
Abstract
Spasticity is a major health problem for patients with traumatic brain injury (TBI). In addition to spasticity, TBI patients exhibit enduring cognitive, balance, and other motor impairments. Although the use of antispastic medications, particularly ITB, can decrease the severity of TBI-induced spasticity, current guidelines preclude the use of ITB during the first year after TBI. Therefore, the present study was performed to quantitate disability in an animal model of closed-head TBI (cTBI; Mararou's model) after ITB treatment. After cTBI, significant deficits in spasticity and gait, cognitive, balance, and anxiety-like behaviors were detected. ITB (Lioresal®) or saline was administered using Alzet pumps (0.8 μg/hour for 4 weeks). Spasticity measures using velocity-dependent ankle torque and ankle extensor muscle electromyography recordings, footprints (gait), balance performance tests, serial learning, and anxiety-like behaviors were performed at multiple post-treatment and -withdrawal of ITB time points. Our data indicated that 1 month of ITB treatment initiated at post-TBI week 1 blocked the early onset of spasticity and significantly attenuated late-onset spasticity and anxiety-like behavior with no significant adverse effects on cognitive and balance performance. This improved spasticity outcome was accompanied by marked up-regulation of gamma-aminobutyric acid (GABA)/GABAb, norepinephrine, and brain-derived neurotrophic factor expression in spinal cord tissue. Early intervention with ITB treatment was safe, feasible, and effective in this cTBI animal model. Collectively, these data provide a strong molecular footprint of enhanced expression of reflex regulation by presynaptic inhibition. The possibility that acute ITB treatment may decrease maladaptive segmental and descending plasticity is discussed. The data provided by the present animal model initiates a pre-clinical platform for safety, feasibility, and efficacy of early ITB intervention after TBI.
Introduction
T
In this regard, development of an experimental animal model of TBI spasticity could provide an opportunity to gain a greater understanding of the neurobiology of TBI-induced spasticity and provide a platform for quantitative evaluation of the safety, feasibility, and efficacy of intervention strategies. We developed an animal model of TBI-induced spasticity using both open- and closed-head TBIs. 14 These studies utilized longitudinal measures of lower-limb joint torques to quantitate longitudinal patterns of spastic hypertonia. 14 –16 The present study was performed to evaluate spasticity, cognitive, vestibulomotor, and locomotor disabilities produced by closed-head TBI (cTBI) and how these disabilities were modified by Lioresal ITB therapy initiated in an acute setting. The potential influence of TBI and treatment on the excitability of voluntary motor pathways was assessed using transcranial magnetic stimulation (TMS) to activate constituents of the executive motor pathway to lower-limb muscles. In addition, spinal cord tissues were studied to assess the expression of immunohistochemical (IHC) markers for agents known to influence the excitability of neural pathways controlling lower-limb motor function, especially spasticity.
Methods
The present studies utilized a clinically relevant laboratory model of cTBI that produces enduring spasticity, cognitive, and vestibulomotor disabilities. A total of 42 young-adult Sprague-Dawley rats (Charles River Laboratories Inc., Wilmington, MA), weighing 260–285 g, were used in these studies. Animals were anesthetized using a combination of ketamine (80 mg/kg) and xylazine (5 mg/kg). Buprenex (0.05 mg/kg) and ketoprofen (5 mg/kg) were used as analgesics during surgery and 1 day postinjury.
Surgical approach for cTBI
This procedure used a standardized weight-drop method to produce controlled levels of TBI, as described previously. 17,18 This method reproduced many of the injury patterns observed in humans after noninvasive TBI. The animal's body temperature was monitored and maintained at 37–39°C using a clean, sanitized, temperature-controlled blanket. The animal and the surrounding surgery field were covered with a sterile drape. cTBI procedures were conducted under aseptic surgical techniques. The dorsal surface of the head was shaved and the incision site was infiltrated with 0.25% bupivacaine (3 mg/kg). The surgical area was then cleaned with repeating cycles of betadine scrub (7.5%), followed by a 70% alcohol wipe and a final application of betadine solution (5.0%). A skin incision was made along the dorsal midline of the cranium. Fascia and soft tissue were retracted to expose the landmarks of the lambda and bregma on the cranium. A stainless disk (10 mm diameter×2 mm height) was attached to the exposed skull with dental acrylic. The animal was then placed on a foam pad, and the cortical impact trauma device was maneuvered over the animal and aligned with the stainless disk. A 450-g impactor, with a 6-mm-diameter tip, was dropped through a Plexiglas tube from a 1.5-m height. The impactor struck the stainless disk affixed to the cranium of the animal to produce moderate-to-severe cTBI. The stainless disk served to diffuse the force of the impactor, thus decreasing the probability of skull fracture. Further, the elastic properties of the foam pad, on which the head was resting, produced a controlled acceleration/rotation closed-head diffuse brain trauma injury. 17 After injury, the stainless disk was removed from the cranium and the skin was closed with sterile sutures.
Osmotic pump implant for ITB delivery
Osmotic pumps were implanted and the intrathecal catheters were placed in the L2 segment in all rats, according to our previous report, at postinjury day 7. 19 Briefly, animals were anesthetized with a subcutaneous (s.c.) injection of xylazine (6.7 mg/kg), followed by intraperitoneal ketamine (100 mg/kg)/glycopyrrolate. 20 –22 Before implant, osmotic pumps (model 2004, 0.25 uL/h; Azla Corp., Palo Alto, CA) were filled with baclofen injection (Lioresal intrathecal) or 0.9% physiological saline. A 3.2-μg/μL Lioresal intrathecal dose, which resulted in a total baclofen delivery of 0.8 μg/h for continuous intrathecal delivery of 4 weeks, was used. This dose is based upon receptor saturation, plasticity, and down-regulation studies by Malcangio and Bowery, 23 and Kroin and colleagues, 24 our previous reflex excitability studies, 19 as well as our pilot dose-response studies in this model (data not shown). Osmotic pumps were incubated overnight in 0.9% saline at 37°C in sterile conditions for immediate delivery of drug or vehicle. Particular efforts were taken to avoid introducing an air space within the infusion catheter. A 4-cm vertical skin incision was made spanning the thoracolumbar juncture, and an s.c. pocket was dissected to accommodate the osmotic pump. Musculature covering spinal processes was retracted from T12-L1, and a T13-L1 laminectomy was performed. The osmotic pump was maneuvered into the s.c. pocket and secured to the surrounding musculature using 6.0 silk sutures. The infusion catheter was tunneled through the rostral bank of dissected musculature and secured by polyacrylamide adhesion (Vetbond™ tissue adhesive; 3M, St. Paul, MN) to an exposed spinal process. Wound margins of muscle and skin were infiltrated with a long-lasting local anesthetic (lidocaine). With the aid of a dissecting microscope, the dura mater was cut and the silastic tubing inserted into the subarachnoid space of the lumbar enlargement. Excess tubing was secured to the surrounding musculature with 6.0 silk sutures. Muscle layers were closed with an absorbable suture (Dexon II; Sherwood, Davis & Geck, Wayne, NJ), and the skin was closed using stainless steel wound clips (Autoclip®; Becton Dickenson, Sparks, MD). Postinjury care included a 5-mL s.c. injection of warmed (37°C) 0.9% physiological saline and overnight assistance with body-temperature regulation using a temperature-controlled heating pad. The pump was removed after 4 weeks of drug delivery by similar procedures, and the infusion catheter was ligated with a 6.0 silk suture and left in situ to avoid catheter-removal–related complications, which include possible leakage of cerebrospinal fluid (CSF) and damage to the cord during surgery.
Behavioral and physiological experimental methodology
Spasticity (motor disability) testing: velocity-dependent ankle torques and electromyography (EMG)
recording. Details regarding instrumentation, animal set-up, and recording procedures have been previously reported. 19,25,26 Rats were immobilized in a custom-designed trunk restraint, without trauma or apparent agitation. All recordings were performed in awake animals. The proximal portion of the hind limbs to the mid-shank was secured in a form-fitted cast that immobilizes the limb while permitting normal range of ankle rotation (60–160 degrees). The lengthening resistance of the triceps surae muscles was measured indirectly by quantitating ankle torque during 12.24-degree dorsiflexion rotations of the ankle from 95 through 83 degrees. Contact with the foot was achieved using a form-fitted cradle aligned with the dorsal edge of the central footpad 2.6 cm distal to the ankle joint. Ankle torque (T) was assessed as the product of gram force (F; 980.7 dynes=1 gf ), the length of moment (torque) arm (l) (2.6 cm distance from ankle joint to stirrup contact point), and sin function of displacement force vector, for example, T=F×l×sin θ (see previous studies 19,25,26 for details of this measure). The angle of contact between the displacement shaft and the moment arm is 95 degrees. The neural activity of the triceps surae muscle was measured using transcutaneous EMG electrodes. The electrode was inserted in a skin fold over the distal soleus muscle just proximal to the aponeurotic convergence of the medial and lateral gastrocnemii into the tendonocalcaneousus. The reference electrode was placed in a skin fold over the greater trochanter. A xylocaine 2% jelly (lidocaine HCl; Astra USA, Inc., Westborough, MA) was applied over the electrode insertion points to minimize pain during recording. A topical antibiotic ointment (a combination of bacitracin, neomycin and polymyxin B; Fougera Altana Inc., Melville, NY) was also applied on these areas after removing the electrodes at the end of each trial to reduce the chance of infection. Before each trial, the urinary bladder was voided to minimize visceral-somatic influence. Controlled dorsiflexion was achieved through the use of an electromechanical shaker (model 405; Ling Dynamic Systems, Royston Herts, UK). A force transducer (LVDT; model FT-03; Grass Instruments, Quincy, MA) was mounted on an axel and configured for recording using red/black spring tension to provide a recording range of 10 mg to 200 g. The output shaft of the ling shaker was joined in series through the transducer to reach the foot cradle that contacts the central footpad. Therefore, because the transducer was free to move, its 1.1 mm of beam travel was only produced by a 200-g resistance, which is typically more than two times greater than any peak force measured. Raw EMG and root mean square (RMS; i.e., a 0.96-DC equivalent of the full wave-rectified AC signals to a digital RMS) of EMG bursts were recorded on additional channels of the signal acquisition system. EMG magnitude was reported as mean RMS magnitude of the EMG bursts time locked to ankle dorsiflexion. Collectively, this arrangement allows simultaneous monitoring of triceps surae EMG, resistive force, and velocity of shaft displacement. Recorded data were processed using a LabVIEW signal acquisition hardware and software system (National Instruments, Austin, TX). Dorsiflexions of 12.24 degrees were performed with 3-sec intervals at 49, 136, 204, 272, 350, 408, 490, and 612 degrees per second (deg/sec).
Recordings were performed at preinjury and at three postinjury intervals (weeks 2, 4, and 9). Treatment (saline control or ITB) was started at postinjury week 1 and lasted 4 weeks. Accordingly, these test intervals correspond to treatment weeks 1-, 3-, and 4-week withdrawal of ITB or saline. These studies provided a longitudinal measure of spasticity outcome induced by TBI and the influence of ITB during and after withdrawal of treatment.
Motor-evoked potential (MEP)
Transcranial magnetic motor-evoked potentials (tcMMEPs) were used (Magstim Ltd., Whitland, UK) as an index of functional motor outcome after TBI and ITB therapy. Magnetic pulse stimulation was applied to the animal's cranium to activate subcortical structures with an electromagnetic figure-of-eight coil (25 mm) placed over the center of the cranium. This stimulation was supplied by a Magstim TMS model using a single magnetic pulse with stimuli intensities from 30 to 70% of maximum stimuli intensities to provide motor thresholds and recruitment curves of motoneuron excitability in different experimental conditions. Magnetic pulses were delivered by computer LabVIEW programming (National Instruments hardware and software) to elicit tcMMEP responses. Onset latency time and amplitude were recorded from the right soleus muscle using intracutanious electrodes placed during the recording of ankle torque and EMGs. These experiments were performed immediately after completion of the velocity-dependent ankle torque and EMGs described above. Rats were immobilized in a custom-designed trunk restraint, without trauma or apparent agitation during this stimulation and recording. All recordings were performed on awake animals at preinjury as well as postinjury weeks 2 (week 1 treatment), 4 (week 3 treatment), and 9 (week 4 withdrawal).
Anxiety-like behaviors
Using the Noldus EthoVision hardware and software (Noldus Information Technology, Leesburg, VA) system, the experimenter recorded the amount of time the animal spent in the open arm, the closed arm, or the center piece of an elevated plus maze (EPM). Additionally, the experimenter manually scored rearing and grooming behaviors during the trial. Each animal was put into the center piece facing the same closed arm, and the experimenter had a 2-sec delay with which to remover themselves from the camera and rat's view. Each animal was tested three times over the course of the experiment with a minimum of 4 weeks between testing per animal. Testing was conducted at 5 weeks postinjury (saline or ITB; 4 weeks of treatment) and 4 weeks after withdrawal of saline or ITB (postinjury week 9). The testing procedure remained the same each time. Between trials, the animal was removed and the whole EPM was wiped down with 70% alcohol and allowed to dry before testing continued.
Footprint analyses
Animals were trained to walk on a 10-cm-wide by 120-cm-long walking surface in a Catwalk apparatus 27 (Noldus Information Technology) in a dark room. Three nonstop runs were acquired for each animal. The enclosed walkway was formed using a glass plate. Green light entered at the long edge of the glass plate, striking the surface below the critical angle for transmission, and was internally reflected. Accordingly, light escaped and was scattered by the contact of the rat's paws with the glass plate. 28,29 The paw prints illuminated during contact were captured by a high-speed video camera positioned below the walkway. Digital images were then transferred to the computer for data analysis using a program (CatWalk) that captures, indexes, stores, and analyzes multiple parameters of gait dynamics according to temporal and spatial patterns of footfalls. 27 During the first walking pass, the bright areas (bright spots that provide a high-resolution image of each footprints) were labeled and indexed. In subsequent passes, these footprints were interactively categorized (right forepaw, left forepaw, right hindpaw, and left hindpaw). The temporal and spatial positions of footprint spots that comprise each footprint, including qualitative measures of size and intensity, along with a tag (indicating which category relative to each paw), were stored in separate files. The CatWalk program utilized algorithms to assign tags to each paw and assessed the temporal and spatial pattern of footfalls. Animals were tested at preinjury and then at weeks 1 and 3 after ITB or saline treatments. A normal age-matched intact control group was also included. Analysis of normal step sequence and interlimb sequence regularity were calculated, and graphical representations of paw placement were generated using EthoVision CatWalk software (Noldus Information Technology).
Cognitive disability testing: Morris water maze (MWM)
An MWM was used to investigate deficits in spatial learning using a serial learning protocol designed to test rodents' ability to navigate to a hidden platform. 30 The MWM was composed of a 182-cm-diameter and 60-cm-high black plastic pool filled with water to a depth of 27-cm. Water temperature was maintained at 23–26°C by a submersible heating coil. A black Plexiglas platform, 10-cm in diameter, was located 2 cm below the water surface to serve as a hidden goal platform. The maze was located in a 3.0×3.0-m room with numerous extra maze cues (posters, pictures, and ceiling-mounted video camera) that remained constant throughout the experiment. The MWM protocol utilized four trials per day for 4 consecutive days, which began after 2 weeks of ITB treatment. For each trial, the animal was placed in the pool by hand at one of four start locations. The start locations were separated by 90 degrees and systematically identified as north, east, south, and west. On each test day, each animal performed trials from each of the four possible start locations, where the order was randomized for each animal on each day. The goal platform was positioned in the southeast quadrant of the maze, 45 cm from the outside wall. Each animal was given a maximum of 120 sec to find the hidden platform. If the rat failed to find the platform within 120 sec, it was placed on the platform for 30 sec by the experimenter. Animals were then placed in a heated incubator (28°C) during the intertrial interval (approximately 12–15 min). All animals were allowed to dry completely before being returned to their home vivarium. Swim speeds for each animal were calculated to insure that latency differences were not the result of impaired motor function. A computerized video tracking system (Noldus EthoVision, version 6.0; Noldus Information Technology) was used to record each animal's latency to reach the goal platform, swim pattern, total distance swum, and cumulative swim distance. In addition to search latency, several other parameters were recorded, including total path length to enable calculation of average swim speed. After the 4-day training session, probe trials were performed. For the probe trials, the hidden platform was removed. The probe trial consisted of placing the animal in the tank in a quadrant opposite the quadrant in which the platform had been located. The animal was given 60 sec to search, during which latency, number of fictive platform crossings, path length, search strategy, and time spent in each respective quandrant were determined. Probe trials were conducted immediately after the end of the 4-day trial to test acquisition of learned behavior (acquisition memory) and at 24 h thereafter to test retention of memory. A discrimination index was calculated using time spent in the goal and opposite quadrants.
Vestibulomotor testing: rotarod
Balance was tested using a rotating rod, on which an animal was challenged to maintain its balance while walking during a test session. The standard 2-min protocol used was one adopted from Hamm. 31 The animal was placed on the walking surface of the rod, which was stationary for the first 10 sec. The rotational speed was then slowly increased to 3 rpm for 10 sec and was steadily increased (automatic preset) by 3 rpm in 10-sec intervals until the maximum rpm of 30 was reached for the final 20 sec of the 2-min test period. The rotarod (Columbus Instruments, Columbus, OH) that was used features a variable-speed rod rotation in the range of 0–99 rpm with four individual electronic timers, with a fall sensor for each animal. Timers were activated by the operator at the moment the animal was placed on the rod and automatically stopped when the animal left the rod. The animal's task was to walk on the rungs as it rotates. The length of time the animal was able to maintain itself on the rotating rod during the 2-min protocol was used to quantitate each animal's maximum balance ability. Animals were tested for three trials per day with a minimum 10-min intertrial interval at pre- and post-TBI 2 and 4 weeks after treatment and 4 weeks after treatment withdrawal (postoperative week 8).
It was not feasible to accommodate all the physiological and behavioral testing at the same postinjury time point. Therefore, as marked, some testing intervals were conducted at different postinjury time points.
Histology and IHC experiments
Standard histology (Luxol Fast Blue and hematoxylin and eosin), Fluoro-Jade C (FJC) histochemistry, and fluorescent IHC were performed using selected cortical, brainstem, and lumbar spinal segments (L3-L5) after TBI-ITB or time-matched TBI saline or intact normal control groups that were euthanized at the week 4 postinjury time point as well as 4 weeks upon withdrawal (postinjury 9 weeks). We have reported on the details of methods recently. 16 Briefly, animals were deeply anesthetized using ketamine (110 mg/kg) anesthesia 21,22 and perfused through the left ventricle with phosphate-buffered saline (PBS; 0.1 M, 35°C, pH 7.4), followed by 4% paraformaldehyde in phosphate buffer (PB; 0.1 M, pH 7.4, 4°C). After postfixation (overnight in the same fresh fixative) and cryoprotection (30% sucrose in PBS), tissue (brain, brainstem, and lumbar spinal cord) specimens were sectioned by cryostat (40-μm thickness) and processed for these experiments. Fluorescent IHC was conducted for glutamic acid decarboxylase (GAD67), gamma-aminobutyric acid (GABAb), dopamine beta hydroxylase (DβH), and brain-derived neurotrophic factor (BDNF) using different sets of sections from same animal. Sections were washed in 0.01 M of PBS and incubated overnight (at 4°C) with monoclonal or polyclonal appropriate primary antibodies (Abs; Chemicon International, Temecula, CA). This procedure was followed by incubation with secondary Abs conjugated to Alexa Fluorochrome (highly cross-absorbed Abs; Molecular Probes, Eugene, OR). In control experiments, the primary Ab was either preabsorbed with an excess amount of the immunogenic peptide (1 μm/mL) or replaced by preimmune serum at the same dilution. Sections were washed thoroughly with PBS (0.01 M; pH 7.4) after Ab incubation. After washing with PB, sections were coverslipped with antifading mounting media (Vectashield; Vector Laboratory, Burlingame, CA). The histochemical marker FJC-staining procedure was conducted using a published protocol using another set of sections that selectively stained degenerating neurons independent of the mechanism of cell death. 32,33 FJC-stained sections were examined using a fluorescence microscope suitable for visualizing fluorescein isothiocyanate. All slides were stored at 4°C, and fluorescent images were captured within 1 week after completion of IHC for light morphometric studies. Because spasticity measurement was performed from hindlimb muscles, lumbar spinal cord segments were also used for IHC. A quantitative evaluation was made.
Statistical analyses
Analysis of variance (ANOVA) and Student's t-tests were used to statistically evaluate significance in behavioral, spasticity, and gait outcomes. Repeated ANOVA was also used to test the effectiveness of ITB on spasticity and/or TBI using the same animals before and after treatments.
Results
Ankle torques and ankle extensor EMGs (spasticity) and gait preinjury recordings
Velocity-dependent ankle torques and ankle extensor EMGs were recorded in all animals before experimental injury at eight test velocities (49, 136, 204, 272, 350, 408, 490, and 612 deg/sec; refer to preinjury data in Fig. 1A,B). At the lowest rotation velocity of 49 deg/sec, a mean value of 76.61 (±6.87) kdynes of ankle torque was observed, and through the next three lowest velocities (136, 204, and 272 deg/sec), small increases in velocity-dependent torque were observed. At these lowest rotation velocities, no systematic, time-locked bursts of EMG activity were observed in the ankle extensor muscles. At the highest four rotation velocities, greater increases in velocity-dependent torque, and EMG amplitudes, that were time locked with ankle extensor muscle stretch, were observed (Fig. 1B). At the highest velocity (612 deg/sec), a mean peak ankle torque amplitude of 116.62 kdynes was recorded.
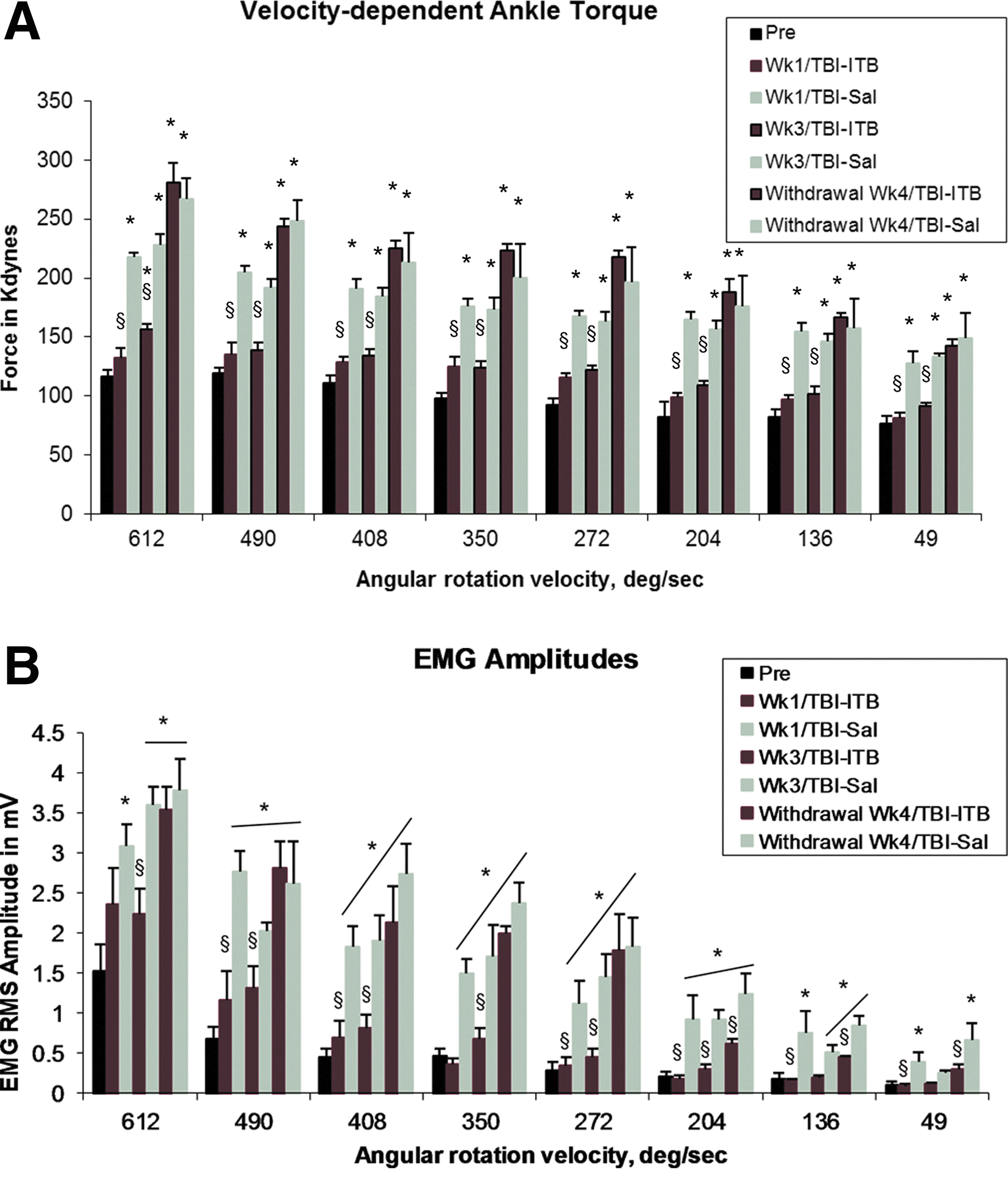
One week of ITB treatment completely blocked spasticity [elevated ankle torques (
Postinjury week 2 (saline treatment week 1)
At 2 weeks (week 2) after injury (1 week after treatment), mean peak ankle torque amplitudes were observed to be significantly increased at each of the ankle rotation velocities in saline-treated animals (week 1/TBI-Sal; Fig. 1A), compared with values recorded in these animals before injury (Fig. 1A). For example, in the TBI-Sal week 1 animals, at the lowest velocity, mean peak ankle torques were observed to be 127.76±10.18 kdynes, compared to 76.61±6.87 kdynes recorded in animals before injury, a mean increase of 57.6%. At the highest velocity (612 deg/sec), mean ankle torques were recorded to be 217.74 kdynes, an 86.7% increase, compared to the 116.62 kdynes recorded in these animals before injury (Fig. 1A). Accompanying these increases in ankle torque were parallel increases in mean EMG burst amplitudes recorded from the ankle extensor muscles (Fig. 1B). These EMG bursts were time locked to the stretch of the ankle extensor muscles. They appeared systematically at rotation velocities beginning at 49 deg/sec and progressively increased in amplitude at higher rotation velocities through 612 deg/sec. In addition to the decreased threshold for stretch-evoked EMGs, the mean EMG amplitudes observed at each of the test rotations were significantly greater than observed preinjury. Collectively, these observations revealed that at week 2 postinjury, significantly increased velocity-dependent stretch-evoked resistance was observed at all test velocities. In addition, the increased amplitude of stretch-evoked EMGs at each test velocity indicated that increased extensor muscle activation contributed to the increased ankle torque recorded at each of these test velocities.
Postinjury week 2 (ITB treatment week 1)
When tested at 2 weeks after injury and 1 week of ITB treatment, mean ankle torques in the ITB group were not significantly different from those recorded in these animals before injury and were significantly less than what was recorded in the TBI-Sal group at all eight test velocities (Fig. 1A). Similarly, mean EMG amplitudes were not significantly different from those recorded in these animals before injury, but were significantly less than that observed in the TBI-Sal group at all eight test velocities (Fig. 1B). Collectively, these observations revealed that compared to TBI-Sal animals, 1 week of ITB treatment significantly reduced or blocked the development of significantly increased velocity-dependent ankle torques and accompanying stretch-evoked activation of the ankle extensor muscle EMGs.
Postinjury week 4 (saline treatment week 3)
When tested at week 4 after injury, the ankle torques and time-locked EMG amplitudes recorded from the lower limb in the TBI-Sal group were still elevated at all eight test velocities (Fig. 1A). They were similar to those observed at postinjury week 2 and were significantly elevated, compared to preinjury control values.
Postinjury week 4 (ITB treatment week 3)
Mean velocity-dependent ankle torque and EMG amplitudes recorded at week 4 (after 3 weeks of ITB treatment; week 3/TBI-ITB) were significantly smaller than those recorded at week 4 from TBI-Sal-treated control animals (Fig. 1A,B). Compared to the TBI-Sal group, the velocity-dependent triceps surae EMG amplitudes time locked to ankle rotation were also significantly smaller in the week 3/TBI-ITB animals (Fig. 2B). However, these ankle torque and EMG amplitudes recorded in the TBI-ITB treatment group were significantly greater from those recorded before injury (34 and 47% greater in ankle torque and EMGs, respectively, at the highest rotation velocity). Collectively, these observations indicated that TBI-ITB treatment significantly blocked (at week 1 post-ITB) or reduced (week 3 post-ITB) the appearance of lower-limb spasticity, compared to observations in the TBI-Sal group.

Graphs of foot placement patterns. TBI-Sal and TBI-ITB animals were tested for gait disability using a three-dimensional footprint analysis protocol (CatWalk). ITB-treated animals showed better gait parameters at post-ITB treatment weeks 1 and 3, compared to saline-treated TBI control animals. TBI, traumatic brain injury; Sal, saline; ITB, intrathecal baclofen.
Treatment withdraw week 4
When tested at 4 weeks after treatment (saline or ITB) withdraw, the ankle torques recorded in the TBI-ITB withdrawal group showed a dramatic increase in the mean velocity-dependent ankle torque amplitudes (Fig. 1A). At 612 deg/sec, mean ankle torques for week 4 ITB withdrawal was 280.75 kdynes, an 80% increase from those recorded after 3 weeks of ITB treatment. This mean was even slightly greater (9%) than the 247.28-kdyne mean recorded in the week 4 (Sal) withdrawal animals at 612 deg/sec. Accompanying these increased velocity-dependent ankle torques at this withdrawal point were parallel increases in mean amplitude of muscle stretch-evoked EMGs recorded from the ankle extensor muscles at all but the three lower rotation velocities (Fig. 1B). Interestingly, the velocity-dependent EMG amplitudes observed in the ITB withdrawal group were significantly smaller, especially in the three lowest rotation velocities, than EMGs recorded at corresponding test velocities in the time-matched Sal group (week 4 withdraw control; Fig. 1B).
Gait analysis
Patterns of limb placement in normal and treated (saline and ITB) animals were analyzed and compared using a CatWalk to determine the step-sequence regularity index (SSRI). The SSRI defines the degree in which the steps taken fall within one of the regular step patterns. When tested at 2 and 4 weeks after injury (1 and 3 weeks of treatments), TBI-Sal animals showed marked irregularities in the step-sequence index (Fig. 2, middle-row panel). By comparison, TBI-ITB-treated animals showed greater regularity of limb movement, compared to TBI-Sal control animals.
Recovery of balance function
Rotarod balance function tested at postinjury (postoperative week 2, 1-week treatment) revealed significant decreases in balance scores for both TBI-Sal and TBI-ITB animals, compared with measures obtained before injury. At this time point, no significant differences in mean balance scores were observed between ITB- and Sal-treated TBI animals, although the mean score of the ITB group trended lower than the Sal-treated injured controls at this time point (Fig. 3). At postinjury week 4 (3-week treatment), the mean rotarod balance score in Sal-treated TBI animals was similar to that recorded at postoperative week 2, whereas the mean score recorded for TBI-ITB-treated animals was observed to be greater and was not significantly different from the mean value recorded in age-matched normal animals. The balance scores recorded in Sal-treated TBI animals at postoperative week 8 (3 weeks of withdrawal) were similar to those recorded at weeks 2 and 4. However, whereas the balance mean score recorded in the ITB-TBI group at week 8 remained lower than observed in normal aged-matched controls, this mean score was significantly greater than recorded in Sal-treated TBI animals (Fig. 3).
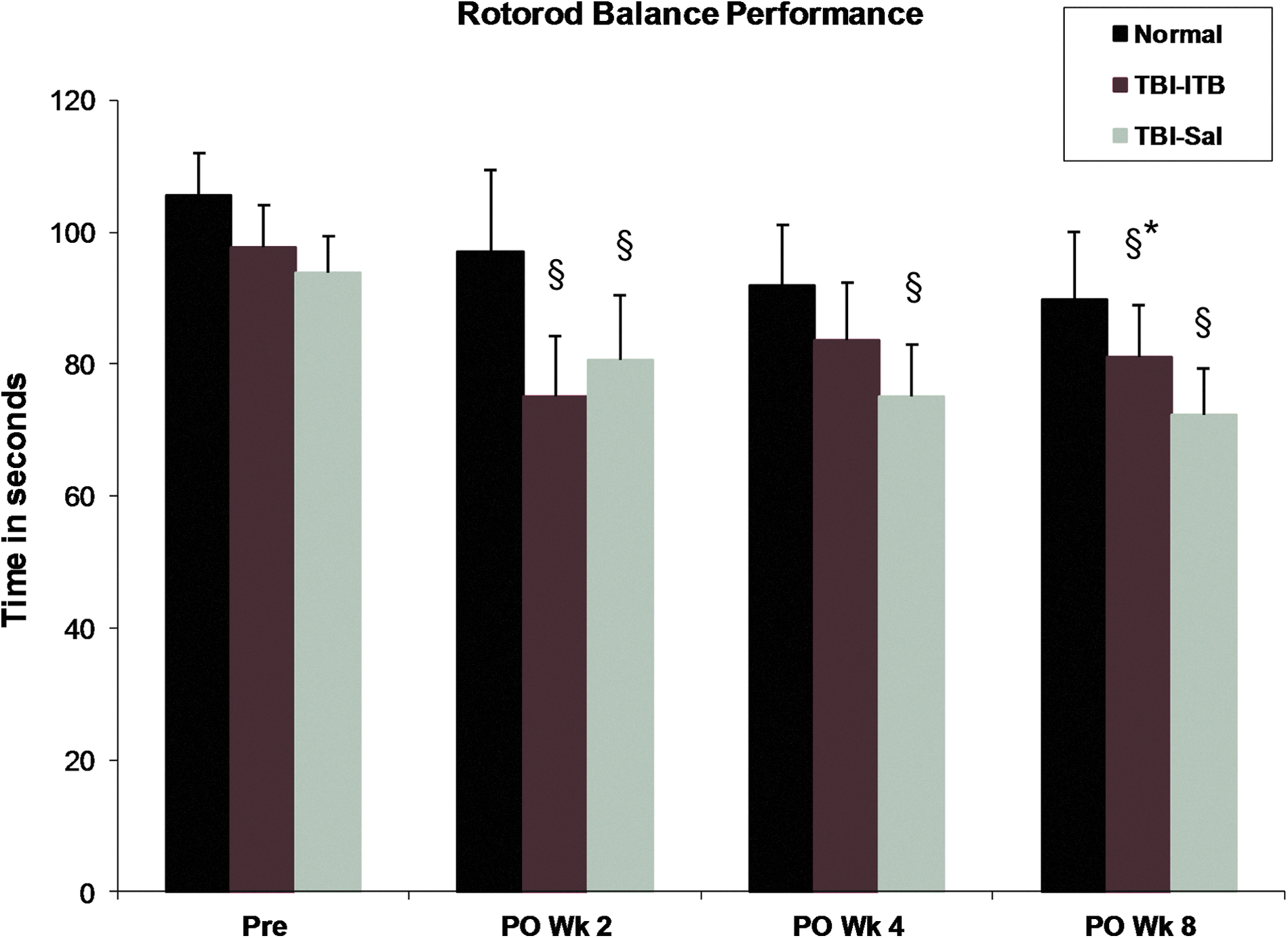
Early intervention with ITB treatment did not adversely affect balance performance. In fact, at ITB treatment week 3 (postoperative week 4), treated animals did not show any balance deficit, compared to age-matched normal animals. § p<0.05, compared to age-matched normal animals; *p<0.05, compared to TBI-Sal animals. ITB, intrathecal baclofen; TBI, traumatic brain injury; Sal, saline; PO, postoperative.
Memory performance
TBI-Sal and TBI-ITB animals were tested for spatial serial learning performance at 3 weeks postinjury and compared to age-matched normal animals (Fig. 4). TBI-Sal animals revealed longer search latencies on test days 3 and 4, with the difference on test day 4 being significantly longe,r compared to mean search latencies observed for normal or TBI-ITB animals. Mean search latencies for TBI-ITB animals were observed to be significantly longer than observed for normal animals only on test day 2 (Fig. 4). Mean search latencies observed on the third and fourth test days were not different from that observed in normal animals. Least-squares linear regression of latencies for platform discovery revealed that the regression slopes (rate of change for platform discovery over the 4 days) for normal and ITB-TBI animals were similar (17.2 and 17.7, respectively) and were greater than observed for TBI-Sal animals.
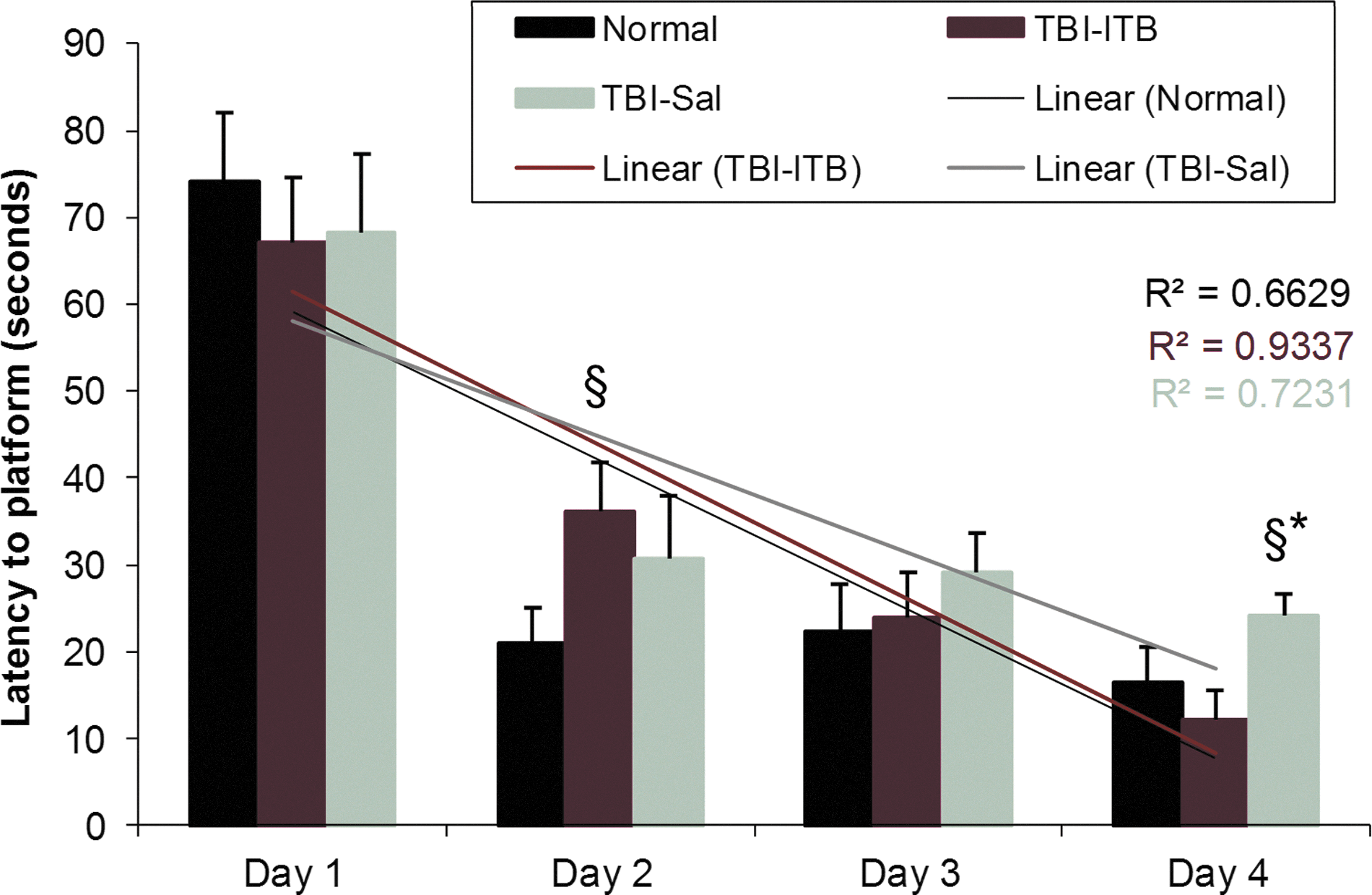
Animals were tested for 4 conjugative days in a MWM for serial learning. At the final day of testing, ITB-treated TBI animals revealed better performance (seek latency to platform) than TBI-Sal controls. ITB-treated TBI animals showed similar learning slope to time-matched normal animals and a different slope pattern than TBI-Sal controls over 4 days. § p<0.05, compared to time-matched normal animals; *p<0.05, compared to TBI-ITB animals. MWM, Morris water maze; ITB, intrathecal baclofen; TBI, traumatic brain injury; Sal, saline.
Retention of spatial learning was tested by comparing the latencies of two probe trials, conducted 24 h apart. The first probe trials for platform latency were conducted immediately after the fourth-day serial learning testing and then 24 h later (fifth day). Normal animals revealed an approximate 10% increase from 23.9 to 26.5 sec, respectively, for the fourth- and fifth-day probe trials. TBI-Sal animals revealed a 32.2% increase, from 29.7 to 39.3 sec, respectively, for the fourth- and fifth-day probe trials. TBI-ITB animals revealed a 38.2% increase, from 26.2 to 36.2 sec, respectively, for the fourth- and fifth-day probe trials (Table 1). No significant differences were obtained in the statistical comparison of the probe latencies for any of the group means.
Acute ITB-treated animals did not show any adverse effect in their acquisition of learned behavior (acquisition memory) at day 4 in probe trial or their retention of memory at day 5, 24 h after their 4-day serial learning.
SEM, standard error of the mean; TBI, traumatic brain injury; ITB, intrathecal baclofen; Sal, saline.
Anxiety-like behavior
Both TBI-ITB and TBI-Sal animals were tested for anxiety-like behavior using an EPM equipped with EthoVision video tracking (Noldus Information Technology) to score time distribution in the open versus closed portions of the maze. TBI-ITB-treated animals distributed significantly more time (20 vs. 5%) in the open areas of the EPM (e.g. showed less anxiety-like behavior) at post-ITB treatment week 4 (postinjury week 5), compared to Sal-TBI animals. However, at the 4-weeks postwithdrawal phase (postinjury week 9), no significant differences were detected between TBI-ITB and TBI-Sal controls (Fig. 5).

ITB treatment resulted in less anxiety-like behavior at post-treatment week 4 (postinjury week 5). The ITB group spent significantly greater time in the open arm of the elevated plus maze. However, at the 4-week postwithdrawal phase (postinjury week 9), no significant differences were detected between ITB- and Sal-treated TBI groups. ITB, intrathecal baclofen; TBI, traumatic brain injury; Sal, saline.
tcMMEPs
The excitability of neural components constituent to the activation of the lower-limb soleus muscle was tested by application of transcrandial magnetic stimulation to the cranium overlying the motor/sensory cortex.
Representative waveforms and mean MEP amplitudes recorded in the soleus muscle in normal, TBI/Sal, and TBI/ITB animals are shown in Figure 6. In normal intact animals, the mean MEP amplitude recorded in the soleus muscle was 59 mV. MEPs recorded at 2 weeks after TBI were observed to be significantly decreased to 13 mV from the values recorded in intact normal animals (pre). The mean amplitude of MEPs that were recorded after 1 week of ITB treatment was 23mV, a 77% improvement, compared to the MEPs recorded in saline-treated TBI animals.
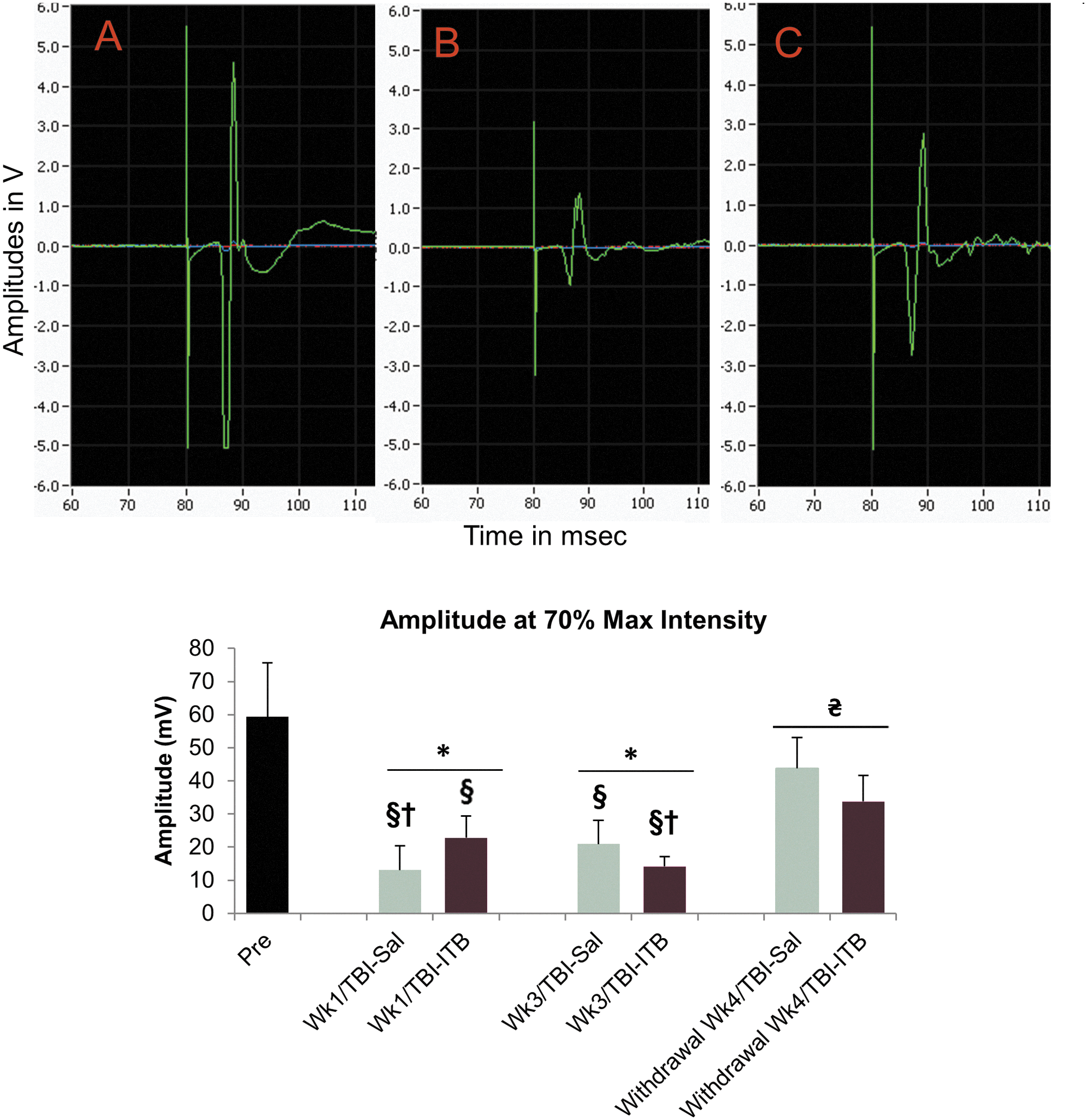
Transcranial magnetic motor-evoked potentials (MEP) amplitude in soleus muscle of normal, TBI-Sal, and TBI-ITB animals [upper panel, raw waveform at 70% maximum (Max) current at ITB treatment:
MEPs recorded at 3 weeks after treatments (Sal or ITB) remained to be significantly decreased to 21 (TBI-Sal) and 14 mV (TBI-ITB), compared to the values recorded in intact normal (pre). Interestingly, the mean amplitude MEPs that were recorded after 3 week of ITB treatment were further suppressed, when compared to the mean amplitude that were recorded after 1 week of ITB treatment (23–14 mV), a 64% reduction in amplitude (Fig. 6).
Interestingly, a significant spontaneous improvement in mean MEP amplitudes were detected at postoperative TBI week 9 in both TBI-Sal or TBI-ITB animals (withdrawal week 4/TBI-Sal or withdrawal week 4/TBI-ITB).
Histology and IHC
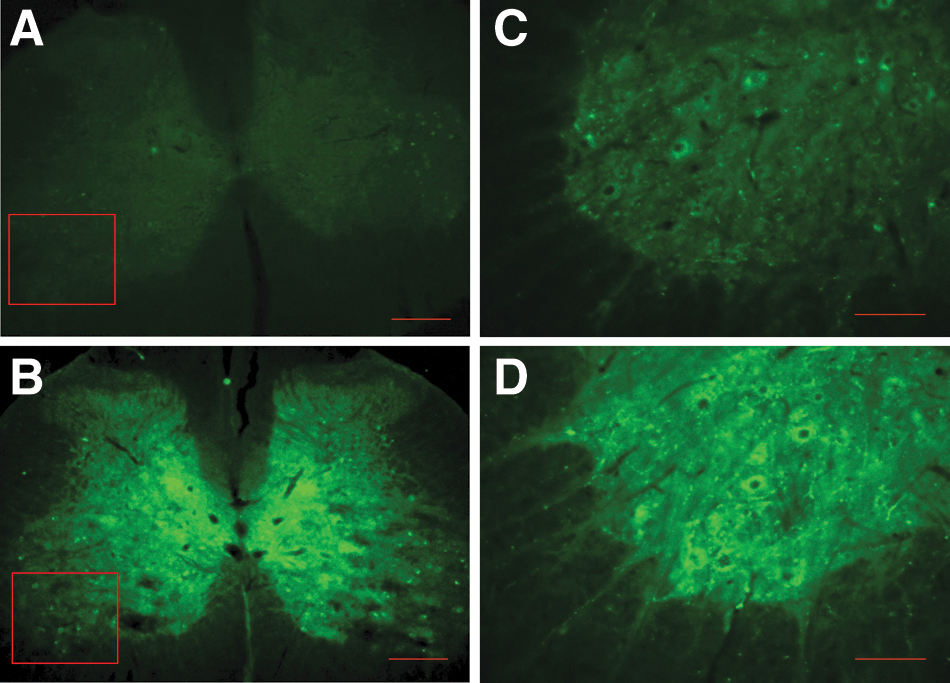
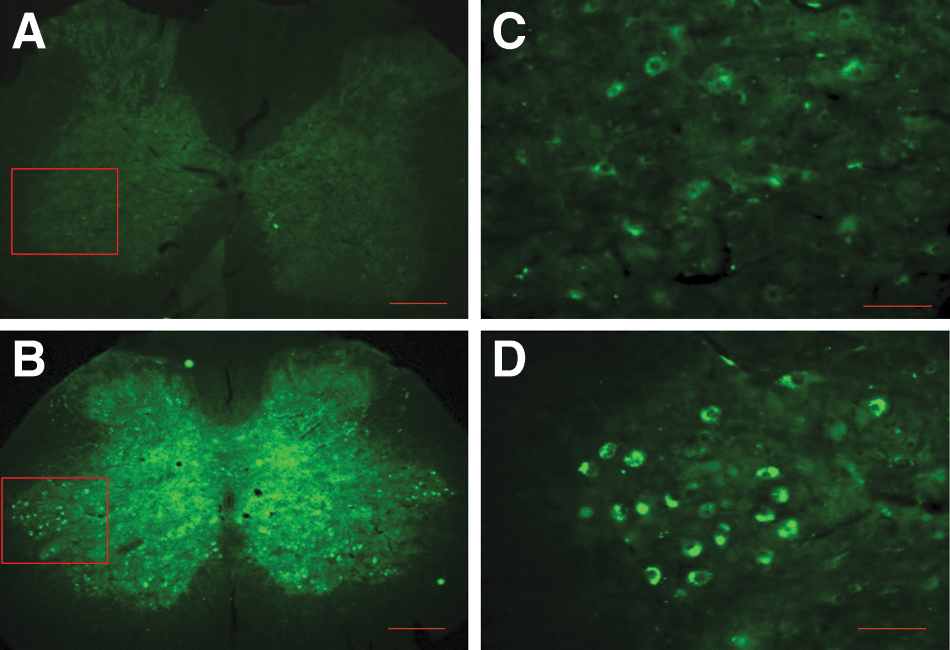
Histological and IHC experiments using brain, brainstem, and spinal cord tissues of TBI specimens indicated diffuse and widespread axonal injury in multiple sites of the brain, including cortex, hippocampus, corpus callosum, and pyramid. Qualitative microscopic examination of hippocampal samples revealed cell loss, shunken cells, and hyperchromatin deposition (Fig. 7C, hippocampal C3 area). FJC-stained lower brainstem samples showed remarkable diffuse axonal injury in pyramid, pyramidal decussation, medial longitudinal fasciculus, tectospinal tract, medullary reticular nuclei, and nucleus solitary tract (Fig. 7B). A normal cortrol sample stained with FJC is shown in Figure 7D, where such diffuse axonal injuries were not detectable. More surprising, IHC of brainstem samples from TBI-Sal or TBI-ITB animals showed clear eveidence of cell loss in the locus ceruleous (LC; Fig. 8B,C, fluorescent photomicrographs of DβH IHC). ITB treatment did not show any differences in TBI-related decreases in the expression of LC cell clusters (Fig. 8B). However, smaller-size perivascular spaces were often detected in ITB-TBI LC samples, when compared to corresponding TBI-Sal samples, where larger perivascular spaces were detected (Fig. 8C). DβH IHC of hipocampus showed a profound reduction/loss of norepinephrine (NE) immuoreactivity in TBI-Sal samples (Fig. 9C, hippocampal C3 area), when compared to corrosponding samples of intact normal control (Fig. 9A). Interestingly, ITB treatment showed a remarkable recovery of NE expression in this CA3 area (Fig. 9B), although this NE expression qualitatively still remained less than corresponding normal CA3 NE expression (Fig. 9A).

Shunken and hyperchromatin deposited cells were identified throughout the hippocampus (

Evidence of cell loss in the locus ceruleous (small arows) in both TBI (

ITB treatment resulted in up-regulation of DβH-positive spared NE fibers in the CA3 areas of the hippocampus (
Qualitative microscopic examination of immunostained cord tissue taken from animals during the ITB treatment phase (3 weeks of treatment) showed marked up-regulation of GAD67 (a GABA isoform; Fig. 10B) and GABAb receptor staining (Fig. 10D). There were no apparent differences in GABAb and GAD67 expressions in lumbar spinal cord between ITB- or saline-treated injury controls at withdrawl phase (4 weeks after treatment withdrawl). In addition, ITB-treated animals showed marked up-regulation of DβH (Fig. 11) and BDNF (Fig. 12) in the lumbar spinal segment (dorsal, ventral, and intermediate gray matter).
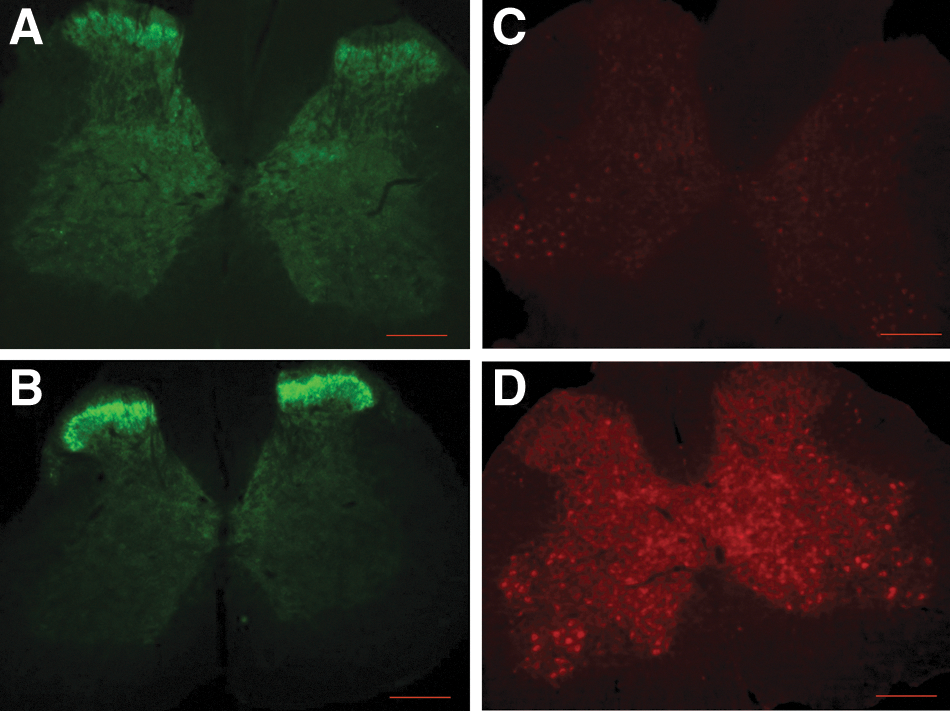
ITB treatment resulted in up-regulation of the neuroprotective molecule, brain-derived neurotrophic factor (BDNF; Fig. 12,
Increased expression of markers for noradrenergic (DβH) and GABAergic (GAD67, GABAb) activity is consistent with increased inhibitory function, which could influence the expression of spasticity in these ITB-treated animals. This indicated that during ITB treatment, the receptors (as well as its ligand) were active and the benifical effects of ITB in controlling spasticity could have been mediated through these receptors. Moreover, this indicated that there were no long-term detectable differences in the expression of these receptors, and thus it would be anticipated that future treatments targeting these receptors would be effective.
Discussion
These studies were conducted to evaluate the development of lower-limb spasticity after cTBI and test the efficacy of early ITB treatment on TBI-induced spasticity. In addition, safety measures were conducted to evaluate the potential negative consequences of antispastic medication delivery to a recovering central nervous system (CNS). These safety measures included evaluation of cognitive performance, balance function, gait regularity (as a measure of the CNS’ ability to coordinate a complex motor task), anxiety-like behavior, and a neurophysiological assesment of the excitabiity of the executive motor system using TMS MEPs recorded from the soleus muscle. IHC studies of spinal cord, hippocampal, and brainstem tissues were performed to highlight the expression molecular markers that are known to influence the excitability of neural elements constituative to lower-limb motor control, congnitive function, and noradrenergic function.
Spasticity
Behavioral results from 11 control (Sal-TBI) animals confirmed that there were significant enduring spasticity disabilities after TBI. Ankle torque and EMG recordings demonstrated progressive changes in ankle extensor muscle stretch reflex activity patterns after TBI. At the first test interval, week 2 postinjury, ankle torques were up to 100% larger that recorded before injury and were observed at each test velocity from 49 through 612 degrees. These increased torque amplitudes were velocity dependent, with the greatest increases in torque observed during rotation at the upper four velocities, compared to the lower four. Because the increased torque amplitudes were time locked to increased EMG burst discharges in the triceps surae muscles, we concluded that these ankle torque and EMG changes were attributed to increased triceps surae muscle stretch reflexes. These altered reflex patterns were definitive of a clinical condition of spasticity (i.e., appearance of exaggerated velocity-dependent lengthening resistance). These significantly increased velocity-dependent ankle torques and EMG burst discharges in the extensor muscles were observed at the postinjury week 4 and 9 test intervals. Accordingly, these robust spasticity patterns were concluded to appear with an early onset after TBI and to have an enduring time course.
There were differences in the amplitude, nature, and time course of the spasticity patterns observed after cTBI, compared to those after mid-thoracic spinal cord contusion. 25 Spinal cord injury (SCI)-induced spasticity appeared with an early onset and, when tested at week 1 postinjury, was significantly increased at all test velocities from 49 to 612 deg/sec. However, the early-onset SCI spasticity was observed to fade into a period of hyporeflexia, during postinjury weeks 2 and 3, and then to return as an enduring spasticity during postinjury week 4. However, this later-onset pattern of SCI-induced spasticity was only observed during the highest four ankle rotation velocities, whereas the TBI-induced spasticity appeared at all test velocities, and this pattern was enduring. In addition, the TBI-induced spasticity showed a greater amplitude of velocity-dependent ankle torques than observed after thoracic SCI. Accordingly, the TBI-induced spasticity was observed as an early-onset, enduring, and clinically relevant robust pattern of spastic disability.
There has been steady progress in our understanding of the changes in pre- and postsynaptic factors that regulate motoneuron excitability in the normal and hyper-reflexic state after SCI. Accompanying progress in understanding changes in presynaptic mechanisms, substantial progress has occurred in the understanding of postsynaptic mechanisms that regulate the input/output gain of motoneuron discharge. 34 –42 Collectively, these studies have revealed that the gain of synaptic inputs can be amplified by brainstem/monoaminergic inputs that regulate dendritic persistent inward currents (PICs) utilizing sodium and calcium channels. The higher the persistent inward current, the higher the synaptic gain and consequent burst rate of the motoneurons. Segmental regulation of PICs occurs through inhibitory mechanisms that regulate afferent inputs. In SCI it has been proposed the acute period of hyporeflexia that follows SCI can be attributed to a reduction in dendritic PICs. Subsequently, after several weeks, motoneurons reacquire PICs that can be easily initiated by segmental inputs. These unregulated PICs are proposed to significantly contribute to clonus and spasms as well as associated amplified bistable properties of motoneurons. 39 In this context, the segmental inhibitory processes, such as presynaptic inhibition, have an even more important role in the regulation of sensory transmission, because it was observed that at clinically relevant doses, baclofen had significantly more influence on 5-enolpyruvylshikimate-3-phosphates initiated by mono- and polysynaptic inputs to motoneurons than on the dendritic PICs themselves. 43 Therefore, the acute ITB treatment might result in more-effective inhibition of sensory afferent transmission to motoneurons because of the absence of progressive maladaptive segmental and descending plasticity (e.g., spontaneous PICs) that typically increase at postinjury weeks 4 and 5 in SCI, and that have been shown to be associated with injury of central descending monoamine (NE and 5-hydroxytryptamine) systems.
When tested after 1 week of ITB treatment, the velocity-dependent ankle torque amplitudes and time-locked burst EMGs were significantly less than observed in the time-matched Sal-TBI animals. Although a small increase was observed at 612 degrees when tested after 3 weeks of treatment, mean ankle torque amplitudes were still reduced by 61%, compared to those recorded in the time-matched saline-treated TBI controls.
When tested at 4 weeks after withdraw of ITB treatment, the mean ankle torques in ITB-TBI animals were observed to be significantly greater at all test velocities than recorded during ITB treatment. Collectively, these data indicated that ITB was effective in suppressing expression of spasticity during treatment, but, once removed, did not prevent the appearance of lower-limb spasticity that was similar in magnitude to that recorded in the untreated Sal-TBI animals.
Gait analysis
These studies revealed that cTBI animals exhibited an irregular gait disability that was detectable by this analysis of footprint sequences (CatWalk; Fig. 2). These step-sequence irregularities were not observed in ITB-TBI animals tested at the end of 1 and 3 weeks of treatment. It is suggested that the step irregularities observed in the cTBI animals was, in part, a result of spastic contractions that were inappropriately interfering with smooth gait progression. It is further suggested that ITB suppression of this velocity-dependent spasticity contributed to smoother gait pattern that was less encumbered with spastic interference.
Vestibulomotor
Balance disorder is a frequent complaint after human TBI. 3,44 Balance instability has also been reported as a frequent, particularly sensitive measure of brain injury in experimental models of TBI in the rat. 31 Previous studies have revealed significant involvement of the noradrenergic system in the setting of vestibulospinal reflex gain. 45 –48 Because we observed significant changes in the nucleus (LC), which provides a principle noradrenergic innervation of the vestibular system, 48 we anticipated that TBI-associated balance deficits would be significant when tested using the ability to maintain balance while walking on the rotating cylinder of the rotarod device. Consistent with previous studies of balance changes induced by experimental TBI, 31 Sal-TBI animals revealed significant, enduring decreased rotarod walk times that were terminated by falling from the rotating cylinder (Fig. 3).
In addition, several studies have identified the specific role of GABAb mechanisms in gain setting as well as adaptive plasticity after vestibular injury. 49,50 When tested after 1 week of ITB treatment, ITB-TBI animals displayed significant balance deficits that trended in the direction of greater than observed in Sal-TBI animals at that interval. However, when tested after the third treatment week and also at 3 weeks after ITB treatment removal, the ITB-TBI animals' rotarod scores were not significantly worse than those recorded in normal animals. It is uncertain how this improvement in rotarod performance occurred. It has been reported that injection of baclofen slightly accelerated vestibular compensation after unilateral vestibular injury. 50,51 Although the ITB cannula tip is situated in the subdural space of the lumbar spinal cord, it is known that the steady-state concentration of baclofen in the fourth-ventricle CSF is 25–30% of the target dose. 24 Pompeiano and colleagues have reported that microinfusion of L-baclofen into LC altered the gain of vestibulospinal reflexes. 46 Therefore, it is possible that the GABAb stimulation that was induced during ITB treatment initially (after 1 week of treatment) decreased vestibulospinal reflex gain and added to the diminished rotarod performance. It is possible, however, that the continued GABAb stimulation contributed to the vestibular compensation that provided the improved rotarod performances in animals after 3 weeks of ITB treatment. These rotarod studies revealed that after 3 weeks of ITB treatment, rotarod balance scores were significantly greater than those recorded in Sal-TBI controls.
Cognitive
Cognitive recovery is typically the hallmark concern related to TBI and, accordingly, is a first measure of safety for experimental therapy. Experimental findings regarding the relationship between TBI and cognitive dysfunction have been central to various laboratory models of TBI, including controlled cortical impact, lateral and mid-line fluid percussion, and impact acceleration animal models (based upon Marmarou's model) of head injury. 17,30,52,53 Laboratory assessment of cognitive function in rats capitalizes upon their intrinsic abilities to navigate mazes. The capacity and efficiency for performing for a maze task is, in part, proportionally related to the ability to create and store a cognitive map. TBI may affect the mechanisms by which visual-spatial memory is formed and recalled and may be associated with marked anterograde amnesia.
The most widely used test for cognitive evaluation in experimental TBI models is the MWM test, originally developed to examine spatial memory in rats with hippocampal lesions. 54 As previously described, the maze is a circular tank filled with opaque water that hides a small, submerged platform that the animal must escape to by using simple visual cues. Depending upon the protocol and the arrangement of the equipment, the MWM can be used to test both anterograde amnesia (learning, using serial trials for platform discovery), as well as retrograde amnesia (memory, using a probe protocol). In addition, because the trials are videotaped, swim speed and distance traveled check for potential confounding variables of motor deficit.
Spatial navigation ability and episodic memory function are believed to be hippocampally mediated, whereas the prefrontal cortex is believed to mediate the executive functions involved in learning search strategies to solve a problem. 55 Sal-TBI animals revealed a consistent pattern of working memory deficits when tested using multiple trials over 4 days to measure MWM search latencies that were consistent with previous reports for spatial learning impairments in experimental TBI. 56,57 It is noteworthy that ITB-TBI animals revealed significant learning deficit only on day 2 of the 4-day series of testing; there were significant differences in mean search latencies between ITB-TBI and Sal-TBI animals on the fourth test day, no significant differences in mean search latencies between ITB-TBI and normal animals were observed by the fourth day, and the overall rate of change in platform latency (learning curve) was similar to that observed for normal animals. This was potentially a surprising finding relative to previous studies that have reported dose-dependent reductions in serial learning in rats that received baclofen by oral dosage, 58 by intracerebroventricular injection, 59 or received by microcannulation directly into the hippocampus. 60 However, there are a few issues that must be considered relative to the influence of chronic ITB on hippocampal function in the setting of TBI. The previous studies of baclofen influences on cognitive performance were typically performed in normal animals, with acute, rather high-intensity doses of baclofen. Spinal cord reflex excitability studies have revealed that baclofen can act differently on neuron excitability in the chronic treatment versus the acute treatment setting. 19
The hippocampal tissues of the ITB animals revealed marked up-regulation of noradrenergic expression, compared to hippocampal tissues of Sal-TBI animals (Fig. 9). The ITB-associated increased NE expression is consistent with previous reports of facilitatory GABAergic influence on central noradrenergic transmission. 61 –63 Because it is known that cognitive performance is dependent upon noradrenergic induced up-regulation of BDNF, 64 it is suggested that this increased NE expression in the hippocampus of ITB-TBI animals could be associated with the improved MWM performance in ITB-TBI animals, compared to Sal-TBI animals.
Anxiety
Anxiety disorder is emerging as an important comorbidity associated with TBI. Accordingly, the influence of experimental treatment on anxiety expression is an important measure of safety, particularly if the treatment induces changes in the expression of anxiety symptoms. It has been reported that anxiolytic agents increase anxiety expression in rats tested by their behavior in an EPM; normal rats spent approximately 45–50% of their session exploring the open areas, whereas anxiolytic-treated rats spent only 20% of the session in the open areas of the maze. 65 Accordingly, the EPM was used to compare behavior in Sal-TBI animals to that in ITB-TBI animals during and after withdrawal of ITB therapy. When tested at postinjury week 5 (post-treatment week 4), TBI animals spent 88% of test session time in the closed regions of the EPM. By contrast, the week 4 ITB-treated animals spent 62% of the session time in the closed arms. This 26% reduction in closed arm place preference is interpreted as an expression of less anxiety in ITB-treated animals. The treatment-associated decrease in the behavioral marker for anxiety in ITB-TBI animals during treatment is consistent with reports of success in treating anxiety and depression in post-traumatic stress disorder patients with baclofen. 66 At 4 weeks after withdrawal of treatment, no significant differences were detected between ITB-treated and saline-treated TBI controls.
MEPs
tcMMEPs have become an important tool to provide an in situ evaluation of the integrity of locomotor pathways as well as a neurophysiological measure of reflex excitability after injury, disease, or therapy. Although the exact neural substrate essential for the TMS-elicited tcMMEPs in lower-limb muscles in rats has not been determined, studies have revealed the strategic importance of the ventromediolateral white matter of the spinal cord for TMS-elicited lower-limb tcMMEPs. 67 –69 In addition, the latency waveforms of spinal-cord–evoked potentials elicited by TMS application to the cranium reveals that, typically, the stimulation in rats produces a direct (vs. indirect, e.g., synaptic) activation of subcortical white matter. 70 Accordingly, TMS-elicited tcMMEPs were recorded to compare motor excitability in Sal-TBI and ITB-TBI animals during and after ITB treatments. Compared to intact normal animals, mean tcMMEPs in Sal-TBI animals were significantly reduced in amplitude (59 vs. 13mV, respectively). This reduction in tcMMEP amplitude is consistent with the extent of descending motor fiber injury shown in Figure 7B. In addition to the marked FJC expression in the pyramidal decussation, it is likely that these injured fibers were only part of the total corticofugal fibers injured. It is known that in this impact/acceleration model of TBI, the dense white matter of the fore- and mid-brain are the particularly vulnerable regions of diffuse axonal injury. Although axonal injury has been observed in a multifocal pattern, it is greatest in the dense compacted parasagittal white matter (corpus callosum, internal capsule, and meso-pontine regions of the superior cerebellar peduncle) that is particularly subject to mechanical shear. 71 –73 At these more rostral levels, the nonsegregated projection path would also include the corticoreticular fibers, which activate fibers of the reticulospinal tract, 74 and which have been suggested to be critical for the expression of the tcMMEP. 67,69 Accordingly, the reduced tcMMEPs in Sal-TBI animals are consistent with diffuse axonal injury of the descending motor system. That the latency of the tcMMEP in Sal-TBI animals was not detectably longer is consistent with the previous findings that demonstrated that smaller-diameter fibers are more vulnerable to diffuse axonal injury (i.e., stretch injury) than the largest fibers. 75 Those tcMMEPs recorded in ITB-TBI animals during week 3 ITB treatment were smaller than those recorded in Sal-TBI animals (21 vs. 14mV, respectively). This decrease in evoked potential amplitude is consistent with the ITB-mediated activation of GABAb receptors known to be located on the terminals of reticulospinal projection fibers and spinal interneurons through which the descending motor volley courses to reach lower limb motoneurons. 76,77 However, when the mean amplitudes of the tcMMEPs were compared at 4 weeks after discontinuation of ITB treatment, no differences in mean amplitudes between Sal-TBI and ITB-TBI MEPs were observed. These data indicate that whereas excitability was reduced during ITB treatment, no enduring change in MEP excitability was detected as a function of treatment.
IHC
The significant benefit of ITB in controlling spasticity after TBI is well correlated with the findings revealed by IHC analysis of the Sal-TBI- versus ITB-TBI-treated tissues, particularly the marked up-regulation of GAD67/GABAb observed in the ITB-TBI tissues. Moreover, changes in neurotrophic factor expression, the IHC expression of BDNF, two factors that are essential for normal patterns of presynaptic inhibition: GABA/GABAb receptors and descending NE fiber sprouting (DβH-positive fibers) in ITB-treated animals' spinal cord tissues are consistent with the ITB-induced plasticity and trophic actions that have been ascribed to enhanced positive neuroplasticity in spinal cord segments. Our present data demonstrated that TBI induced significant changes in the excitability of the ankle extensor stretch reflex pathways to triceps surae muscles.
It is known that transection of the rat spinal cord reduced the binding of [3H]GABA by 80%. 24 The decrease in GABA binding below the level of SCI suggests that a decrease occurs in the number of GABA receptors. After TBI, we also observed decreased receptor expression in the rat lumbar spinal cord sections. Most of the GABAb receptors, a metabotropic receptor, in the spinal cord, are presynaptic and are located on descending axons, incoming dorsal root afferent axons, and interneurons. 78 –80 Normally, incoming dorsal root information, as well as subsequent interneuronal information, is subject to presynaptic GABA inhibition, which can reduce the amount of primary and secondary excitatory neurotransmitter release. 78 Thus, after ITB, the observed robust increase in immunoexpression of GABAb receptors and GAD67 could, accordingly, enhance the presynaptic inhibition and aid in the reduction of excitatory neurotransmitter release. Our previous studies showed that GAD67 and BDNF colocalize in the same ventral horn motoneurons 16 and further supports the idea that BDNF might enhance synaptic plasticity after ITB.
One of the most consistent findings after experimental TBI has been a prolonged reduction in central NE as well as NE turnover. 81 This is consistent with our previous observations that after experimental TBI, we observed an approximately 56% reduction in DβH-expressing neurons in LC. 14 Moreover, a profound reduction in DβH intermediate reticular fibers in the TBI lumbar spinal cord was also observed. 14 Because the LC provides the noradrenergic support for many CNS neural circuits, many disparate CNS disabilities, including motor, cognitive, and balance functions, are coherently tied to this decrease in central NE level. 82 Descending NE has been reported to decrease the activity of gamma motoneurons acting on group II primary afferents, premotor interneurons, and on gamma-motoneurons themselves. 83,84 Therefore, loss of NE modulation of these afferents, interneurons, and gamma-motoneurons would result in an increase in the excitability of the tonic and dynamic muscle stretch reflexes. Thus, descending noradrenergic projections are known to directly and indirectly modulate motoneuronal excitability by presynaptic inhibition of primary afferent and interneuronal activation of limb motoneurons, 85 as well as modulation of motoneuronal dendritic currents. 86,87 Up-regulation of the central noradrenergic system could increase the modulation of reflex excitability in ITB-treated animals, compared to Sal-TBI controls. Moreover, neuronal circuitry in the spinal cord involved in pattern generation can mediate complex motor function, including walking, and receive tonic stimulation by the bulbospinal NE system. 88 Therefore, up-regulation of spinal NE fibers after ITB could contribute effective regulatory functions of these reflex pathways and contribute significantly to reducing spasticity and enhancing recovery of normal gait patterns. However, up-regulation of NE fiber sprouting observed throughout the lumbar spinal gray matter in ITB-treated animals is a surprising finding, because neurons are not known to coexpress GABA and NE in the spinal cord, although LC neurons colocalize with a number of peptides and BDNF, which can be released along with the principle neurotransmitter, NE. Accordingly, a pharmacological cointeraction between these two systems might exist.
Although many convergent mechanisms are possible, these data suggest that a better outcome could be mediated by up-regulation of the neuroprotective molecule, BDNF, and other inhibitory molecules, such as GABA/GABAb and NE, which play important roles in the regulation of afferent pathways to motoneurons.
In our studies, significantly increased velocity-dependent ankle torques and EMG burst amplitudes and lower-limb spasticity were observed in TBI animals at all postinjury test intervals. Gait-pattern instability was observed when tested at 1 and 3 weeks postinjury using a SSRI analysis. Rotarod testing at 2, 4, and 8 weeks postinjury revealed significant balance disability. Serial learning deficits were observed when tested using an MWM. Our data indicated that ITB treatment blocked early-onset spasticity (tested after 1 week of treatment; 2 weeks postinjury) and significantly attenuated late-onset spasticity (more than 61%) when tested after 3 weeks of treatment. Although we observed two instances of potentially transient adverse effects, overall, these treatment benefits were not accompanied by significant adverse effects on cognition and balance performance. These transient decreases in performance included a drop in performance in the day 2 data for MWM testing and a decrease in rotarod perfomance during the first week of treatment. Because it is known that GABAb receptors down-regulate in the presence of ligand, 23 these transient adverse effects of initial treatment impact may have been softened by receptor down-regulation. Interestingly, acute ITB-treated animals in other testing periods showed a trend of improved scores for serial learning, improved gait performance, and revealed less anxiety-like behavior, when compared to saline-treated TBI controls. Early intervention with ITB treatment was safe, feasible, and effective in this cTBI animal model.
Footnotes
Acknowledgments
This work was supported by Medtronic Inc. and the Department of Veteran Affairs (Rehabilitation R&D Merit Review B6570R; R&D Merit B78071; R&D RCS B7345S).
The authors appreciate Linda M. Page, PharmD, Medtronic Neuromodulation (Minneapolis, MN), for her valuable suggestions.
Author Disclosure Statement
No competing financial interests exist.