
Abstract
Progesterone is an anti-inflammatory and promyelinating agent after spinal cord injury, but its effectiveness on functional recovery is still controversial. In the current study, we tested the effects of chronic progesterone administration on tissue preservation and functional recovery in a clinically relevant model of spinal cord lesion (thoracic contusion). Using magnetic resonance imaging, we observed that progesterone reduced both volume and rostrocaudal extension of the lesion at 60 days post-injury. In addition, progesterone increased the number of total mature oligodendrocytes, myelin basic protein immunoreactivity, and the number of axonal profiles at the epicenter of the lesion. Further, progesterone treatment significantly improved motor outcome as assessed using the Basso-Bresnahan-Beattie scale for locomotion and CatWalk gait analysis. These data suggest that progesterone could be considered a promising therapeutical candidate for spinal cord injury.
Introduction
A
Effective treatments should ideally act on part or all of those events to finally spare tissue and preserve axonal tracts, oligodendrocytes, and myelin sheaths. No gold standard therapy for SCI has been established, 4 –6 although clinical trials with methylprednisolone (NASCIS II and III) have demonstrated modest therapeutic benefits. 7,8
Progesterone (PROG) could represent a good candidate for therapy, because it is neuroprotective, promyelinating, and anti-inflammatory in pathologies of peripheral and central nervous systems. 9 –12
Specifically, in experimental brain trauma, PROG reduces edema and inflammatory cytokines, prevents neuronal loss and mitochondrial dysfunction, and improves functional outcomes. 13 –15 This has permitted the development of two Phase II clinical trials that have recently shown significant improvements in patients with traumatic brain injury receiving PROG. 16 –18
In our previous studies, we demonstrated beneficial effects of PROG after complete section of spinal cord: it restores the expression levels of several molecular markers, subsides chromatolysis in motoneurons, 19,20 enhances the differentiation of oligodendrocyte precursor cells by increasing the expression of pro-oligodendrogenic factors such as Olig2 and Nkx2.2, 21 and decreases the activation and proliferation of astrocytes and microglial cells. 22
In this study, we have assessed histological and functional effect of PROG with multiple approaches, including magnetic resonance imaging (MRI), stereological cell counting, open field based Basso-Bresnahan-Beattie (BBB) scale for locomotion, CatWalk gait analysis, Hargreaves plantar test, and dynamic Von Frey.
Our investigation provided compelling evidence that PROG significantly improved tissue preservation and functional outcome after spinal cord contusion. These results showed that PROG could be considered a strong candidate for the therapy of SCI, beause it has been considered for traumatic brain injury.
Methods
Animals
Young adult male Wistar rats (300–335 g, 12 weeks old) obtained from Harlan-interfauna Ibérica (Barcelona, Spain) were maintained in a 12:12 h light:dark cycle, and received food and water ad libitum. Rats were handled in accordance with the guidelines published in the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals, the principles laid out in the Guidelines for the Use of Animals in Neuroscience Research published by the Society for Neuroscience, and European Union guidelines (Council Directive 86/609/EEC). Experimental procedures were approved by the Ethical Committee for Animal Welfare at the National Paraplegics Hospital (CEBA). Special care was taken to use the minimum number of animals needed for statistical accuracy.
SCI
Male Wistar rats were submitted to a moderate-severe contusive spinal cord injury as described previously. 23,24 Briefly, animals were anesthetized with an intraperitoneal injection of sodium pentobarbital (45 mg/kg, Normon Veterinary Division, Madrid, Spain) and Xilagesic (2% xylazine, 10 mg/kg, Calier, Barcelona, Spain). Once the absence of reflexes had been checked, the rats were injected with a low dose of atropine (50 μg/kg body weight; Brown Medical, Barcelona, Spain) to reduce salivary and bronchial secretions and to avoid the induction of bradycardia and possible cardiac arrest by the surgery or xylazine. Artificial tears were applied to the eyes to prevent corneal abrasion and infection.
We performed a laminectomy of T8 vertebra, and the vertebral column was stabilized by clamping spiny processes of T7 and T9 vertebrae. Spinal cord contusion was performed with the Infinite Horizon™ device (Precision Systems and Instrumentation, Lexington, KY), applying a force of 200 Kdyn without any additional dwell time. 25 Force and displacement curves generated by the Infinite Horizon™ were checked to confirm that injuries were performed with similar intensities and profiles, showing no artifacts indicative of an erroneous/abnormal lesion, therefore ensuring consistency and reproducibility of the injury. This method enabled us to reduce the number of lesioned animals to a minimum, avoiding animal suffering without compromising the statistical accuracy. The injury produced by this method is equivalent to that described as moderate-severe by other groups. 26,27 Sham operated animals received the same protocol of laminectomy but without contusion.
Postoperative care included a subcutaneous injection of Buprex (buprenorphine, 0.05 mg/kg; Schering Plough, Madrid, Spain) and a prophylactic subcutaneous antibiotic injection 1 h after the lesion and on the following day (Baytril, Enrofloxacine, 1 mg/kg; Bayer, Kiel, Germany). The animals were fed with wet extruded rodent food, and manual bladder expression was used until they were self-voiding (within 10 days). The animals were monitored for hydration and eventual infections until the end of the experiment.
PROG treatment
Injured animals received daily subcutaneous injections of natural PROG (16 mg/kg/day, Sigma Aldrich, n=8, SCI+PROG group) or vehicle (castor oil, Sigma Aldrich, n=8, SCI group) for 60 days until sacrifice. PROG was given to awake animals 1 h after injury. This dose of PROG has been shown to prevent edema and neuronal loss and to improve cognitive responses after brain-contusion injury, 28 to induce oligodendrogenesis and remyelination after spinal cord injury, 21,29 and to decrease reactive gliosis. 22 Sham-operated rats (n=8, CTL) received a laminectomy but not SCI.
Magnetic resonance data acquisition and analysis
Rats were anesthetized and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer 60 days after injury. The spinal cord was dissected out and post-fixed for 4 h in the same solution at 4°C. Spinal cords were placed in Fluorinert (Sigma, Madrid, Spain), and MRI was performed on a 4.7-T Bruker BioSpec system (Bruker, Karlsruhe, Germany) with a H 1 surface coil. Three dimensional T2-weighted Fast Spin Echo images (T2W-3D) were acquired with the following parameters: repetition time (TR) 1650 msec, echo time (TE) 70 msec, number of averages (NA)=1, slice thickness 0.5 mm, 16 sagittal slices per sample, field of view (FOV) 2.56×1.28×0.8 cm2, and data matrix 256×128×16. T2W-3D hyperintense lesions in the spinal cord were identified and the volumes and lengths were measured with Image J software (NIH, Bethesda, MD). 23,30
Tissue processing for immunohistochemistry and Luxol Fast Blue (LFB) staining
After MRI studies, spinal cords were immersed in a solution of 30% sucrose in 0.1 M phosphate buffer during 72 h at 4°C. Tissue blocks of 5 mm including the epicenter of the spinal cord lesion in the middle of the block were then embedded in optimum cutting temperature compound (Tissue-Tek, Sakura Finetek, Zoeterwoude, NL), frozen on dry ice, cut into 50-mm thick serial coronal sections on a cryostat, and sequentially collected on slides. After immunohistochemistry and LFB staining, cross-sections were examined under a light microscope, at 40× or 60× magnification, equipped with a digital camera Panasonic GPKR2 22 connected to an Olympus BH2 microscope. Image analysis was performed using Bioscan Optimas II software or Image J, NIH image analysis software. All slides were assessed blindly with respect to treatment.
LFB staining
Sections were treated with 95% ethanol and left in LFB solution (0.1 mg% LFB in 95% ethanol with 10% acetic acid) at 60°C for 18 h. After several washes, sections were immersed in lithium carbonate and then 70% ethanol, rinsed in distilled water, dried, and mounted with Permount.
Immunohistochemistry
Immunostaining was performed as described previously 21 using primary antibodies against myelin basic protein (MBP 1:100, Abcam, Cambridge, UK), neurofilament H (NF-H 1:1000, Abcam, Cambridge, UK), or adenomatus polyposis coli (APC) clone (clone CC1 1:100, Calbiochem, Darmstadt, Germany), and secondary biotinylated anti-rabbit immunoglobulin G (IgG) (1/200 Vector Laboratories, Burlingame, CA) or biotinylated anti-mouse IgG (1/200 Vector Laboratories). After the incubation with secondary antibodies, sections were incubated with avidin-biotin complex (ABC) for 30 min (ABC kit, Vector Laboratories) and finally revealed with diaminobenzidine tetrachloride (0.50 mg/mL, Sigma, St. Louis, MO) in the presence of 0.01% H2O2 for 7 min in the dark. The sections were given a final rinse in phosphate buffered saline, dehydrated in graded ethanols and xylene, and mounted with Permount. CTL experiments were performed to rule out interference of non-specific staining. These experiments were run in parallel and involved the incubation of tissue without primary antibodies.
Quantification of tissue sparing
Serial sections were taken every 0.5 mm from 2.5 rostral up to 2.5 mm caudal to the epicenter and were assayed for LFB staining. The spared area of white and gray matter in each section was quantified using NIH image analysis software Image J. The white matter was determined to be spared if LFB staining was grossly normal in appearance and density (lacking cysts and degeneration) and spared gray matter estimation was based on comparison with normal gray matter cytoarchitecture observed in naïve animals. 25,31 We calculated the volume of spared white matter (SWM) and spared gray matter (SGM) by multiplication the spared area of each section by section thickness. The volume of spared tissue was normalized as a percentage of the volume of white and gray matter measured at the same spinal levels in intact rats.
Optical density measurement of MBP
Optical density of MBP staining was measured as described previously. 21 Briefly, inmmunostaining intensity was determined in areas (50,000 μm2) covering three white matter regions: dorsal funiculus, lateral funiculus, and ventral funiculus, anatomically delimited by the method described in Labombarda and associates. 21 Computer-assisted image analysis (Bioscan Optimas II) was used to transform differences in color intensity of immunopositive areas into gray differences, and results were expressed as the mean±standard error of the mean inverse logarithm of gray intensity per unit area (mm2) (ILIGV/area=LIGV). 20 Sections were processed simultaneously under identical light beam, wavelength, and gray-scale threshold throughout the experiment and quantification analysis. LIGV was averaged for each section and expressed as a percentage of CTL intact animals.
Stereological APC (CC1) counting
Serial sections were taken every 0.5 mm from 2.5 rostral up to 2.5 mm caudal to the epicenter and were assayed for APC immunostaining. The number of total APC+cells was estimated using the optical dissector method with total section thickness as the dissector height and a counting frame of 60 mm×50 mm. 32 Section thickness was estimated using the fluorescent properties of hematoxylin, so counterstained sections were examined under a Nikon Eclipse E 800 confocal scanning laser microscope. A total of 240 counting frames (24 per section covering dorsal, lateral, and ventral white matter funiculus) were assessed per animal. Knowing the volume of the dissector in each section, we calculated the number of APC+cells/mm3. To obtain the total number of APC+cells, we multiplied the number of cells/mm3 per the total volume of spinal cord (the summation of cross-sectional areas×thickness section×distance between slices).
Quantification of axonal profiles
We obtained micrographs of ventral, dorsal, and lateral white matter with a bright field microscope using a 60×objective. The final area of the picture was 25,000 mm2, with a final pixel size of 0.239 microns×0.179 microns. We performed a semi-automated quantification of axonal profiles using NIH image analysis software Image J. Pictures were transformed in 8-bit grayscale images, then converted to binary images using manual threshold. Dilate, Watershed and Erode filters were applied to the images, and finally the automated counting tool included in Image J was used. To obtain total axonal profile in white matter, we multiplied axonal profile density (axonal profile/mm2) per white matter area of dorsal, ventral, and lateral funiculus. Finally, the number of axonal profiles per funiculus was added, and axonal preservation was expressed as percentage of total axonal profile in white matter with respect to CTL animals.
Open-field locomotion
Open-field locomotion was evaluated by using the BBB locomotor scale according to the instructions published in the original articles. 26,27 Briefly, animals were gently placed to the open field during several sessions. Then, two trained observers (blind to hormone treatment) scored rats for 4 min at the same time of the day at 3, 7, 14, 30, and 60 days after SCI. One researcher recorded all the data on a score sheet, and the other one kept the animal moving in the open field. Movements elicited by the touch of an examiner were not scored. A score of 0 points defines no movement of the hindlimbs (HL), and the maximum of 21 points defines normal locomotion as observed in unlesioned rats. Score includes criteria such as joint movements, weight support, forelimb-hindlimb (FL-HL) coordination, and tail position.
Because interlimb coordination can be quite difficult to assess in the open field, we corrected open field BBB scoring using an objective assessments of coordination with the CatWalk system (CatWalk-based BBB score) according to Koopmans and colleagues. 33 We acquired and studied five uninterrupted runs with the CatWalk system on the day of open field testing, considering “consistent coordination” when Regularity Index (RI) (Table 1) scored 100% in 5/5 runs; “frequent coordination” when RI scored 100% in 3/5 or 4/5 runs; “occasional coordination” when RI scored 100% in 1/5 or 2/5, and “no coordination” when RI scored 100% in 0/5 runs. We also used the BBB subscoring scale to improved the sensitivity of the BBB by scoring the higher motor functions (i.e., toe clearance, predominant paw position, trunk stability, and tail position) regardless of the other BBB parameters, adding them together to yield a single score (maximum score=13). 34
RF, right forelimb; RH, right hindlimb; LF, left forelimb; LH, left hindlimb.
CatWalk-automated quantitative gait analysis
While the general locomotor performance was evaluated using the BBB scale, a more detailed analysis of locomotion was performed using the CatWalk gait analysis system in rats that were able to maintain weight support with their HL (BBB score above 9). CatWalk measures a large number of both static and dynamic gait parameters and enables quantitative assessment of locomotion. 33,35 –39
Briefly, the CatWalk system consists of a glass runway that contains light from a fluorescent tube. Internal reflection causes the light to be restricted to the glass surface plate under normal circumstances. When an object touches the glass surface, however, the light exits the glass, thereby only illuminating the contact area. Hence, placement of the rat paws on the glass floor lights up the corresponding areas on the glass floor. The run of the animal across the glass runway is detected by a video camera (Pulnix TM 765E, Pulnix Inc., U.K.) positioned underneath the glass plate. The signal is digitized (50 frames/sec) by a Picolo Diligent frame grabber board. The data are acquired, compressed, stored, and eventually analyzed by the CatWalk 7.1 software program. 35
One week before the surgery, animals were trained to walk across the CatWalk, baseline scores were recorded, and rats were then tested on days 14, 30, and 60 post-surgery (when animals regained at least frequent weight supported stepping). For locomotor analysis, the following criteria concerning walkway crossing had to be met: (1) the rat crossed the walkway without any interruption or hesitation with a consistent pace, (2) a minimum of five correct crossings with about four step cycles (i.e., each paws positioned four times) were acquired and analyzed per animal, and (3) to avoid a high variability in gait velocities, only runs with a stable crossing time of the runway between 3 and 4 sec were included.
Because no left-right differences were present in any of the gait parameters investigated, corresponding left and right gait parameters were averaged before analysis. The analyzed parameters are described in Table 1. For a full description of the locomotor parameters, see Hamers and coworkers. 36
Sensory function
Sixty days after injury, rats were brought into the behavior room 1 h before the test session to allow them to habituate to the environment. Animals were placed alone in each behavioral apparatus for 15 min before testing, and trials of a particular sensory test were separated by at least 1 hr. They received sugared cereal rewards throughout testing to keep them from attending to the movements and procedures of the examiner. The average value of three readings was used as the tolerance threshold for each hindpaw.
Mechanical sensitivity
The sensitivity to touch stimuli was assessed by using the dynamic plantar aesthesiometer (Ugo Basile, Varese, Italy), an automated apparatus based on the Von Frey filament principle. Rats were placed in a raised cage with a wire mesh floor over the stimulator unit. The metal filament was applied to the center of the palmar surface of the hindpaw, and upward force was increased from 1 to 50 g over 15 sec. Force at withdrawal was recorded for both hindpaws. 40,41
Thermal sensitivity
The Plantar test (Ugo Basile, Varese, Italy) was used to evaluate thermal hyperalgesia according to the Hargreaves method. 42 Rats were placed in a cage with a glass floor, over a movable infrared light source. The light source was positioned under the center of the palmar hindpaw, and it was turned off automatically when the rat lifted the paw, allowing the measurement of time between the beginning of the light beam and the elevation of the foot. To prevent tissue damage, a 20-sec cutoff time was set. Thermal withdrawal latency was recorded for both hindpaws. 40,41
Statistical analysis
All the statistical analysis and graphs were performed with Prism 5.0 software (
One-way analysis of variance followed by the Newman Keuls post-test or t test was used to determine statistical differences between three or two treatments, respectively, at 60 days. The number of rats was used as the n number (n=8 for all treatments). A p<0.05 was considered to be significant.
Results
SCI
The real force at the impact site and the mean amount of spinal cord displacement, two key parameters in defining the severity of injury, did not show significant differences between SCI and SCI+PROG groups, ensuring consistency and reproducibility of the injury (force: SCI 204.3±1.42 Kdyn vs. SCI+PROG 204.9±1.66 Kydn; displacement: SCI 1188±53.47 mm vs. SCI+PROG 1212±33.56 mm). After surgery, animals showed an initial drop in body weight during the first postoperative days, increasing afterward until the end of the study. There were no differences between the groups (data not shown).
PROG reduced secondary damage and increased spared tissue
We determined the effect of PROG on the extension of the lesion 60 days after injury by quantifying ex vivo the edema-related hyperintense signaling in T2W-3D MRI, which mainly reflected the cyst already formed. 43 –45 PROG reduced volume and length of T2 hyperintense signals when compared with SCI rats (Fig. 1A, B, C). In addition, the non-hyperintense region and the total volume of spinal cord at the epicenter region were significantly increased in PROG+SCI rats 60 days after the injury (Fig. 1D, E).
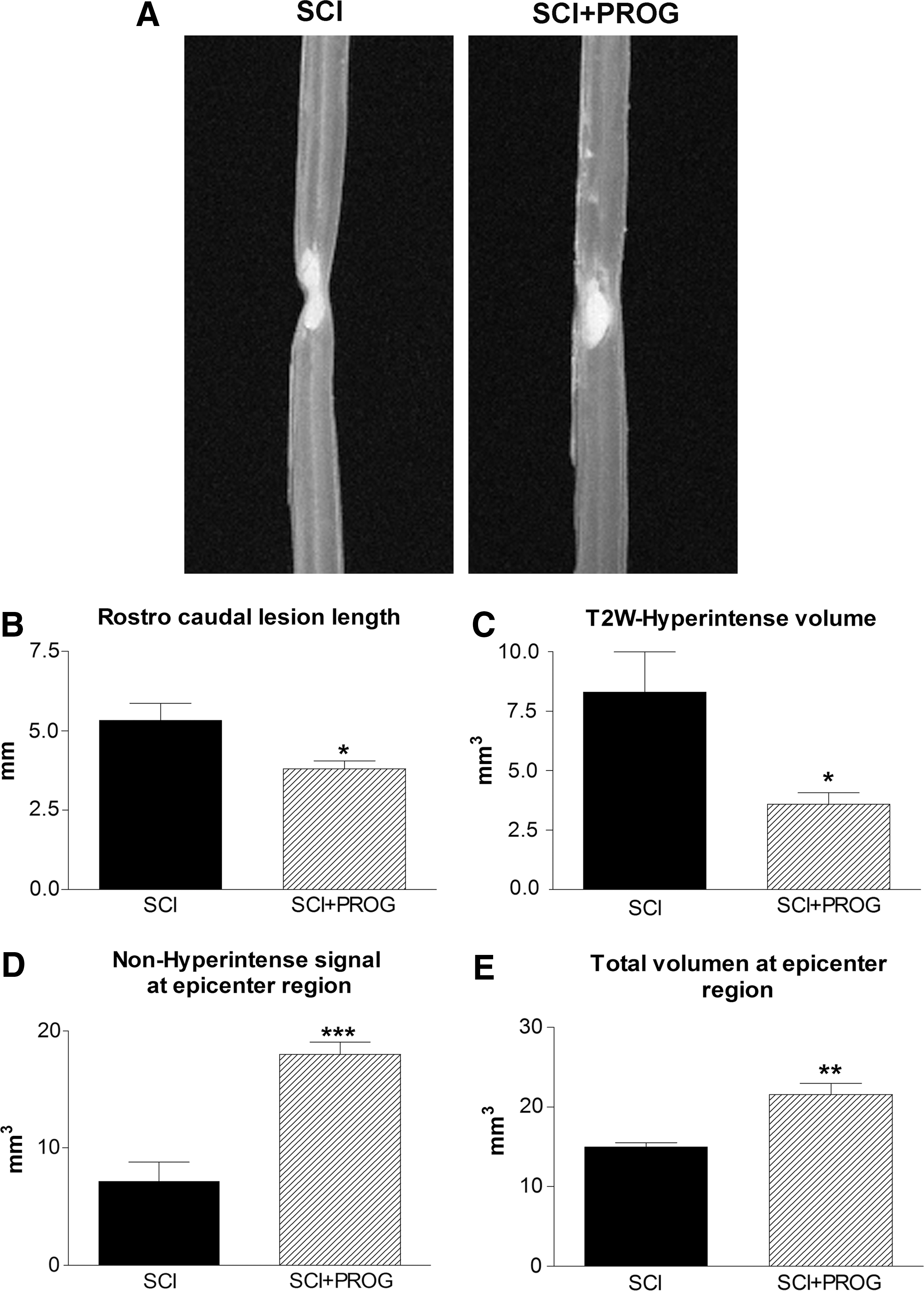
Representative T2-weighted three-dimensional magnetic resonance imaging (T2W-3D MRI) showing smaller hyperintense region in spinal cord injury plus progesterone (SCI+PROG) rats (
Next, we studied the effect of PROG administration on the extension of the lesion and tissue sparing at the histological level. We performed LFB staining on serial coronal sections taken between 2.5 mm rostrally and 2.5 mm caudally from the epicenter, and we quantified SWM and SGM (Fig. 2).

Effect of progesterone (PROG) on white and grey matter damage measured at 60 days after lesion. An increase in spred white matter (SWM) preservation was observed in spinal cord injury (SCI)+PROG rats at all distances measured from the epicenter (
After 60 days, injured animals lost approximately 77% of white matter in segments located 2.5 mm rostral and caudal to the epicenter, while PROG treatment reduced white matter decrease to about 42% (i.e., 2.5 mm rostral: SCI: 22.99±3.03%; SCI+PROG: 58.60±4.06, p<0.001, Fig .2A). The amount of white matter preservation at the epicenter itself was approximately 7% in SCI rats, while in SCI+PROG animals, it was about 16% (SCI: 7.29±1.74%; SCI+PROG: 16.19±2.17%, p<0.05, Fig. 2A). The preservation of SWM was observed between SCI and SCI+PROG groups at all distances measured from the epicenter (Fig. 2A).
On the other hand, no statistical differences were observed between the SCI and SCI+PROG rats in SGM (SCI vs. SCI+PROG, p>0.05, Fig. 2B).
Additional quantitative assessment of the area of white matter at lateral and ventral funiculus was performed at the epicenter itself, because this parameter is related to supraspinal control. 46,47 This area was significantly larger in contusioned animals treated with PROG than in the vehicle group (Fig. 2C, SCI+PROG: 0.38 mm2±0.034 vs. SCI: 0.27 mm2±0.067, p<0.05).
PROG administration increased oligodendrocyte numbers and reduced myelin damage after SCI
We studied the effect of PROG administration on oligodendrocyte preservation. We stereologically quantified the number of CC1+cells in SWM within a 5 mm segment of the lesion epicenter.
After 60 days of spinal cord contusion, lesioned animals showed a 93.46% reduction in the total number of CC1+cells throughout the white matter, while PROG-treated rats showed a higher number of cells, losing about 65% of oligodendrocytes compared with sham-operated rats (CTL: 425 103±13.18 103, SCI: 27.83 103±4.30 103, SCI+PROG: 127.4 103±13.18 103; p<0.001 SCI vs. CTL; p<0.001 SCI vs. SCI+PROG; Fig. 3A, B).
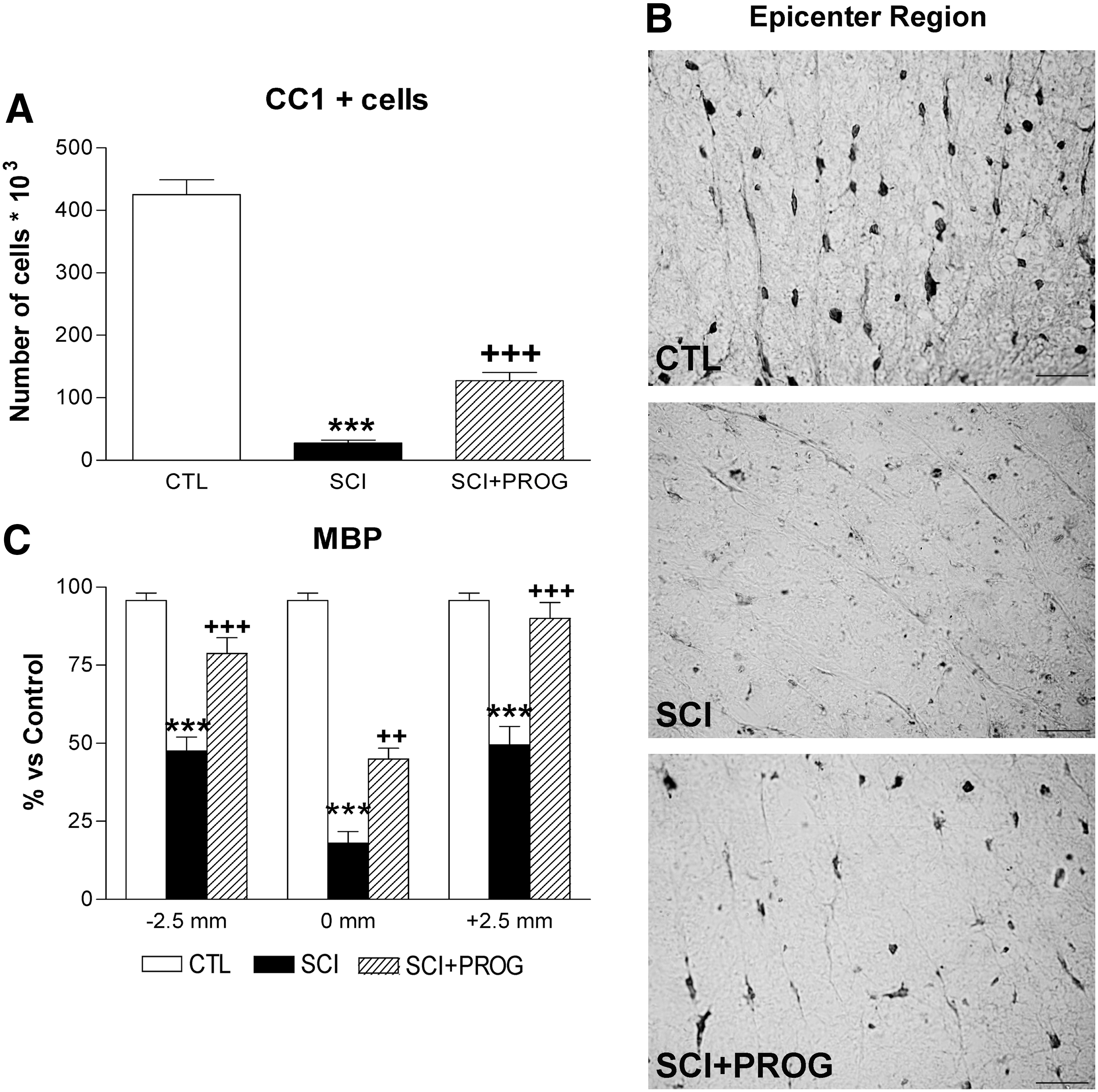
Progesterone (PROG) increased the oligodendrocytes number and reduced myelin damage 60 days after lesion. Spinal cord injury (SCI) animals showed a reduction in the total number of CC1+cells throughout the white matter compared with control (CTL) rats, while the in SCI+PROG group, the reduction was lower (
Next, we measured MBP immunoreactivity in the SWM of 2.5 mm rostral, 2.5 mm caudal, and the midpoint of the epicenter region (Fig. 3C). MBP immunoreactivity diminished dramatically in SCI rats at all studied regions but was partially preserved in SCI animals treated with PROG (SCI vs. SCI+PROG, p<0.01 at all regions, Fig. 3C). Therefore, increased oligodendrocyte density in the PROG-treated group was accompanied by better myelin preservation.
PROG improved axonal preservation after spinal cord contusion
We also evaluated the number of axonal profiles on sections immunostained against NF-H. Quantification revealed a higher axonal preservation in PROG-treated animals compared with vehicle-treated ones (SCI+PROG vs. SCI, p<0.05 or less at all regions, Fig. 4A, B). This axonal protection was verified along the whole 5-mm segment.

Progesterone (PROG) improved axonal preservation 60 days after injury at 2.5 mm rostral, 2.5 mm caudal, and at the midpoint−0mm- of the epicenter (
PROG improved locomotor outcome after SCI
Open-field locomotion
The general locomotor performance was evaluated using the BBB scale, but interlimb coordination was corrected with the RI value obtained in the CatWalk gait analysis (CW-BBB) (see Methods, Table 1).
All rats manifested significant bilateral HL paralysis immediately after injury. After 14 days, SCI rats improved their scores, showing consistent weight-supported HL steps but no FL-HL coordination (CW-BBB score=11), while PROG-treated rats demonstrated consistent coordinated FL-HL movements (CW-BBB score=14) (p<0.05 SCI vs. SCI+PROG, Fig. 5A). This pattern was maintained for the duration of the assessment period. Behavioral improvements seen in PROG-treated animals were present at each time point after 14 days (p<0.05 at all time points, Fig 5A).

Integration of the CatWalk(CW)-based coordination into the Basso-Bresnahan-Beattie (BBB) locomotor scale results in the CatWalk-based BBB, which showed that spinal cord injury plus progesterone (SCI+PROG) animals regained better locomotor function when compared with SCI rats (
As indicated in Methods, the BBB subscore evaluates aspects of fine motor control regardless of the other BBB parameters. 34,48 After 14 days, the BBB subscore of SCI animals became stable at a score of 2, whereas PROG-treated animals eventually reached a higher score between 5 and 8 (Fig. 5B). At 60 days post-injury, PROG rats presented better subscores than the SCI group: 75% of PROG rats had the tail consistently up, 50% had consistent trunk stability and parallel paw position of HL during the stance, and 62% showed consistent toe clearance. In the SCI group, only 37% of rats had the tail up, none of the rats reached trunk stability and parallel paw position, and 12% showed consistent toe clearance.
CatWalk-automated quantitative gait analysis
Fourteen days post-injury, all rats in both groups reached a BBB score higher than 9, allowing us to test their locomotor performance also by CatWalk gait analysis. Importantly, gait velocity, which strongly affects the variations in gait parameters, is controlled using the Cat-Walk 36 by training the animals to make runs with a stable crossing time. During this study, no significant differences were found between animal groups with respect to crossing time or average body weight, another parameter that affects gait (data not shown). There were no significant differences in any studied parameter for the functions of the FL between SCI and SCI+PROG rats.
Base of support (BOS) of the hindpaws
The distance between the hindpaws during locomotion is an indicator of trunk stability of the animal. BOS was higher in SCI rats throughout the test period compared with sham control animals (p<0.001 at all time points, SCI vs. CTL, Fig. 6A). PROG-treated rats showed a decrease in BOS with respect to SCI rats (p<0.05 or less at all time points, SCI vs. SCI+PROG, Fig. 6A). During the study, BOS increased slightly over time in all groups, probably because of the normal increases in body weight (CTL, SCI, SCI+PROG; Time: F (2, 42)=5.24, p<0.05, Fig. 6A).
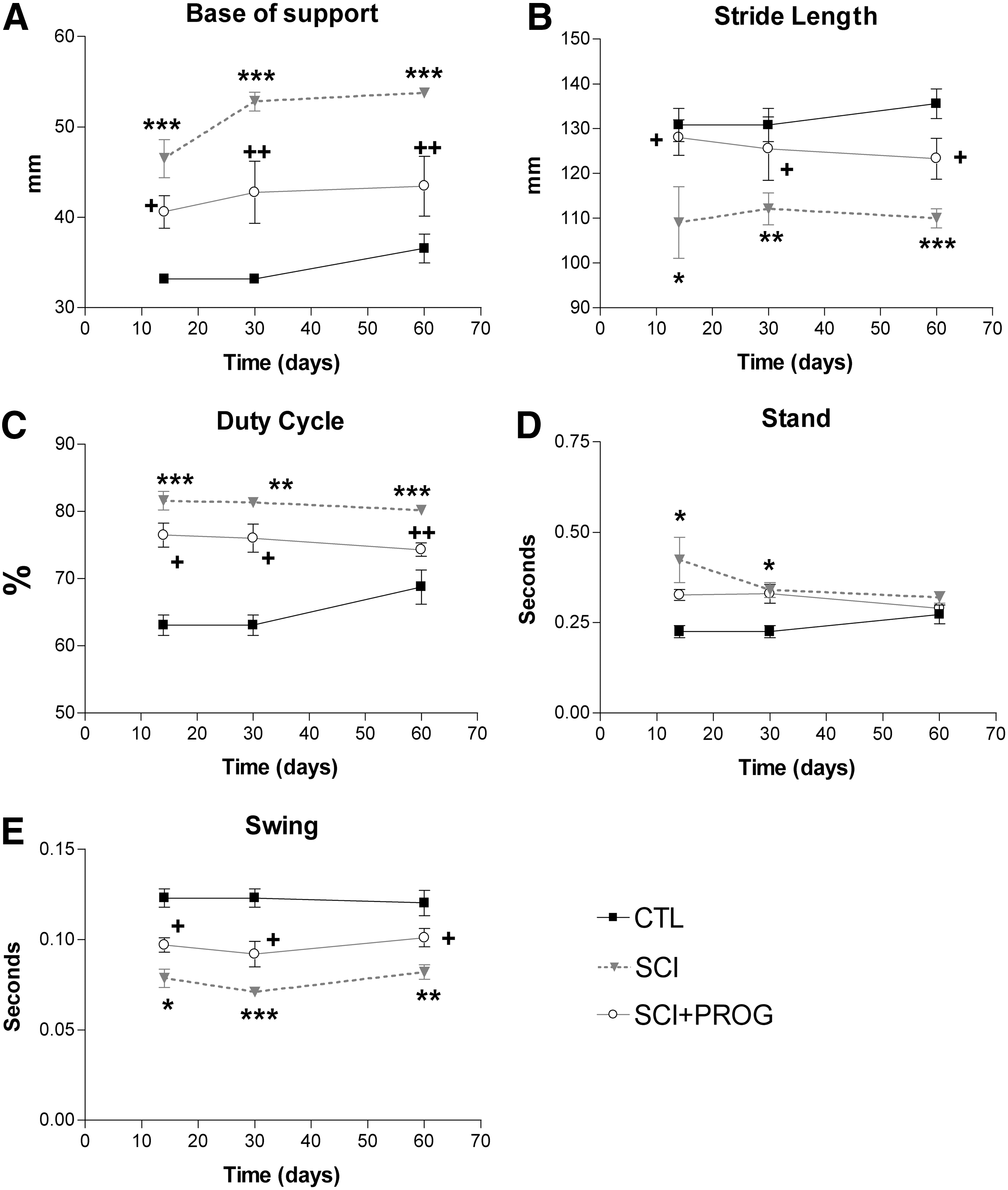
CatWalk gait parameters analysis showed that progesterone (PROG) improved overground locomotion. Base of support (
Stride length
Stride length decreased significantly in injured rats compared with sham-operated animals (p<0.05 or less at all time points, SCI vs. CTL, Fig. 6B). PROG-treated rats, however, showed an increased stride length compared with SCI rats (p<0.05, SCI vs. SCI+PROG, Fig. 6B) and remained closer to sham-operated values (ns, SCI+PROG vs. CTL, Fig. 6B)
Duty cycle, Stance, and Swing
The Duty cycle is the percent of the total step cycle during which a paw is on the ground (Table 1). The Duty cycle of the hindpaws increased in injured rats (p<0.05 or less at all points, SCI and SCI+PROG vs. CTL, Fig. 6C), but this increase was smaller in rats treated with PROG (p<0.05 or less at all time points, SCI vs. SCI+PROG, Fig. 6C).
The time that the hindpaw was not in contact with the glass floor (Swing phase) decreased in all injured animals, but to a lesser extent in PROG-treated animals throughout the study (p<0.05 at all time points SCI vs. SCI+PROG, Fig. 6E). The time that the hindpaws were in contact with the floor (Stance phase), however, was significantly increased in both groups of lesioned animals only after 14 and 30 days (p<0.05 SCI and SCI+PROG vs. CTL, Fig. 6D).
Print area, Max area, and Paw angle
Even though the duration of the Stance phase did not change among groups, we evaluated additional characteristics of the Stance by studying Print area, Max area, and Paw angle (see Table 1). After 14 and 30 days, both injured groups showed statistically significant increases in Print area and Max area compared with sham, but without significant differences between them (data not shown). After 60 days, however, the Max Area and Print area values remained similar in SCI rats while in SCI+PROG animals, both parameters decreased, reaching values similar to those of sham-operated rats (Table 2). Paw angle increased throughout the study in injured animals, but to a lesser extent in those treated with PROG (data not shown). Data at 60 days are shown in Table 2.
CTL, control; SCI, spinal cord injury; PROG, progesterone.
p<0.05 vs. CTL; *** p<0.001 vs. CTL; # p<0.05 vs. SCI. One-way analysis of and Student Newman Keuls post-test.
RI
This parameter reflects FL-HL coordination, correlating the order of paw placements of the animal with the normal step sequence patterns. 35,36 In sham-operated animals, all runs reached a RI of 100%, showing perfect interlimb coordination. In injured animals, mean RI was reduced in both groups from 2 weeks after injury (p<0.05 vs. CTL, Fig. 7), but the decrease was smaller in PROG-treated animals at all time points (p<0.05 SCI vs. SCI+PROG, Fig. 7). These PROG-treated animals recovered interlimb coordination reaching RI values similar to control rats at 30 days (95.72±1.64, ns SCI+PROG vs. CTL) and 60 days (96.76±1.19, ns SCI+PROG vs. CTL, Fig. 7).
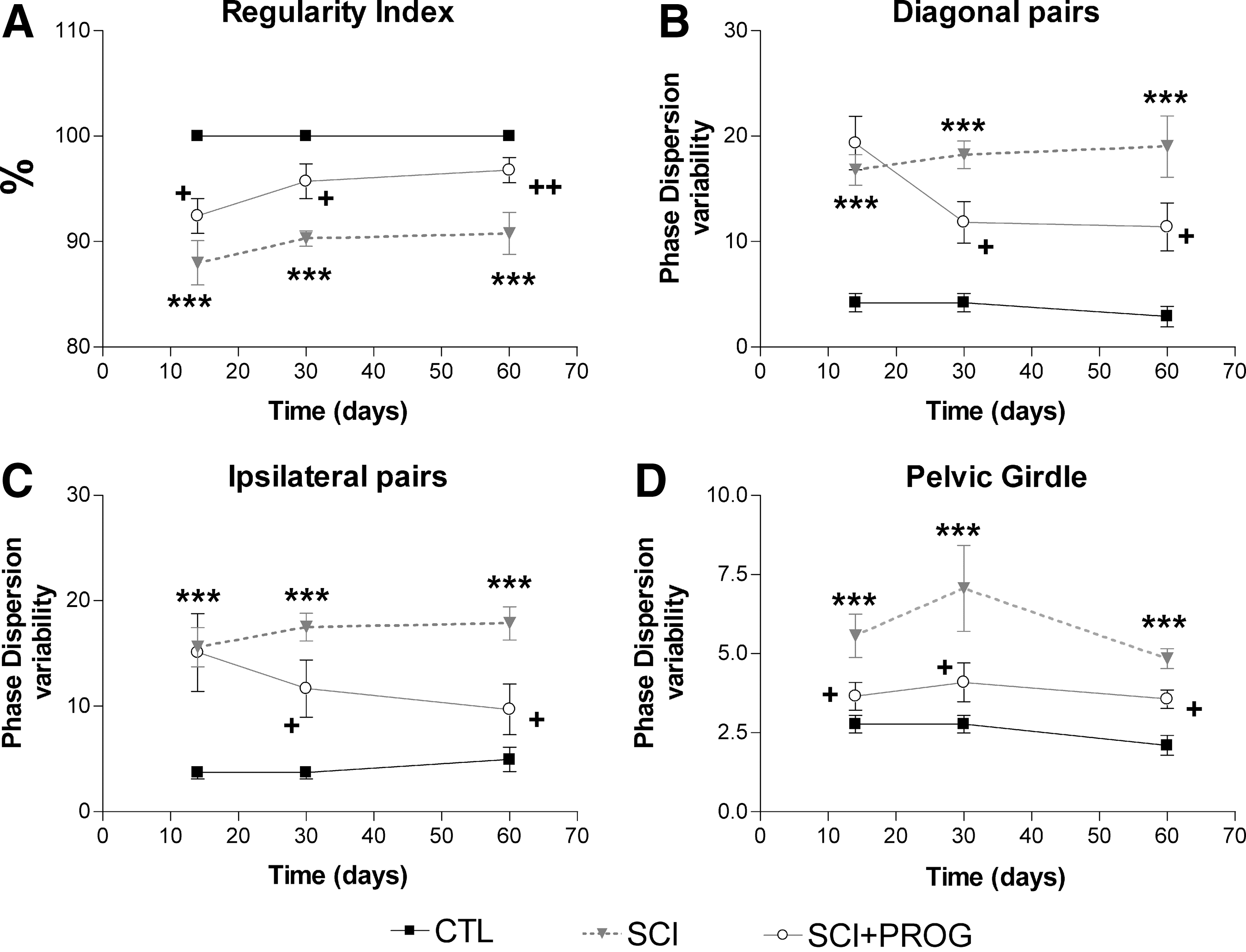
PROG treatment improved interlimb coordination after injury. Spinal cord contusion decreased RI in SCI rats compared to CTL animals, but in a lesser extent in SCI+PROG
Phase dispersions (PD)
Another parameter used to estimate coordination is the measurement of PD, which, in contrast to the RI, determine the paw placements with respect to timing (Table 1). Changes in PD variability represent a measure of interlimb coordination: during uninterrupted locomotion, coordinated limb pairs show very small phase dispersion variability, whereas limbs that move largely autonomously display a high variability. 33,49
We averaged the values of complementary pairs to perform statistical analysis: PD diagonal pairs is the average between left forward-right hindlimb (LF-RH) and right forward-left hindlimb (RF-LH) and PD ipsilateral pairs is the average between right forward-right hindlimb (RF-RH) and left forward-left hindlimb (LF-LH). We also estimated the PD variability of pectoral (LF-RF) and pelvic (LH-RH) girdles.
Injury severely increased PD variability in all pairs of paws compared with sham animals (ipsilateral pairs: F2,42=42.26, p<0.001, Fig. 7B; diagonal pairs: F2,42=38.27, p<0.001, Fig. 7C; pelvic girdle: F2,42=31.37, p<0.001, Fig. 7D). Significant effects of PROG treatment became apparent after 14 days, however. The PD variability was significantly lower in SCI+PROG than in SCI rats in the ipsilateral and diagonal pairs and in the pelvic girdle (p<0.05 SCI vs. SCI+PROG Fig. 7B, C, D). The value of PD variability was stable inside each group over time, not existing statistically significant differences from 14 onward (ipsilateral pairs: F2,42=2.43, ns, Fig. 7B; diagonal pairs: F2,42=1.02, ns, Fig. 7C, pelvic girdle: F2,42=3.50, ns, Fig. 7D).
On the other hand, changes in PD mean value represent changes in the step patterns during gait. 49 Lesioned animals showed changes in the mean PD for ipsilateral and diagonal pairs vs sham-operated controls, but no difference was observed between SCI and SCI+PROG rats (data not shown)
Sensory function
Animals were tested for mechanical and thermal sensitivity to explore sensory functions after 60 days of spinal cord contusion.
Mechanical sensitivity
There was no significant difference in the filament force necessary to elicit a hindpaw withdrawal between the SCI rats and sham-operated animals, indicating no significant mechanical allodynia in our model of contusion (Fig. 8A). In addition, chronic PROG administration did not change the threshold of response to mechanical stimulation (Fig. 8A). This was supported also by the absence of differences in some CatWalk parameters such as the duration of Stance phase (Fig. 6D) and the pressure applied during Stance (Print intensity) and by the increase in the area contacted by the paw during the stand (Print area) and the increase of the area contacted at the moment of maximum paw-floor contact (Max area) (Table 2). Decreases in those parameters have been previously related to mechanical allodynia. 50,51

Neither PROG nor spinal cord injury induced mechanical or thermal allodynia after 60 days of spinal cord contusion. Dynamic Von Frey
Thermal sensitivity
As for mechanical stimulation, no statistically significant differences were observed between groups, showing that our model did not produce thermal hyperalgesia and that PROG administration did not induce changes in the nociceptive behavior (Fig. 8B).
Discussion
The current study demonstrated that PROG treatment after spinal cord contusion was beneficial at the anatomical, histological, and functional levels. Because at present there are no universally accepted treatments for this neurological disorder, our present investigation identifies PROG as a promising candidate for clinical trials.
PROG reduced secondary injury and white matter pathology
By MRI techniques, we observed that the volume of hyperintense signal was smaller in rats treated with PROG, suggesting that the steroid effectively reduced edema formation after spinal trauma. This is in agreement with previous reports showing that PROG reduces edema formation after traumatic brain injury 16 by decreasing the expression of aquaporin-4 at the zone of injury 52 and by stabilizing the blood–brain barrier with its antioxidant effects. 53 The improvement in tissue preservation and total spinal cord volume may imply a secondary damage reduction, an effect expected given the already described effects of PROG in decreasing inflammation and reactive gliosis 13,22,54,55 reducing oxidative stress 53 and decreasing excitotoxicity via indirect conversion into allopregnenalone. 9,56 –58
Further, PROG notably reduced the extension of the hyperintense signal in a rostrocaudal direction. This would imply that PROG may be beneficial for injuries located near the lumbar or cervical enlargements, where limiting the expansion of the primary damage would preserve levels containing, for example, central pattern generators for locomotion or breathing control.
Our histological measurements support and extend our MRI findings: SWM values were higher at all distances measured in PROG-treated rats in accordance with the MRI data. A remarkable agreement across studies indicates that only 5–10% tissue sparing is necessary for the recovery of plantar stepping (BBB score=10–11). 27,59 –61 Accordingly, SWM was about 7% at the epicenter of lesioned rats that showed a consistent stepping (mean BBB score=11) 60 days after the lesion. Interestingly, PROG treatment preserved 16% of SWM at the epicenter, and rats demonstrated consistent plantar stepping with FL-HL coordination (mean BBB score=14). This confirms that a small increase in SWM may result in a substantial recovery of locomotor function, probably by preserving more supraspinal and propiospinal inputs. 27,49,61
In this regard, after PROG treatment, the areas of dorsolateral and ventrolateral funiculus were significantly larger at the epicenter site. White matter tracts that occupy the outer rim of the cord such as the rubro-, raphe-, reticulo- and vestibulospinal tracts are part of the descending brainstem system that appear to provide important, if not essential, input to the central pattern generator (CPG) to initiate locomotion and produce complex and precise locomotor patterns. 46,47,62 Thus, axon sparing of these tracts by PROG treatment may underline the described locomotor recovery.
PROG and its reduced metabolite, allopregnenalone, are well recognized promyelinating factors in the peripheral and central nervous systems in several diseases. 9 –12,63 –67 We have demonstrated in previous works that chronic PROG administration increases the expression of transcription factors that define oligodendrocyte linage, such as Olig2 and Nkx2.2 after spinal cord transection. 21,29 In the present work, we report that PROG treatment reduced the loss of oligodendrocyte numbers and MBP immunoreactivity produced by SCI. This effect could be because of preservation of the myelin-producing cells, but also to an increase in oligodendrocyte numbers by differentiation of oligodendrocyte precursors into mature myelinating oligodendrocytes, promoting the remyelination process. Either way, the effects on oligodendrocyte number could contribute to the reported PROG actions in locomotor outcome, because therapeutical strategies that limit white matter damage and oligodendrocyte death have been shown to improve recovery after SCI, 68,69 and spontaneous functional recovery correlates well with mature oligodendrocyte number in white matter after spinal cord contusion. 70 PROG could have acted directly on oligodendrocytes, or indirectly by modulating the inflammatory environment, because PROG treatment decreases gliosis and cytokine production after spinal cord transection 22 (author unpublished results). Further studies should be performed to answer this question.
We completed the evaluation of SWM with the study of axonal profile preservation at the epicenter. Quantification revealed a higher axonal preservation in PROG-treated animals along the whole 5-mm segment compared with SCI rats. PROG could protect axons by reducing inflammation, oxidative stress, reactive gliosis, edema, and excitotoxicity, but also preserving the integrity of the myelin sheath. 71 –73 Otherwise, PROG could have promoted axonal regeneration of supraspinal or propiospinal neurons.
Finally, PROG produced no significant changes in SGM. This is in accordance with previous reports, which described that functional deficits after midthoracic injuries are mainly attributed to white matter damage. 74 –76
PROG improved locomotor outcome after spinal cord injury
Behavioral recovery is the highest goal in the search for therapeutically relevant repair strategies for SCI. An important aspect of the observed locomotor outcome is the recovery of coordination. As it was demonstrated by others, 27,35,49,77 rats after thoracic spinal cord trauma exhibited no interlimb coordination (CW-BBB score=11, high PD variability of ipsilateral and diagonal pairs of HL). The group treated with PROG, however, regained better locomotor function presenting consistent FL-HL coordination (CW-BBB score≥14, and values close to control animals in coupling between FL-HL, RI, and PD variability of the ipsilateral and diagonal pairs). It is generally accepted that coordinated gait can only be generated when there is a functional connection between the CPGs of the individual limbs located in the lumbar and cervical spinal cord. 78 –81
The recovery of FL-HL coordination in the PROG-treated rats indicated that communication between FL and HL CPGs has improved. Possible mechanisms underlying the recovery may be: (1) the preservation of descending brainstem systems, which are important in activation of CPG (rubro-, vestibulo-, reticuloraphe- and propiospinal tracts) 46,47,62,78 ; (2) the formation of new intraspinal circuits that might contact long propiospinal neurons bridging the lesion and arborizing on lumbar neurons 46,82 ; (3) neuroprotection and CPG plasticity. In this regard, we have published that PROG increases the synthesis of BDNF after spinal cord transection, 19 which, in turn, like IGF-1, may induce synaptogenesis and CPG plasticity. 33,83 Additional tract-tracing experiments to verify theses hypothesis would be needed, however.
The greater trunk stability in PROG-treated rats could produce the improvement of the following gait parameters: BOS, Swing phase, Print- and Max area, and Stride length. Particularly, an increased BOS is a common feature of SCI 36,84,85 and has been explained as an adaptation for an instable gait. 35 Thus, recovery of a normal hindlimb BOS, 60 days post-injury in PROG-treated rats, could be related to improvements in trunk stability. These data agrees with the BBB subscore, which indicated that in SCI+PROG group, 50% of the rats showed trunk stability, while none of the rats showed trunk stability in the SCI group. Alternatively, pronounced external hindpaw rotation might increase BOS measurements. 36 In this regard, PROG also decreased the hindpaw rotation after injury, possibly reducing also the BOS. Indeed, the BBB-subscore showed that 50% of the animals in the SCI+PROG group reached a normal parallel paw position, while none of the rats reached the appropriate position in the SCI group.
In accordance with previous reports, Duty cycle increased after SCI,
35,84
because of the reduction of the Swing phase. Conversely, after PROG treatment Duty cycle almost reached control values, because Swing phase was increased. That means that the rat paw falls to the ground faster in vehicle-treated rats than in PROG-treated rats. Even though the duration of the Stance phase did not change among groups, we evaluated additional characteristics of the Stance by studying Print and Max areas. Both parameters increased after SCI as
Stride length normally decreases after injury 36,38,86 ; however, PROG treatment raised this parameter, and rats tended to recover the ability to step a normal distance. These results might be the consequence of an improvement in BOS, the increase of Swing phase, and greater trunk stability.
Treatments that produce beneficial effects after SCI also may present undesired effects. In our current study, we checked whether PROG effects on locomotion were accompanied by the development of mechanical allodynia or thermal hyperalgesia. In the spinal contusion model, it has been described that normal sensory function is supported by as little as 10% of white matter sparing. 49 Accordingly, we did not find mechanical allodynia or thermal hyperalgesia in rats with SCI, and we assessed that PROG did not induce such behaviors either, neither measured by classical tests, such as hot and cold plate and the Hargreaves method, nor with CatWalk parameters normally decreased in animals in which allodynia is developing (Stance, Print Intensity, Print and Max area). 50,51
Data obtained in the present study demonstrate that PROG improved locomotor outcome after thoracic contusion. There is no general consensus about the functional effects of PROG after SCI, however. While improvement of motor and histological outcome has been reported in one study, 87 no recovery is observed in another. 88 The discrepancies between our present investigation and the report by Fee and colleagues 88 may be mainly because of the duration of PROG treatment. They performed an acute treatment of 5 days, while we treated the rats with PROG during all the experiment (60 days), suggesting that a long-term treatment is needed to observe the behavior outcome.
Conclusion
The current study adds further evidence to support PROG as a potential effective treatment after SCI, as it has been used for traumatic brain injury.
We suggest that beneficial actions of PROG on locomotor outcome could be related to the reduction of secondary damage and the preservation or regeneration of axons and myelin of the descending pathways, and with the increase of oligodendrocytes number, either by inhibiting their apoptosis or promoting their differentiation. Most of the clinical trials for SCI are directed to limiting secondary injury to prevent neurological function loss and to provide the anatomical substrate for further reparation. 89 In this line of evidence, PROG treatment holds promise for success. In fact, multi-potential drugs such as PROG, which have pleiotrophic sites of action and modulate multiple injury mechanisms, could be the focus of future clinical trials in spinal cord trauma.
Footnotes
Acknowledgments
We thank Dr. Angel Arevalo-Martin for his generous assistance with MRI interpretation and his contribution to the development of some quantification procedures. We also are grateful to Dr. Jorge Collazos-Castro for his selfless help and generosity in sharing his knowledge. We thank Javier Mazarío for his generous assistance with thermical and mechanical sensitivity tests. We thank Marién Fernandez, David Castejón, and Palmira Villa (CAI de Resonancia Magnética, Instituto Pluridisciplinar, Universidad Complutense de Madrid) for technical assistance with the MRI. This work was supported by the University of Buenos Aires (UBACyT 2011–2014: 20020100200053 - / UBACyT 2011–2014: 20020100100089). B. Paniagua was partially funded by Wings for Life Foundation.
Author Disclosure Statement
No competing financial interests exist.