
Abstract
The biochemical and cellular events that lead to secondary neural damage after traumatic brain injury (TBI) contribute to long-term disabilities, including memory deficits. There is a need to search for single and/or combined treatments aimed at reducing these TBI-related disfunctions. The effects of citicoline and of voluntary physical exercise in a running wheel (3 weeks), alone or in combination, on TBI-related short-term (3 h) and long-term (24 h) object recognition memory (ORM) deficits and on neurogenesis and neuroprotection were examined using a rodent model of TBI (controlled cortical impact injury). Citicoline improved memory deficits at the two times tested, while physical exercise only in the long-term test. Physical exercise had a clear neuroprotective effect as indicated by reduced interhemispheric differences in hippocampal formation and lateral ventricle volumes and in density of mature neurons in the hilus of the dentate gyrus and the perirhinal cortex. Physical exercise also increased cell proliferation and neurogenesis in the granular cell layer of the dentate gyrus. Some degree of neuroprotection of citicoline was suggested by reduced interhemispheric differences in the volume of the hippocampal formation. Contrary to what was expected, the effects of citicoline and physical exercise did not sum up. Further, a negative interference between both treatments was found in several behavioral and histological variables. The promising profiles of both treatments as therapeutic tools in TBI when applied singly underscore the need to perform further works looking for other combined treatment regimens that increase the benefit of each treatment alone.
Introduction
T
The specific pathological features that ensue after TBI can be categorized as primary injury, which results from immediate mechanical damage, and secondary injury, which evolves over a period of hours to days and even months after the primary insult and is the result of biochemical and physiological events, such as ischemia, inflammation, altered neuronal and glial functions, loss of membrane integrity, etc., that ultimately lead to neuronal cell death. 5 Therefore, reduction of secondary injury, and the subsequent cognitive deficits associated with it, should be a highly relevant target of therapeutic approaches. Another treatment target should be related to the promotion of plasticity-related mechanisms and neurogenesis.
Physical exercise has been found to improve cognition in healthy animals 6 and to reduce memory deficits after brain insults, including TBI, in animals. 7 –9 Some evidence suggests that it can also confer benefits in humans. 10 These benefits seem to be mediated by a variety of mechanisms such as increased angiogenesis, cell proliferation and neurogenesis, enhanced synaptic plasticity, and regulation of inflammatory responses. 9,11 –15 Thus, physical exercise seems to be a promising tool in promoting functional recovery after TBI.
On the other hand, citicoline, the exogenous sodium salt of cytidine 5′-diphosphocholine (CDP-choline), seems to be capable of protecting, repairing, and rebuilding cell membranes, promoting synaptogenesis, decreasing apoptosis, regulating cerebral phospholipids, and supporting brain energy metabolism. 16 –18 Citicoline has been reported to decrease cognitive impairment 19 and improve neurological recovery 19,20 in rat models of TBI. There are also various reports of positive effects of this compound in patients with brain damage, 18 although negative results have also been reported. 21
It seems reasonable to assume that greater benefits for TBI-associated deficits may be conferred by combining two or more treatments endowed with either additive or complementary neuroprotective and neuroreparative actions compared with the use of a single treatment. One of these therapeutic approaches consists of combining a pharmacological agent such as citicoline, which can be administered shortly after the initial insult, with a long-term treatment such as physical exercise.
In this study, we tested the efficacy of post-injury citicoline and physical exercise, alone and in combination, to reduce TBI-induced object recognition memory (ORM) deficits in rats. We also examined the effects of both treatments on cell proliferation and neurogenesis in the dentate gyrus (DG). The putative neuroprotective effects of both treatments were also determined by quantifying the density of adult neurons in two brain areas related to ORM, 22,23 the perirhinal cortex (PRHc), and the hippocampal formation (specifically, the hilus of the DG-HDG), and by examining gross morphological alterations in the brains.
Methods
Ethics and animal welfare
All procedures were performed in compliance with the European Community Council Directive for care and use of laboratory animals (86/609/EEC), and with the related directive of the Autonomous Government of Catalonia (DOGC 2073 10/7/1995).
Forty-eight male Sprague-Dawley albino rats (Prolabor, Barcelona, Spain), 6-weeks old on their arrival to the laboratory, were used. On arrival, they were kept in the quarantine room for 1 week. Therefore, the age of the animals at the beginning of the experimental procedures was 7 weeks, and their mean initial body weight was 251.18 g (standard deviation [SD]±31.32). Food and water were available ad libitum. The animals were kept under conditions of controlled temperature (20–22°C) and humidity (40–70%), and maintained on a 12-h light-dark cycle (lights on at 8:00 a.m.).
Experimental groups
The animals were randomly assigned to one of five experimental conditions, according to whether they were sham operated (Sh) or received TBI (Lesion, L), and according to the treatment administered: citicoline (C) vs. vehicle (V), and exercise (E) vs. sedentary (S) conditions. Animals in the ShVS group (n=11) were sham operated, received vehicle injections, and were sedentary. Animals in the remaining groups underwent TBI and received either no treatment (LVS group; n=11), citicoline (LCS group, n=9), exercise (LVE group, n=9), or the combination of both treatments (LCE group, n=8).
Stereotaxic surgery and TBI model
Controlled cortical impact injury (CCI) was used as the animal model of TBI. For stereotaxic surgery, anesthesia was induced with 5% isoflurane (Forane, Abbot Laboratories, SA, Madrid, Spain) in oxygen (2 L/min) in a Plexiglas chamber (20×13×13 cm) for 7 min. The animals were then placed in a stereotaxic frame (David Kopf Instruments, Tujunga, CA), and the anesthesia was continued by delivering 2% isoflurane in oxygen (1 L/min) through a nose mask. The scalp was incised on the midline, and after the skull was exposed, a craniectomy (4 mm diameter) was performed over the right hemisphere (4.5 mm posterior to bregma and 3 mm from midline).
According to the parameters used by Yu and colleagues, 24 the pneumatically operated TBI device (Pittsburgh Precision Instruments, Inc, Pittsburgh, PA) with a 3-mm tip diameter impacted the brain at a velocity of 6.0 m/sec reaching a depth of 2.0 mm below the dura mater layer, and remained in the brain for 150 msec. The impactor rod was angled 15 degrees to the vertical to maintain a perpendicular position in reference to the tangential plane of the brain curvature at the impact surface. A transducer connected to the impactor measured velocity and duration to verify consistency. Thereafter, the scalp was sutured. To control for postoperative pain, a single 0.2 mL subcutaneous injection of buprenorphine (Buprex, Schering-Plough, SA, Madrid, Spain) was administered. Animals of the ShVS group underwent operation in a similar way, except that no impact was applied.
Citicoline vs. vehicle condition
Animals received intraperitoneal (ip) injections of either citicoline (Ferrer Interquim SA, Barcelona, Spain) or vehicle (saline) starting 4 h after surgery, and thereafter every 24 h until completing five injections. Citicoline (200 mg/kg) was dissolved in physiological saline (volume 2 mL/kg).
Physical exercise versus sedentary condition
After the last citicoline or vehicle injections, and thus 4 days after surgery, animals in the exercise condition were placed in a cage (48×26×20 cm) connected to a 37-cm diameter running wheel (Rat Wheel W/Brake, ENV-042, Med Associates, Inc, St. Albans, VT), while rats in sedentary groups were kept in their home cages (52×28×18 cm). Each wheel had a counter that allowed recording of daily running distance and running time. The rats remained in these conditions until the end of the experiment (i.e., for a total of 20 days).
Neurological assessment
The animals underwent neurological assessments at two different times: 24 h and 20 days after surgery. Neurological assessments consisted of the grip strength test and a composite neurological score (neuroscore, adapted from Marklund and associates 25 ). For the grip strength test (Bioseb, Paris, France), which assesses muscular strength of rats, the animal forepaws were placed on a bar connected to a dynamometer. Thereafter, animals were pulled out from the bar by an experimenter. The maximal peak force displayed by the animal to keep gripping the bar was recorded.
The neuroscore was composed of forelimb reflex, hindlimb reflex, and lateral pulsion tests. The forelimb reflex was tested by suspending the animals by the tail with the forepaws near a flat surface (a table). Intact rats stretch both forepaws at a level higher than the nose to stay on the table. The hind limb reflex test examined the strength of a backward extension of the hind limbs when the animal's tail was lifted and the forelimbs remained on a hard surface. Lateral pulsion evaluated the animal's ability to resist hand pressure to move from left to right, and vice versa.
For the three tests, separate scores were taken for the right side of the body (ipsilateral to the cortical impact), and the left side (contralateral to the impact). A score from 0 to 4 was given for each task according to the following criteria:
Forelimb reflex
4, fully extends the limb toward the table; 3, extends forelimb toward the table, but without enough strength to prevent nose from touching surface; 2, incomplete extension of forelimb so that the nose touches surface before the paws; 1, minimal extension of forelimb; 0, no forelimb extension.
Hindlimb reflex
4, normal backward and outward extension of the hindlimb; 3, partial extension of the hindlimb; 2, extension is initiated, but not maintained; 1, no hindlimb extension; 0, retraction of the hindlimb toward the abdomen.
Lateral pulsion
4, normal resistance to lateral push; 3, low resistance to lateral push, but without falling; 2, low resistance to lateral push with fall, but regaining normal position rapidly; 1, the animal falls and regains normal position with difficulty; 0, the animal falls and is unable to regain normal position.
Thus, a maximum score of 12 could be obtained for each side.
Bromodeoxyuridine (BrdU) injections
To assess the degree of cell proliferation in the DG, the animals received five ip injections, one daily, of BrdU (Sigma-Aldrich, Madrid, Spain) at a dose of 200 mg/kg and a volume of 20 mg/mL, dissolved in phosphate buffer saline (PBS). BrdU is a substance that is incorporated into the DNA during S phase of the cell cycle, thus allowing the immunohistochemical identification of new cells. 26 The first injection was given on day 11 of the exercise/sedentary condition. This moment of administration was chosen because cell proliferation increases induced by exercise seem to peak during the second week of treatment 27 as well as to minimize any interference with a possible increase in proliferation from TBI. 28
ORM
The ORM procedures started on day 18 of the exercise/sedentary condition, and 3 days after the last BrdU injection. Training was carried out in an open box (65.5 cm width×65.5 cm length×35 cm height) made of a conglomerate covered with brown melamine, and enclosed in a sound-attenuating cage (72 cm width×72 cm length×157 cm height) made from white melamine, and ventilated by an extractor fan. The illumination on the floor of the box was 30 lux. The objects used varied in shape, color, and size, and consisted of Lego pieces, a hanger, and a drinks can. They were fixed to the floor of the box with double-sided adhesive tape so that the rats could not move them.
The objects were not known to have any ethological significance for the rats, and they had never been seen by the animals. A prior pilot study had shown that rats of the same strain and age had no spontaneous preference for any of them. The objects for the recognition task were available in duplicate copies.
All behavioral sessions were recorded with a video camera mounted above the experimental apparatus. Tapes were analyzed off-line by a trained observer who was unaware of the treatment condition. Throughout the experiment, exploration of an object was defined as directing the nose to the object at a distance ≤2 cm or touching it with the nose. Turning around or sitting on the object was not considered exploratory behavior. To avoid the presence of olfactory cues, the apparatus and objects were thoroughly cleaned with a solution of 70% alcohol in distilled water and dried before placing the first rat and after testing each animal.
Habituation to the apparatus
To habituate the animals to the experimental box, three habituation sessions were performed (two on the same day, separated by a 2-h interval, and the third one on the following day). The animals were introduced into the recognition memory box, under the same lighting and sound conditions as during training, but without any objects, and were allowed to explore it for 12 min.
Neophobia test
To test possible anxiety reactions to novel objects, a neophobia test was conducted 2 h after the last habituation session. 29 An unfamiliar object was exposed in the center of the open box. The animals were placed in the box facing away from the object and allowed to explore for 10 min. Total time exploring the object was recorded.
Acquisition and memory tests
ORM training began 24 h after the neophobia test. During the acquisition session, two identical objects were placed near the two corners at either end of one side of the cage. The rat was placed in the experimental apparatus, facing the center of the opposite wall, and was allowed to explore for 15 min. The time spent in exploring each object was recorded.
Two memory tests were performed, the first one 3 h after the sample trial, and the second one 24 h after it (that is, 21 h after the first test). In both retention tests, one copy of the object used on the acquisition session (familiar object) and a new object, different for each retention session, were placed in the same locations as the stimuli used during the training trial. The specific objects used as either familiar or novel, as well as their locations, were used in a balanced manner to reduce potential biases because of preferences for particular location or object.
The rats were placed in the open box for 5 min. Time spent exploring each object was recorded. To analyze cognitive performance, a discrimination index that made it possible to adjust for any differences in total exploration time was calculated ([(time exploring the novel object–time exploring the familiar object)/total time spent on both objects]×100). Because ORM is based on the natural tendency of rats to explore novelty, an index significantly higher than zero (i.e., animals exploring the novel object more than the familiar one) is considered a good recall of the familiar object, whereas an index close to zero (i.e., animals exploring both objects similarly) is considered a lack of recall. 29
Brain processing
Twenty-four hours after the second memory test, the animals were killed with an overdose of sodium pentobarbital (Dolethal, 200 mg/kg; Vetoquinol SA; Madrid, Spain) and intracardially perfused with 4% paraformaldehyde (PFA; Sigma-Aldrich, Madrid; Spain) in PBS. The brains were then extracted and submerged in a 4% PFA solution. Three hours later, they were rinsed with phosphate buffer three times every 20 min, and stored at −80°C in a cryoprotective solution (sucrose 30% in phosphate buffer). Coronal slices, 40 μm width, were obtained using a cryostat (Shandon Cryotome FSE, Thermo Electron Corp, Waltham, MA), and stored in Olmos antifreeze solution.
Nissl staining
To examine the macroscopic effects of TBI, 1 of every 10 coronal sections throughout the extent of brain tissue where the lesion cavity was visible were stained with cresyl violet in all the animals of the four TBI groups. These sections were digitalized with a scanner (HP Scanjet G4050). Using Fiji image analysis software, 30 digital images were calibrated, and the areas of the following regions in the hemispheres ipsilateral and contralateral to the cortical impact were measured: lesion cavity, hippocampal formation, and lateral ventricle. For volume calculations, the areas obtained in each slice were multiplied by 0.04 mm (slices width) and by 10 (number of sections until the next slice analyzed).
For each section and structure (except for the lesion cavity), an interhemispheric ratio score was computed ([ipsilateral area/contralateral area]×100). The mean ratio scores for all the sections in each rat were used for a more standardized comparison between groups of animals. Given that these ratio scores are expected to be significantly similar to 100 if there is no volume loss because of brain damage, and significantly different from 100 if otherwise, one-sample t tests were also used to determine whether interhemispheric ratio scores for each group were statistically different from 100.
BrdU, doublecortin (DCX), and neuronal nuclei (NeuN) immunohistochemistry
Triple immunofluorescent staining was performed for BrdU, DCX, and NeuN. BrdU allows quantifying the amount of cell proliferation along the period of BrdU administration (between days 11 to 15 of the exercise or sedentary conditions). DCX, which is a microtubule associated phosphoprotein that is selectively expressed by immature neurons, 31 was used to determine the amount of new cells that had developed into neurons. Finally NeuN, which is a nuclear marker of mature neurons, 32 was used as a measure of neuroprotection.
Five coronal slices of 40 μm (600 μm between slices) were taken starting at AP coordinate −2.52. 33 For DNA denaturation, the sections were submerged into a 1 M HCl solution for 1 h at 37°C. They were then submerged in 0.1 M borate buffer (pH 8.5). To reduce autofluorescence, the sections were rinsed with 0.3 M glycine (Sigma Chemical, Madrid; Spain) for 30 min. To block nonspecific binding, they were then washed in 10% fetal bovine serum (Sigma-Aldrich, Madrid, Spain) in PBS-T. The sections were incubated overnight at 4°C and 1 h at room temperature with primary antibodies to BrdU (1:2000 dilution; rat anti-BrdU ab6326 Abcam), DCX (1:4000 dilution; rabbit anti-DCX ab 18723 Abcam), and NeuN (1:1000 dilution; mouse anti-NeuN Mab 377 Chemicon).
Fluorescent secondary antibodies to BrdU, DCX, and NeuN (1:4000 dilution; goat anti-rat IgG Dylight 488 Jackson ImmunoResearch, goat anti-mouse IgG Dylight 594 Jackson ImmunoResearch, and goat anti-rabbit IgG Dylight 649 Jackson ImmunoResearch, respectively) were used to incubate the slices for 2 h at room temperature. Sections that were not incubated with the primary antibodies were used as negative controls. Finally, the sections were incubated with 4,6- diamidino-2-phenylindole (DAPI, Sigma Chemical; Madrid; Spain) for 5 min. The sections were mounted on slides with Fluoromount mounting media (Sigma Chemical, Madrid; Spain).
Image processing and cell quantification
The number of BrdU+ cells was counted in the granular cell layer of the DG of the ipsilateral and contralateral hemispheres in each of the five coronal slices processed for immunohistochemistry. The slices were examined using a confocal microscope (Olympus Fluoview 1000) and a 20x magnification. Photographs were taken, and the number of BrdU+ cells of the granular cell layer was manually counted in each hemisphere.
To quantify the number of double-labeled BrdU/DCX cells, images of oil immersed sections were taken with a confocal microscope (Olympus Fluoview 1000) at 60x magnification. For each hemisphere, BrdU+ cells were evaluated for double staining with DCX. In those animals with more than 25 BrdU+ cells per hemisphere, 25 positive cells were randomly selected through the five sections, and the number of BrdU+-DCX+ cells was estimated from percent BrdU+ cells that were also stained for DCX. NeuN+ cell density in HDG was quantified in two slices randomly chosen from those used to quantify BrdU+ cells.
A Fiji macro was designed ad hoc and used for all the images, and an ROI was drawn around the HDG. For each slice, the density of NeuN+cells in the ipsilateral and contralateral HDG was quantified and an interhemispheric ratio value ([ipsilateral NeuN density/contralateral density]×100) was calculated. The mean ratio was taken as a measure of the amount of neuronal loss. A ratio statistically similar to 100 indicates that the density of NeuN+ cells in both hemispheres is similar, while a ratio statistically lower than 100 indicates that the density of NeuN+ cells in the ipsilateral hemisphere is significantly lower than that of the contralateral hemisphere.
The process to quantify NeuN+ cells in the PRHc was similar to that used in HDG, except that a single image at 10x magnification was obtained from two slices randomly chosen from those used to quantify BrdU+ cells, excluding the first slice (Bregma −2.52 mm), because this structure is not visible in this coordinate. The Fiji macro was adjusted for this region, and a circular ROI situated inside the PRHc and identical for all the subjects was considered for all the images. Density ratio values as explained for HDG NeuN+ cells were also calculated for the PRH and used for the statistical analyses.
Statistical analyses
The statistical package SPSS (IBM statistics, Chicago, IL) was used for data analyses. The data were mainly analyzed with either a mixed design analysis of variance (MANOVA, with a between-group factor, composed of five categories, each of the five experimental groups, and a within-group factor, the repeated measures taken), or a univariate analysis of variance (a single between-group factor), when a single dependent variable was tested.
Paired comparisons (Bonferroni) for ANOVA, or simple contrast analyses for MANOVA, were performed when the between-group factor was significant. Polynomial contrasts were also used in MANOVA when the within-group factor had more than two levels and was significant. One-sample t tests were used to determine whether the values of a specific group were statistically different to a given reference value. Finally, correlation analyses were also used for several measures. Statistical significance was set at the level of p≤0.05.
Results
Neurological assessment
The first neurological assessment was performed 24 h after surgery—i.e., when the exercise condition had not yet been introduced. Therefore, for the statistical analyses of the first neurological assessment, data from LCS and LCE groups were pooled, and the same was done with the data from LVS and LVE groups. The analyses of the second neurological assessment were performed in the same manner, comparing data from the five experimental groups.
ANOVA for the first neurological assessment indicated significant differences between conditions with regard to ipsilateral neuroscore [F(2, 45)=14.51; p<0.001] and contralateral neuroscore [F(2,45)=10.93; p<0.001], while no differences were found for grip test between conditions. Post hoc paired comparisons (Bonferroni) indicated that the ipsilateral neuroscore was significantly lower in TBI-vehicle (p<0.001) and TBI-citicoline (p<0.001) conditions compared with sham-vehicle-sedentary animals, while no statistical differences were found between TBI-citicoline and TBI-vehicle conditions.
With regard to contralateral neuroscore, TBI-citicoline condition had significantly lower values than TBI-vehicle (p<0.010) and sham animals (p<0.001). No significant difference has been observed between sham and TBI-vehicle condition. ANOVA for the second neurological assessment indicated the lack of between-group differences on any measurements.
Running behavior
For the statistical analyses, mean values of running distances and running times were calculated for the following three periods: days 1 to 6, days 7 to 13, and days 14 to 20. With regard to running distances, MANOVA did not find statistical differences between LVE and LCE groups. No significant interaction was observed between group×exercise period. In contrast, exercise period factor was significant [F (2,30)=27.97; p<0.001].
To analyze the evolution of running distances across the three periods, polynomial contrasts were performed. These contrasts indicated that running distances adjusted to a linear function (p<0.001), with running distances increasing across exercise periods in both groups. Mean (SD) daily running distances pooled for both exercising groups were 1369.61 (±785.46) m on days 1–6, 4329.58 (±3032.12) m on days 7–13, and 6289.07 (±3958.73) on days 14–20.
Similar results were found for mean running time. Thus, MANOVA indicated that exercise period factor was significant [F(2,30)=31.14; p<0.001], while neither group, nor the group×exercise period interaction were significant. Polynomial contrasts indicated that mean running time increased linearly across the three exercise periods p<0.001). Mean (SD) daily running times pooled for both exercising groups were 47.87 (±26.66) min on days 1–6, 126.66 (±76.32) min on days 7–13, and 170.18 (±95.26) min on days 14–20.
Recognition memory
Exploration during neophobia test, acquisition session, and retention trials
No significant differences among groups were found in ANOVA for the amount of exploration in the neophobia test and acquisition trial.
With regard to the retention tests, MANOVA indicated that neither the main factor group nor the group×session interaction were significant. In contrast, the session factor was significant [F(1,41)=10.39; p=0.002], indicating that exploration times were higher in the first retention trial compared with the second retention trial.
Discrimination indices in the retention trials
Two animals (one in LVS and another one in LCE group) failed to explore any of the objects during the retention trials. Because object recognition memory is based on exploration measurements and discrimination indices cannot be calculated when total exploration time is 0, these animals were excluded from the statistical analyses. Therefore, the number of subjects used for the analyses of retention trials was 46.
As indicated in Figure 1, the discrimination index of the ShVS group was significantly different from zero (indicating a significant recall) on both the first (3 h) [t(10)=4.24; p=0.002], and the second (24 h) [t(10)=5.32; p=0.002] retention tests. In contrast, the discrimination index of LVS group was not significantly different from zero in any of the two sessions, indicating a lack of recall. With regard to drug-treated TBI animals, the discrimination index was significantly different from zero only in the first retention test, but not in the second retention test, in LCS [first retention test: t(8)=4.45; p=0.002], and LCE [first retention test: t(6)=3.87; p=0.008] groups. In contrast, the discrimination index of LVE animals was significantly different from zero only in the second retention [t(8)=3.94; p=0.004], but not in the first retention test.
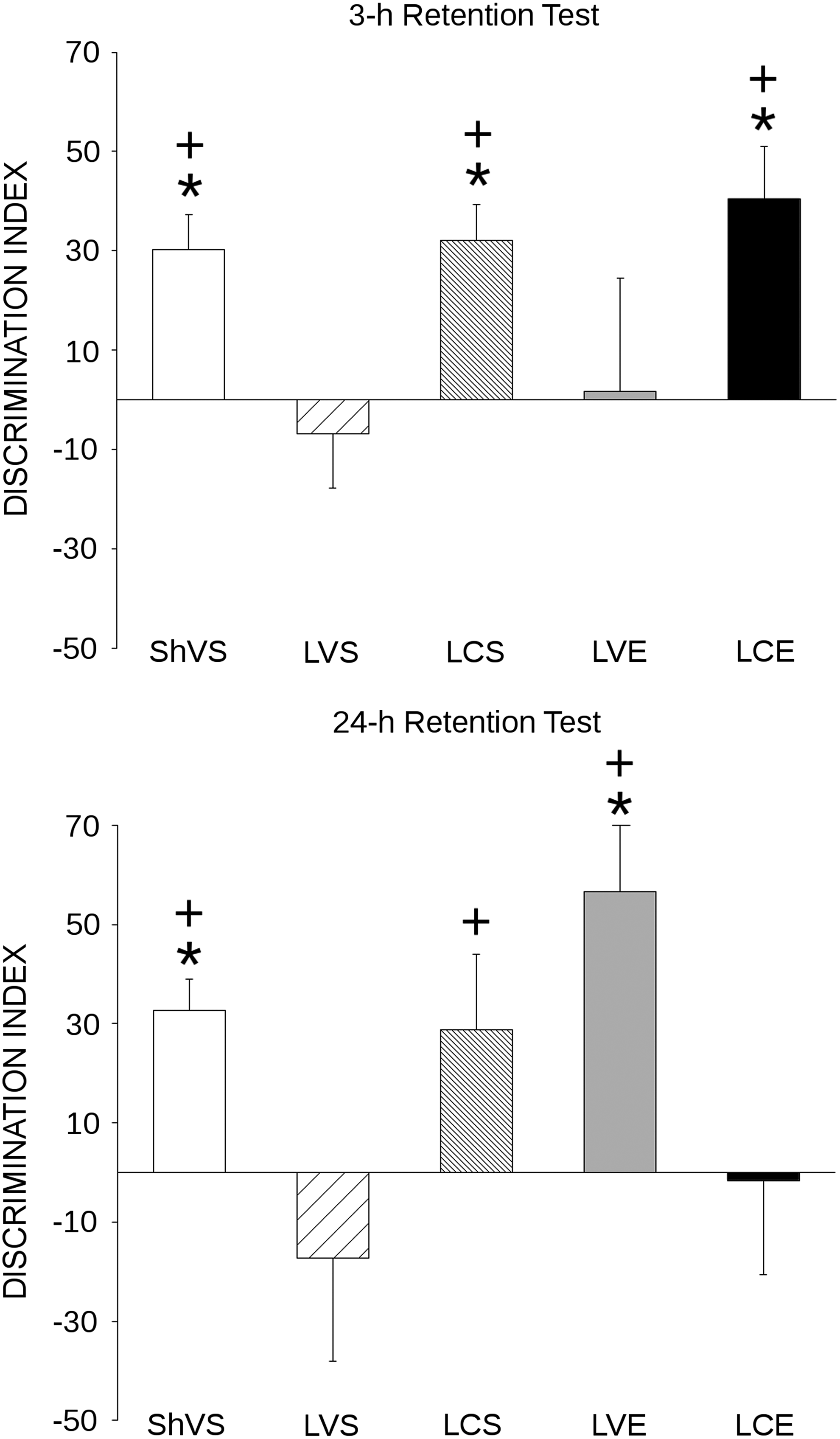
Recognition memory. Mean values of discrimination indices for each experimental group in the 3-h (top) and 24-h (bottom) object recognition memory tests. Animals in sham-operated vehicle sedentary (ShVS) group remembered the familiar object in both retention tests, while animals in the lesion vehicle sedentary (LVS) group (rats with traumatic brain injury [TBI] that did not receive any treatment) failed to remember the familiar object in both retention tests. Citicoline improved object recognition memory (ORM) in both tests (lesion citicoline sedentary [LCS] and lesion citicoline exercise [LCE] groups), while voluntary physical exercise (lesion vehicle exercise [LVE] group) reversed the TBI-related memory deficit on the second, but not on the first, retention test. Finally, there was a negative interaction between citicoline and physical exercise on ORM on the 24-h retention test (LCE group). (+Different from LVS group, p<0.05; *different from 0; p<0.05).
Mixed MANOVA showed a significant main effect of group [F(4,41)=2.93; p=0.032] and a significant group×session interaction [F(4,41)=3.58; p=0.014], while the main session factor was not significant. To better determine the direction of this interaction, simple contrast effects were calculated within each of the two retention sessions to compare each experimental group with the untreated TBI animals (LVS group), which were expected to have the worst performance.
These contrasts indicated that the discrimination index of the ShVS group was significantly higher than that of the LVS group both in the first [F(1,41)=4.72; p=0.036] and in the second [F(1,41)=5.98; p=0.019] retention sessions. The performance of LCS group was also significantly higher than that of LVS group in both retention sessions [first retention session: F(1,41)=4.74; p=0.035; second retention session: F(1,41)=4.59; p=0.038], while the discrimination index of the LCE group was higher than that of LVS group only in the first retention session [F(1,41)=6.05; p=0.018], but not in the second session. With regard to the LVE group, it had a discrimination index that was significantly higher than that of the LVS group only in the second retention session [F(1,41)=11.78; p=0.001], but not in the first retention session.
Neuroprotection-related variables
Cresyl-violet staining
In one of the subjects of the LVE group, it was not possible to observe the hippocampal formation and lateral ventricle in violet-stained sections because of the extreme expansion of the lateral ventricle and to extremely aberrant morphology of the hippocampal formation. Therefore, statistical analyses were performed with 36 animals in TBI groups for the cresyl-violet data.
Figure 2A shows a photograph of one coronal slice stained with cresyl violet in a representative animal with TBI. Besides a lesion cavity, this figure shows the characteristic increase in volume of the lateral ventricle, as well as altered morphology in the hippocampal formation found in the ipsilateral hemisphere.
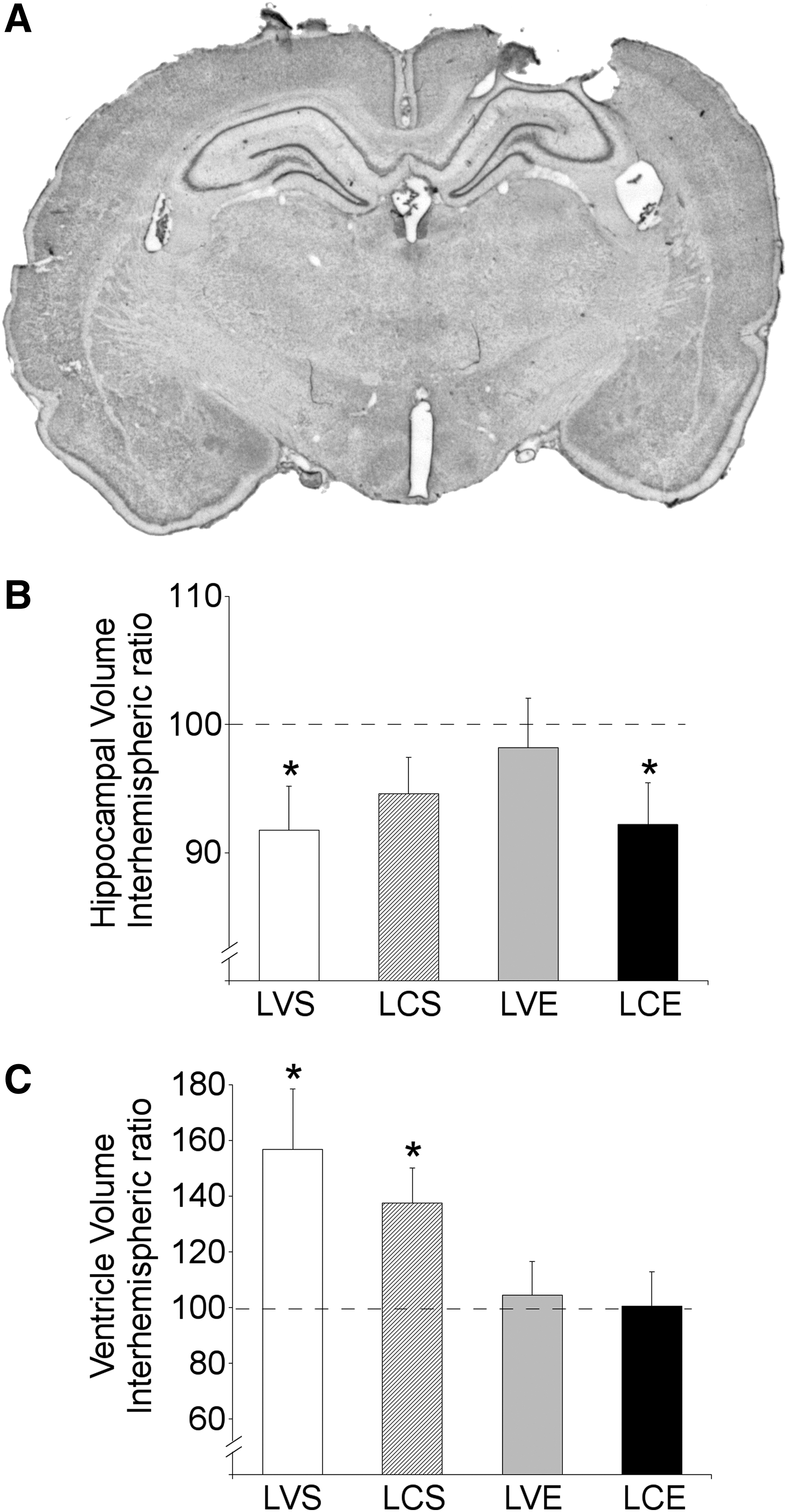
Gross anatomical assessment of neuroprotection. (
ANOVA did not show any statistical difference between TBI groups in mean volumes of lesion cavity, as well as in interhemispheric ratios for hippocampal formation (Fig. 2B), and lateral ventricle volumes (Fig. 2C).
One-sample t tests were performed separately for each group to examine whether interhemispheric ratio volumes were statistically different from 100 (the expected value if there was no tissue loss). These ratio values were significantly different from 100 for both the hippocampal formation and the lateral ventricle in the LVS group [hippocampus: t(10) = −2.41; p=0.037, lateral ventricle: t(10)=2.63; p=0.025]. In the LCS, group the interhemispheric ratio was only significantly different from 100 for the lateral ventricle [t(8)=2.97; p=0.018], but not for the hippocampus, while in the LCE group, the interhemispheric ratio was significantly different from 100 for the hippocampal formation [t(7) = −2.40; p=0.047], but not for the lateral ventricle. Finally, the LVE group ratio values for the hippocampal formation and the lateral ventricle did not significantly differ from 100.
NeuN+ cells in the HDG
Quantification of NeuN+ cells in the HDG was not possible in one animal of the LVE group because of an extremely aberrant morphology of the hippocampal formation. Therefore, statistical analyses for this variable were performed with 47 subjects.
Figure 3 depicts a sample ROI used to determine the boundaries of the HDG (Fig. 3A) and photographs (Fig. 3B) of HDG stained for NeuN of a representative animal in each group. The mean ipsilateral/contralateral ratio values for NeuN+ cell densities in the HDG for each experimental group are depicted in Figure 3C.

Cellular assessment of neuroprotection in the hilus of dentate gyrus (HDG) showing an important cell loss in lesion groups, especially in the sedentary groups (lesion vehicle sedentary [LVS] and lesion citicoline sedentary [LCS]). (
One-sample t tests indicated that all TBI groups had ratios significantly lower than 100, indicating that the NeuN+ cell density was lower in the ipsilateral than in the contralateral hemisphere: LVS [t(10) = −6.86; p<0.001]; LCS [t(8)=−4.09; p=0.003], LVE [t(7)=−2.67; p=0.032], and LCE [t(7)=−5.67; p=0.001]. In contrast these values were not significantly different from 100 in ShVS animals.
ANOVA indicated the existence of significant differences between groups [F(4, 42)=4.91; p=0.002]. Post hoc paired comparisons (Bonferroni) indicated that the ratio of HDG NeuN+ cell density was significantly higher in the ShVS group than in the LVS (p=0.002), and LCS (p=0.024) groups, while no significant differences were found between ShVS and LVE groups. Finally, the difference between the ShVS and LCE groups tended to approach significance (p=0.073). No other significant paired comparisons were found.
NeuN+ cells in the PRHc
Figure 4 shows photographs stained for NeuN of a representative animal in each group (Fig. 4A), and the interhemispheric ratio for mean NeuN+ cell densities in the PRHc for each experimental group (Fig. 4B).

Cellullar assessment of neuroprotection in the perirhinal cortex (PRHc). (
One-sample t tests indicated that LVS [t(9)=−2.53; p=0.032], and LCS groups [t(8)=−5.77; p<0.001] had ratios significantly lower than 100. In contrast, the ratio in ShVS, LVE, and LCE groups did not differ statistically from 100. ANOVA did not find significant differences between groups.
Cell proliferation and neurogenesis in the DG
Figure 5A shows photographs of immunofluorescent sections of the DG labeled for DAPI, BrdU, and DCX for one representative animal of each experimental group. The mean total number of BrdU+ and of BrdU+-DCX+ cells for each group are depicted in Figures 5B and 5C, respectively.

Cell proliferation and neurogenesis in the dentate gyrus (DG). (
No significant differences between groups were found with regard to the proportion of BrdU+ cells that coexpressed DCX, either in the ipsilateral or in the contralateral DG.
ANOVA showed significant between-group differences in the number of BrdU+ and of BrdU+-DCX+ cells both in the ipsilateral [BrdU+: F(42,4)=4.59; p=0.004; BrdU+-DCX+: F(42,4)=4.98; p=0.002], and in the contralateral DG [BrdU: F(42,4)=3.16; p=0.023; BrdU+-DCX+: F(42,4)=3.83; p=0.001], as well as in the total number of BrdU+ [F(42,2)=4.04; p=0.007] and BrdU+-DCX+ [F(42,4)=4.47; p=0.004] cells.
Post hoc paired comparisons (Bonferroni) indicated that the number of BrdU+ cells in the ipsilateral DG, as well as the total number of BrdU+ cells, were significantly higher in the LVE group compared with ShVS (ipsilateral: p=0.036; total: p=0.014), LVS (ipsilateral: p=0.006; total: p=0.021), and LCS (ipsilateral: p=0.005; total: p=0.015) groups. The LVE group also had a significantly higher number of BrdU+ cells in the contralateral DG than ShVS group (p=0.025), while the differences between LVE and LCS (p=0.070), and between LVE and LVS groups (p=0.065) only tended to approach significance. The remaining paired comparisons were also not significant.
With regard to BrdU+-DCX+ cells, post hoc paired comparisons indicated that the LVE group had significantly higher values in the ipsilateral and contralateral DG, as well as in the total number of these cells, compared with ShVS (ipsilateral: p=0.038; contralateral: p=0.018; total: p=0.015); LVS (ipsilateral: p=0.003; contralateral: p=0.017; total: p=0.006), and LCS (ipsilateral: p=0.004; contralateral: p=0.038; total: p=0.013) groups. No other significant paired comparisons were found.
A positive significant correlation (Pearson, unilateral) was found between the discrimination index in the 24-h retention test and each of the following variables: total number of BrdU+-DCX+ cells (r=0.267; p=0.038), number of BrdU+-DCX+ cells in the contralateral DG (r=0.269; p=0.037), percent BrdU+ cells that also expressed DCX in the ipsilateral (r=0.034; p=0.010) and the contralateral (r=0.315; p=0.018) hemispheres. No significant correlations were found between the discrimination index in the first retention test and any of the values related to cell proliferation and neurogenesis.
In summary, physical exercise increased cell proliferation and neurogenesis in the granular cell layer of the DG in TBI animals, compared with sham animals, untreated TBI rats, and TBI animals in the sedentary condition that had received citicoline. Moreover, there was a correlation between the number of newly born neurons and ORM in the second retention test.
Discussion
There is a need to search for single and/or combined treatments aimed at reducing TBI-related chronic disruption of memory and other cognitive functions. In this work, the efficacy of chronic voluntary physical exercise (wheel running) and the agent citicoline, either alone or in combination, to reduce ORM deficits has been tested in a rat model of TBI.
TBI induced severe deficits in both short- and long-term ORM 3 weeks post-injury. We also observed an impairment of neurological function 24 h after surgery that was no longer apparent after 20 days. These data suggest that the specific CCI parameters and location of impact had caused a severe cognitive deficit and a mild to moderate sensoriomotor impairment.
Both citicoline and physical exercise reversed the severe ORM deficits induced by TBI. The effect of citicoline, however, was higher in the 3-h retention test, while the effects of physical exercise were only found in the 24-h retention test. Any influence of either treatment on total exploration times and neophobia can be disregarded as mediating mechanisms for improved performance.
Physical exercise also increased cell proliferation and neurogenesis in the DG, and a positive correlation was found between neurogenesis and memory in the 24-h retention test. This treatment also had a neuroprotective effect, as evidenced by the following results: (1) while all TBI groups showed a reduced density of NeuN+ cells in ipsilateral vs. contralateral HGD, physical exercise attenuated this neuron loss, because no significant differences where found between sham-operated animals and the exercising groups; (2) in both exercising groups, NeuN density ratio in the PRHc did not show the neuronal loss seen in TBI sedentary groups; and (3) physical exercise also attenuated the TBI-related reduction of hippocampal volume and the TBI-related expansion of the lateral ventricle.
Citicoline attenuated TBI-related reduction of ipsilateral hippocampus volume in sedentary animals, suggesting a small neuroprotective effect. In contrast, this compound neither increased cell proliferation and neurogenesis in the DG, nor normalized the NeuN+ cell density interhemispheric ratios in the HDG and the PRHc. It also did not reduce expansion of the lateral ventricle. Contrary to what was expected, however, the combination of physical exercise and citicoline failed to induce additional benefits compared to each treatment alone. Further, a negative interaction between both treatments was found in the 24-h memory test and in some histological measurements.
Neurogenesis and neuroprotection may have mediated the benefits of voluntary physical exercise on ORM in animals with TBI
As has been found in healthy animals, 34 physical exercise increased cell proliferation (number of BrdU+ cells) and neurogenesis (number of BrdU+-DCX+ cells) in the granular cell layer of the DG in animals with TBI. Cell proliferation and neurogenesis in the perilesional area have also been reported after TBI in exercising animals. 35 A work conducted on mice 14 found that voluntary physical exercise increased cell proliferation and neurogenesis in the granular cell layer of the DG, but only when physical exercise was initiated 5 weeks after TBI induction and not when initiated with a delay of only 1 week. This is in contrast to the enhancing effects of neurogenesis found in the present work with an exercise regimen initiated 5 days after injury.
It has been suggested that new immature neurons in the DG dentate are relevant for memory formation, with 1- to 2-week–old new neurons playing an essential role in these processes (see Aasebø and associates 36 for a review). Regarding ORM, several articles show that very young neurons (less than 10 days old) could be relevant for long-term memory formation. 37 –39 In our experiment, the BrdU+-DCX+ cells were 5 to 9 days old on the acquisition session, but it has to be taken into account that exercise not only increases cell proliferation and survival of the new neurons, but also speeds up the maturation and the incorporation of the neurons into functional circuits. 40
In view of these data, the positive correlation found between the number of BrdU+ -DCX+ cells and the discrimination index in the 24-h retention test (but not in the 3-h retention test, which was not improved by exercise) suggests that novel neurons born during exercise may have contributed to cognitive recovery. This is in line with the fact that immature neurons of the subgranular cell layer of the DG (which are DCX+) exhibit hyperexcitability and enhanced synaptic plasticity, two properties that may give them advantages over mature neurons for selective formation of synaptic connections and for information processing. 41
As stated above, the data on gross volumetric measurements and the NeuN+ density ratios in the HDG and the PRHc suggest a neuroprotective effect of physical exercise on two regions involved in ORM. Neuroprotective effects of both forced and voluntary physical exercise have also been reported in other works, mainly when this treatment is applied before TBI. 42 There are also a few reports that physical exercise applied after brain damage may reduce neuron loss and inflammatory responses. For example, both forced and physical exercise reduced neuron loss after TBI 9,43,44 and after hippocampal domoic acid damage. 45 Piao and colleagues 14 have also reported neuroprotective effects of voluntary physical exercise after TBI in mice.
In contrast to the present results, neuroprotection was only observed when exercise was initiated 5 weeks after trauma, while an earlier initiation (1week) led to increased inflammatory responses. These contradictory data underscore the need to better determine the variables that mediate the timing-related effects of physical exercise on cognition, neurogenesis, and neuroprotection after TBI.
In any case, it seems that the cascade of events that, although initiated early after trauma, gives rise to secondary neuron loss during extended periods 46 can be arrested or attenuated with physical exercise. 9,14 This is of outstanding importance, given the chronic nature of most TBI sequels and the feasibility of physical exercise as a long-term treatment in humans.
The effects of physical exercise on hippocampal formation might be related to the fact that this treatment only improved ORM in the 24-h test, but not in the 3-h retention test, because the specific training procedures used favored a higher involvement of the hippocampus in the second retention test. Thus, it seems that this structure (a region highly affected by physical exercise) plays a more crucial role in ORM under certain circumstances, such as when there is a long delay between sample and testing, or when spatial components are emphasized. 22,23
Clarke and coworkers 47 found that plastic modifications in the hippocampus were crucial for ORM when in the sample phase two different objects were used, one of which was replaced in the retention test, probably because in this circumstance the animal has to make more spatial or relational associations. 22 The first retention test of the present work could be considered as a new sample phase with similar characteristics (two different objects were used, one of them being presented again in the second retention test, but in a different position). Both the training-testing delay and the specific procedures used may thus have made the contribution of the hippocampus more important in the second retention test.
The finding by Piao and associates, 14 however, that physical exercise initiated 1 month, but not 1 week, after TBI improved short-term ORM (1-h) in mice, also points to the possibility that the effects of physical exercise on short-term vs. long-term ORM tests may vary depending on timing of exercise initiation.
Citicoline improved memory in animals with TBI, but the mechanisms mediating this effect have not been clearly elucidated
Citicoline improved 3-h and 24-h ORM after TBI, although it seemed to be more effective in the first retention test, because while the discrimination index of citicoline-treated animals was higher than that of untreated animals with TBI at the two times tested, this index was only significantly higher than zero in the 3-h retention test. Citicoline has been found to improve spatial memory after brain ischemia 48 and CCI 20 in rats, but the present one is, to our knowledge, the first report of citicoline benefits on ORM after an experimental model of TBI. In contrast, mixed results have been obtained in human patients with TBI. 21
The results found in the present article give only a few hints about the plausible mechanisms that may have mediated the cognitive enhancing action of this compound. Thus, the interhemispheric ratio of hippocampal volume did not show significant tissue loss, suggesting a certain degree of neuroprotection in sedentary rats treated with citicoline. These effects are consistent with findings of other works with animal models of TBI that have found that citicoline attenuates brain edema and blood-brain-barrier breakdown, 49,50 reduces myelin loss, 50 and increases the number of CA1, CA2, and CA3 surviving neurons. 50,19
These neuroprotective effects may be mediated, in part, by a reduction of oxidative stress and apoptosis-related mechanisms. 50 We have not found, however, an attenuated loss of NeuN+ cell density in either the HDG or the PRHc of the ipsilateral hemisphere, or increased cell proliferation and neurogenesis in the DG. The possibility that citicoline may have exerted neuroprotective effects in other brain areas, including some hippocampal regions not examined here, cannot be disregarded.
While increased cell proliferation and neurogenesis in the DG 37 and in the perilesional areas 37,38 has been found in rat models of stroke after citicoline administration, there are not previous reports examining the neurogenic effects of this compound after TBI. Because the timing of BrdU injections was designed to label new cells born during physical exercise, the present results do not allow determination of whether there may have been enhanced neurogenesis close to the timing of citicoline administration.
On the other hand, discrepancies among works studying the efficacy of this drug after TBI may be attributed to differences in the doses used, the time of treatment initiation after injury, the route of administration, the duration of the treatment, etc, because optimal parameters of citicoline administration have yet to be clearly settled. 19
Combination of citicoline and physical exercise may be detrimental under certain circumstances
Contrary to our expectations, the effects of physical exercise and citicoline on memory and on histological parameters did not sum up. Further, while physical exercise did not hamper the effect of citicoline in the first retention test, there was nevertheless a negative interaction between physical exercise and citicoline in the second retention test, because animals that underwent combined treatment failed to demonstrate any recall and had a performance similar to that of untreated TBI animals in this test. This suggests that the combination of citicoline and physical exercise may be detrimental under certain circumstances.
A negative interaction between both treatments was also found in some histological parameters. Thus, in contrast to what was found in animals treated with physical exercise alone, no attenuation of TBI-related hippocampal volume loss was evidenced in animals that received both treatments. Moreover, cell proliferation and neurogenesis in the DG of animals with the combined regimen did not statistically differ from either those of the animals treated with exercise alone, or from sham rats, suggesting that the combined treatment attenuated the enhancing effect of physical exercise on neurogenesis. The combined treatment, however, did not prevent the attenuated expansion of the lateral ventricle or the reduction of interhemispheric differences in NeuN+ cells densities in the PRHc (and to a lesser extent, in the HDG) found in animals treated with exercise alone.
The negative interaction of citicoline and physical exercise in the 24-h memory test and in several histological parameters is intriguing. The two treatments were not given simultaneously, but sequentially. Specifically, citicoline was administered early after TBI, while physical exercise was initiated after the last citicoline injection because initiation of physical exercise too early after TBI may be detrimental. 8
In contrast to the present results, there are reports of additive positive effects of citicoline when given in combination with other neuroprotective agents for the treatment of brain ischemia, 51 as well as of exercise when given with a supplemented diet. 52 There are other works, however, showing no additive 53,54 or even negative 55,56 effects when combining different pharmacological or nonpharmacological treatments. Interestingly, negative interferences between physical exercise and amphetamine have also been reported. 57
These results highlight the need to publish both positive and negative data from animal studies, as well the importance of pre-clinical tests combining different treatments that have beneficial effects when administered separately. 58 The latter issue is of special importance in the case of those treatments that have few or any negative side effects such as physical exercise and citicoline, because the probability of being used together for human therapeutic purpose without previous clinical trial is not negligible.
With regard to citicoline and physical exercise, the possibility is suggested that some of the metabolic changes initiated by the former (for example, phospholipid synthesis) might have recruited metabolic resources needed for physical exercise to exert its beneficial effects. If this were the case, perhaps a longer delay between the end of citicoline administration and the onset of physical exercise (and thus, also, a longer delay of exercise initiation after TBI) could be associated with additive benefits of both treatments.
Conclusions
Citicoline and physical exercise exerted benefits on object recognition memory after TBI when given alone, but combined treatment led to some negative interference. Given the promising profiles of both treatments as therapeutic tools for the treatment of TBI-related deficits, further works need to be performed to study whether other schedules of combined treatments could be beneficial by acting through additive and/or complementary mechanisms.
Undoubtedly, significant progress in the treatment of TBI-related deficits would ensue if it were possible to potentiate the benefits of a pharmacological agent, such as citicoline, by combining it with a long-term treatment that can be easily incorporated into the daily routines of patients, such as physical exercise. In the meantime, the present data suggest that the combined treatment with citicoline and physical exercise should be avoided in patients after TBI.
Footnotes
Acknowledgments
A.J.S. was a recipient of a pre-doctoral fellowship from the Ministerio de Educación y Ciencia (BES 2008_007827).
This work was supported by the Ministerio de Educación y Ciencia (SEJ2006-14226-PSIC) and by the Ministerio de Ciencia y Tecnología (PSI 2009-08034).
We thank Ferrer Internacional, S.A. for kindly donating citicoline.
Author Disclosure Statement
No competing financial interests exist.