
Abstract
Chronic subdural hematoma (CSDH) is considered to be an inflammatory disease. Eosinophils are frequently expressed in the outer membrane of CSDH and are major sources of transforming growth factor beta (TGF-β). The mothers against decapentaplegic (Smad)-signaling pathway, which is activated by TGF-β, has been shown to be involved with fibrosis. In the present study, we compared the concentrations of eotaxin-3, eosinophil-specific chemoattractant, and TGF-β between CSDH fluid and cerebrospinal fluid (CSF) from control patients. We also explored the expression of the Smad-signaling pathway in the outer membrane of CSDH. Eight patients whose outer membrane and 12 whose CSDH fluid were successfully obtained during trepanation surgery were included in the study. Concentrations of eotaxin-3 and TGF-β were measured by enzyme immunoassay kits. Expression levels of Smad2, phosphorylated Smad3, Smad3, Smad4, and actin were examined by Western blot analysis. In addition, expression of Smad3 was also examined by immunohistochemistry. Concentrations of eotaxin-3 and TGF-β in CSDH fluid were significantly higher than those in CSF. Smad2, Smad3, phosphorylated Smad3, and Smad4 were detected in all cases. Smad3 was shown to be present in fibroblasts. These findings indicate that eotaxin-3 is expressed in CSDH fluid, inducing eosinophils into the outer membrane and resulting in elevation of TGF-β with the Smad pathway activated by TGF-β. These data suggest a potential mechanism for CSDH formation and growth.
Introduction
C
Previous studies have shown that eosinophils infiltrate the outer membrane of a CSDH, 8 –10 suggesting that eosinophils might play a role in CSDH growth. Eosinophils can secrete plasminogen and induce fibrinolysis, thus propagating the pathophysiology of CSDH. 9,10 Eosinophils play an important role in processes of repair and healing 8 and are considered to be major effector cells of tissue fibrosis in a variety of eosinophil-associated allergic diseases, including asthma 11 –13 and idiopathic pulmonary fibrosis. 14 However, the role of eosinophils in CSDH outer membranes is not fully understood. The present study was therefore undertaken to assess whether the proteins related to eosinophils were present in CSDH fluid or CSDH outer membranes through implementation of immunoblot and immunohistochemical (IHC) analysis.
Methods
Patients
A total of 12 patients (8 men and 4 women; age, 58–79 years; mean age, 69) suffering from CSDHs, as confirmed by computed tomography or magnetic resonance imaging, were enrolled in this study. All patients had a history of head injury, and none had any hemostatic disorder or were receiving antiplatelet or -coagulation therapy. All patients underwent burr hole irrigation surgery under local anesthesia at the Aichi Medical University Hospital (Aichi, Japan). The ethics committee of Aichi Medical University approved this clinical experiment. Informed consent was obtained from each patient.
Analysis of eotaxin-3 and transforming growth factor beta 1
CSDH fluids were sampled during trepanation surgery in all cases. As a control, cerebrospinal fluid (CSF) samples were obtained from 6 patients undergoing neck clipping for unruptured cerebral aneurysm. After collection, all samples were immediately centrifuged and supernatant fluids were stored at −80°C until analysis. Eotaxin-3 and TGF-β1 were measured using an enzyme immunoassay (R&D Systems, Minneapolis, MN), according to the manufacturer's instructions. The limits of detection of these assays were 2.3 pg/mL for eotaxin-3 and 4.6 pg/mL for TGF-β1.
Western blot analysis
Samples of the outer membranes from CSDHs were also obtained from 8 patients during trepanation surgery. Outer membranes were homogenized using a homogenizer in 80 μL of homogenization buffer containing 50 mmol/L of Tris base/HCl (pH 7.5), 0.1 mmol/L of dithiothreitol, 0.2 mmol/L of ethylenediaminetetraacetate, 0.2 mmol/L of ethyleneglycol-bis-aminoethylether tetraacetate, 0.2 mmol/L of phenylmethylsulfonyl fluoride, 1.25 μg/mL of pepstatin A, 0.2 μg/mL of aprotinin, 1 mmol/L of sodium orthovanadate, 50 mmol/L of sodium fluoride, 2 mmol/L of sodium pyrophosphate, and 1% Nonidet P-40. Homogenates were later centrifuged at 12,000g at 4°C for 10 min. Protein concentrations of supernatants were determined by Bradford's assay, using bovine serum albumin as the standard. Crude samples (25 μg of protein each) were subjected to 7.5% sodium dodecyl sulfate polyacrylamide gel electrophoresis, and proteins were subsequently transferred to polyvinylidene difluoride membranes. Membranes were incubated overnight at 4°C with primary polyclonal antibodies (Abs) against actin (Sigma-Aldrich, St. Louis, MO), Smad3 phosphorylated at Ser423/425 (Cell Signaling Technology, Danvers, MA), Smad2 (Cell Signaling Technology), and Smad 4 (Cell Signaling Technology) at a dilution of 1:500. After washing, membranes were incubated with secondary Abs conjugated to horseradish peroxidase (Cell Signaling Technology) at a dilution of 1:3000 for 30 min at room temperature. Reactions were developed with enhanced chemiluminescence (ECL) or ECL plus (GE Healthcare, Buckinghamshire, UK). Ser423/425 phosphorylated Smad3 immunoblots were stripped and reblotted overnight at 4°C with primary polyclonal Smad3 (Cell Signaling Technology) at a dilution of 1:500. Reblotted membranes were developed with ECL plus.
Histological examination
To study the cellular localization of Smad3, IHC staining, using the avidin-biotinylated peroxidase complex (ABC) technique, was performed at room temperature (n=3). Outer membranes were fixed in 10 mL of ice-cold 4% paraformaldehyde in 0.1 mol/L of phosphate buffer (pH 7.4) for 3 h. Serial axial cryostat sections (10 μm) were collected on slides for staining. After hematoxylin and eosin (H&E) staining (n=3), nonspecific immunoreactivity was blocked with goat serum for 30 min. Samples were incubated overnight at 4°C with primary Ab against Smad3 (Cell Signaling Technology) at a 1:50 dilution. After washing, biotinylated anti-rabbit immunoglobulin G was applied for 1 h, followed by a 1-h incubation with ABC. Sera for the blocking step, biotinylated Abs, and ABC were purchased from Vector Laboratories (Burlingame, CA). Reaction products were developed by incubation in 0.05% 3,3′-diaminobenzidine tetrachloride and 0.01% H2O2 in 50 mM of Tris-HCl (pH 7.5) for 10 min. Chronic subdural membranes without primary Ab were used as negative controls.
Statistical analysis
Data are expressed as the mean±standard error values. Significant differences between groups were assessed by Mann-Whitney's U test, and significance was accepted when p<0.05.
Results
Concentrations of eotaxin-3 and transforming growth factor beta 1
In control CSF samples, the mean value of eotaxin-3 was 9±2 pg/mL. The mean concentration of eotaxin-3 in CSDH fluids from 12 patients was 3,245±1,047 pg/mL, which was significantly higher than control CSF (Fig. 1A). Concentration of TGF-β1 in CSDH fluids was 23,781±2,969 pg/mL, which was also significantly higher, compared to control CSF (71±12 pg/mL; Fig. 1B).
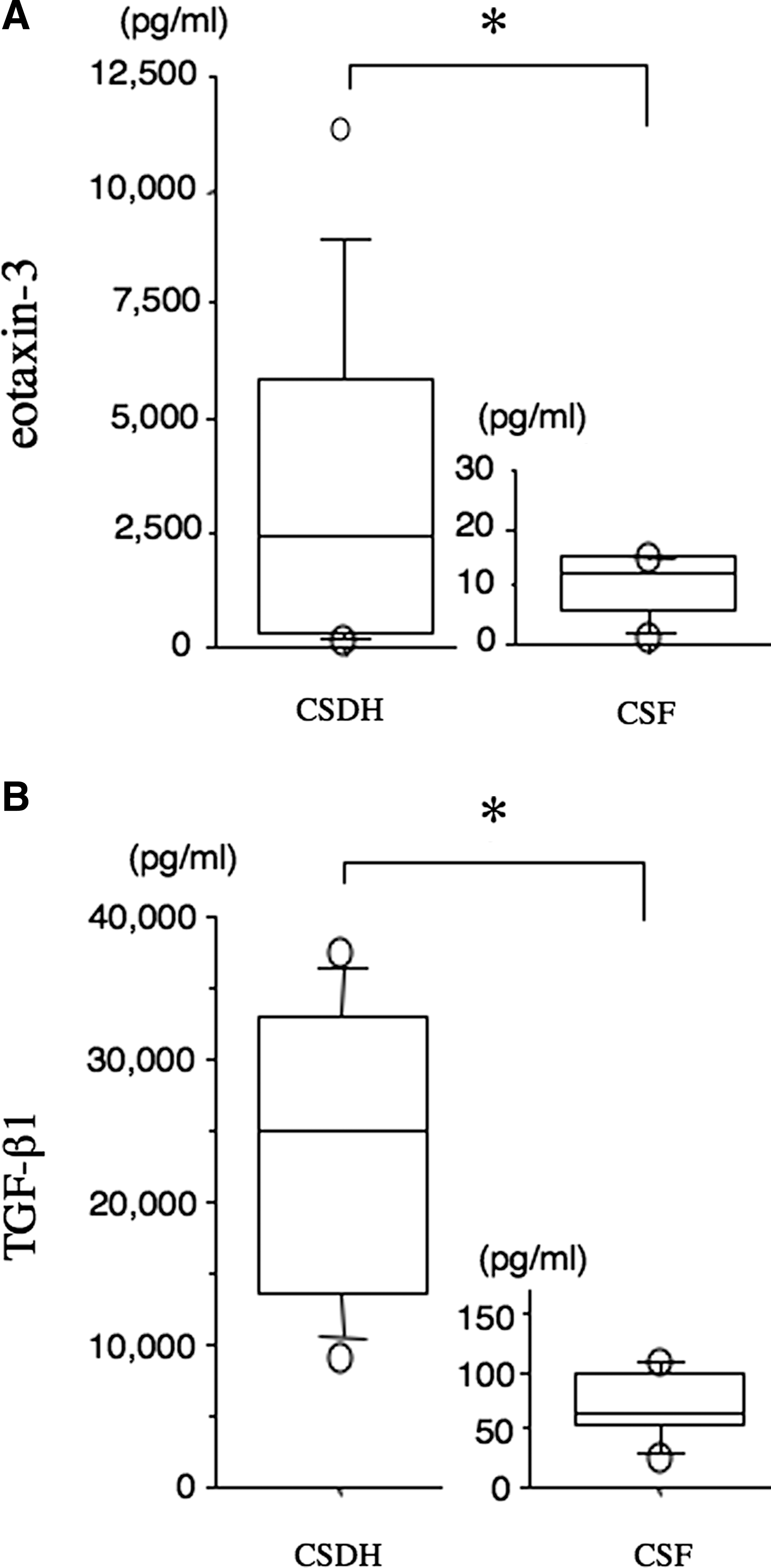
Concentrations of eotaxin-3 and transforming growth factor-β1 (TGF-β1;
Western blot analysis of mothers against decapentaplegics
Nearly consistent amounts of actin were detected on all blots, showing that equal levels of protein were loaded on gels. Smad2, Smad3, and Smad4 were detected in all cases, and Smad3 was phosphorylated in all cases (Fig. 2A).
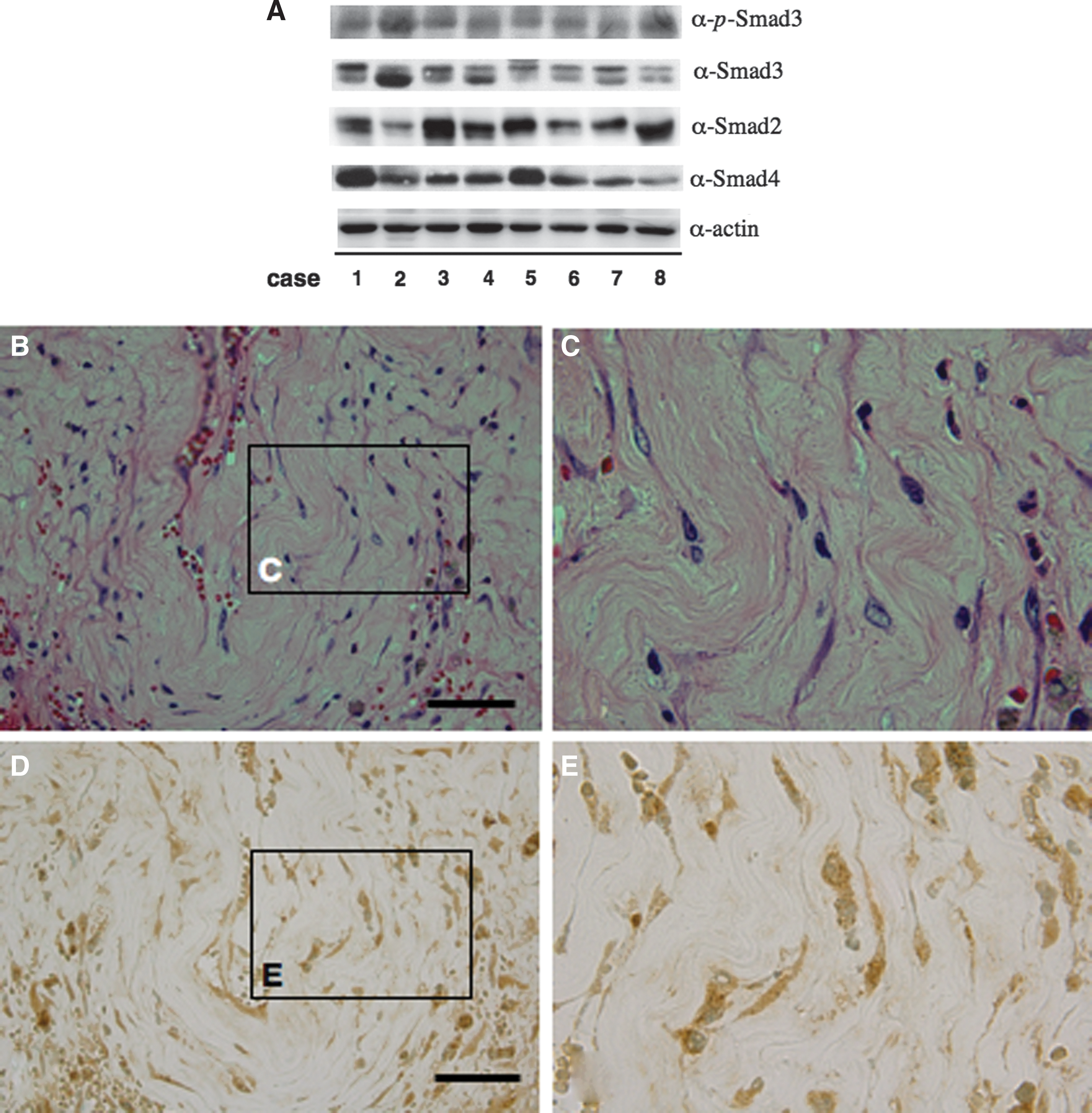
Outer membranes of chronic subdural hematomas from 8 patients homogenized in Laemmli sample buffer were subjected to Western blot with anti-actin (α-actin), anti-Smad2 (α-Smad2), anti-Smad3 (α-Smad3), anti-Smad4 (α-Smad4), and anti-phosphorylated Smad3 at Ser423/425 (α-p-Smad3) antibodies (
Histological observations
H&E staining showed that fibroblasts were well developed and were observed between collagenous fibers in CSDH outer membranes (Fig. 2B,C). Smad3 immunoreactivity was observed mainly in fibroblasts (Fig. 2D,E). IHC staining of chronic subdural membranes without primary Abs was consistently negative (data not shown).
Discussion
In this study, we clarified that eotaxin-3 and TGF-β were significantly increased in CSDH fluid. Smad2, Smad3, Smad4, and phosphorylated Smad3 were expressed in CSDH outer membranes, and Smad3 was expressed in fibroblasts of the CSDH outer membrane.
Subdural effusion is one of the main complications of brain trauma, which evolves into CSDH. The dural border cell layer, which is continuous with the dural mater externally and the arachnoid internally, is weaker and therefore even mild brain injury may cleave the cell layer, resulting in the formation of subdural effusion or CSDH. 15 Formation of CSDH is considered a self-perpetuating inflammatory process that is composed of coagulation, fibrinolysis, and repetitive bleeding. This process enlarges the hematoma cavity and induces the formation of neomembranes within the hematoma. Inflammatory cytokines and chemokines are elevated in the CSDH and may induce expressions of lymphocytes and eosinophils within the CSDH membranes. 1,2,5,16
Eotaxin, a potent eosinophil chemoattractant cytokine, has been detected in a guinea pig model of allergic airway inflammation. 17,18 Eotaxin selectively induces eosinophil, but not neutrophil, accumulation at sites of allergic inflammation. In epithelial cells, eotaxin was found at significantly higher levels in patients with severe asthma. 19 There are three potent eosinophilic chemoattractant isoforms: eotaxin-1, eotaxin-2, and eotaxin-3. In eosinophilic esophagitis, eotaxin-3 levels are strongly correlated with the extent of esophageal tissue eosinophilia. 20 Both eotaxin-2 and eotaxin-3 have differential profibrogenic effects on lung fibroblasts; eotaxin-2 promotes fibroblast proliferation and collagen synthesis, whereas eotaxin-3 induces fibroblast migration. 21 Eotaxin-3 is more effective at inducing the migration of eosinophils in asthmatics than eotaxin-1 and eotaxin-2. 22 Their different roles have not been fully defined, but our data suggest that eotaxin-3 may induce the expression of eosinophils and that activated eosinophils play a role in inflammation in CSDH outer membranes.
The multi-functional protein, TGF-β, plays a central role in the pathogenesis of chronic inflammation and fibrosis. Eosinophils have been identified as the major TGF-β-producing cell in lungs of asthmatics. 12 Eosinophils from patients with blood eosinophilia also express TGF-β, which may contribute to the development of the fibrosis observed in eosinophil-associated lesions. 23 Approximately 50% of the eosinophils infiltrating upper airway tissues express the TGF-β1 gene, 24 which is correlated with bronchial airway fibrosis and asthma severity. 12 Treatment with anti-TGF-β Ab has been shown to prevent the deposition of collagen through direct modulation of TGF-β signaling, suggesting that TGF-β is a profibrotic mediator. 25 Thus, the significantly increased TGF-β1 present in CSDH fluid may be produced by eotaxin-3 induction of eosinophils in CSDH outer membranes.
TGF-β1 signals through a heterodimeric complex of type I and II serine/threonine kinase receptors. Upon TGF-β1 binding to these receptors, activation of Smad2 or Smad3 occurs at the C-terminus. Smad2 or Smad3 form heterodimers with Smad4, and this Smad2/3-Smad4 complex translocates to the nucleus, where it regulates the transcription of target genes by binding directly to promoter regions or by associating with other transcription factors. 26 This Smad pathway is the main signaling pathway for TGF-β1. TGF-β1 has been reported to be involved in the scarring response in the injured rat brain. 27 Bleomycin-induced pulmonary fibrosis depends on increases in phosphorylation and nuclear accumulation of Smad2/3. 28 The TGF-β/Smad3 pathway is central to the pathogenesis of interstitial fibrosis of obstructive nephropathy. 29 The increased expression of TGF-β1 and phosphorylated Smad2/3 induced in pediatric eosinophilic esophagitis suggests a potential mechanism of fibrosis. 30 Smad3 deficiency attenuates bleomycin-induced pulmonary fibrosis in mice, further suggesting that disruption of the TGF-β/Smad3-signaling pathway may be a therapeutic target against fibrosis. 31 These previous data suggest that the TGF-β/Smad pathway contributes to fibrosis in different organs. As a whole, our immunoblot and IHC data suggest that the TGF-β/Smad-signaling pathway may play an important role in fibrosis in CSDH outer membranes. Moreover the TGF-β/activin receptor-like kinase 1–signaling pathway has been recently detected to be involved with angiogenesis in the CSDH outer membrane. 32 TGF-β plays multi-functional roles within CSDH outer membranes.
Conclusion
The present investigation revealed, for the first time, expression of eotaxin-3 in CSDH fluid. Eotaxin-3 induces expression of eosinophils in CSDH outer membranes, resulting in expression of TGF-β, which activates the Smad signaling pathway. The TGF-β/Smad pathway may be involved in the growth of CSDHs (Fig. 3). However, we have not ruled out the role of additional molecules in the activation of the TGF-β/Smad signaling pathway in CSDH outer membranes. Further studies using inhibitors of the TGF-β/Smad pathway, such as targeting the inhibition of eotaxin-3, might more precisely clarify the roles of eosinophils in CSDH outer membranes. We hope one could design a therapeutic strategy by understanding this growth mechanism of CSDHs.

Schematic drawing of the eotaxin/transforming growth factor-β1 (TGF-β1)/Smad-signaling pathway in chronic subdural hematoma (CSDH). Expression of eotaxin-3 in CSDH fluid induces eosinophils in CSDH outer membranes. Activated eosinophils produce TGF-β1 in CSDH fluid. TGF-β1 activates the Smad-signaling pathway in fibroblast within CSDH outer membranes. This signaling pathway might be involved in the growth of CSDHs.
Footnotes
Acknowledgment
The authors thank Hisae Inui for her technical assistance.
Author Disclosure Statement
No competing financial interests exist.