
Abstract
Cell therapy for spinal cord injury (SCI) is a promising strategy for clinical application. Mesenchymal stem cells (MSC) have demonstrated beneficial effects following transplantation in animal models of SCI. However, despite the immunoprivilege properties of the MSC, their survival in the injured spinal cord is reduced due to the detrimental milieu in the damaged tissue and immune rejection of the cells. The limited survival of the engrafted cells may determine the therapy success. Therefore, we compared two strategies to increase the presence of the cells in the injured spinal cord in rats: increasing the amount of MSC transplants and using immunosuppressive treatment with FK506 after transplantation. Functional outcomes for locomotion and electrophysiological responses were assessed. The grafted cells survival and the amount of cavity and spared tissue were studied. The findings indicate that immunosuppression improved grafted cells survival. A cell–dose effect was found regarding locomotion recovery and tissue protection independent of immunosuppression. Nevertheless, immunosuppression enhanced the electrophysiological outcomes and allowed filling of the cavity formed after injury by new regenerative tissue and axons. These results indicate that MSC transplantation combined with immunosuppression prolongs the survival of engrafted cells and improves functional and morphological outcomes after SCI.
Introduction
T
With regard to the feasibility of cell transplants, the possibility of autologous transplantation has been proposed as an important consideration for the translation to the clinical setting. However, there are some difficulties for the autologous application of these cells since the characteristics of the MSC, such as expansion growth rate, 14 cytokine expression profile and response to inflammatory stimulus, 15 as well as the capability to stimulate axonal growth and produce beneficial effects after SCI, 16 are subjected to donor variation. On the other hand, the time needed for the in vitro expansion of the cells make impossible the autologous transplant during the acute phase of the injury. Allogeneic cell transplantation would solve these issues, provided that host rejection is avoided by an accurate selection of the donor and immunosuppression for ensuring the success of the treatment.
It has been reported that MSC are able to modulate the immunological activation of immune cells in vitro, including the suppression of T-cell proliferation and the inhibition of dendritic cells differentiation. 17 –20 In addition, MSC are hypoimmunogenic due to their low expression of the major histocompatibility complex (MHC) class I and no expression of co-stimulatory molecules, 18 thus not inducing the proliferative response of lymphocytes. 17,18 Indeed, the possibility of universal donor MSC for therapeutic applications has been suggested. 19 Nevertheless, the immunomodulatory properties of MSC in vivo have not been fully addressed. Allogeneic MSC transplanted into the intact spinal cord of rats have short-term survival 8,21 that may be prolonged by immunosuppressive treatment, 21 indicating an immunogenic response against the grafted MSC. In addition, we have recently found an increased expression of immunogenic reaction associated genes a few days after allogenic MSC transplantation in the injured spinal cord. 22 Thus, despite the relative immunoprivileged properties of MSC, the injured spinal cord of the host appears to induce an immune rejection against the transplanted cells. This exacerbated immune response may explain the fast disappearance of the grafted allogeneic MSC reported after SCI in rats 6,8,9 and dogs, 23 as well as that of xenogenic human MSC in injured rats. 10
Therefore, the reduced survival of the grafted MSC after SCI seems to be an important factor limiting the success of the transplant to produce reparative effects. The aim of this work was to investigate if the amount of cells present in the injured spinal cord is important for the success of the transplant. Thus, two concentrations of allogeneic MSC were used—a low amount of cells (∼half a million), previously reported to provide some benefits in a similar SCI model, 8,13 and a higher number of MSC (∼1 million). 13,24,25 In addition, the two types of transplants were combined with immunosuppressive treatment to prolong the graft survival. For this aim, we selected tacrolimus (FK506), a FDA-approved immunosuppressant drug that in addition to efficiently preventing allograft rejection also exerts neuroprotective and pro-regenerative actions. 26 –28 In experimental animals, FK506 treatment consistently has been found to increase axonal regeneration after peripheral nerve injury, 29 –31 allowing the use of allogeneic cells for repairing long nerve defects. 32 After SCI in rats, FK506 exerts some protective effects against the secondary injury and enhances axonal sprouting. 33 –36
Methods
MSC culture and characterization
Primary cultures of MSC were set up from P22 male Sprague-Dawley rats. The animals were euthanized with CO2. From each animal, tibias and femurs were dissected, placed in cool phosphate buffered saline (PBS) and epiphyses removed. The diaphyses of bones were flushed with PBS using a syringe and the marrow was homogenized. The extract was filtered through a 70-μm nylon mesh and recovered by centrifugation for 10 min at 1500 rpm (220 g). The pellet was re-suspended in growth medium α-MEM with L-glutamine (Life Technologies, Grand Island, NY), supplemented with 20% heat-inactivated fetal bovine serum (FBS, Lonza, Verviers, Belgium), 2mM L-glutamine (Life Technologies) and 100 units/mL penicillin-streptomycin (Life Technologies, 100×), and plated in 100-mm culture dishes (Iwaki, Asahi Technoglass, Chiba, Japan) at a density of 5 · 106 cells/cm2. After 24 h, the supernatant containing non-adherent cells was removed and fresh medium was added. When the culture was near confluence, every 4–5 d, the cells were detached using PBS with 0.05% trypsin (Life Technologies) and 0.04% EDTA (Sigma, St. Louis, MO) and re-plated at 5,000 cells/cm2 8 . Cells were passaged three to four times, expanded to 80–90% of confluence.
For phenotype characterization, after four passages in culture, MSC were labeled for 30 min using antibodies against CD90, CD29, CD11b and CD45, conjugated with FITC (BD Bioscience, Erembodegem, Belgium; 1:200 v/v). After incubation, the cells were centrifuged for 5 min at 1300 rpm and re-suspended in PBS. Analysis of surface markers was performed using flow citometry (FACSCalibur, Becton Dickinson, Madrid, Spain) and FACSDiva software (BD Bioscience). The capacity of the MSC for differentiation to adipogenic and osteogenic cells was assessed as described previously. 8,37 For analysis of adipogenesis, cells were fixed for 20 min with 4% paraformaldehyde in phosphate buffer (PB); the adipogenic cells were labeled using 60% Oil red O stock solution (0.5% Oil red O in isopropanol, Sigma) for 15 min and washed with distilled water. For osteogenic labeling, cells were fixed using 70% ethanol pre-cooled for 1 h at 4°C, washed and incubated during 30 min with 0.1 mg/mL Alizarin red solution (Sigma) in distilled water.
For identification of the cells after grafting, MSC were transfected with a lentiviral vector encoding for the green fluorescence protein (GFP) under EF1α promoter. Cells in passage 2 were plated at 2000 cells/cm2 and incubated with lentiviruses during 48 h. Then, the medium was changed and the cells cultured as described above before transplantation. The infected cells were checked using an antibody against GFP (1:200, Abcam, Cambridge, United Kingdom). The efficacy of infection was measured by flow cytometry (FACSCalibur, Becton Dickinson) and FACSDiva software (BD Bioscience). We found that around 90% of the cells in the infected culture were positive for GFP.
Spinal cord injury and cell transplantation
Adult female Sprague-Dawley rats (9 weeks old; 250–300g) were used. The animals were housed with free access to food and water at a room temperature of 22±2°C under a 12:12 light-dark cycle. The experimental procedures were approved by the ethical committee of the Universitat Autònoma de Barcelona in accordance with the European Directive 86/609/EEC.
Under anesthesia with ketamine (90mg/kg) and xylacine (10 mg/kg) and aseptic conditions, a longitudinal dorsal incision was made to expose T6-T10 spinous processes. A laminectomy of T8-T9 vertebra was made and a cord contusion of 200 Kdyns was induced using an Infinite Horizon Impactor device (Precision System and Instrumentation, Kentucky, USA). The animals were transplanted acutely, 30 min after the contusion, and divided in eight groups. Four groups of rats received an injection in the spinal cord (see below) of a low dose of cell suspension or vehicle (VHC), with or without FK506 co-administration. Other four groups of rats were injected with a high dose of cell suspension or vehicle, with or without FK506 co-administration. In addition, for studying cell survival, four additional groups were transplanted with GFP+ MSC (gMSC). Table 1 shows all the experimental groups included in the study.
MSC, mesenchymal stem cells; i.p., intraperitoneal; VHC, vehicle; gMSC, green fluorescence protein+MSC.
Spinal cord injections were made using a glass needle (100 μm internal diameter, Eppendorf, Hamburg, Germany) coupled to a 10 μL Hamilton syringe (Hamilton #701, Hamilton Co, Reno, NV). Intraspinal injections were performed at the epicenter of the contusion and at 2 mm rostrally and caudally. In the low-dose groups, 3 μL of cell suspension (50,000 cells/μL) or vehicle (L15) were injected at each indicated point for a total of 450,000 cells per rat. In the high-dose groups, 4 μL of cell suspension (75,000 cells/μL) or vehicle (L15) were injected in both caudal and rostral injection points and 6 μL in the epicenter, for a total of 1,050,000 cells per rat. The cell concentration was increased in the high-dose groups in order to inject a volume only slightly larger than in the low-dose groups (total volume of 14 μL vs. 9 μL), and do not compromise the survival of cells through the injection process (viability >95% of the cells after re-suspension and injection through the needle). A perfusion speed of 2 μL/min was controlled by an automatic injector (KDS 310 Plus, Kd Scientific, Holliston, MA), and the tip was maintained inside the tissue 3 min after each injection to avoid liquid reflux.
The wound was sutured by planes and the animals allowed recovering in a warm environment. An intraperitoneal (i.p.) bolus of saline or FK506 (2 mg/kg) solution was administered immediately after the transplantation and additional injections of saline or FK506 (1 mg/kg) were given once a day until the end of the follow-up. To prevent infection, amoxicillin (500 mg/L, Normon, Madrid, Spain) was given in the drinking water for one week. Postoperative analgesia was provided with buprenorphine (0.05 mg/kg). Bladders were expressed twice a day until reflex voiding was re-established.
Functional assessment
Open-field locomotion: motor behavior was tested before surgery and at 3, 7, 14, 21, 28, 35 and 42 d post-operation. Animals were placed individually in a circular enclosure and allowed to move freely for 5 min. Two observers evaluated locomotion during open-field walking and scored the hindlimb performance according to the BBB-scale, ranging from 0 (no movement) to 21 (normal movement). 38
Treadmill locomotion: maximal walking speed, gait pattern, and locomotor coordination were assessed at end time point using the DigiGait Imaging system (Mouse Specifics Inc., Boston, MA) as previously described. 39 Briefly, images captured by a video camera mounted below a transparent treadmill belt were digitized at 140 frames per second, and a minimum of eight sequential step cycles analyzed. For the maximal speed, the treadmill speed was progressively increased from 0 cm/sec until the animal was not able to maintain running. For gait and coordination the rats were recorded at 20 cm/sec, if the animals were able to run at this speed. The start and ending times of the stance phase of each stride were extracted and a foot gait diagram was depicted. 39,40 Quantitative values for the gait parameters (stride duration, stance duration and swing duration) were calculated. To assess the coordination of locomotion the regularity index (RI), the correct alternation between different pairs of limbs and the phase dispersion (PD) were calculated as previously described. 39
Electrophysiological tests
For the electrophysiological tests, the animals were anesthetized with pentobarbital (30 mg/kg i.p.), placed prone onto a metal plate and skin temperature maintained above 32°C. An electromyograph (Sapphyre 4ME, Vickers Healthcare Co., Woking, United Kingdom) was used.
Motor evoked potentials (MEPs) were elicited by transcranial electrical stimulation delivered by two needle electrodes placed subcutaneously over the skull, with the anode over the sensorimotor cortex and the cathode at the nose. Single electrical pulses of supramaximal intensity (25 mA, 100 μs) were applied, and the MEPs were recorded with needle electrodes from tibialis anterior (TA) and gastrocnemius medialis (GM) muscles. 41,42 Peripheral motor nerve conduction tests were performed by stimulating the sciatic nerve with single electrical pulses (100 μs at supramaximal intensity) delivered by needles at the sciatic notch, and recording the compound muscle action potentials (CMAPs) of TA and GM muscles by means of needle electrodes. The recording active electrode was inserted on the belly of the muscle and the reference at the fourth toe. The latency and the onset-to-peak amplitude of the maximal MEP and M waves were measured. 42
Histology
The end time-point of the animals for evaluation of treatment was 42 d after injury. The rats were deeply anesthetized (pentobarbital 60 mg/kg i.p.) and intracardially perfused with 4% paraformaldehyde in PBS. The spinal cord segment from 1cm rostral to 1cm caudal of the injury epicenter (2 cm total length) was harvested and post-fixed in the same fixative solution for 24 h and cryopreserved in PB-30% sucrose.
For evaluation of the injury transversal spinal cord sections 30 μm thick were cut with a cryotome (Leica CM190, Leica Microsystems, Wetzlar, Germany) and distributed in 15 series of 24 sections (separated by 450 μm) each. One series of sections was re-hydrated in water, immersed in hematoxylin Harris solution (Fluka, Sigma) for 5 min, washed in water two times followed by 1% HCl in ethanol solution during 30 sec. The sections were washed with water again and stained with Eosin Y (Merck Millipore, Dramstadt, Germany) during 6 min.
The sections were dehydrated and mounted with DPX (Sigma). Analysis of spared tissue and injury size was made using 19 transversal cord sections (separated by 450 μm between pairs) of each animal. Images of transversal sections were taken at 40x (Olympus BX51) through a digital camera (Olympus DP50). The area of spared tissue, of the cavity and the total spinal cord section were delineated and measured using ImageJ software (NIH, Bethesda MA). Volumes of the graft, spared tissue, cavity and total spinal cord injured segment were calculated using the Cavalieri's correction of morphometric volume. 43
For neurofilament immunohistochemistry sections were blocked with PBS 0.3% with Triton 5% in fetal bovine serum, and incubated for 24 h at 4°C with primary antibodies rabbit anti-NF-L (neurofilament light chain, 1/500, Millipore) and chicken anti-NF-H (neurofilament heavy chain, 1/1000, Millipore). After washes, sections were incubated for 2 h at room temperature with donkey anti-rabbit-AlexaFluor 488 and goat anti-chicken-AlexaFluor 594 (1/200, Jackson Immunoresearch, West Grove, PA). Slides were dehydrated and mounted with Citoseal 60 (Thermo Fisher Scientific, Madrid, Spain). Images of the spinal cord injured epicenter section were taken at 100× with the same setting. Microphotographs were transformed to a gray scale and analyzed using ImageJ software. The NF-H labeled area for each section was measured after defining a threshold for background correction. The percentage of NF-H positive area with respect to the injured area was measured for each animal.
Cell survival assessment
Groups of animals injected with GFP-MSC were sacrificed at 2, 7, 14, 21, and 42 d after injection (n=3 for each time point). Perfusion and processing of the spinal cord was as above. Longitudinal sections of 30-μm thickness of the spinal cord segment were cut and distributed in 12 series of eight sections (separated by 360 μm) each. Sections were blocked with PBS 0.3% with Triton 5% in fetal bovine serum and incubated for 24 h at 4°C with primary antibody rabbit anti-GFP (1/200, Life Technologies). After washes, sections were incubated for 2 h at room temperature with biotinylated conjugated donkey anti-rabbit antibody (1/200, Vector, Burlingame, CA), and then with AlexaFluor 488 conjugated streptavidine (1/200, Jackson Immunoresearch, West Grove, PA). Slides were dehydrated and mounted with Citoseal 60 (Thermo Fisher Scientific, Madrid, Spain).
Analysis of the GFP+ area was performed on eight spinal cord sections (separated by 360 μm between pairs) of each animal. Consecutive images of the spinal cord injured segment were taken at 100×with the same setting and the total section was mounted using Photoshop software (Adobe Systems Inc.). Microphotographs were transformed to a gray scale and analyzed using ImageJ software. The GFP labeled area for each section was measured after defining a threshold for background correction. The total volume of GFP signal for each spinal cord was calculated.
Data analysis
Quantitative data from white matter spared tissue, mean cavity, stance/swing ratio, RI, and electrophysiology tests were analyzed by t-Student or one-way analysis of variance (ANOVA) when more than two groups were compared. Data of graft volume were analyzed using two-way ANOVA. Statistical analyses of BBB score and percentage of spared tissue was performed by two-way ANOVA of repeated measures. Bonferroni's post hoc test for compared pairs of groups was used where needed. Data from maximal treadmill speed was evaluated as survival curve by log-rank test. For the parameters in which no statistical differences between vehicle groups were found, these animals were grouped for the comparative analysis with MSC-treated animals (see supplementary Fig. S1 and Fig. S2; online only). A p value lower than 0.05 was considered significant.
Results
Immunosuppression with FK506 enhances MSC survival in the injured spinal cord
Figure 1 shows the localization of the grafted cells in the spinal cord injured segment after transplantation. GFP labeled cells were observed inside the cord parenchyma around the injection sites and in the tissue injured area (Fig. 1A). To study graft survival, the spinal cords were analyzed at 2, 7, 14, 21, and 42 d post-operation (Fig. 1B) and the stereological volume of GFP immunofluorescence signal was calculated (Fig. 1C). All the transplanted animals had a large volume of GFP signal at 2 and 7 d after injury with a pronounced decrease at following time points. Comparison of the cell dosage (Fig. 1C, top graph) showed a significantly higher GFP signal in animals that received the high dose of cells than those with the low dose at 2 d post-transplantation. Although at longer times after transplantation no statistical differences were found between low and high cell dose groups, at the last analyzed day we found GFP+ cells only in rats transplanted with the high dose. For both low (Fig. 1C, middle graph) and high (Fig. 1C, bottom graph) MSC dose grafts, we observed increased GFP signal at 21 and 42 d after grafting in animals administered FK506. Thus, these results suggest that the higher dosage of cells allowed grafted cells to survive at least until 42 d and importantly, immunosuppression improved graft survival at late time.
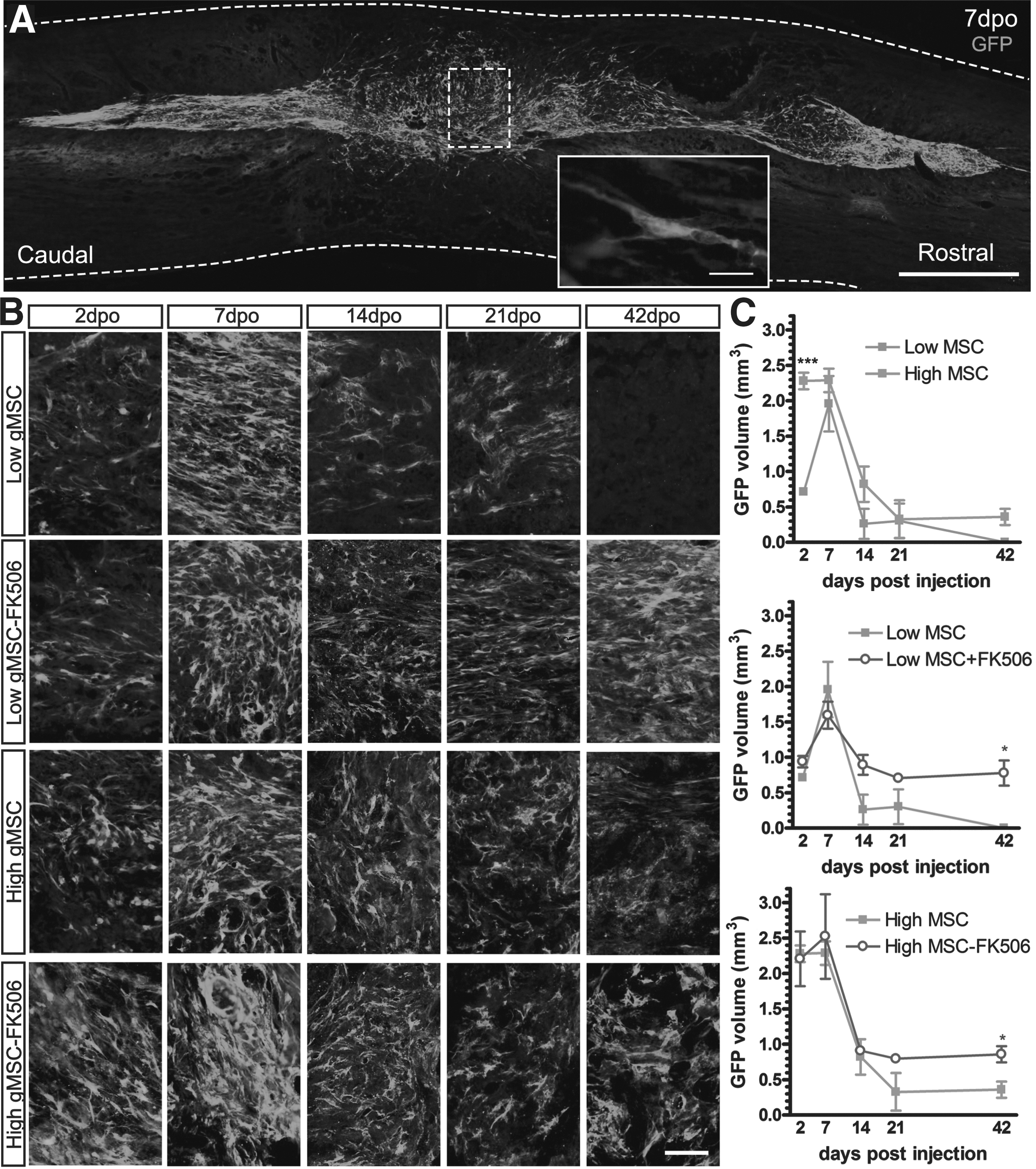
Grafted mesenchymal stem cell (MSC) survival in the injured spinal cord. Green fluorescence protein (GFP)+ MSC grafted cells were localized 7 d after transplantation into the spinal cord
MSC transplant improves functional outcome after SCI
Locomotor function was analyzed by open field voluntary locomotion test using the BBB score and BBB sub-score (Fig. 2), and by kinematic analysis on a treadmill (supplementary Fig. S3). All contused rats displayed immediate but temporary hindlimb paralysis (BBB 0 score) followed by a fast recovery during the next two weeks reaching a plateau phase. In the cell dose analysis (Fig. 2A and B), only the animals with the high dose of MSC had higher BBB score than the control groups at the plateau phase, independently of the immunosuppressive treatment. Regarding the FK506 effect, there were no differences between groups with or without FK506 administration for both low (Fig. 2C) and high MSC graft dose (Fig. 2D). The dose effect was better observed using the BBB sub-score that provides more resolution in the middle-high part of the BBB scale. Again, the two groups with the high dose of MSC reached higher scores than the corresponding control groups, but without differences between them (Fig. 2). In contrast, co-administration of FK506 improved also the locomotion in the animals grafted with the low dose of MSC (Fig. 2).
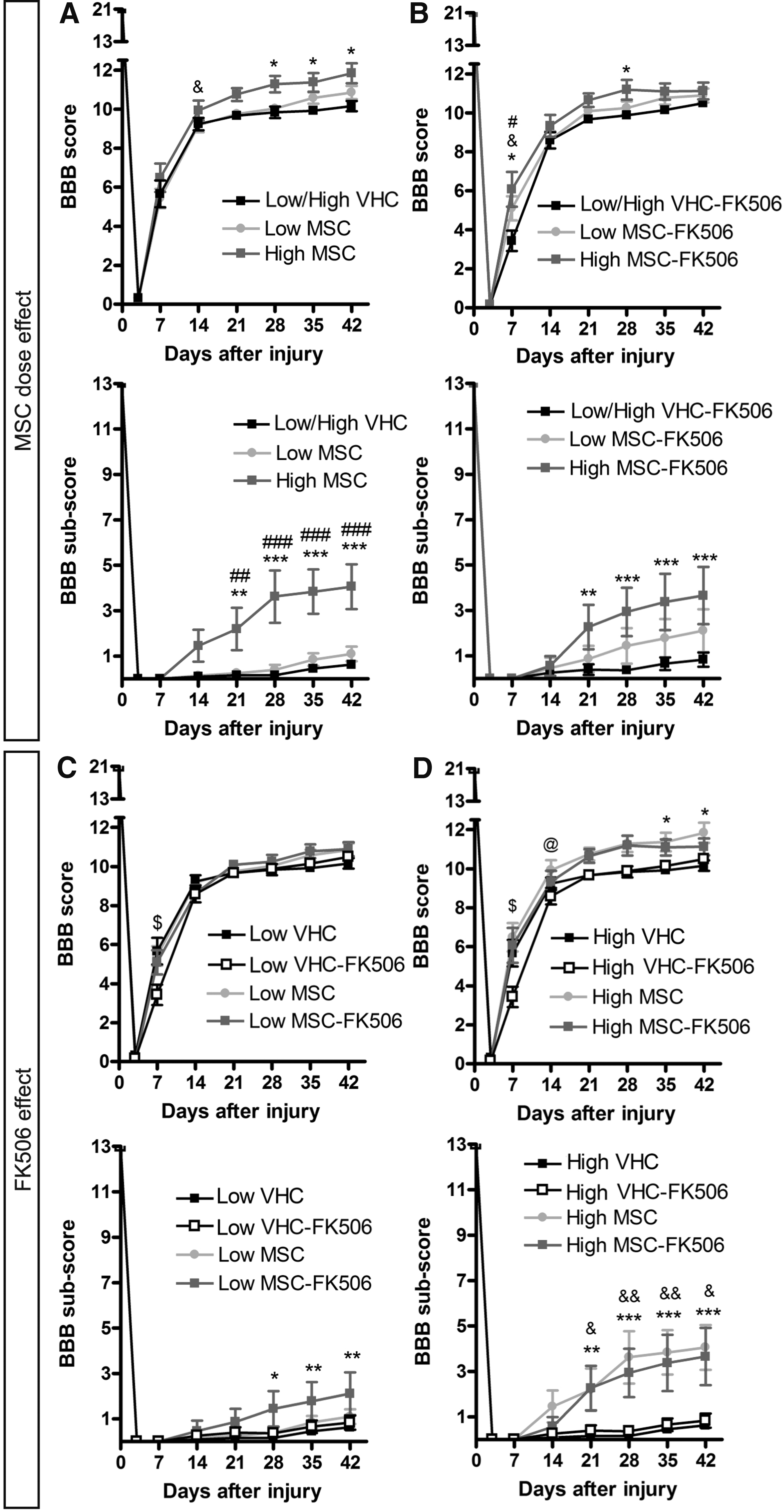
Open field evaluation after spinal cord injury (SCI) and mesenchymal stem cell (MSC) transplantation. The open field locomotion was evaluated weekly after injury using the BBB score and sub-score. The comparative graphs are divided according to MSC dose effect without immunosuppression
In the test of locomotion on the treadmill, we analyzed the maximal speed that animals were able to run with weight support (supplementary Fig. S3) and the gait and coordination performance of animals at 20 cm/sec (supplementary Fig. S4). At the end of the follow-up, all the rats were able to support body weight in stance and run at least at 3 cm/sec. The proportion of animals able to walk decreased with increasing treadmill speed. Animals transplanted with the high dose of MSC were able to run significantly faster than controls, independently of the FK506 administration (supplementary Fig. S3). With regard to gait analysis, the rats increased the forelimb and reduced the hindlimb step number following SCI. As a consequence, the stance and swing times for each step decreased in forelimbs, while the stance time increased in hindlimbs, resulting in an increase of the stance/swing ratio for both fore and hindlimbs in comparison with intact rats. There were no significant differences between treatment groups in the gait parameters.
On the other hand, the regularity index for alternating step pattern, used as a measure of coordination, was markedly reduced in all the groups after injury (supplementary Fig. S4D). The uncoordinated walking was also reflected by impairment of the alternation stepping between ipsilateral and diagonal pairs of limbs (supplementary Fig. S4E). The contralateral alternation of hindlimbs was also reduced in the VHC groups, compared with all the MSC-treated groups (supplementary Fig. S4G), indicating improvement of the correct step sequence of the hindlimbs in MSC-transplanted groups. These changes were corroborated with the phase dispersion measurements, an indicator of the step time delay between two pairs of limbs. After SCI, antero-posterior ipsilateral and diagonal coordinations were affected as shown by progressive increase in the delay or phase dispersion (supplementary Fig. S4F). As a result a significant increase of the slope of linear regression in comparison with normal values was observed in all SCI groups. In contrast, the hind contralateral phase dispersion increased slightly after injury (supplementary Fig. S4G).
The slope of the linear regression was significantly higher in the vehicle injected groups with respect to intact rats, and also compared with Low MSC-FK506, High MSC, and High MSC-FK506 groups. Moreover, the coefficient of variation of the contralateral hindlimbs phase dispersion, a measure of the regularity in the coordination, 39 increased after injury but the Low MSC-FK506, High MSC, and High MSC-FK506 groups showed significantly less increase than the other injured groups. All these results denote a significant improvement of hindlimb coordination by the MSC transplantation that is better with the high dose of cells.
With regard to the electrophysiological results, whereas the CMAPs amplitude remained normal (supplementary Fig. S5), the MEPs were completely abolished in all the rats after SCI, and only a few animals presented small responses (less than 50 μV in amplitude) of normal latency at 42 d post-operation (Fig. 3A–3D). A late MEP (with latency around ∼20 msec) appeared after injury in TA and GM muscles. The proportion of animals that presented MEPs was higher in the MSC-transplanted groups compared to VHC-injected groups. In general, all the transplanted groups showed higher MEPs amplitude than the control groups. The mean amplitude of the second MEP component was higher in the animals grafted with high dose of MSC than in the untreated control groups for the TA muscle (Fig. 3E, 3F), whereas for the GM muscle, the cell dose effect was only significant in animals treated with FK506 (Fig. 3J). With regard to the FK506 effect, animals with immunosuppression had higher MEPs amplitude than injured controls for TA muscle in both the low (Fig. 3G) and the high (Fig. 3H) cell dose, and for GM muscle only in the high cell dose (Fig. 3L).
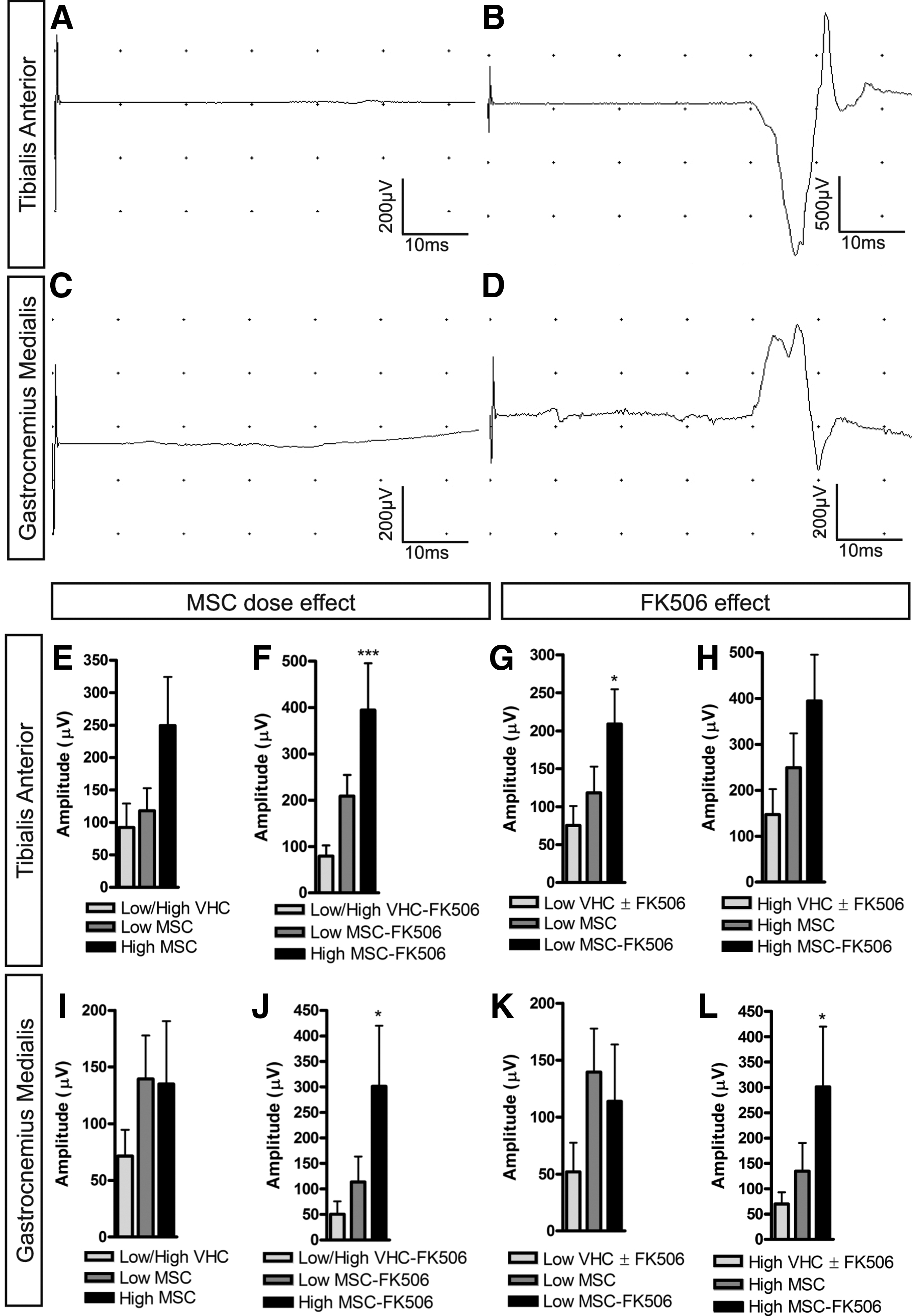
Electrophysiological results after spinal cord injury (SCI) and mesenchymal stem cell (MSC) transplantation. Representative recordings of motor evoked potentials (MEPs) in tibialis anterior (TA)—
All these results indicate that functional recovery induced by MSC transplantation was dependent on the amount of cells grafted and that combination with immunosuppressive treatment slightly enhanced the improvement.
MSC transplantation and FK506 co-administration exert neuroprotection, reduce the cystic cavity and promote axonal growth in the injured tissue
To determine the capacity of the cell graft to provide tissue protection, we evaluated the amount of spared tissue at 42 d after SCI. The cross-section area of the spinal cord was progressively reduced from both rostral and caudal segments to the epicenter of the contusion injury. The amount of spared tissue area was higher in all the MSC groups with respect to vehicle injected groups (Fig. 4). Regarding the MSC dose effect, both Low and High MSC groups had more percentage of spared tissue than the control group in rostral and caudal segments but only the High MSC group showed significant preservation in the epicenter area (Fig. 4G). This dose effect was not observed with co-administration of FK506 (Fig. 4H) since both Low and High MSC-FK506 groups had a similar amount of preserved tissue. Indeed, animals treated with FK506 showed higher proportion of spared tissue than non-immunosuppressed animals, and significantly with respect to control groups at rostral, caudal and epicenter areas (Fig. 4I, 4J).
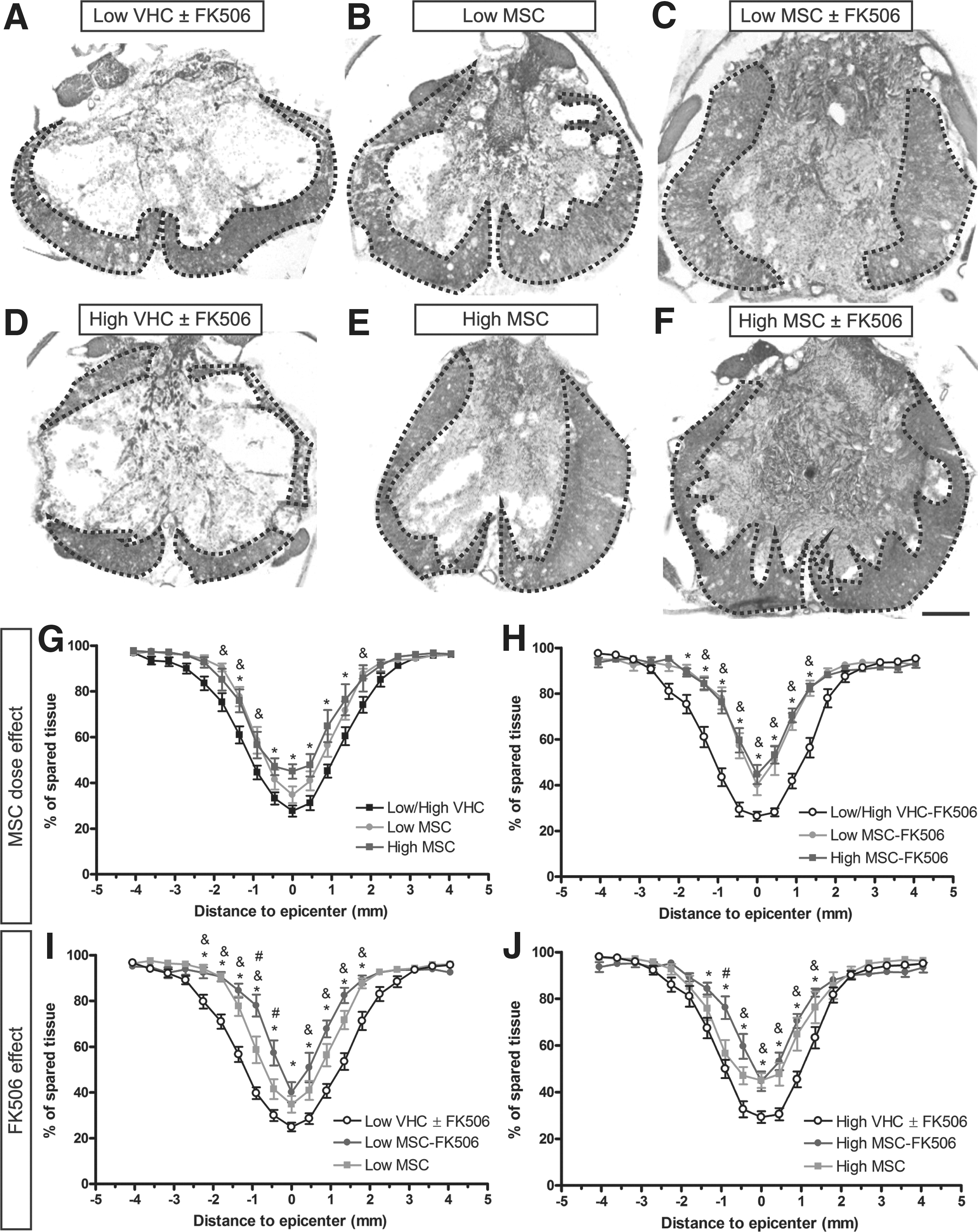
Spared tissue in the injured spinal cord after mesenchymal stem cell (MSC) transplantation. Measurements of spared tissue were made on hematoxylin-eosin stained transversal sections of the injured spinal cord segment (
Moreover, the amount of spared white matter in the epicenter showed similar results to the total spared tissue in the same area (supplementary Fig. S6). Thus, in the case of dose effect without FK506 administration only the High MSC group achieved significant differences versus control groups (supplementary Fig. S6A), while with FK506 treatment, both Low and High MSC-FK506 had more spared white matter than the respective controls (supplementary Fig. S6B). Analyzing the FK506 administration effect, administration of the immunosuppressant significantly increased the amount of spared white matter in the low cell dose compared with controls (supplementary Fig. S6C). In contrast, both High MSC and High MSC-FK506 groups showed similar white matter preservation that was significantly larger than in the respective controls (supplementary Fig. S6D). These findings indicate a neuroprotective effect of MSC transplantation dependent on the amount of grafted cells and immunosuppression.
The stereological estimation of the cavity formed in the injured spinal cord (Fig. 5) revealed a significant reduction of the empty space after MSC transplantation (Fig. 5D–5F) compared to control injured groups (Fig. 5A–5C). In addition, in rats receiving FK506 after MSC transplantation (Fig. 5F) a dense tissue was observed filling the cavity. The reduction of the cyst formation was highest in both Low (Fig. 5I) and High MSC-FK506 (Fig. 5J) groups, indicating an effect associated with the immunosuppression, while no difference between cell doses was observed (Fig. 5G, 5H). The immunohistochemical staining against neurofilaments revealed a high number of axons growing inside the dense tissue filling the cavity in all the animals that received MSC transplantation (Fig. 6), more markedly in animals also treated with FK-506 (Fig. 6J). The NF-H staining in longitudinal sections of the cord of High gMSC-FK506 animals showed a network formed by axons inside the filled cavity (Fig. 7A). The presence of NF-H positive axons was lower in the area occupied by the graft (Fig. 7B, 7C) than in the injured areas where the number of gMSC was small (Fig. 7D).
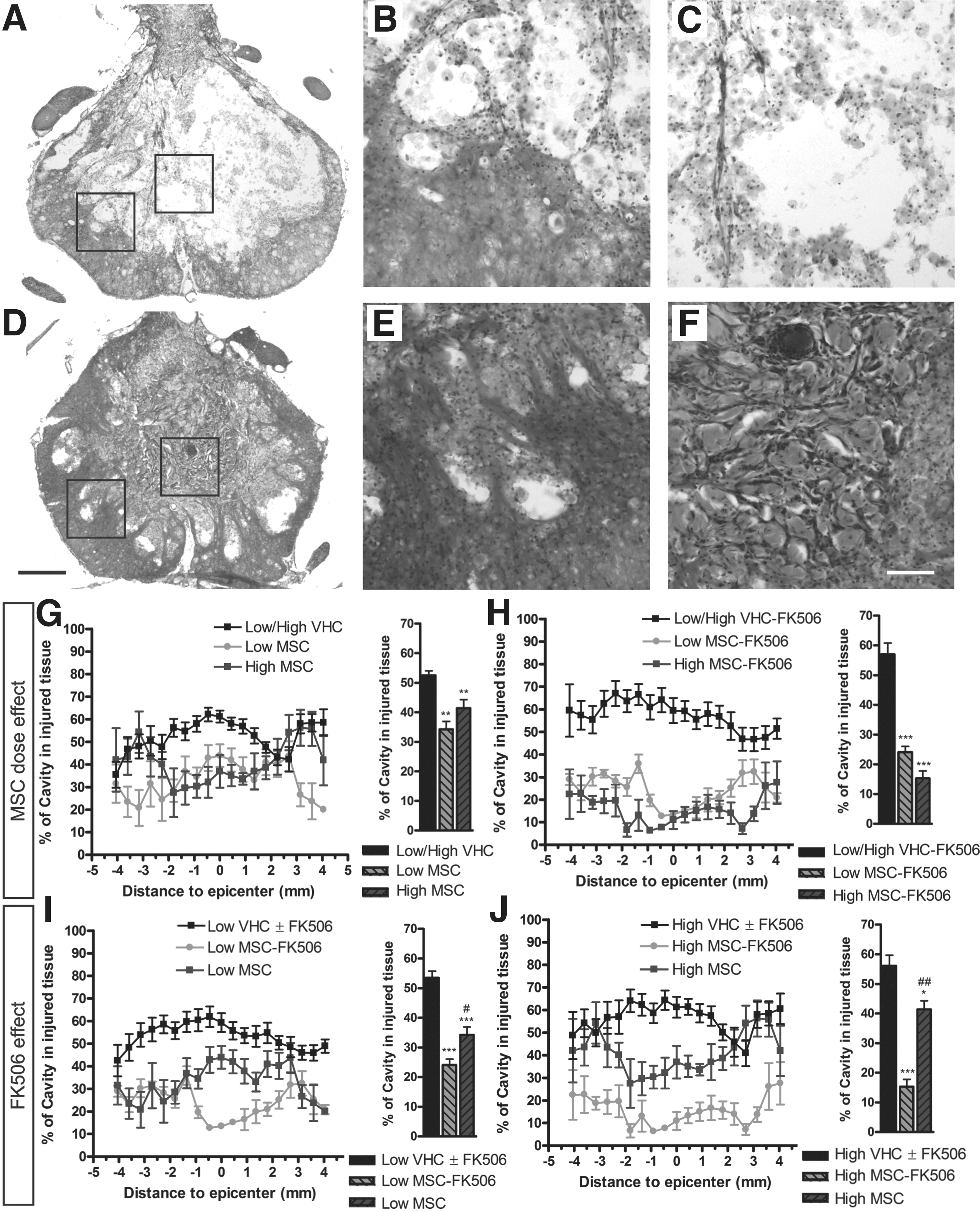
Cystic cavity analysis after spinal cord injury (SCI) and mesenchymal stem cell (MSC) transplantation. Representative hematoxylin-eosin stained transversal sections of the injured spinal cord segment of vehicle injected groups

Neurofilament immunostaining of the spinal cord tissue after spinal cord injury (SCI) and mesenchymal stem cell (MSC) transplantation. Immunohistochemistry for neurofilament heavy chain (NF-H; red) and neurofilament light chain (NF-L; green) performed to detect axons in transversal sections of the spinal cord 42 d after injury. Representative images of vehicle groups

Axonal regeneration within the injured spinal cord tissue after mesenchymal stem cell (MSC) transplantation. Immunohistochemistry for neurofilament heavy chain (NF-H; red) to detect axons and for green fluorescence protein (GFP; green) to detect GFP+MSC (gMSC) in longitudinal spinal cord section of the High gMSC-FK506 group 42 d after injury
Interestingly, some GFP positive MSCs emitted prolongations wrapping NF-H positive fibers (Fig. 7E, 7F), suggesting that engrafted cells contributed to axonal regeneration. All together, these observations indicate that the MSC graft allowed the rescue of some damaged tissue independently of immunosuppression, while combined MSC and FK506 treatment also increased the filling of the spinal cord cavity supporting axonal growth in the new tissue.
Discussion
MSC transplantation has demonstrated potential therapeutic applications in CNS disorders, including traumatic SCI. 2,4,44,45 However, the immunogenic rejection of engrafted MSC after SCI has received less attention. 19 Our study shows longer survival of engrafted MSC after SCI by immunosuppressive treatment with FK506. As a result of this prolonged survival, we observed slight improvement of functional recovery and reduction of damaged cord tissue provided by the graft, and the formation of compact tissue inside the cavity resulting from the spinal cord contusion that allowed axonal growth.
Immunosuppression increases allogeneic MSC graft survival
Survival of allogeneic MSC grafted in the injured spinal cord of immune competent rats was significantly compromised from early time points. This limited survival of the grafted cells is dependent on the amount of transplanted cells since a low concentration of cells (around half a million) resulted in total graft loss by six weeks after transplantation, while injection of a higher number of MSC (around one million) resulted in partial survival during the six weeks of follow up. In contrast, immunosuppression with FK506 resulted in longer survival of the engrafted cells at both doses of MSC grafts. Although a number of works noted that MSC have immunosuppressive properties in vitro, 17 –19 it is not clear yet that this capability is sufficient to prevent their rejection by the host after transplantation in a hostile environment such as the injured spinal cord.
Experimental results suggest that MSC may also behave as non-professional antigen-presenting cells (APC), inducing an immune response. 46 –48 This reaction against the engrafted MSC from rat 21 and human donors 49 has been also observed in the intact spinal cord, with an important recruitment of macrophages, that lead to a dramatic reduction in the number of cells between two and four weeks after injection in comparison to a syngeneic transplant. 21 Further, administration of cyclosporine A, an immunosuppressive agent, prolonged the survival of the MSC grafts, 21,49 confirming the immune rejection induced in the intact spinal cord. After SCI, a limited survival of MSC has been reported for xenogeneic human MSC 13 and allogeneic transplants in contused rats 6,8,9 and dogs. 23
We recently found an increased expression of immune reaction associated genes a few days after allogeneic MSC transplantation in the injured spinal cord. 22 The long-term survival of the MSC graft boosted by immunosuppression is indicative of graft rejection induced by the immune system. Interestingly, immunosuppression prolonged the long-term survival of the grafted MSC but did not seem to affect survival of the MSC during the first week following transplantation (Fig. 1), suggesting that rejection of the cells starts after a few days of injection, coincident with the increase of immune genes expression. 22
Positive effects of MSC transplant combined with immunosuppression after SCI
After SCI, disruption of the spinal cord architecture severely compromises neural functions below the level of the injury. Our detailed functional analysis showed initial complete paralysis of the hindlimbs in all the injured rats, followed by partial recovery of motor skills, and loss of MEPs. The transplantation of MSC after SCI protected the injured tissue and improved the functional recovery, more in animals with the high dose of MSC than with the low dose, as reflected by improvements in the BBB score and sub-score and by the increased speed walking and hindlimbs coordination pattern. In contrast, functional recovery was not enhanced by the combination of FK506 daily treatment to the MSC graft. Thus, a larger amount of MSC during the early times after injury by injecting more cells seemed to be more positive than the prolongation of graft survival by immunosuppression. However, co-administration of FK506 improved the neuroprotective effect exerted by the cells, comparatively more when the number of transplanted cells was low, and resulted in larger MEP responses demonstrating improved descending information across the injury at the lumbar cord segments. Altogether, our observations suggest that the most beneficial effects would be provided by a large amount of transplanted cells early after injury, and by prolonging their presence during more time by the addition of immunosuppression.
It has been reported that MSC transplantation at acute and subacute phases of SCI results in improvement of locomotor performance in mice, 50 rats 7,8,10 –12 and dogs. 23,51 However, such recovery is limited by the restricted survival of the grafted MSC. Our findings demonstrate enhanced recovery when the presence of the grafted cells was increased. The better survival of autologous versus allogeneic MSC grafts resulted in enhanced locomotor skills in SCI dogs, 23 confirming the importance of long-term graft survival for recovery. This conclusion also was reported for olfactory ensheathing cells transplantation after SCI, 52 as well as for peripheral nerve allografts, 31 in which immunosuppression with FK506 improved the functional recovery induced by the grafted cells.
Besides the neuroprotection exerted by the transplanted MSC, administration of FK506 allowed the formation of a dense tissue filling the cavity formed after the cord contusion, which might be constituted by grafted MSC and extracellular matrix. 23 Moreover, we observed axons growing inside this dense tissue filling the cavity that may provide a scaffold for axonal growth, 11,53 improving the chances for late recovery. Interestingly, the presence of MSC influenced the axonal growth inside the new tissue. Axons were not entering delimited areas where a dense mass of MSC was present, probably because the density of cells leaves no room for the growth of axons. In contrast, in sites with a low density of MSC they were ensheathing axons, suggesting a role in the guidance of axonal regeneration.
MSC grafts used for repairing peripheral nerve injury 54 and root avulsion injury 55 also demonstrated positive effects on axonal regeneration. Co-cultures of MSC with neurons 53 and with spinal cord slices 22 enhanced neurite growth in vitro, suggesting that factors expressed by these cells promote axonal regeneration, even in a non permissive substrate. 53 In fact, MSC may express adhesion molecules and receptors, such as Ninjurin1 and 2, Netrin 4, 56 axonal guidance factors, such as robo1 and 4, 57 metalloproteinases MMP1 and MMP2 that can degrade the extracellular matrix, 58,59 and also fibronectin, laminin and collagen, offering the possibility to act as a cell bridge and to reduce the inhibitory environment for axonal regeneration.
Potential effect of FK506 for recovery after SCI
Besides the immunosuppressive effects as an immunophilin ligand, FK506 has diverse neuroprotective actions after central and peripheral neural injuries. 26,35,60 After SCI, the beneficial effects of the FK506 appear to be dependent on the injury model. Thus, improvement of locomotor function by FK506 administration was reported following photochemical 36,52 and hemisection 35 lesions but not after contusion 61 and compression 62 of the spinal cord. Further, the neuroprotective effects of FK506 also are dependent on dose and timing of administration. 60,63,64 We selected a dose of 1 mg/kg/d according to previous reports, since this dose has cell graft protective action 64,65 but does not induce functional and histological benefits in a SCI model similar to ours. 62 Our findings confirm that the dose of FK506 used improved graft survival but without marked effects in locomotor recovery and tissue sparing in non-transplanted animals (supplementary Fig. S1 and Fig. S2).
In conclusion, the findings of this study support the notion that allogeneic MSC grafts trigger an immune response in the host and that immunosuppression may rescue engrafted MSC from rejection. Nevertheless, the longer graft survival only resulted in a limited enhancement of functional recovery, and increased tissue sparing when a low number of cells was transplanted. In addition, the cell–dose effects observed are indicative of the importance of the amount of transplanted cells. On the other hand, the tissue formed filling the cavity with MSC under immunosuppression could act as a bridge for axonal regeneration, an interesting feature for combined therapies in long-term treatments. Considering these results, sufficient quantity of allogeneic MSC transplant plus immunosuppressive treatment may be a feasible strategy to improve the outcomes in regenerative therapies for SCI.
Footnotes
Acknowledgments
We thank Monica Espejo, Jessica Jaramillo, and Marta Morell for technical help. The GFP-viral vector was kindly provided by Dr. Francisco J. Rodríguez from the research center of Hospital de Parapléjicos de Toledo, Spain. This work was supported by grants from the Ministerio de Ciencia e Innovación (grant SAF2009-12495), and funds from CIBERNED and Red de Terapia Celular (TERCEL), Instituto de Salud Carlos III of Spain.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.