
Abstract
Astrogliosis occurs following injury to the zebra finch brain. To date, only estrogen synthase (aromatase) has been identified in injury-induced astrocytes. The expression of other steroidogenic enzymes or their receptors remains unknown in the avian brain. However, in mammals, an upregulation of androgen receptors has been identified in glial cells. The aim of this study was to determine if the androgen receptor is upregulated following injury in adult zebra finches. Finches were given a single penetrating injury and brain tissue was collected 24 or 72 h later. Expression of androgen receptor was examined using immunohistochemistry and quantified using quantitative polymerase chain reaction (qPCR) analysis. Androgen receptors were localized to astrocytes versus neurons, further solidifying the role for astrocytes in neural recovery.
Introduction
N
An example of this divergent vertebrate response following injury is the upregulation of glial aromatase (converts androgens to estrogens). Increased glial aromatase expression is neuroprotective, and functions to modulate apoptosis around the injury site and, therefore, mitigates neurodegeneration 1,20 –22 in both the mammalian 20,23 and avian 24,25 brain. Following injury in rodents, glial aromatase expression is upregulated 24 h following injury, and is undetectable by 30 days post-injury. 26 –28 By contrast, in zebra finches (ZF), increased expression of astrocytic aromatase is observed as early as 6 h following injury, and has been observed as long as 40 days post-injury. 25 Recently, the study of injury-induced aromatase expression has been studied in fish. Interestingly, aromatase B expression was significantly reduced 48 h and 7 days after the injury, in contrast to what is observed in mammals and birds. 29 Given that estrogen response following injury varies across vertebrates, we sought to identify if androgen signaling was also regulated differentially.
Previous research into the role of androgens following injury has focused primarily on aromatizable androgens presumably being converted to estrogens following damage. 21,30 On the other hand, the role for non-aromatizable androgens, such as dihydrotestosterone (DHT), remains elusive. DHT has no effect on reactive astrogliosis after injury; 14 however, it is upregulated in microglia after injury. 31 It is capable of rescuing avian spinal motor neurons from injury-induced cell death by acting directly on these cells via an androgen receptor (AR)-mediated mechanism, 32 and is neuroprotective following diabetic neuropathy in rats. 33 Furthermore, in the uninjured rat brain, DHT increases neuron survival after proliferation within the dentate gyrus during normal development. 34 Conversely, DHT has no influence on hippocampal neuronal survival following injury to mature rats; 30 however, it increases spine synapse density in CA1 hippocampal neurons in female rats. 35 Together, these data suggest a more complex mechanism for androgen-mediated neuroprotection than had been previously thought.
Therefore, another site of estrogen-mediated neuroprotection may be through the regulation of AR and increased androgen signaling. This is further supported by data in the rat brain that show that both estrogen and AR are upregulated following injury. 31 Previous research has shown that estradiol increases androgen signaling via an increase in AR expression in the ZF brain. 36 In the present study, we examined the expression of AR following injury. Adult male and female ZF were given an unilateral penetrating injury to the entopallium. The entopallium is a commonly targeted area for penetrating injuries because of the lack of constitutive steroid hormone production. 24 AR expression was examined using both quantitative polymerase chain reaction (qPCR) and immunohistochemical analysis.
Methods
Birds
Adult male and female ZF (>90 days post-hatching) were obtained from a breeder (Magnolia Bird Farms; Anaheim, CA) and housed in the animal facility at Vassar College. The Vassar College Institutional Use and Animal Care Committee (IACUC) approved all animal procedures.
Injury
Subjects were anesthetized (0.03–0.05 mL/15 g of Nembutal [pentobarbital sodium salt] from Sigma-Aldrich, St. Louis, MO, 25 mg/mL in a 20% propylene glycol, and 5% ethanol solution) and positioned in a stereotaxic apparatus with the head angled at 45 degrees relative to the bifurcation of the Y-sinus. Following a small scalpel incision of the skin, the cranium was exposed and an 18 G needle was used to create two parasagittal openings of the skull on either side of the midline. These openings were 2 mm anterior to the pineal gland and 3 mm lateral to the midline. Injuries were targeted toward the entopallial nucleus 3 mm ventral to the brain surface. A 50 μL 22-s Hamilton syringe needle (Hamilton Company, Reno, NV) was positioned at the surface of the brain at an angle of 45 degrees relative to the Y-sinus, and lowered 3 mm ventrally where it resided for 60 sec. The needle was extracted and the procedure repeated on the other side of the midline. This procedure resulted in one discrete penetrating injury on both sides of the midline. The scalp was repositioned over the cranium and the incision sealed with Collodion Flexible (EM Science, Gibbstown, NJ). After surgery, the birds recovered from anesthesia under a heat lamp.
Immunohistochemistry (IHC)
Whole brain sections from adult birds were removed from the skull and fixed in 5% acrolein at either 24 or 72 h following penetrating injury. Tissue was cut on a freezing microtome at 30 μm after being gel embedded and stored in cryoprotectant at −20°C until use. Following standard IHC protocols, free-floating sections were washed 6×10 min in 0.1M phosphate buffer (PB). The tissue was then treated with 10% sodium borohydride in 0.1M PB. Tissue was then placed in 10% normal goat serum and incubated with primary antibody in 0.1M PB with 0.3% Triton-X (Sigma-Aldrich, St. Louis, MO) for 48h at 4°C. AR N-20 was used at a concentration of 1 μg/3 mL. 37 Tissue was then washed 10×6 min in 0.1M PB with 0.1% Triton-X (PBT) and placed in biotinylated rabbit secondary antibody for 60 min (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA), washed again 3×10 min in 0.1M PBT and then placed in A/B solution (VectaStain, Burlingham, CA) for 60 min. After incubation and a series of washes in 0.1M PBT and 0.175 M sodium acetate sections were then placed in a chromagen solution containing diaminobenzidine tetrahydrochloride (Sigma-Aldrich, St. Louis, MO), hydrogen peroxide, and nickel sulfate (Sigma-Aldrich, St. Louis, MO) to visualize the reaction. Sections were mounted on slides and then cover-slipped after serial dilutions in alcohol and zylene (Fisher Scientific, Pittsburgh, PA). IHC was performed simultaneously on brains to control for any between-run differences in staining procedures.
Co-expression of glial markers and AR
To determine the nature of cells expressing AR, we used double-label immunocytochemistry. Vimentin (Developmental studies Hybridoma Bank, Iowa City, IA) has been previously used as a glial marker in the avian brain. 38 Sections were washed 6×10 min in 0.1M PB, then tissue was treated with 10% sodium borohydride in 0.1 M PB. Tissue was then placed in 10% normal goat serum and incubated in a primary antibody cocktail containing 1:2500 AR and 1:10 anti-vimentin in 0.3% PBT (48 h, 4°C). Tissue was then washed 10×6 min in 0.1M (PBT) to remove excess primary antibody and then incubated in a secondary antibody cocktail containing 1:50 mouse-adsorbed, goat anti-rabbit cyanine-5 (CY-5) and 1:50 rabbit-adsorbed, goat anti-mouse cyanine-2 (CY-2; Jackson Immunochemicals, West Grove, PA) in 0.3% PBT for 2 h at room temperature under foil. Sections were then washed in 0.1% PBT for a total of 2 h, mounted, dehydrated, and cover-slipped.
Confocal microscopy
Sections stained with antibodies against vimentin and AR were examined under a scanning confocal microscope (Leica SP5). Sections were observed under 10× or 63×/1.4 N.A. Plan Apo objectives. Argon ion and 633 HeNe lasers were used to generate the 488 (CY-2) and the 633 (CY-5) lines used for excitation.
Western blotting
The antibody used to detect AR has been extensively characterized. 37 Birds were collected and decapitated after an overdose of isoflurane. The telencephalons were rapidly dissected, quickly frozen in a dry ice methanol bath, and placed at −80°C until use. Brain tissue was homogenized in Hepes buffer (10 mM Hepes, 10 mM KCl, 0.1 mM EDTA, and 200 μL of 10% IGEPAL/5mL of buffer A, 1 μL of protease inhibitor [Sigma-Aldrich, St. Louis, MO]) and centrifuged at 4°C at top speed (15,000–16,000g) for 30 min and placed on ice. Supernatant was immediately removed and placed in −20°C until use. Protein from each sample (20 μL) was then run on 4–12% Tris-HCl gels (Bio-Rad, Hercules, CA) and transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA). Membranes were blocked in 5% nondairy milk for 3 h and incubated overnight at 4°C in primary antibody at 1:1000. Goat anti-rabbit secondary (Jackson ImmunoResearch Inc., West Grove, PA) was applied at 1:3,000 and incubated at room temperature for 1 h, followed by incubation in A/B for 1 h. After a series of washes in Tris-buffered saline, the reaction was visualized with DAB.
qPCR
Birds (n=8) were decapitated 24 h following treatment, and each telencephalic lobe was rapidly dissected out into 1 mL of Trizol reagent (Invitrogen, Carlsbad, CA) and completely homogenized. Total RNA was isolated from the samples using the methods suggested by the manufacturer. Samples were analyzed on a ND-1000 spectrophotometer (NanoDrop, Wilmington, DE), and only those samples that had a 260/280 ratio that exceeded 1.85 were used. For every qPCR experiment, 1 μg of total RNA were reverse transcribed with an iScript cDNA synthesis kit (Bio-Rad, Hercules, CA). For qPCR, 1 μL (or 5% of the total first strand synthesis reaction) of the resulting cDNA was amplified with iQ Syber Green supermix (Bio-Rad, Hercules, CA) in 25 μL of total reaction volume. Assays were performed in 48 well optical plates, and each sample was amplified in triplicate (MiniOpticol Real Time (RT) PCR Detection system, Bio-Rad, Hercules, CA). In every run, wells without the RT product were included, in order to detect any external contamination. Amplicons were generated against exons of the ZF androgen receptor transcript, as well as the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Statistical analysis
mRNA expression data were analyzed using a t test, with treatment as the main variable. Post-hoc analysis was used to determine individual differences. JMP statistical analysis software (JMP, Cary, NC) was used to generate analysis. For all test results, a predetermined significance level of α=0.05 was used.
Results
AR immunoreactivity
Following injury, there was an upregulation of non-neuronal AR-immunoreactive (ir) cells (Fig. 1). Anatomical structural analysis and co-expression studies (Fig. 2) suggest that these cells are astrocytes, because of their differences in shape and size when compared with both neuronal and microglial cells (Fig. 1 B–D). Expression of glial AR-ir cells was localized around the injury, but not found on the ipsilateral side of the brain. Glial AR-ir cells were found in both males and females following injury, suggesting that both males and females are able to upregulate glial AR expression following injury. Western blot analysis revealed a single band of the appropriate size, detected for both male and female injured tissue (Fig. 3).

Photomicrographs illustrating the upregulation of glial androgen receptor expression following injury.

Photomicrographs depicting the overlap of vimentin-positive cells and injury-induced androgen receptor cells. A small population of vimentin-positive cells were also positive for androgen receptors, suggesting that increased expression of androgen receptors following injury is glial and not neuronal. Color image is available online at
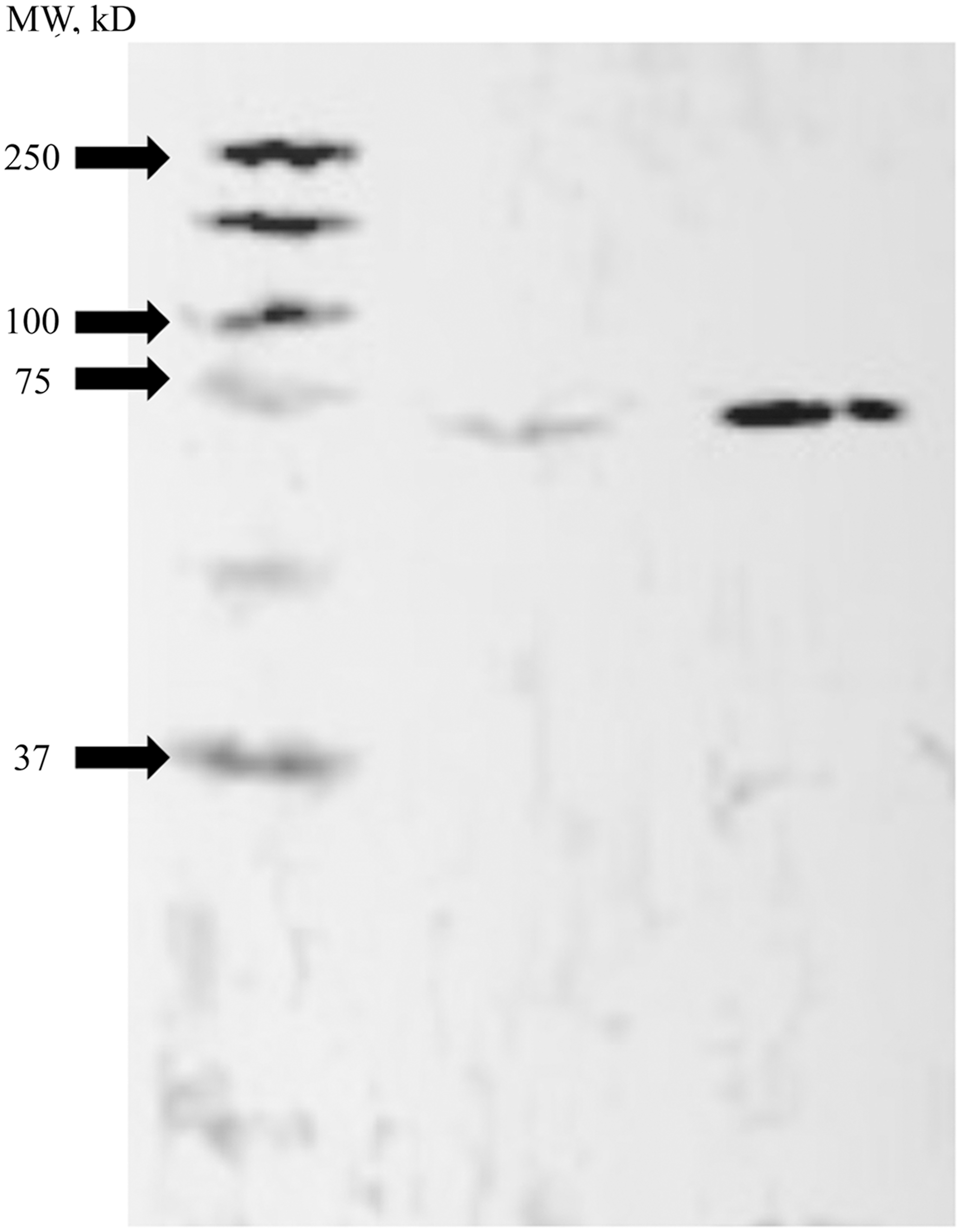
Representative Western blot analysis for injury induced androgen receptor. Western blots were used to confirm the presence of a single androgen receptor band in both females (L) and males (R) following injury.
AR mRNA expression after injury
AR mRNA expression was greater on the injured side of the brain than on the uninjured side (Fig. 4) in males, but not in females. Twenty-four hours after inducing an injury, the injured groups had significantly higher fold changes in AR expression than did uninjured controls (t[13]=2.868, p<0.0100; Fig 4A). However, at 72 h following injury, this effect is no longer significant.

Androgen receptor mRNA expression following injury. Androgen receptor mRNA expression following injury in males and females, analyzed using quantitative polymerase chain reaction (qPCR) at 24 h (L) and 72 h (R). Fold change in expression was calculated using the double delta Ct method assuming 100% efficiency. *Denotes a significant difference between injury and non-injury.
Discussion
In this study, we examined the possible upregulation of AR following injury in the songbird brain. No role for androgens or their receptors have been identified in the avian brain following injury. We found that androgen receptors were upregulated following injury (between 24 and 72 h), and that they were upregulated in astrocytes and not neurons. This is the first time that an increase in AR expression following injury has been presented in the avian brain, indicating that despite circulating levels of androgens remaining unchanged, increased expression of ARs may account for the role that androgens play in mediating neuroprotection following injury.
These data are in agreement with studies by Garcia-Ovejero et al., showing an upregulation of AR following injury. 31 However, in that study, upregulation of AR only occurred in microglia and not in astrocytes as shown here. 31 The difference in cell types is perplexing, and suggests a possible difference in their roles. The role for microglia following injury can be characterized as both neuroprotective and neurotoxic, whereas reactive astrocytes appear to promote neuroprotection alone. This increased neuroprotection is accomplished by regulating neurotransmitter levels, repairing the extracellular matrix, controlling the blood–central nervous system (CNS) interaction as well as transport processes and trophic factors supporting damaged cells. 1,5,39 The upregulation of ARs in astrocytes rather than microglia may reflect a greater role for androgens in neural repair in the avian brain versus initiation of immune process in mammals. Future studies will examine the role that androgen signaling may play in neuroprotection within the avian brain.
Interestingly, both aromatase upregulation and, presumably, the subsequent increase in estrogens occurs prior to the increase in AR expression shown here. The data provide a possible mechanism for the upregulation of AR. 24 However, more work on estrogen mediation of AR following injury is necessary and ongoing to further elucidate this mechanism. These data suggest that non-aromatizable androgens are an important site for neuroprotection, and should be further studied, especially within the avian brain. Also, further clarifying the mechanisms of action between estrogens and androgens is pivotal for our understanding of how the brain naturally repairs itself after injury. Future studies hope to elucidate the role that non-aromatizable androgens may have on rates of neural repair and cognitive recovery.
Footnotes
Acknowledgments
This work was supported by Vassar College.
Author Disclosure Statement
No competing financial interests exist.