
Abstract
The availability of markers able to provide insight into protein changes in the central nervous system after fatal traumatic brain injury (TBI) is limited. The present study reports on the semi-quantitative assessments of the immunopositive neuroglial cells (both astrocytes and oligodendrocytes) and neurons for S100 protein (S100), as well as neuronal specific enolase (NSE), in the cerebral cortex, hippocampus, and cerebellum with regard to survival time and cause of death. Brain tissues of 47 autopsy cases with TBI (survival times ranged between several minutes and 34 d) and 10 age- and gender-matched controls (natural deaths) were examined. TBI cases were grouped according to their survival time in acute death after brain injury (ABI, n=25), subacute death after brain injury (SBI, n=18) and delayed death after brain injury (DBI, n=4). There were no significant changes in the percentages of S100-stained astrocytes between TBI and control cases. The percentages of S100-positive oligodendrocytes in the pericontusional zone (PCZ) in cases with SBI were significantly lower than in controls (p<0.05) and in the ABI group (p<0.05). In the hippocampus, S100-positive oligodendrocytes were significantly lower in cases with ABI and SBI (both, p<0.05), compared with controls. It is of particular interest that there were also S100-positive neurons in the PCZ and hippocampus in TBI cases after more than 2 h survival but not in ABI cases or controls. The percentages of NSE-positive neurons in the hippocampus were likewise significantly lower in cases with ABI, compared with controls (p<0.05) but increased in cases with SBI in PCZ (p<0.05). In conclusion, the present findings emphasize that S100 and NSE-immunopositivity might be useful for detecting the cause and process of death due to TBI. Further, S100-positivity in neurons may be helpful to estimate the survival time of fatal injuries in legal medicine.
Introduction
T
The molecular mechanisms after TBI still are not fully understood. The brain undergoes profound remodeling in an attempt to restore the most important functions after TBI. The injury causes a local restricted neuronal cell death, as well as glial activation and related neurotoxicity that contribute to subsequent neurological dysfunction. 8 Afterwards, complex interactions between glial cells and neurons in a cascade of secondary events in TBI start to happen. 5,7,9 These processes require a complex interaction between damaged neurons releasing alerting signals and responding glial cells. 10 Further, remodeling and restorative brain plasticity processes begin, such as angiogenesis, axonal remodeling, neurogenesis and synaptogenesis. 11,12
To refine already established methods in cases of unknown survival times, we examined brain tissue samples from autopsy cases with lethal TBI and other causes of death immunohistochemically for S100 protein (S100) and neuronal specific enolase (NSE), and via immunofluorescence for S100.
S100 is a multigenic family of non-ubiquitous Ca2+-modulated proteins expressed in vertebrates exclusively and implicated in intracellular and extracellular regulatory activities, modulating the activity of effector proteins or cells. 13 Calcium-ions as universal intracellular second messengers play a regulatory role in processes such as conduction and transmission of nerve impulses, cell motility, cell growth and differentiation, gene expression, communication between various enzyme systems, apoptosis, and necrosis. 14 S100 constitutes one of the various protein families that act to regulate cytosolic calcium levels or transduce the intracellular calcium signal. S100 is expressed in cytoplasm, in cell and nuclear membrane of brain tissue, and in different peripheral tissues, including cardiomyocytes (especially S100A1), adipocytes, chrondrocytes, and in certain neuroendocrine neoplasms, although the concentrations found in the central nervous system (CNS) are much higher than those found elsewhere. 13,15
Among S100 protein family members, S100B is primarily expressed in and secreted by astrocytes and other glial cells (e.g., oligodendrocytes) within the CNS in high concentrations, whereas S100B reflects about 95 % of the total S100 pool. 16 –19 Hence, S100 has been considered a glial marker protein, and it also is detected in numerous other cell types in the CNS, such as ependymal cells, the choroid plexus epithelium, and a few neurons. 17 Early studies have shown that S100 exerts neurotrophic effects in nanomolar concentrations on neurons, stimulating neurite outgrowth and regeneration and enhancing survival of neurons during development and after injury. 16 Nevertheless, higher concentrations result in opposite effects, such as cell death by necrosis and apoptosis. 20 These studies suggest that S100 is involved in processes underlying the differentiation of several cell types in the human brain. S100 already has been investigated immunohistochemically in autopsy cases with regard to the cause of death, the severity of TBI and the survival times. 21,22
As a major form of the enolases, NSE catalyzes the interconversion of 2-phosphoglycerate to phosphoenolpyruvate in the glycolytic pathway. It is highly enriched in cytoplasm of mature neurons and in cells of neuronal origin but also can be found in thrombocytes and erythrocytes. 23,24 The enzyme is localized mainly in nerve cell bodies and axons 25 ; it is not found, however, in any glial cells. 26 NSE immunostaining has been studied previously in comparison to necrotic changes and causes of death. In these cases NSE-positive neurons indicate vital reaction after TBI. 27,28 Also, NSE has been defined as an effective marker of axonal injury within the CNS in its early stages 29 and of brain stress in general. 30
Both S100 and NSE are clinical biomarkers used in patients with TBI. S100 and NSE levels in cerebrospinal fluid and serum can be used more or less satisfactorily to predicate the existence, absence and severity of CNS injury or brain death and the outcome of TBI patients. 31 –38 Additionally, even postmortem cerebrospinal fluid levels of both proteins S100 and NSE can be used as specific markers of assessing the severity of brain damage due to a lethal TBI. 39 Hence, both proteins–S100 and NSE–were chosen for the presented forensic study.
To our best knowledge, no combined immunohistochemical studies are available for S100 and NSE. The purpose of the present study was therefore to determine whether the percentage of S100 and NSE positivity in neuroglial cells like astrocytes and oligodendrocytes, as well as in neurons of different brain regions, were associated with the cause of death and the survival time after TBI.
Methods
Sample collection
In 47 autopsy cases with TBI and known survival times between several minutes and 34 days, samples of fresh brain cortex tissue with microscopically verifiable contusion signs were submerged in neutral buffered 4% formalin for fixation. The locations of these cortical contusions were frontal (n=25), temporal (n=14), parietal (n=4), and occipital (n=4). A sample from the hippocampus (ipsilateral to the contusion) in 35 of these cases and from the cerebellum (ipsilateral to the contusion) in 31 cases also were taken in order to gather information on the time course of generalized secondary changes in the CNS, such as hypoxia/ischemia, located outside of the contusion zone. Further, hippocampus samples were taken, as this anatomical region is extensively studied and due to the fact that previous studies showed significant changes in the immunopositivity of brain cells for S100 and NSE after TBI or necrotic changes. 22,27 The cognitive outcome of TBI survivors also depends on the extent of molecular alterations in hippocampus. In addition, samples of cerebellum were taken because of their protected location beyond the tentorium cerebellum in cases of TBI and promising results in our own recently-published study. 40 The ages of the individuals at the time of their death ranged from 18 to 85 years (average age, 46.2 years). There were 31 males (66.0 %) and 16 females (34.0 %).
According to the given survival times, the cases with TBI were divided into three subgroups comparable to related studies 2,39,40 : acute death after brain injury (ABI; survival time <2 h; n=25); subacute death after brain injury (SBI; survival time 2 h – 4 d; n=18); and delayed death after brain injury (DBI; survival time >4 d; n=4). For the control group, brain tissue was taken from the frontal cortex, the hippocampus and the cerebellum of ten autopsy cases without head trauma. Basic characteristics of all cases are given in Table 1.
m, men; w, women; TBI, traumatic brain injury.
The postmortem interval (PMI; i.e., the time interval between estimated time point of death and tissue sampling during autopsy) of TBI and control cases ranged from 5 h to 6 d (average PMI, 2.7 d) with continuously adequate cooling (approximately 4°C).
Histological and immunohistochemical methods
After a maximum of 24-h fixation, the brain tissue samples were embedded in paraffin. Serial sections with a thickness of 2 μm were produced since this has been shown to yield the best staining results. The sections were deparaffinized in xylene and rehydrated. The tissue sections were also stained using hematoxylin and eosin to evaluate morphology and possible traumatic changes.
Immunohistochemical staining was performed using the indirect polymer method (Dako REAL™ EnVision™ Detection System, DakoCytomation, Glostrup, Denmark) as described elsewhere. 41 The formerly used avidin-biotin-method was avoided because of the ubiquitous occurrence of biotin and the effect of this substance on background staining in brain tissue. Further, other studies have confirmed that our method leads to higher sensitivity and greater consistency of results. 41
The antibodies required pretreatment by microwave heating at 70°C (S100) and at 43°C (NSE) for 20 min, in citrate buffer, pH 6.0. To block the endogenous peroxidase, the sections were immersed for 10 min in peroxidase blocking solution (DakoCytomation).
The primary antibodies were polyclonal rabbit anti-S100 (S100 isolated from cow brain; DakoCytomation; reaction with human S100B strongly, S100A1 weakly, and S100A6 very weakly, no relevant reaction with other tested S100 protein members S100A2, S100A3 and S100A4) and monoclonal mouse anti-NSE (γγ-enolase purified from human brain, labels human NSE, DakoCytomation). They were applied at a concentration of 333 μg/L (S100) and 524 μg/L (NSE) for 30 min at room temperature. The object plates were locked into the Thermo Shandon Coverplate™ system over the entire period of immunohistochemical staining. A Dextran coupled with peroxidase and secondary antibody molecules (DakoCytomation) was then used before 3,3‘-diaminobenzidine in hydrogen peroxide/phosphate-buffered saline served as chromogen. Counterstaining was performed with hematoxylin (Mayer‘s).
Histomorphological evaluation and semi-quantitative analysis
Evaluation was conducted microscopically using the Carl Zeiss Axiolab optical microscope (magnification, 50 ×−400 ×). The total number of neurons and neuroglial cells, as well as the numbers of neurons and neuroglial cells in which immunoreactivity was detected, were respectively counted in ten high power fields (400×magnification) in two distant sections measuring at least 30 μm. Neuroglial cells were characterized as astrocytes and oligodendrocytes. Cytological classification of cells as astrocytes, oligodendrocytes, or neurons was performed according to established cytomorphological criteria. This especially applies to the morphology of the nucleus, the location of the nucleolus in the nucleus, and the spatial proximity of oligodendrocytes to neurons (satellitosis). The percent positivity was estimated as (number of positive cell type/total number of cell type)×100.
True to the idea of establishing basic knowledge for a routine use of S100 and NSE, we used routine samples of TBI cases with their typical disadvantages of forensic autopsy material. In order to ensure the highest possible reliability of our method, we only counted cells that could be allocated most certainly to one cell type.
We evaluated the changes in astrocytes in layer I and the white matter of the pericontusional zone in the cortex (PCZ, near to the histologically-visible injured area of the contusion zone), in the stratum pyramidale and radiatum of the CA3 region of the hippocampus, and in the stratum moleculare, as well as in the medullary layer of the cerebellum. Layer I was observed because of the important role of astrocytes in the blood–brain barrier in forming the glia limitans. The CA3 region was chosen for its vulnerability of neurons against experimental traumatic injuries in rats, 42,43 which is most likely related to the mechanical heterogeneity of hippocampal areas. 44
We evaluated the changes in oligodendrocytes in layers III-V, the white matter of the PCZ, and in the same regions of hippocampus and cerebellum, as we did with astrocytes. For the evaluation of the changes in neurons, the layers III-V of the PCZ, the stratum pyramidale of the CA3 region of the hippocampus, as well as cerebellar Purkinje cells, were observed. Layers III-V were observed for neurons and oligodendrocytes because of reported morphological changes in neurons after cortical compression in rats. 45
To compare changes in the PCZ of TBI cases with the control group, the given specifications for PCZ were applied to the frontal cortex samples of the control group. See Figure 1 for visualization for the location of countings. Exemplarily examples of in this study evaluated cases are shown in Figure 2.

Samples of hematoxylin-eosin stained brain tissue sections.
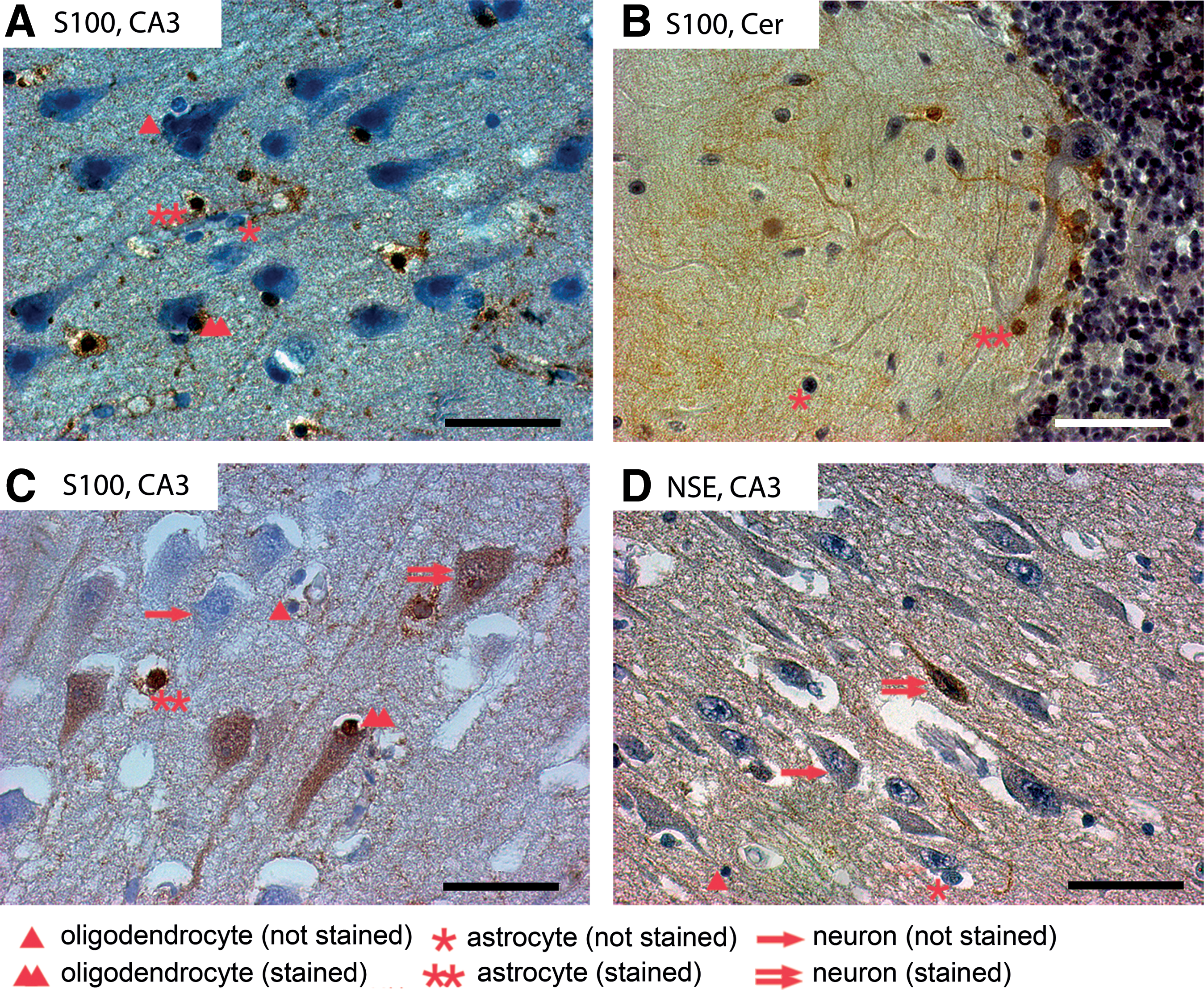
Immunohistochemically stained sections of different traumatic brain injury cases. Every cell type is highlighted only once per picture. CA3, hippocampal CA3 region; Cer, cerebellum.
Immunofluorescence methods
For immunofluorescence labeling using paraffin-embedded tissue sections, the treatment procedure was performed as previously described. 11 In brief, after the sections were deparaffinized, they were treated with citrate buffer (DakoCytomation) followed by microwave heating and washing in 0.1 M Tris buffered saline. After the pre-incubation with 5 % fetal calf serum in Tris buffered saline containing 0.1 % Triton X-100 for 30 min, the slices were incubated with a mixture of rabbit anti-S100 (1:400; DakoCytomation) and mouse anti-MAP2 (microtubule-associated protein 2, 1:200; Merck Millipore, Billerica, MA) in 5 % fetal calf serum in Tris buffered saline containing 0.3 % Triton X-100 overnight at 4°C. The visualization of the primary antibodies was performed with Cy2-conjugated donkey anti-rabbit (1:400) and Cy5-conjugated donkey anti-mouse immunoglobulin G (1:200; Jackson ImmunoResearch, West Grove, PA), incubation for 2 h at room temperature. In addition, slices were stained with the nucleic acid probe Hoechst 33342 (1:1000, Molecular Probes, Leiden, The Netherlands) with auto-fluorescence under ultraviolet light to identify the cell nuclei.
After intensive washing, all stained sections were dehydrated in a series of graded ethanol, processed through n-butylacetate, and covered with Entellan (Merck, Darmstadt, Germany). Critical control experiments were carried out without primary antibodies.
Confocal microscopy
The immunofluorescence was investigated by a confocal laser scanning microscope (LSM 510 Meta, Zeiss, Oberkochen, Germany) using excitation wavelengths of 488 nm (argon laser, yellow-green Cy2-immunofluorescence), 543 nm (helium/neon1, red Cy3-immunofluorescence) and 633 nm (helium/neon2, Cy5-labelling, color-coded in blue), whereas an ultraviolet laser (362 nm) was used to evoke the blue-cyan Hoechst 33342 auto-fluorescence.
Statistical analysis
Statistical analysis was performed using SPSS Statistics (version 21, 2012 IBM, Armonk, NY) and OpenOffice Calc (Version 3.4.1, 2012 Apache Software Foundation, Forest Hill, MD). Correlations between immunopositivity percentages and gender, PMI, and among percentages of cell positivity in different locations and for different cell types were computed using Spearman correlation. The correlation coefficient (rs), the significance (p) and the number of used cases (n) is specified for each correlation.
The groups of survival time were tested for statistically significant differences in immunopositivity percentages using the Mann-Whitney-Wilcoxon-Test (non-parametric). For each comparison, there is the difference of percentages points (%p), the significance (p) and the number of used cases (n).
Resulting p values of both methods were checked for multiple testing applying the Explorative Simes-Procedure (Benjamini-Hochberg-Procedure). Only results withstanding this procedure were listed: p values of less than 0.05 were considered statistically significant.
Results
No correlation between the positivity of the cells and PMI (rs =−0.27 to 0.15; p=0.1 to 0.96) or gender (p=0.07 to 0.98) could be demonstrated. Due to an often delayed death of older individuals and hence a significant positive correlation between survival times and age of the individuals (rs=0,33, p<0.05), no meaningful correlation between the age of the individuals and the positivity of brain cells could be performed.
There were no significant differences between the locations of the PCZ in the cortex concerning positivity of neurons and neuroglial cells (p=0.07 to 0.97). No significant differences were observed in the results while combining percentages of neuroglial cells of the gray and white matter in cortex samples, in cases of the CA3 region of hippocampus, stratum pyramidale and stratum radiatum, or in cases of the cerebellum, stratum moleculare, and medulla. In the following evaluation we therefore only refer to the region PCZ, CA3 or cerebellum. For general comparison and quality assurance, the numbers of total counted cells over all categories of survival times were registered and are shown in Table 2. There were no significant differences between the three different survival time groups in cases of TBI. However, the total cell count of neurons in the PCZ and the hippocampal CA3 region of TBI cases was lower than in the control groups.
Standard error is shown via±. All numbers were rounded off to whole numbers.
TBI, traumatic brain injury.
Differences between groups of survival time
Astrocytes show the known morphological characteristics and intensive immunopositive staining with anti-S100 antibody (for visual illustration see Fig. 2A, 2B, 2C). S100 positivity decreases in the group of SBI, compared with the ABI and control groups. It seems to regain control group levels in the group of DBI. There were no significant changes in the percentages of S100-stained astrocytes in the PCZ or hippocampal CA3 region between cases with TBI and control cases (data not shown). Also, no significant differences in the cell populations of neuroglial cells within the cerebellum were observed that could withstand the Explorative Simes Procedure.
In general, the total numbers of S100-positive oligodendrocytes decreases in all three investigated survival-time categories after TBI, compared with controls. The percentages of S100-stained oligodendrocytes in the PCZ (Fig. 2A, 2C) were significantly lower in cases with SBI than in cases with ABI (18.2%p; p<0.05; n=37), as well as in the control group (26.7%p; p<0.05; n=25). These percentages increased in cases with DBI. In the CA3 region of the hippocampus, the percentages of S100-stained oligodendrocytes were lower in cases with ABI (13.6%p; p<0.05; n=31) and in cases with SBI (24.7%p; p<0.05; n=20) than in the control group (Fig. 3).
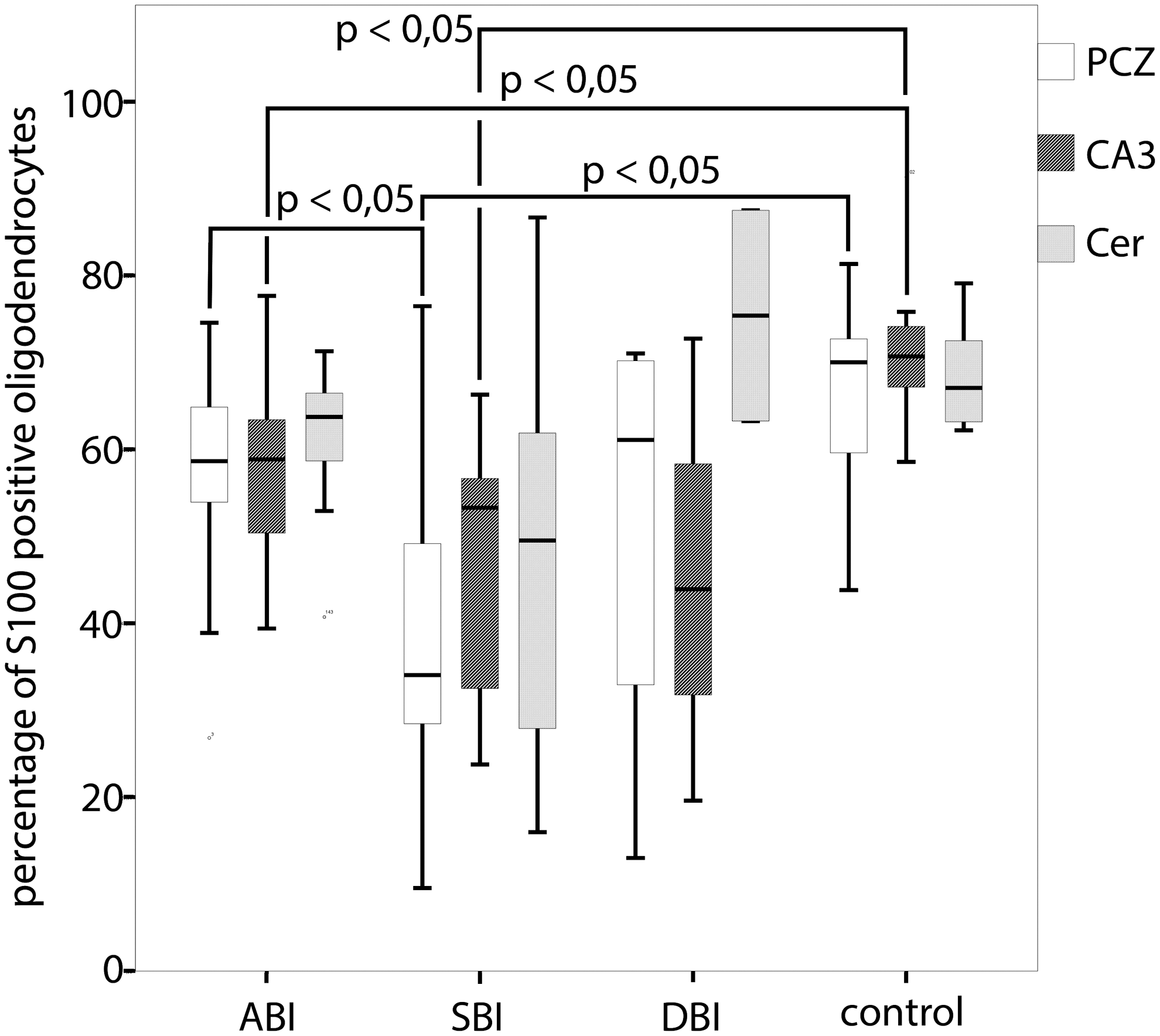
The percentages of S100-positive stained oligodendrocytes in the pericontusional zone, the hippocampal CA3 region and in the cerebellum in relation to categories of survival time. Statistical analysis by the Mann-Whitney-Wilcoxon-Test, checked for multiple testing by the Explorative Simes-Procedure. PCZ, pericontusional zone; CA3, hippocampal CA3 region; Cer, cerebellum; ABI, acute death after brain injury (≤2 h); SBI, subacute death after brain injury (2 h – 4 d); DBI, delayed death after brain injury (>4 d).
The percentages of NSE-stained neurons in the CA3 region of the hippocampus (Fig. 2D) were higher in cases with SBI than in cases with ABI (15.8%p; p<0.05; n=32), though significantly lower in cases with ABI than in the control group both in the PCZ (20.4%p; p<0.05; n=33) and in the CA3 region (17.8%p; p<0.05; n=31). Following the clear decrease in NSE-positive neurons after TBI in the cases of ABI, NSE-positive neurons start to increase over the categories of SBI and DBI in both regions, where the rates of NSE-positive neurons in the PCZ are generally higher than in the hippocampal CA3 region. Interestingly, the neurons of the PCZ and the CA3 region of the hippocampus seem to recover from their loss of NSE-positivity and reach a comparable level with the control groups in time interval of SBI (Fig. 4). In none of the cases were there any NSE-positive Purkinje cells in the cerebellum.
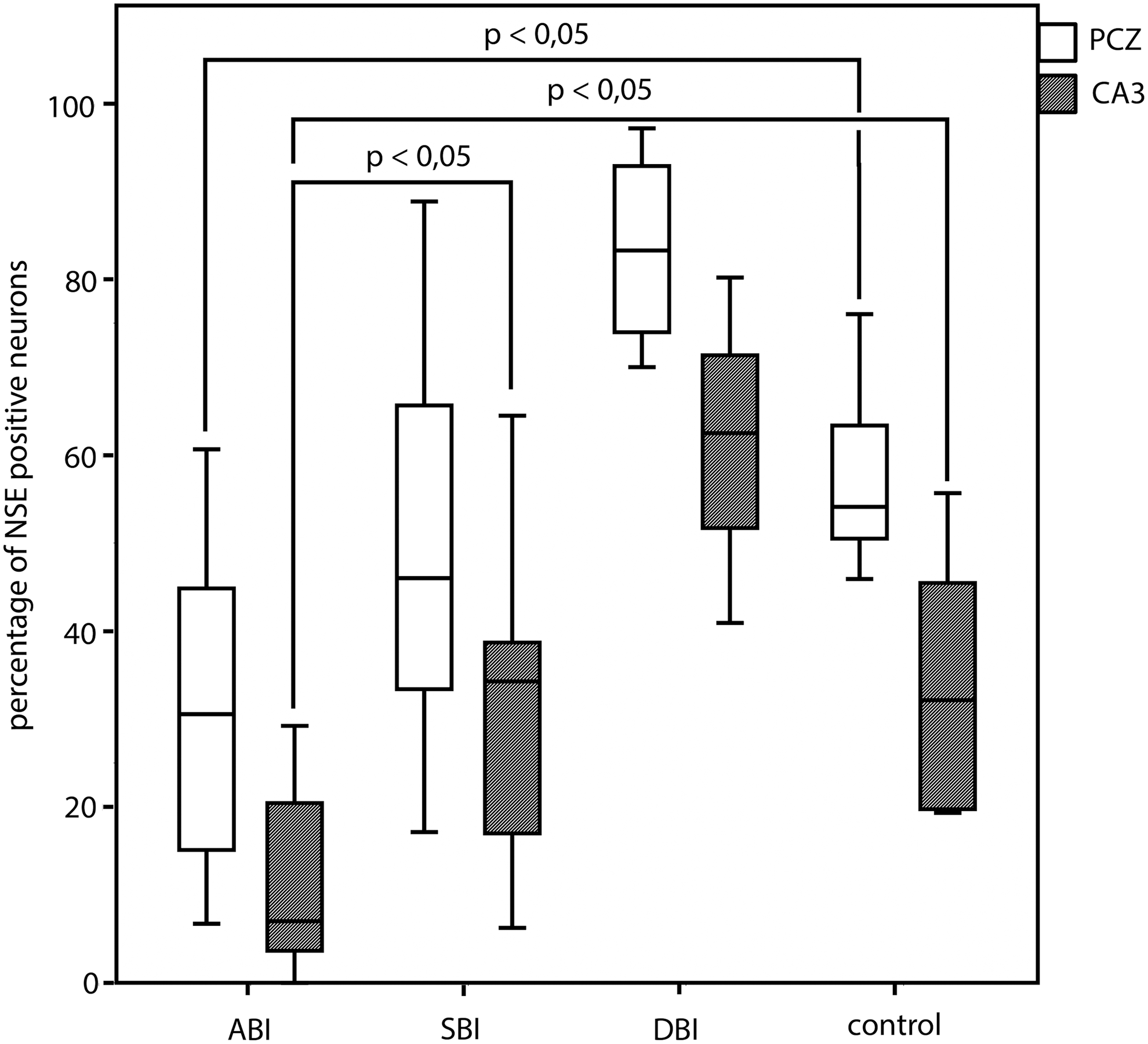
The percentages of neuronal specific enolase (NSE)-positive stained neurons in the pericontusional zone and the hippocampal CA3 region in relation to categories of survival time. Statistical analysis by the Mann-Whitney-Wilcoxon-Test, checked for multiple testing by the Explorative Simes-Procedure. PCZ, pericontusional zone; CA3, hippocampal CA3 region; ABI, acute death after brain injury (≤2 h); SBI, subacute death after brain injury (2 h – 4 d); DBI, delayed death after brain injury (>4 d). Data for cerebellar region are not shown because none of the Purkinje cells reacted with anti-NSE.
Surprisingly, there also were S100-positive neurons in cases with SBI and DBI of the PCZ and the CA3 region (Fig. 2C). In most cases, a diffuse neuroplasmatic staining of the neurons was found in the hippocampal area, as in the PCZ the neurons were stained in a more juxtanuclear way. For both regions, we detected an unsystematic distribution of S100-positive neurons in spatial proximity to S100-positive glial cells. No S100-positive neurons were found in the cases with ABI or in controls (Fig. 5). In every case in which positive S100-stained neurons in the PCZ could be found, they also could be seen in the CA3 region of the hippocampus.
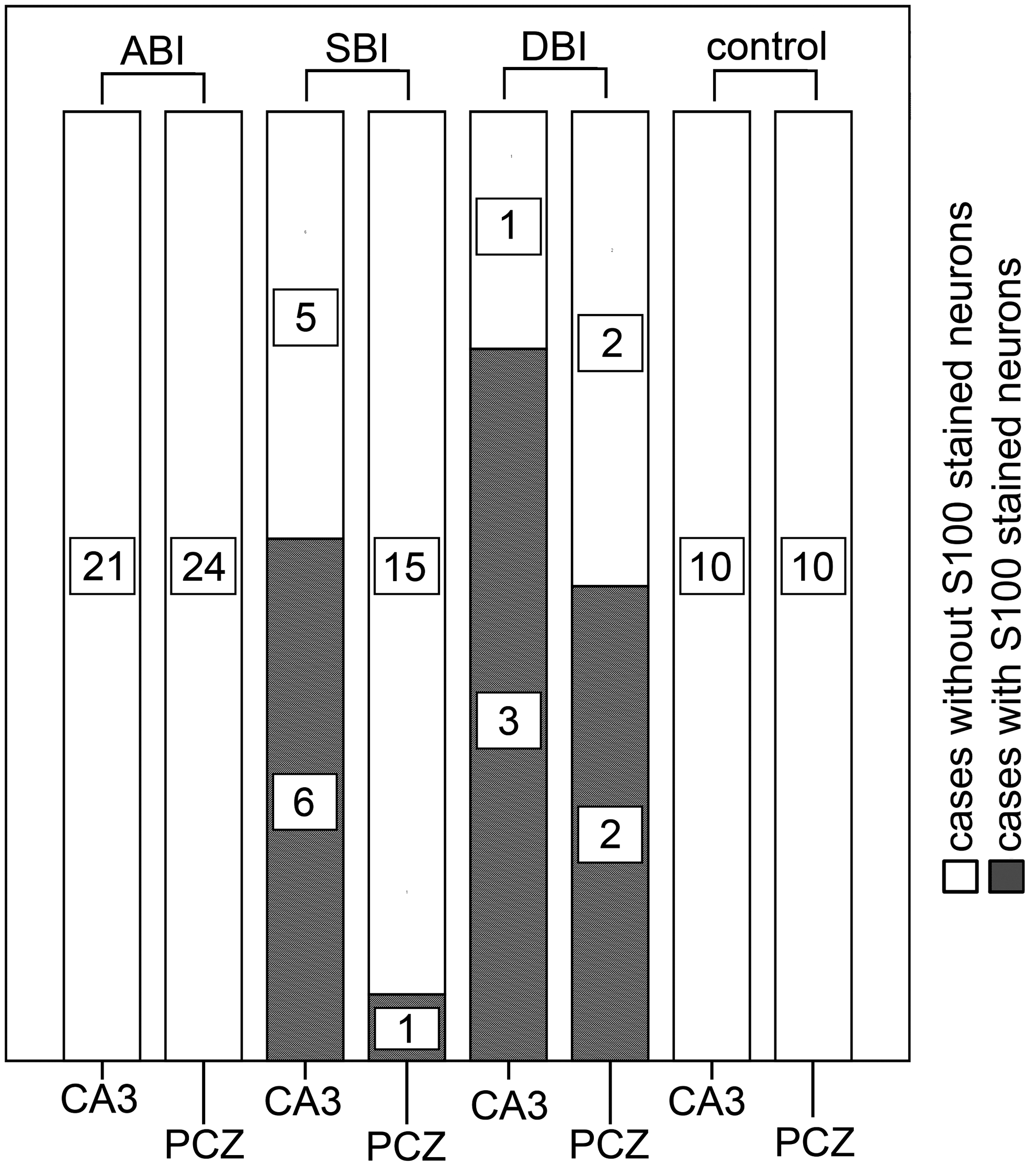
Cases with S100-positive stained neurons in the pericontusional zone and the hippocampal CA3 region in relation to the categories of survival time. PCZ, pericontusional zone; CA3, hippocampal CA3 region; ABI, acute death after brain injury (≤2 h); SBI, subacute death after brain injury (2 h – 4 d); DBI, delayed death after brain injury (>4 d).
To confirm these results and to rule out staining artefacts, immunofluorescence methods were performed using confocal microscopy, where we approved suspected cells to be of neuronal origin by MAP2-staining, as was done previously. 11 During this procedure, additional aspects were detected: S100-positive neurons were found in MAP2-positive, as well as in MAP2-negative neurons. Neuronal cells, whose morphology is typical for both triangular and bipolar architecture, are stained by S100 in a similar intensity. S100-positivity in neurons is not necessarily associated with apoptotic or aging processes of neuronal cells (Lipofuscin-positive). Immunofluorescence results are presented in Figure 6.
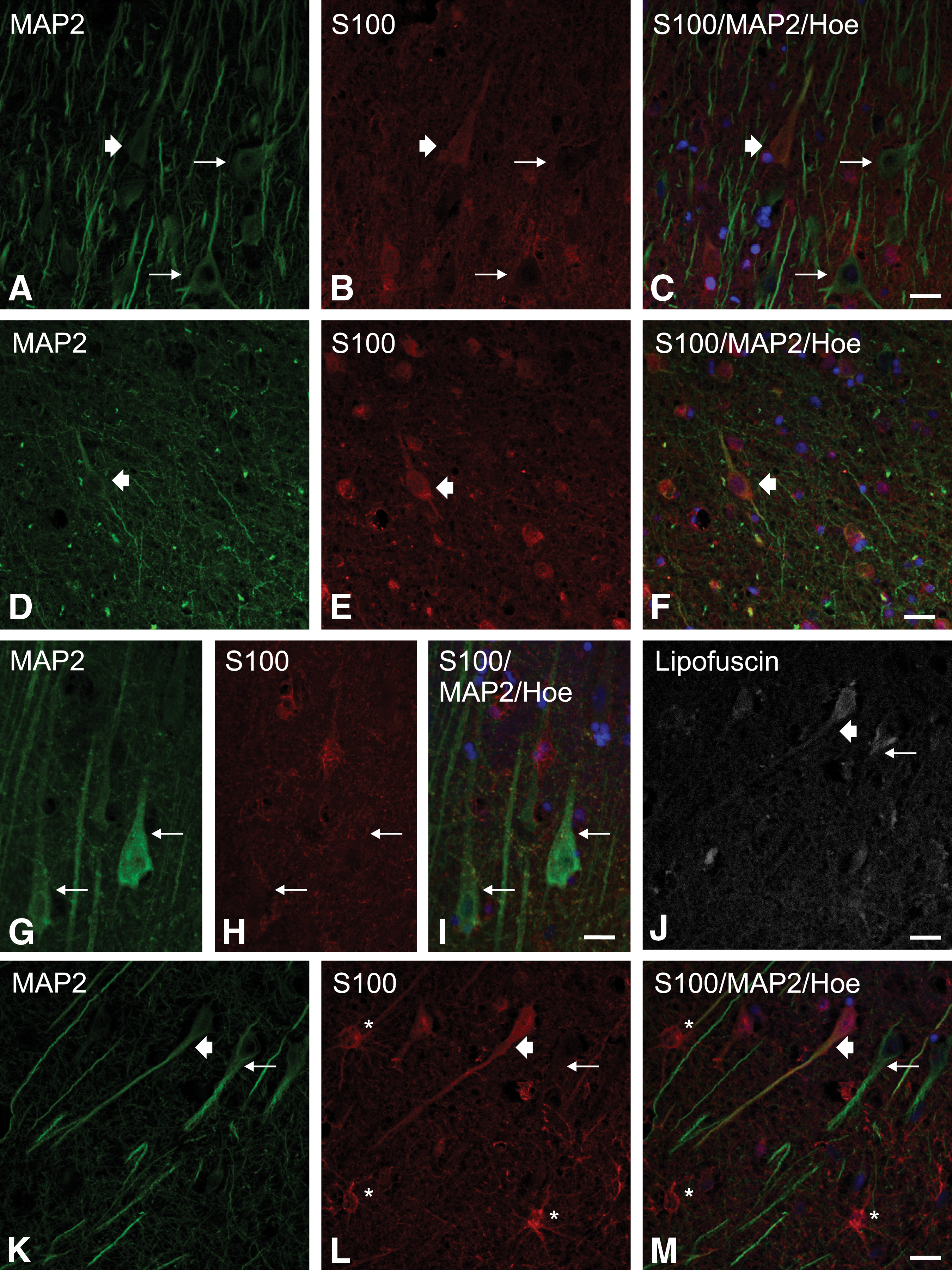
S100-positive stained neurons in the hippocampal CA3 region using immunofluorescence methods with confocal microscopy.
Correlations between individual cell percentages
A positive correlation could be found in the percentages of S100-stained glial cells (astrocytes and oligodendrocytes) comparing their positivity in the gray and white matter of PCZ and cerebellum and in case of the hippocampal CA3 region in the stratum pyramidale and the stratum radiatum (Table 3.1). Also, we could demonstrate positive correlations between the percentages of positively stained glial cells in the PCZ and the CA3 region of the hippocampus (Table 3.2). By evaluating percentages of positively NSE–stained neurons, we were able to demonstrate a positive correlation between those in the PCZ and those neurons in the stratum pyramidale of the hippocampal CA3 region (Table 3.3).
rs, Spearman's rank correlation coefficient; PCZ, pericontusional zone; CA3, hippocampal CA3 region; NSE, neuronal specific enolase.
Discussion
The aim of this study was to establish an immunohistochemical method to determine the cause of death and the survival time after TBI using the proteins S100 and NSE. Main findings suggest that S100 and NSE immunopositivity in neuroglial cells and neurons in different brain regions were associated to the cause of death and the survival times after TBI.
We did find a positive correlation between positively S100-stained glial cells as well as positively NSE-stained neurons in the PCZ and within the hippocampus. This suggests that our observations are more part of a global process in the brain during secondary events after TBI than only of the injured area (contusion zone). PMI was shown to have no effect on cell positivity under the used conditions. Because PMI is an influential aspect for forensic work, the described method seems to be useful within the realm of forensic medicine.
Changes in the positively S100–stained glial cells in dependence of survival time
Contrary to findings of Li and colleagues, 21,22 where causes of death and survival times were connected to S100-stained astrocytes, we did not find any significant differences in their staining, compared with controls and different categories of survival time. Astrocytic swelling was observed even after survival times of only a few minutes, which could be interpreted as cytotoxic edema immediately after the traumatic impact, as was previously reported. 46 S100 seem to be specifically involved in this process by regulation of calcium fluxes and stimulation of astrocyte proliferation. 47 Also, percentages of stained astrocytes were much higher in our overall investigations than in cited works. 21,22 This could be related to the use of the polymer instead of the avidin-biotin-method and the higher number of cases used by Li and associates, especially in the group of DBI. Astrocytes play a vital and active role in the CNS, including maintenance of cellular homeostasis and reparative responses. 48
Besides that, neuroglial cells themselves are able to trigger the cell death among each other, which likely leads to the leakage of S100 from dying cells after traumatic stress but to the possibility of stimulation of glial cell proliferation. 11,49 It was shown that S100 is expressed in and released from oligodendrocytes in humans. 21,50 Interestingly, in our samples a significant difference in the staining of oligodendrocytes was found when compared with controls and survival times. The study by Li and colleagues 21 instead shows no significant change in the positivity of the oligodendrocytes. In addition to using the avidin-biotin instead of the polymer method, another possible reason for this difference could be the usage of slightly different categories survival time and different layers of the cortex in which they were counted.
Since no evident reduction of S100-positive neuroglial cells was detected in PCZ for cases of ABI, immediate lethal brain dysfunction due to the primary impact independent of any relevant hypoxic/ischemic brain damage could be suggested. The loss of S100-positivity in oligodendrocytes in cases of SBI could be related to an active release from traumatically damaged oligodendrocytes next to processes of remyelinisation after injury and could be caused by hypoxia and acidosis accompanied by brain edema. 51,52 Also, S100 was found to be at least partially responsible for the demyelinisation of oligodendrocytes and has been reported to cause apoptosis. 20,53,54 We assume that our finding of reduced oligodendroglial S100-immunostaining in cases of SBI is caused rather by an increased release of S100 than by reduced cellular S100 expression, because of the known increase in S100 levels in CSF and serum after TBI. 39
The observed trend of increasing rates of S100-positive oligodendrocytes in DBI cases might be due to counterregulatory regeneration effects. Interestingly, oligodendrocyte precursor cells reacquire features of multipotent precursors after injury, and might represent a source for new neurons or glia replacing damaged cells. 11 Recent studies show that the content of S100 in various brain areas is regulated by serotonin and therefore the loss of S100 can be reversed by clinical treatment with a 5-HT1A agonist. 55
Changes in the positively S100–stained neurons in dependence of survival time
In this study, the known glial cell marker S100 also stained neurons. The distribution of S100 proteins and their presence in neurons has been discussed in several animal studies before. 56,57 An early study in human brains showed that subpopulations of neurons (pyramidal cells) apparently contain S100A1 but not S100B in the hippocampus area. 58 It might be possible that S100 occurs in neurons only under certain conditions and that neurons might contain much less S100 than glial cells.
To our knowledge, in combination with TBI, this phenomenon only has been described by Li and colleagues. 21 While S100-positive neurons could only be observed in the cases with SBI and DBI of the stratum pyramidale of the CA3 region of hippocampus and the PCZ, one could assume that the survival time needs to be at least 2 h before S100 positive neurons can be detected in the hippocampus and even longer when S100 positive neurons are supposed to be seen in the PCZ. Those findings correlate with observations of Li and coworkers, 21 in which S100 positive neurons also were found in cases of head injuries with more than 6 h survival time.
On the one hand, subpopulations of neurons might have the possibility to synthetize S100 in pathological situations like TBI, as it was believed before for rats. 56 On the other hand, the S100-staining of neurons possibly could be explained by the role of neuroglial cells in repairing and neuroprotection processes after injury. 54,59 Additionally, after secretion partially via the surface receptor for advanced glycation end products, there is the extracellular, cytokine-like effect of S100, which plays an important role in the development and maintenance of neuronal functions. 60,61 S100 modulates the proliferation and differentiation of neurons, 62 has a neuroprotective function, 63,64 and stimulates dendrite and axonal growth. 65,66 After trauma, S100 can be released early and actively from neuroglial cells or can be leaked from damaged cells during traumatic or metabolic stress. 67 Some hippocampal glial cells may secrete lethal amounts of S100 for neighboring neurons without showing any peripheral signal that this is occurring. This assumption is backed up by our findings of lower percentages (16.7%p; n=45; did not withstand Explorative Simes Procedure) of positively S100-stained oligodendrocytes in the hippocampus in cases with S100-positive neurons.
Either the extracellular S100 binds to neuronal membrane structures because of its affinity with cytoskeleton components or it is taken up by neurons when used as tracer of axonal transport. 68 Maybe traumatic deletion of S100 exacerbates neuronal loss and tends to a dysregulation of the neuroprotective metallothioneines, a phenomenon that alters the neuronal metabolism. 69 The fact that the hippocampus is affected early after TBI is most likely a result of the higher vulnerability to hypoxia/ischemia of hippocampal neurons due to an early loss of supporting glial cells compared to the neurons of other brain regions. 3,70
Diffusion of S100 from glial cells into neurons during fixation has never been reported before and would not be compatible with numerous S100-negative neurons next to the S100-positive ones. A technical artifact in the staining pattern via immunohistochemistry was ruled out by the use of an additional immunofluorescent method with exact comparable results. That S100-positive but MAP2-negative neurons were detected might be because the used neuronal marker MAP2 does not stain all neurons consequently. 71 Our results suggest a reactive neuronal staining only, while none of the neurons were positive for S100 in control cases and ABI. Hence, the combination of our results proves that distinct amounts of S100 occur in some neurons after TBI. However, the exact molecular mechanisms remain to be identified. Published data regarding the long-term regeneration of damaged neurons and their staining pattern for S100-immunhistochemistry is generally lacking.
It is possible that the appearance of S100 positive neurons in the hippocampus and the PCZ could determine a minimum survival time. A more exact cut-off-point may be constructed using more postmortem cases with survival times between 2 h and several days.
To phrase it conservatively, our data may be interpreted as an introduction of S100 as a therapeutic target for neuroregeneration after trauma. This assumption is supported by experiments in rats in which S100 has been proven as a “therapeutic drug.” 72,73 Therefore, the immunohistochemical distribution of S100 should be considered when testing the effects of treatments after TBI. Those could be heparinization, 74 the use of arundic acid to suppress S100 expression in rats, 75 or the intraperitoneal application of S100 in rats. 76
Changes in the positivity of NSE-stained neurons in dependence of survival time
Considering the cellular specifity of this neuronal protein, the presented changes of NSE expression could be a result of a direct loss of NSE from the damaged neurons, as the known high cerebrospinal fluid and serum levels of NSE (in clinical trials as well as postmortem) after a TBI suggest. 29,39,77 These results are accompanied by similar findings from Nogami and colleagues, 27 where NSE staining in the CA3 region of the hippocampus decreased with an increasing histological necrosis index observed in different causes of death.
After reaching the lowest NSE-positive percentages in ABI cases, an increase starts in cases of SBI up to an extension in the DBI group beyond the measurements for control cases. Perhaps, these findings could be interpreted by neuroregeneration after TBI in combination with an excess expression of NSE to gain the deficiency of neuronal cell deaths.
The immunohistochemical investigation of cerebellar samples seems to be not useful to detect survival times and causes of death in forensic routine. The Purkinje cells of the cerebellum showed no immunoreactivity for NSE in TBI as well as control cases. This outcome is comparable to similar findings in humans as well as in rats, where no or only poor staining for NSE was found. 27,78,79 One could assume to observe changes in injured cerebellums: Four of our cases showed morphological signs for a distinct injury of the cerebellum within a contre-coup zone. However, the Purkinje cells of these samples also were not stained for NSE.
Conclusion
In conclusion, the observations suggest S100 and NSE in combination as a good forensic immunohistochemical method to estimate unknown survival times of TBI victims, especially in cases with S100-stained neurons in the PCZ or the CA3 region of hippocampus. Our findings can therefore be used to get an overview of the immunohistochemical changes in the investigated time intervals after fatal TBI and are useful for detecting the cause and process of death due to TBI.
Further combinations of this method with postmortem biochemistry, molecular pathology and genetics could lead to even more exact information and are already in progress.
Footnotes
Acknowledgments
We wish to thank Cornelia Pietschmann (Institute of Legal Medicine, Leipzig) and Katrin Becker (Rudolf Boehm Institute of Pharmacology and Toxicology, Leipzig) for their skillful technical assistances in laboratory for histology and immunofluorescence.
This study was approved by the local ethic commission of the Medical Faculty of the Leipzig University (AZ: 117-12-23012012). Following the approval of the ethical committee the public prosecutor's office, Leipzig University allowed us to analyze the brain tissue samples to determine the cause and manner of death according to the guidelines from the central ethic commission of the federal medical association.
Author Disclosure Statement
This work was partly supported by the German Ministry of Defense. The authors declare that no competing financial interests exist.