
Abstract
Tamoxifen (TMX) is a selective estrogen receptor modulator that can mimic the neuroprotective effects of estrogen but lacks its systemic adverse effects. We found that TMX (1 mg/day) significantly improved the motor recovery of partially paralyzed hind limbs of male adult rats with thoracic spinal cord injury (SCI), thus indicating a translational potential for this cancer medication given its clinical safety and applicability and the lack of currently available treatments for SCI. To shed light on the mechanisms underlying the beneficial effects of TMX for SCI, we used proteomic analyses, Western blots and histological assays, which showed that TMX treatment spared mature oligodendrocytes/increased myelin levels and altered reactive astrocytes, including the upregulation of the water channels aquaporin 4 (AQP4), a novel finding. AQP4 increases in TMX-treated SCI rats were associated with smaller fluid-filled cavities with borders consisting of densely packed AQP4-expressing astrocytes that closely resemble the organization of normal glia limitans externa (in contrast to large cavities in control SCI rats that lacked glia limitans-like borders and contained reactive glial cells). Based on our findings, we propose that TMX is a promising candidate for the therapeutic treatment of SCI and a possible intervention for other neuropathological conditions associated with demyelination and AQP4 dysfunction.
Introduction
A
Estrogen receptors (ER) are expressed both in neurons and glia, 6 including astrocytes 7 and oligodendrocytes. 8 TMX and/or its metabolites bind to ERs and induce tissue-specific changes in their three-dimensional conformation, 9,10 allowing a tissue- and cell- selective recruitment of different transcriptional cofactors, 11 –14 which explains why TMX acts as an ER agonist in the brain and an ER antagonist in others tissues (e.g., breast). In the CNS, TMX also can have diverse effects depending on the cell type, it can mimic estrogen's effects on oligodendrocytes 15 but can antagonize estrogens' effects on astrocytes. 16 Further, TMX can stimulate signaling pathways that are independent of ER activation. 17 For example, TMX inhibits protein kinase C in an estrogen-independent fashion in the brain, and because of this activity, is currently in clinical trials for bipolar disorder. 18 TMX also inhibits swelling-activating anion channels 19 in an ER-independent fashion, which is believed to contribute to its neuroprotective effects in brain ischemia. 20 Beneficial effects of TMX also have been demonstrated in such other neuropathological conditions as traumatic injury to the central and peripheral nervous system, stroke, multiple sclerosis, Parkinson's disease, Alzheimer's disease, cognitive decline, and mood disorders, 21 including SCI. 22,23
Our goal in this study was to test the translational potential of TMX in a rat model of moderate contusion SCI, and to use proteomic, biochemical and histological analyses to shed light on the mechanisms underlying TMX's beneficial effects in injured spinal cords.
Methods
Contusion injury
Male Sprague-Dawley rats (Harlan, 225–250 g) were anesthetized with pentobarbital (50 mg/kg, intraperitoneal); our procedure was detailed in an article by Durham-Lee and colleagues. 24 Animals were contused at the tenth thoracic segment (T10) with an Infinite Horizons Impactor using a force of 150 kdynes (1 sec dwell time). All procedures complied with the recommendations in the National Institutes of Health's Guide for the Care and Use of Laboratory Animals and were approved by the University of Texas Medical Branch (UTMB) Animal Care and Use Committee. Control age-matched animals were not subjected to any part of the surgical or post-surgical care protocols. We use only naïve rats as controls, as we discussed in Durham-Lee and colleagues. 24
Tamoxifen treatment
Timed-release tamoxifen pellets (Innovative Research of America; Catalog #E-351) were surgically implanted subcutaneously (on the lateral side of the neck between the ear and shoulder) 2 h after SCI as a clinically relevant time for drug administration. Drug administration via pellets was advantageous because it significantly reduced the distress of animals versus prolonged daily intraperitoneal or intravenous injections; one of our goals was to test different durations of TMX delivery. In addition, subcutaneous pellets would have an advantage even for clinical applications. The TMX pellets were designed for a constant delivery rate of 1mg/day for 14 days or 28 days. Tamoxifen pellets have been used in several other animal studies (listed on the manufacturer's web site:
Assessing possible adverse effects of tamoxifen
TMX can cause liver cancer in rats, 27 unlike in humans. Although TMX doses that can cause adverse effects in normal rats are 10 times higher than used in our study, 27 we assessed TMX's tumorigenic activity in SCI rats, since their drug metabolism is altered, and thus their susceptibility to drugs' adverse effects higher. Therefore, we weighed the livers in both groups of SCI rats (35 days after SCI), but we found no significant differences between groups. The average weights of livers in naïve (n=10; standard deviation), control SCI, and TMX-treated SCI rats are in the table below. Further, gross histological assessments of liver and intestinal organs performed by a UTMB veterinarian showed no pathological alterations.
Locomotor assessment
Hind-limb movement was assessed using the Basso, Beattie, and Bresnahan (BBB) Scale. 28 BBB scores were collected daily on the first 14 days after injury, and once weekly thereafter.
Protein extraction
Animals were anesthetized by intraperitoneal injection of pentobarbital (150 mg/kg) and sacrificed by transcardial perfusion with 0.9% saline containing heparin. T10 (lesion epicenter) and lumbar (L1-L5) spinal cord segments were removed and stored at −80°C. For protein extraction, we used ProteinExtract™ complete Mammalian Proteome Extraction Kits (Calbiochem Cat No. 539779). Protein concentrations were determined using the RC DC Protein Assay Kit (BioRad cat #500-0122).
Proteomic separations and analysis
Proteins were analyzed by two-dimensional gel electrophoresis as described previously. 29,30 See supplementary material for a detailed description.
Electrophoresis and Western blotting
As detailed by Guptarak and colleagues, 31 samples containing 40 (g of protein were mixed with an appropriate volume of 6×super-denaturing sample buffer and equal amounts of protein were loaded onto a sodium dodecyl sulfate-polyacrylamide gel; the samples were not boiled to avoid aggregation of membranous proteins.
Immunofluorescence
The method is detailed by Guptarak and colleagues. 31 Spinal cords were removed, postfixed overnight at 4°C in 4% paraformaldehyde, and cryoprotected in phosphate buffer with 30% sucrose for three days. The sections were washed in 0.05M TBS mounted on gelatin-coated glass slides, and dried before the addition of mounting medium with the nuclear stain 4',6-diamidino-2-phenylindole, dihydrochloride (DAPI; Vector Laboratories H-1200, Iowa City, IA).
Primary antibodies
Primary antibodies (Table 2) used were: mouse anti-CC1 (Abcam # ab16794 1:100, Cambridge, MA), a mature oligodendrocyte marker APC (or CC1) that is an antagonist of β-catenin and results in a reduction in Wnt signaling 32 ; rabbit anti-aquaporin 4 (AQP4; Millipore/AB 3594 1:10,000, Billerica, MA); mouse anti-3CB2 (Hybridoma bank; 1:1000, Iowa City, IA), an intermediate filament-associated protein specifically expressed in embryonic radial glia 33 ; and mouse anti-Iba-1 (Wako Pure Chemicals 1:1000, Osaka, Japan), an ionized calcium binding adaptor molecule 1 that is specifically expressed at low levels in resting microglia but at high levels in activated microglia/macrophages.
Image acquisition and analysis
Images were collected using a Zeiss LSM-510 META confocal microscope with a 10×0.3 numerical aperture (NA) lens and a 63×oil 1.4 NA objective (Optical Microscopy Core at UTMB). The images were obtained using excitation lines at 364, 488, and 543 nm and three different channels of emission with sequential acquisition. 3D rendering was done using Imaris Premier 7.5 software. See supplementary material for a more detailed description.
Statistical analysis
All statistical tests were evaluated at the α level of 0.05, with two-tailed t-test, using the SPSS program. For multiple-group comparisons, data were analyzed using analysis of variance. The LSD multiple comparisons post-hoc test was used to determine p values (<0.05).
In all our graphs, “#” is used to denote significant difference (p<0.05) between uninjured rats and control SCI rats and “*” to denote significant difference (p<0.05) between control SCI rats and TMX-treated SCI rats.
To identify differentially expressed gel spots in the proteomic analyses we used t-tests to compare the (log 2 transformed) intensity of spots. The comparisons were first made for uninjured versus SCI, with p values produced for each compared spot. Because of the large number of the spots (1119), we adjusted the p values for multiple-testing procedure, using the Benjamini-Hochberg model. 34 In brief, the method first sorts all p values in ascending order. Then every p value is modified to have a new (interim) value, i*p_value/N, where, i is the index (in the sorted data) of the original p value, and N is total number of the null hypothesis. The adjusted p values are determined as cumulative minima in the array of interim p-values. The BH adjustments to the p-values were done using the p.adjust function of R. 35
Results
Acute TMX administration
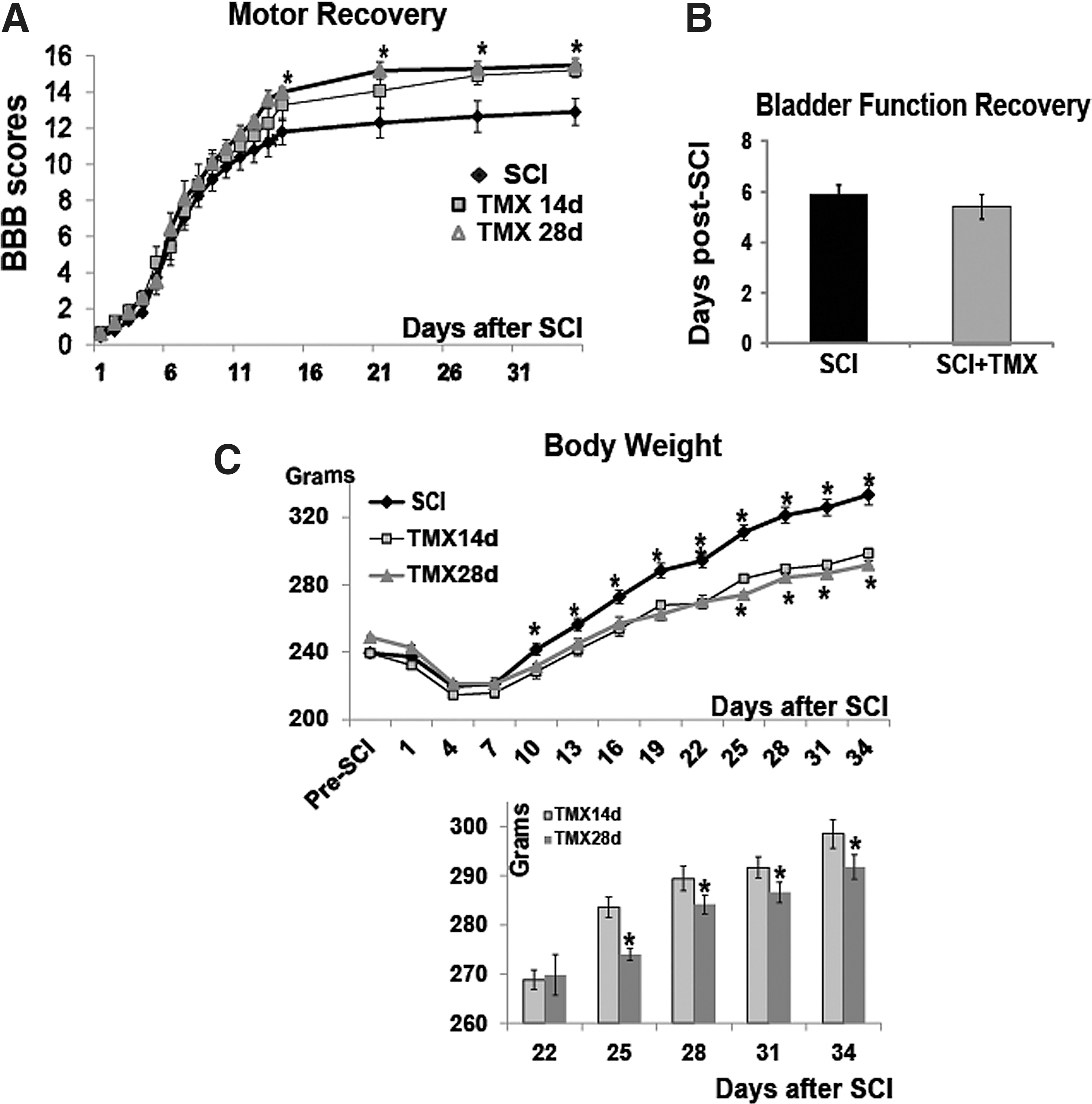
TMX improved the locomotor recovery of SCI rats. SCI rats were divided into three experimental groups: (1) SCI rats that received no treatment (n=10); (2) control SCI rats that received a placebo pellet releasing vehicle (n=10); and (3) SCI rats that received TMX pellets (n=20). We measured locomotor recovery of vehicle- and TMX-treated SCI rats using the BBB test (Fig. 1A). We did not find statistically significant differences in the BBB scores between SCI rats that received no treatment and those that received placebo pellets. Therefore, those two groups of SCI rats were combined into one control group (n=20). In all graphs presented here, control group of SCI rats is labeled as “SCI,” while SCI rats treated with TMX are labeled as “TMX.”
Tamoxifen

TMX was delivered daily (1 mg/rat) for either 14 days or 28 days; (n=10/group). As shown in Figure 1A, both groups of TMX-treated SCI rats (14 and 28 day delivery), demonstrated a delayed, but significant improvement in hind-limb locomotion. Although improvements in BBB scores were similar, the 28-day TMX delivery group of SCI rats appeared to achieve faster locomotor recovery than the 14-day delivery group. The maximal improvement of ∼3 BBB score units was achieved 35 days after SCI in both TMX-treated groups. In contrast to control SCI rats, all TMX-treated SCI rats showed a coordinated gait at 35 days after SCI. The same difference in the BBB scores at 28 and 35 days after SCI was confirmed by another experimenter, also blinded to drug treatments.
Bladder function recovery occurred at about six days post-SCI, in control SCI and all TMX-treated SCI rats (14-day and 28-day delivery groups were plotted together; Fig. 1B) Therefore, TMX did not affect bladder recovery of SCI rats.
It is well established that continuous administration of TMX induces a reduction in body weight. 36 –39 Therefore, we measured the body weight of all SCI rats as an indicator of the effectiveness of TMX delivery via pellets. TMX-treated SCI rats had markedly and significantly reduced body weight versus control SCI rats (Fig. 1C). Further, weight gain in SCI rats treated with TMX for 14 days was accelerated beyond 22 days post-SCI (shown in the bar graph in Fig. 1C), compared with SCI rats treated with TMX for 28 days, indicating that this TMX effect remained for about one week after TMX pellets ceased to release the drug.
Delayed TMX administration
TMX did not affect the locomotor recovery of SCI rats. TMX pellets (1 mg/day/rat) were subcutaneously inserted 42 days after SCI, and delivered TMX for 14 days (until 56 days post-SCI). Similarly injured SCI rats were divided into three groups 42 days after SCI: 1) SCI rats that received no treatment (n=10); 2) control SCI rats that received a placebo pellet (n=15); and 3) SCI rats that received TMX pellets (n=19).
In contrast to acute TMX administration, delayed TMX delivery did not improve locomotor recovery (Fig. 1D; SCI rats that received placebo pellet and SCI rats that received no treatment were again combined into one group). Similar to acute TMX intervention, delayed delivery of TMX also suppressed the weight gain in SCI rats (Fig. 1E), indicating that the lack of beneficial TMX effects on locomotor recovery of SCI rats did not result from ineffective TMX pellets.
Proteomic analyses
TMX affected the abundance of proteins expressed in oligodendrocytes and astrocytes. Proteomic analysis was performed with proteins (total cell extracts) isolated from the lesion site (T10) 35 days after SCI in three experimental groups: 1) SCI rats treated with TMX for four weeks (n=5), 2) control SCI rats (no treatment; n=5); and 3) naïve age-matched rats that received no treatment (n=5). The protein spots from all gels were matched, and an abundance value for group 1 calculated versus naïve and control SCI rats. Fluorescence staining revealed 1118 spots; only spots whose abundance satisfied the following criteria were selected for identification by mass spectrometric analysis (MS): 1) abundance changed significantly after both SCI (uninjured vs. SCI; p<0.05), and TMX treatment (SCI vs. TMX; p<0.05), or 2) abundance did not change significantly after SCI (uninjured vs. SCI; p≥0.05) but it did after TMX treatment (SCI vs. TMX; p<0.05).
Our statistical analysis identified 25 spots that were excised and processed for MS identification; no proteins were found where abundance was affected by TMX but not affected by SCI, suggesting that TMX predominantly affected the abundance of proteins altered by SCI. The identities of the 25 selected spots are shown in Table 2. The abundance of the first ten proteins in Table 2 was significantly decreased in SCI rats (fold change SCI vs. naïve <1), and significantly increased in TMX-treated SCI rats (fold change TMX vs. SCI >1). We also identified 15 proteins (see Table 2) where levels were significantly increased in SCI rats (fold change SCI vs. naïve >1), and then significantly changed in TMX-treated SCI rats.
Ab, antibody.
MW, molecular weight; No., number; SCI, spinal cord injury; N, naive; TMX, tamoxifen; NCBI, National Center for Biotechnology Information.
Among proteins where abundance was significantly decreased after SCI and then partially restored by TMX, we found markers of oligodendrocytes, such as 2′,3′ -cyclic nucleotide 3′-phosphodiesterase (CNPase), 40 and of oligodendrocyte progenitors (stathmin), 41 as well as myelin (enoyl co-A). 42 These findings indicate a beneficial effect of TMX on oligodendrocytes and myelin levels, of which loss at the lesion site substantially contributes to functional impairment after SCI.
Among proteins that were significantly elevated after SCI and then decreased by TMX treatment, we found markers of astrocytes and their activation, such as glial fibrillary acidic protein (GFAP isoforms), fascin (intermediate filaments bound to GFAP 43 or D-3-phosphoglycerate dehydrogenase, 44 which suggests that TMX ameliorates the activation of astrocytes at the lesion site. To confirm the results of our proteomic analysis we performed Western blots (Fig. 2 and Fig. 4).

Tamoxifen (TMX) restores myelin loss at the lesion site.
TMX increased myelin levels at the lesion site
In Figure 2A, we show representative images of 2D gels: A1, age-matched uninjured T10; A2, T10 in control SCI (35 days after SCI); and A3, T10 in SCI rats treated with TMX (35 days after SCI). Figure 2B presents an enlarged region containing spot #498 (identified as CNPase, a myelin-associated enzyme). The bar graph in Figure 2C shows quantitative analysis of normalized CNPase abundance levels obtained from proteomic analysis in 15 2D gels (n=5/group), indicating a significant decrease in CNPase levels at the lesion site 35 days after SCI (#; p<0.05) and a partial but significant reversal in TMX-treated SCI rats (*; p<0.05). We also used Western blots to measure CNPase protein levels at T10 in naïve, control SCI, and TMX-treated SCI rats (n=5/group; Fig. 2D and Fig. 2E). TMX treatment was identical in all SCI rats used for both proteomic and Western blot analyses. All Western blot analyses were repeated twice by two experimenters, and only results confirmed in two independent experiments are presented here. Consistent with our proteomic analyses, Western blots also showed SCI-induced CNPase decreases (an ∼40 kDa band, Fig. 2D) at T10 35 days after SCI (#; p<0.05) and a partial but significant reversal with TMX treatment (*; p<0.05; Fig. 2D and Fig. 2E). We found that CNPase levels were decreased at the lesion site by ∼50% in proteomic analyses (Fig. 2C), and by ∼60% in Western blot analyses (Fig. 2D and Fig. 2E). TMX treatment significantly increased CNPase levels at the lesion site by 30% in Western blots, and by 20% in the proteomics study.
To further confirm the beneficial effect of TMX on myelin loss after SCI, we also measured protein levels of myelin basic protein (MBP), the most abundant myelin protein in mature oligodendrocytes/axons. Using Western blots, we found a ∼50% decrease in MBP levels at the lesion site 35 days after SCI (#; p<0.05; Fig. 2F), consistent with our CNPase analyses (Fig. 2C and Fig. 2E). We also found that TMX significantly increased MBP levels by 20% (*; p<0.05, Fig. 2D). The late-appearing myelin oligodendrocyte glycoprotein (MOG) is a marker of the final steps in the myelination processes. MOG levels were significantly decreased by >60% at the lesion site 35 days after SCI (#; p<0.05; Fig. 2 D and Fig. 2G), and significantly restored in TMX-treated SCI rats, by ∼20% (*; p<0.05; Fig. 2D and Fig. F), in agreement with the results in Figures 2C, 2E, and 2F.
We also tested if TMX treatment affected myelin levels in lumbar segments (pooled L1-L5). Lumbar segments directly control hind-limb locomotion. 45,46 Therefore, in addition to myelin changes at the lesion site, myelin changes in lumbar segments also could affect the recovery of hind-limb function after SCI. As shown in Figure 2H, we analyzed MOG levels in lumbar segments of the same SCI rats whose T10 segments were used for the analyses shown in Figure 2D. MOG levels in the lumbar segments of SCI rats were not significantly affected by either SCI or TMX, suggesting that TMX does not affect myelin levels in the absence of injury-induced processes, in agreement with our proteomic analysis showing no effect of TMX on proteins unaltered by SCI.
TMX spared oligodendrocytes
The CC1 antibody recognizes adenomatus polyposis coli, and is a marker of oligodendrocyte cell bodies without labeling myelin. 47 CC1-labeled oligodendrocytes were assessed in three groups of rats: 1) naïve rats (Fig. 3A); 2) SCI rats that received vehicle Fig. 3B); and 3) SCI rats that received TMX (1 mg/day) for 14 days (Fig. 3C; n=5/group). We found significantly increased CC1 immunolabeling in all TMX-treated SCI rats (Fig. 3D), indicating an ∼20% increase in the number of mature oligodendrocytes/μm2 in TMX-treated SCI rats, consistent with ∼20% preservation of myelin (Fig. 2). Here, we show increased CC-1 labeling in TMX-treated SCI rats even when cavities at the lesion site had sizes similar to those in control SCI rats (Fig. 3B and Fig. 3C), suggesting that TMX had a sparing effect on oligodedendrocytes independent of reduced cavitation. Our quantitative assessment of cavity sizes in AQP4-immunolabeled lesion sites (AQP4 delineated cavity borders; see Fig. 5) showed significant (*p=0.0035) 1.83-fold smaller lesion sizes in TMX-treated injured spinal cords (n=5/group; Fig. 3F).
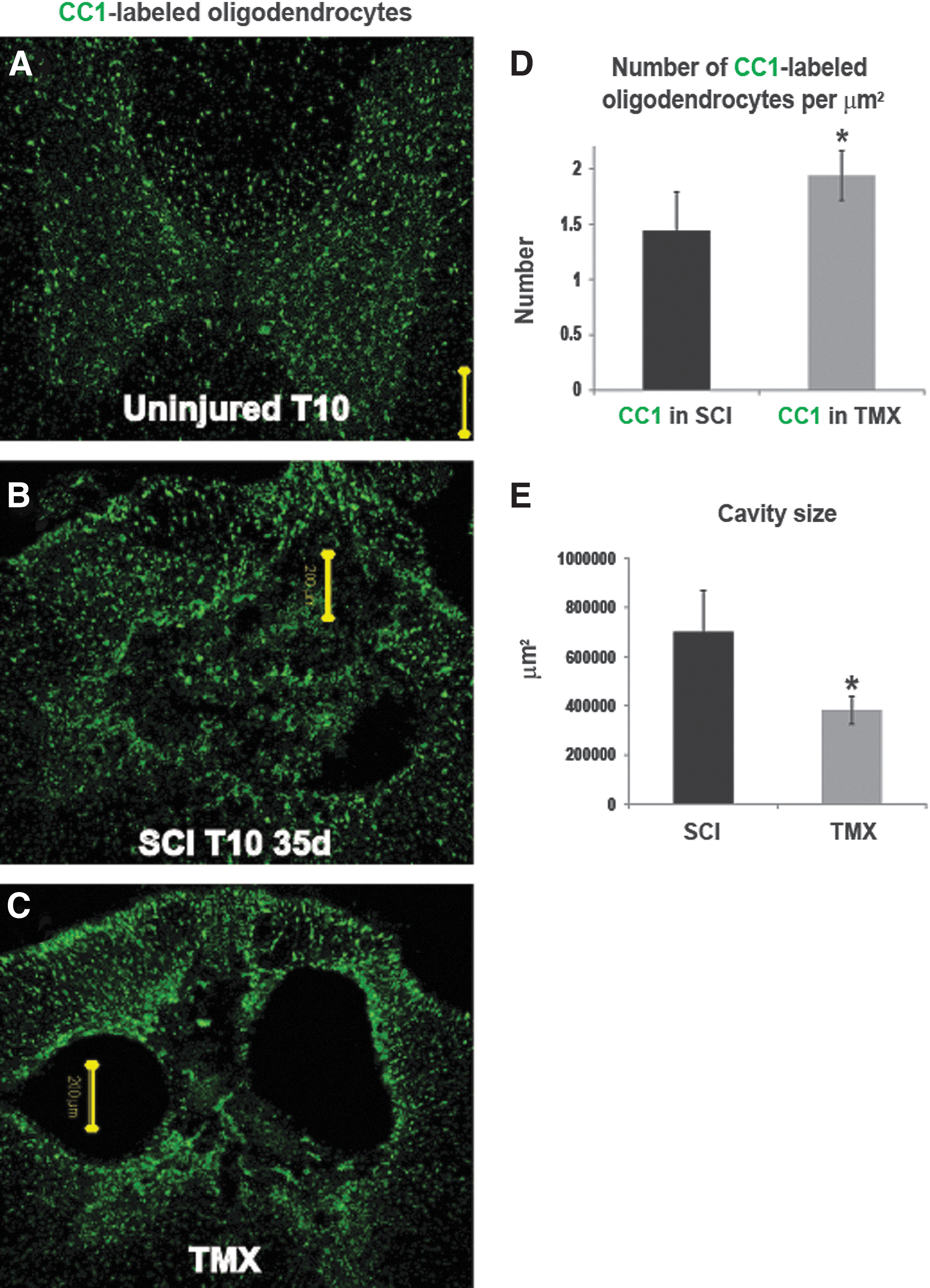
Tamoxifen (TMX) spares oligodendrocytes and reduces cavitation. Representative confocal images of the CC1-immunolabeling at the 10th thoracic segment (T10), 35 days after spinal cord injury (SCI), in three experimental groups, an age-matched uninjured rat,
TMX altered astrocytic activation at the lesion site
Proteomic analysis identified 48 and 49 KDa GFAP bands (Table 2, Fig. 4A and Fig. 4B) as being significantly increased after SCI by ∼80-120% (#; p<0.05; Table 2; Fig. 4B), and decreased by ∼50% in TMX-treated SCI rats (*; p<0.05; Table 2; Fig. 4B). As shown in Figure 4B, GFAP48kDa levels in TMX–treated SCI rats were indistinguishable from those in uninjured spinal cords, suggesting that TMX treatment abolished astrocytic activation at the lesion site. Western blot analysis (Fig. 4C) confirmed the proteomic findings. SCI-induced increases in several GFAP bands, including the ∼50kD band (marked with black arrow) and GFAP bands with lower molecular weight (MW; between 45 and 50 kDa, marked with gray arrow) were seen in the same Western blots with a longer exposure time. We quantified all GFAP bands together (Fig. 4D). SCI induced significant increases in GFAP bands, by ∼70% (#; p<0.05). TMX treatment abolished the SCI-induced increases in GFAP levels (*; p<0.05; Fig. 4D), although it more prominently affected the intensity of the lower MW bands (<50kDa; Fig. 4C). Interestingly, vimentin, another intermediate filament protein strongly increased in activated astrocytes after SCI (Fig. 4C), was not affected by TMX, suggesting that TMX selectively affects different targets in reactive astrocytes in injured spinal cords.

Tamoxifen (TMX) affects reactive astrocytes.
AQP4 is another exclusively astrocytic protein that is significantly upregulated in activated astrocytes in chronically injured spinal cords, 48 consistent with our findings here (Fig. 4E, 4F, and 4G). We also found that AQP4 levels were even more increased in TMX-treated SCI rats (Fig. 4E, 4F, and 4G). As shown in Figure 4F, we found that TMX treatment elevated levels of the AQP4 M23 isoform by ∼50%, and of the M1 isoform by ∼20% (Fig. 4G) at the lesion site 35 days after SCI.
In contrast to unchanged myelin levels in lumbar cord segments, GFAP levels in L1-L5 were mildly but significantly increased 35 days after SCI (Fig. 4H). However, TMX did not alter the GFAP levels, indicating that pathways upstream of the GFAP increases in lumbar segments are likely different from the signals that trigger GFAP increases at the lesion site. AQP4 levels in lumbar segments were unaffected by SCI (Fig. 4I), again suggesting that GFAP and AQP4 do not share the same upstream processes. AQP4 levels in lumbar segments were not altered by TMX (Fig. 4I), consistent with TMX affecting only injury-induced processes.
TMX altered the organization of cavity walls delineated by AQP4-expressing astrocytes
The most surprising result of the AQP4 immunolabeling at the lesion sites in control versus TMX-treated SCI rats was the difference in the shape of the cavities delineated by AQP4-labeled astrocytes. Figures 5B-B2 and 5C-C2 represent three sections 40 microns apart taken from the same T10 segments of control (B-B2) and TMX-treated SCI rats (C-C2). Cavities in TMX-treated SCI rats were more rounded, with walls consisting of densely packed AQP4-expressing astrocytes (white arrow in Fig. 5C1), whereas the cavities in control SCI rats were irregularly shaped with unorganized, partially opened walls (white arrow in Fig. 5B1). The same result was obtained in all analyzed control- and TMX-treated SCI rats (n=5/group). High magnification images and the corresponding 3D renderings of lesion wall segments (labeled with white squares in Fig. 5B2 and 5C2) in control- and TMX-treated SCI rats (Fig. 5E and Fig. 5F) show well-organized cavity walls in TMX-treated SCI rats, with densely packed astrocytes abundantly expressing AQP4 that closely resembled the organization of the glia limitans externa (Fig. 5G).

Aquaporin 4 (AQP4) and the organization of the lesion wall.

TMX prevented spreading of cells into cavities
Well-organized lesion walls in TMX-treated SCI rats were associated with cavities devoid of cells (with DAPI- stained nuclei; Fig. 5C-C2 and Fig. 6B, 6D, and 6F), in contrast to cavities in control SCI rats (Fig. 5B-B2 and 6A, 6C, and 6E). We quantified DAPI-labeled nuclei only within the lesion in control- (n=5) and TMX-treated SCI rats (n=5; see Fig. 6E and 6F), and found that control SCI rats had ∼3 fold more nuclei/μm2 than TMX-treated SCI rats (Fig. 6G). Figures 6E and 6F are representative examples of cell infiltration within cavities in control and TMX-treated SCI rats, respectively. More interestingly, we found that cavity walls characterized by high AQP4 levels and glia limitans-like organization (thin white arrows in Fig. 6B) were associated with little or no cell penetration into the lesion cavity, not only in TMX-treated SCI rats (thin white arrow in Fig. 6F), but also in control SCI rats (thin white arrows in Fig. 6A and 6E), although such an organization of cavity walls was rare in control SCI. Conversely, segments of disorganized lesion walls in TMX-treated SCI rats that were not characterized by high AQP4 levels (thick white arrows in Fig. 6B and 6F) also were associated with infiltration of cells into the lesion. Although we have not performed a detailed characterization of cells infiltrating into the cavities, we identified CC-1 labeled cells (see Fig. 3B), AQP4-labeled cells (Fig. 6A) but also immature radial glia–like reactive astrocytes labeled with 3CB2 antibody and reactive microglia labeled with Iba-1 (Fig. 6H).

Spreading of cells within the lesion.
Discussion
Translational potential of tamoxifen
Based on the criteria outlined in the review by Tator and colleagues 49 and Guest and colleagues, 50 we summarized our results in Table 3, which suggest that TMX may be a promising intervention for the treatment of SCI. For example, TMX readily crosses the blood-brain barrier, 51 –53 and therefore can be systemically administered, which has a clear clinical advantage. Further, the safety profile of TMX has been well established in the past four decades. TMX is approved by the United States Food and Drug Administration not only for the long-term (five-year) daily treatment of breast cancer patients but is also safe enough to be approved as a long-term preventive agent in women at high-risk for breast cancer. Adverse effects of TMX (increased risk of endometrial cancer and thromboembolic diseases; ∼1.2 cases per 1000 women) strongly depend on the duration of its use. 54 Therefore, the short-term TMX therapy proposed here may be considerably safe, particularly in male SCI patients, who constitute ∼80% of all SCI patients. Our analyses also supported the safety of 1mg/kg TMX administered for two weeks to SCI rats; it did not cause adverse liver changes and it did not affect proteins in uninjured spinal cords.
SCI, spinal cord injury; T10, tenth thoracic segment; SQ, subcutaneous; BBB, Basso, Beattie, and Bresnahan locomotive rating scale; WB, Western blot.
Even though the same TMX intervention paradigm used in our experiments was not repeated by another research group (reproducibility increases translational potential of pharmacological interventions) 49 beneficial effect of a similar TMX dose (5mg/kg) on motor recovery of SCI rats has already been shown by Tian and colleagues, 23 who administered one TMX injection 30 min post-SCI. Although our study tested the effect of TMX in a more clinically applicable time window (starting 2 h after SCI), the results of both studies suggest that TMX has a clear translational potential as a pharmacological intervention for SCI. Further, pharmacological interventions currently in clinical trial for SCI (minocycline or riluzol) or interventions that have translational potential (nimodipine, magnesium sulfate, or glyburide) 49 have demonstrated either smaller or equivalent improvements in locomotor recovery as TMX. However, multimodal therapeutic targets of TMX (estrogen receptor and non-estrogen receptor-mediated pathways) are not affected by either of the promising pharmacological interventions mentioned above, and may explain a unique neuroprotective effect of TMX, discussed below.
Proteomic analysis of TMX treatment. Given that TMX can affect many cell types in the CNS, and stimulate a plethora of signaling pathways (as described in the Introduction), we used a global proteomic analysis to shed light on the mechanisms underlying beneficial effect of TMX in SCI. The results of our proteomic analysis (summarized in Table 2) are consistent with the proteomic analyses of TMX effects in breast cancer cells, which, similar to our study, also identified stathmin, protein disulfide isomerase A3, peroxiredoxin, and GFAP as TMX targets, 55 thus validating our results. However, the initial goal of our proteomic analysis was to identify markers of cell populations affected by TMX, rather than to identify a complete list of proteins affected by TMX because of the inherent limitations of the proteomic method that generates a large number of false negatives. For example, we found that TMX significantly affected MBP, MOG and AQP4 levels when we used Western blots but not when we analyzed proteomic data. This can be explained by either the well-known lower sensitivity of the proteomic analysis versus Western blots or by the stringency of our statistical analysis. Because we used corrections for multiple comparisons, we eliminated from the analysis a large number of proteins (1118-25=1093 spots), in part because of the intra-group variations, which if analyzed for individual proteins, might have rendered some differences between treatments groups significant. As our proteomic analysis identified several oligodendrocytic and astrocytic proteins as being significantly affected by TMX (see Table 2), we focused our further analyses on those two cell populations, although the effect of TMX on other cell types present at the lesion site 35 days after SCI cannot be ruled out.
TMX effects on oligodendrocyte/myelin levels can explain the improvement in BBB scores. We have shown that loss of myelin levels at the lesion site in chronically injured spinal cords is correlated with BBB scores, 56 in agreement with other findings. 57 Further, we have also demonstrated that even modest increases in myelin (i.e., MOG) levels by ∼12% at the lesion site lead to substantial improvements of locomotor recovery. 56 Therefore, our finding that TMX increases levels of all major myelin proteins (MBP, CNPase, MOG), and oligodendrocyte numbers by ∼20% at T10 can explain the observed improvement in hind-limb motor recovery (by ∼3 BBB scores).
We have previously shown that acute loss of myelin at the lesion site (∼40% decrease) occurs within the first 24 h after SCI, followed by additional >40% decrease that is finalized by seven days after SCI, 56 consistent with a delayed wave of oligodendrocyte death. 58 –60 Given the delayed locomotor improvement of TMX-treated SCI rats (14 days), and a lack of a TMX effect on the bladder function recovery (occurred at ∼6 days), it is likely that TMX reduced the delayed loss of oligodendrocytes, and the resultant loss of myelin and axonal demyelination/dysfunction that is critically important for the functional recovery after SCI. 57 Although the effect of TMX on oligodendrocyte survival after trauma has not been studied, Lee and colleagues, 61 have shown that 17β-estradiol reduces oligodendrocyte cell death by inhibiting JNK3 activation, a pathway that is also implicated in oligodendrocyte apoptosis after SCI. 62 Given that TMX mimics the effects of 17β-estradiol on oligodendrocytes, 15 it is likely that TMX prevented delayed apoptotic death of oligodendrocytes after SCI.
Novel TMX effect on astrocytic AQP4
It has been shown that TMX decreases the activation of astrocytes in different conditions, 63 including SCI. 23 However, our data suggest that TMX did not decrease overall astrocytic activation as vimentin levels were not affected while AQP4 levels were further increased, consistent with TMX having complex effects on reactive astrocytes that can be either ER-mediated (agonistic 64 or antagonistic 16 ) or non-ER-mediated. 20
Although the role of estrogen in the regulation of AQP4 expression has not been studied, there are reports that would indicate that estradiol (presumably through the estrogen receptor-mediated pathways) can affect AQP4 levels. For example, estrogen withdrawal results in reduced AQP4 levels in rat brains. 65 However, exogenous administration of estradiol also decreases AQP4 levels in ischemic brain. 66 Given that published findings linking AQP4 and ER are scarce and controversial and that our study is the first to report the effect of tamoxifen on AQP4 levels in injured spinal cords, it remains to be determined if TMX effects on AQP4 levels are ER- or non–ER-dependent.
We have already demonstrated that higher AQP4 levels in chronically injured spinal cords are associated with better functional recovery after SCI, 67 in agreement with an AQP4 deletion study showing that a lack of AQP4 impairs locomotor recovery in SCI mice. 68 However, the role of AQP4 increases in chronically injured spinal cords is not known. Our data indicate that AQP4 increases in TMX-treated chronically injured SCI rats were associated with the formation of a glia limitans–like organization of the cavity walls and smaller fluid-filled cavities, a novel finding. We hypothesize that increased AQP4 may facilitate specific “stacking” arrangement 69 of the glia limitans subtype of astrocytes, 70 which could prevent fluid accumulation and/or spreading of reactive glia (see Fig. 6H), and thus limit progressive cavitation after SCI. Interestingly, Kimura and colleagues 68 have shown that AQP4 knockout SCI mice not only recover locomotion more poorly than wild type SCI mice but also form fluid-filled cavities, 68 while wild type SCI mice never form cysts, further supporting our hypothesis that TMX-mediated AQP4 increases limit the expansion of the fluid-filled cavities.
Our data also suggest that TMX increased levels of the M23 isoform ∼2.5 times more than levels of the M1 AQP4 isoform. In contrast to M1, M23 isoform is capable of assembling into large and immobile orthogonal arrays of particles (OAP) 71,72 that may contribute to the specific stacking arrangement possibly through intercellular OAP interactions among glia limitans–forming astrocytes. It is also interesting that irregularly shaped, “open” cavities containing cells, as described in our control SCI rats, closely resembled lesions in humans SCI associated with astrocytes not expressing AQP4, 67 suggesting that more profound dysfunction of AQP4 after SCI in humans may be one of the factors contributing to the vastly poorer functional recovery than in rodent models of SCI characterized by AQP4 upregulation.
Together, our data strongly suggest that TMX delivery should last for two weeks after SCI, and that during the first week post-injury, TMX alleviates protracted death of oligodendrocyte and thus prevents debilitating demyelination, while in the second week, it limits cavitation by upregulating AQP4—all processes leading to the improved locomotor recovery of SCI rats. Therefore, we propose TMX as a promising therapeutic intervention for SCI. However, we also believe that the effect of TMX on myelin levels and its novel effect on AQP4 may also be potentially useful for the treatment of other demyelinating diseases particularly if associated with the loss of both myelin and AQP4, such as neuromyelitis optica (Devic's disease). 73
Footnotes
Acknowledgments
We thank Dr. Jason Vellano for the gross morphological and histological assessments of the liver and GI tract in our experimental animals. We also thank Dr. David Konkel for critically editing the manuscript.
Author Disclosure Statement
This work was supported by Mission Connect, a program of the TIRR Foundation. No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.