotion following red nucleus lesioning. 41 There is a high degree of correlation between stepping ability and spared white matter tissue in the region of the RST in the rat, indicating that the ventrolateral funiculus is more important for general locomotion than the DLF. 21,42 Notably, unilateral pyramidotomy in the mouse also leads to a bilateral decrease in stride length in both forelimbs and hindlimbs but has no effect on other parameters in CatWalk gait analysis, 28 suggesting a similar function of the CST and DLF in overground locomotion. Although overground locomotion was not overtly effected by DLF lesioning, our results in the horizontal ladder task are indicative of a role of this circuitry in hindlimb locomotion. More specifically, the increase in cumulative error score in the left hindpaw on this task following C4 DLF lesioning suggests that the RST may play a role in sensorimotor integration, which is required for successful placement of the hindpaw onto the ladder beam.
Assays that use animal behavior to ascertain locomotor function have an inherent variability, and every effort should be made to minimize this variability whenever possible. We recommend at least 1 week of daily animal handling and environmental enrichment for successful implementation of this behavioral model, acknowledging that week-to-week variability in the outcome of any particular behavioral test can occur. It is for this reason that the assessment of animals across a battery of behavioral tests (instead of only one) is also recommended.
Conclusion
In conclusion, we have characterized a DLF crush model in adult mice that harbors several significant advantages in the assessment of potential treatment options and motor recovery following SCI. Because this model was developed in mice, it is of particular use in the assessment of molecular therapeutic targets reliant on transgenic mouse technologies. As a DLF lesion results in complete ablation of the RST, it permits the assessment of axonal regeneration and functional recovery without severe deficits in bladder or respiratory function, keeping animal care demands low. Finally, and in particular, because the cervical region of the spinal cord is the most common injury area in cases of human SCI, 43 this DLF crush injury leads to motor function loss in an area of special importance to SCI patients.
Footnotes
Acknowledgments
This work was supported by funding from the Wings for Life Foundation. We thank Dr. Femke Streijger for initial help with behavioral testing, and members of the Tetzlaff Lab for constructive feedback on the data. Dr. Hilton is supported by a Canadian Institutes of Health Research (CIHR) Transplantation Training Program Award. Dr. Assinck is supported by a CIHR Doctoral Award. Dr. Duncan is supported by a Multiple Sclerosis Society of Canada Doctoral Award. Dr. Tetzlaff holds the Rick Hansen Man in Motion Chair in Spinal Cord Injury Research (Rick Hansen Foundation).
Author Disclosure Statement
No competing financial interests exist.