
Abstract
Spinal cord injury (SCI) is associated with rapid and sustained impairments in cardiovascular function that ultimately cause an early onset of cardiovascular disease. We know remarkably little about the temporal progression of cardiovascular disturbances, but such an understanding is critical to inform clinical management and develop appropriate intervention strategies. To characterize the cardiovascular response to SCI, six male Wistar rats were instrumented with telemetry and assessed for continuous arterial blood pressure (BP), core body temperature, and heart rate (HR) 7 days before and up to 28 days after T3 SCI. Hemodynamic variables were averaged day by day and hour by hour. Spontaneously occurring autonomic dysreflexia (AD) was characterized by applying a novel algorithm to continuous BP and HR data, and induced AD was assessed weekly via the BP response to colorectal distension. Systolic BP was reduced at all time points after SCI compared with before SCI (p<0.003), except at 4 and 6 days post-injury. Core body temperature was reduced at 2 days post-SCI only (p=0.001). The nocturnal dip in BP and temperature observed pre-SCI was absent during the first 14 days post-SCI, but returned from 21 days post-SCI on (p<0.024). The frequency and severity of spontaneously occurring AD events were significantly less between days 6 and 10 post-SCI compared all other time points (p<0.037). The pressor response to colorectal distension was greater at 14, 21, and 28 days post-SCI compared with at 7 days post-SCI (all p<0.004). In conclusion, SCI induces rapid and profound alterations in basal hemodynamics and diurnal rhythms that partially recover by 14 days post-SCI. AD, on the other hand, is acutely present post-SCI, but the frequency and severity of AD events increase substantially from 14 days post-SCI on.
Introduction
Spinal cord injury (SCI) at or above the major splanchnic sympathetic outflow is associated with a myriad ocardiovascular disturbances that may result in end organ damage and an early onset of cardiovascular disease. 1,2 Although chronic cardiovascular complications have been well documented, 3 –5 we know remarkably little about the temporal development of abnormal cardiovascular control in the acute and subacute phases of SCI. An improved understanding of cardiovascular disturbances during this critical time window is expected to inform clinical management and allow for appropriate intervention strategies to be developed. The innovation of clinically relevant rodent SCI models and high-fidelity telemetric blood pressure (BP) transducers circumvents many of the complexities of conducting such time-course studies in humans, and allows for a robust approach to conducting serial assessments of cardiovascular function across time. In the current study, we employed this approach to investigate the hemodynamic response pre- and post-SCI, with a particular focus on the development of autonomic dysreflexia (AD) and alterations in diurnal rhythms.
“Autonomic dysreflexia,” a term that describes potentially life-threatening episodic hypertension, has been reported to occur in both acute and chronic SCI patients. 6,7 In animal models, the progression of AD has traditionally been assessed by conducting serial measures of BP and heart rate (HR) in response to some form of physiological stimuli (i.e., bladder stimulation or colorectal distension [CRD]). 8,9 Using this “induced” method, AD has been reported to occur acutely (1–2 days post-SCI), 10 although the “typical” dysreflexic response is not fully developed until 4–6 weeks post-SCI. 4,8 More recently, with the advancement of telemetry devices, it is now possible to investigate the incidence of spontaneously (i.e., naturally) occurring AD by applying an “AD detection algorithm” to continuous BP data. 5,11 Only two studies have taken this approach and generally agree that there is an initial spike in AD frequency in the acute period post-injury, followed by a slight decline in AD frequency during the subacute period, and a re-emergence of a high daily incidence of AD in the chronic period. 5,11 Although the frequency of spontaneously occurring AD was well characterized, the duration and severity of spontaneously occurring AD in addition to the time of day at which AD occurs is yet to be elucidated. Understanding the “complete AD phenotype” with respect to incidence, duration, severity, and time of day is a critical first step in determining the potentially deleterious effects of repeated episodes of AD on the cardiovascular system, and to investigating treatments aimed at reducing AD.
Diurnal rhythmicity in BP has been investigated in both the clinical SCI population and in animal models of SCI. In the clinical population, there is general agreement that individuals with complete cervical SCI exhibit no circadian oscillations in BP or HR, 12 –15 whereas individuals with low-thoracic SCI exhibit normal circadian oscillations characterized by the presence of a “nocturnal dip” in BP and HR. 13,15 In a mid-thoracic animal model of SCI, we have previously shown that circadian oscillations in BP were absent in the acute period following SCI (0–6 days), but returned between 6 and 10 days post-SCI. 3 Circadian oscillations in BP are thought to resemble those of sympathetic nerve activity; 16 hence, the recovery of circadian BP oscillations in our previous study 3 is likely caused by the subsidence of neurogenic shock and the partial preservation of descending sympathetic control that accompanies a T5 model. In this respect, circadian oscillations in BP control in an animal model with a more rostral injury are yet to be elucidated. Also, no study has investigated how circadian oscillations change from acute-to-chronic SCI or the effect that SCI has on other hemodynamic variables that are usually under sympathetic control, such as core body temperature.
In the present study, we extended previous work by concurrently investigating alterations in multiple hemodynamic variables from acute-to-chronic SCI. Using high-fidelity telemetric blood pressure monitoring, we circumvented the limitations of fluid-filled catheter approaches and were able to track beat-by-beat hemodynamics and core body temperature in the same animal pre-to-post SCI. Finally we explored the progression of AD from acute-to-chronic SCI by applying our novel AD detection algorithm to beat-by-beat BP and HR data, and by investigating the hemodynamic response to CRD on a weekly basis.
Methods
Experimental design
We conducted a within animal pre- and post-SCI design to specifically investigate the cardiovascular and autonomic changes that accompany SCI. Experiments were conducted on six male Wistar rats (250–300 g; Harlan Laboratories). Three weeks prior to SCI surgery, rats were fitted with a radiotelemetric device for the subsequent continuous (24 h/day) assessment of arterial BP, HR, and core body temperature. To ensure that rats had adequately recovered from telemetry implantation, intact values were averaged between days 14 and 17 post-implantation (i.e., days 7–4 pre-SCI). These time points also ensured that any prophylactic SCI surgery medication did not interfere with intact cardiovascular indices (see later section SCI surgery and animal care ). Responses at various time points post-SCI were compared with pre-SCI values. All procedures were conducted in strict accordance with the Canadian Council for Animal Care and approved by the University of British Columbia Animal Care Committee.
Surgical procedures
Telemetry implantation
The telemetry device (TRM54P, Millar, Auckland, New Zealand) was prepared for implantation according to manufacturer guidelines. Rats were pretreated for 3 days with prophylactic enrofloxacin (Baytril; 10 mg/kg, s.c., Associated Veterinary Purchasing; AVP, Langley, Canada). On the day of surgery, rats were anesthetized with isoflurane (initial chamber induction at 5% isoflurane with 2 L/min oxygen, followed by maintenance on a Bain system at 1.5% isoflurane with 1.5–2 L/min oxygen). The abdomen was shaved and rubbed with iodine. A midline abdominal incision was made and the descending aorta was exposed via blunt dissection. The aorta was briefly occluded with 4-0 silk and punctured 1–2 mm anterior to the iliac bifurcation. A 20 gauge curved needle was used to guide the tip of the telemetry device into the aorta, after which it was advanced rostrally such that the tip was just distal to the renal arteries. The catheter was fixed in place using a small amount of tissue adhesive. The body of the telemetry device was secured to the abdominal wall using 4-0 silk sutures. The abdominal wall was then closed using 4-0 Vicryl subcuticular sutures. Animals received warmed lactated Ringer's solution (5 mL, s.c.) and recovered in a temperature-controlled environment (Animal Intensive Care Unit, Los Angeles, CA). Enrofloxacin (10 mg/kg, s.c.), buprenorphine (0.02 mg/kg, s.c.), and ketoprofen (5 mg/kg, s.c.) were administered once a day for 3 days postoperatively.
SCI surgery and animal care
Rats were pretreated for 3 days with prophylactic enrofloxacin (Baytril; 10 mg/kg, s.c., AVP). On the day of surgery, rats were anesthetized with ketamine hydrochloride (Vetalar; 70 mg/kg, i.p.; AVP) and medetomidine hydrochloride (Domitor; 0.5 mg/kg, i.p.; AVP). A dorsal midline incision was made in the superficial muscle overlying the C8-T3 vertebrae. The dura was opened at the T2-T3 intervertebral gap and the spinal cord was completely transected using microscissors. Complete transection was confirmed by two surgeons via visual separation of the rostral and caudal spinal cord stumps, and Gelfoam (Pharmacia & Upjohn Company, Pfizer, New York) was placed between the stumps to achieve hemostasis. The muscle and skin were closed with 4-0 Vicryl and 4-0 Prolene sutures, respectively. Animals received warmed lactated Ringer's solution (5 mL, s.c.) and recovered in a temperature-controlled environment (Animal Intensive Care Unit, Los Angeles, CA). Enrofloxacin (10 mg/kg, s.c.), buprenorphine (0.02 mg/kg, s.c.), and ketoprofen (5 mg/kg, s.c.) were administered once a day for 3 days postoperatively. The bladder was manually expressed three times (8 a.m., 2 p.m., 10 p.m.) daily until spontaneous voiding returned (∼10 days post-injury). Daily animal monitoring was conducted between 8 a.m. and 10 a.m.
Hemodynamic monitoring
Beat-by-beat arterial BP, HR, and body temperature were continuously monitored 24 h/day, for 3 days pre-SCI (days 14–17 after transmitter implantation) and every other day post-SCI until day 28. This protocol was designed to allow “odd” days to be used for the assessment of CRD-induced AD (see later section Induced AD ) and to reduce the enormous amount of data generated from beat-to-beat analyses. Hemodynamic variables were monitored at 1000 Hz during each 24 h period, and the mean for the entire day was reported. Because of the need for animal monitoring, we deemed it necessary to remove the following hemodynamic data from all subsequent analyses. For the first 10 days post-SCI, rats were monitored three times per day, but the bulk of the monitoring (e.g., medications, cage changes, weighing) was conducted in the morning (8–9:30 a.m.); hence we completely eliminated this time from the analyses. The other monitoring times were for bladder expression only, and lasted no longer than 2–3 min per animal. For these checks, those 10 min before and 30 min after bladder expression were also removed. From day 10 onwards, reflexive micturition occurred, and rats were only checked once a day in the morning. CRD assessments never took place on the same day as the 24/7 data recordings.
Assessment of diurnal rhythmicity in hemodynamics
On days 4 and 8 (acute SCI), and on days 14, 20, and 28 (chronic SCI), hemodynamic variables were additionally averaged on an hour-by-hour basis such that the diurnal rhythmicity in hemodynamic variables could be assessed. For these analyses, hemodynamic variables during night hours were compared against hemodynamic variables during daytime hours. For core body temperature, we also calculated the peak-to-trough variation. 17
Spontaneously occurring AD
We developed a novel AD detection algorithm that was capable of screening beat-by-beat BP and HR data to detect the incidence, severity, and duration of spontaneously occurring AD (MATLAB, The MathWorks, Inc., Natick, MA). Our algorithm extended previous work by Rabchevsky and coworkers 5 and Zhang and coworkers, 11 who had previously used a basic algorithm to detect abnormal spikes in BP and troughs in HR that resembled the hypertension and bradycardia typically associated with AD. A potential limitation of the precedent approaches, however, is that BP and HR raw data were down-sampled from their initial sampling frequency, which may have resulted in an underestimation of the true peak pressor response during AD. Furthermore, AD was defined as a 10 mm Hg increase in mean arterial pressure (MAP) above baseline, coupled with a 10 bpm drop in HR, the latter of which seemed inappropriate to define bradycardia, given the high resting HR of a rodent. We circumvented these potential limitations by recording beat-by-beat systolic BP (SBP) and HR telemetry recordings 24 h/day for 3 days pre-SCI and every other day post-SCI until day 28 (Lab Chart; AD Instruments. Inc., Dunedin, New Zealand). Beat-by-beat SBP and HR (calculated from R-R interval) were extracted from the raw data using commercially available software (Lab Chart) and imported into MATLAB. SBP and HR were screened for non-physiological values, and any data point that had a HR<180 or >625 bpm was excluded from the analyses. Next, a moving average window of 240 sec was created. An “AD threshold” was then set at 20 mm Hg above the moving average baseline in order to detect sudden spikes in SBP. A cluster of SBP spikes was defined as an AD event in which the following conditions were met: 1) SBP peaks exceed the “AD threshold” for a duration>10 sec, and 2) there was a corresponding mean HR drop of ≥40 bpm during the last 75% of the AD event relative to the mean HR during the first 10% of time during the potential spontaneous AD event. The 40 bpm was chosen to reflect a similar percentage drop from baseline to that typically observed clinically. If more than one AD event was identified within 120 sec of the preceding event, these events were amalgamated and counted as one AD event. For each identified AD event, we extracted the pressor response, maximum SBP, average SBP, minimum HR, average HR, duration of the event, and time of day. An sample AD event detected using this algorithm is presented in Figure 1.
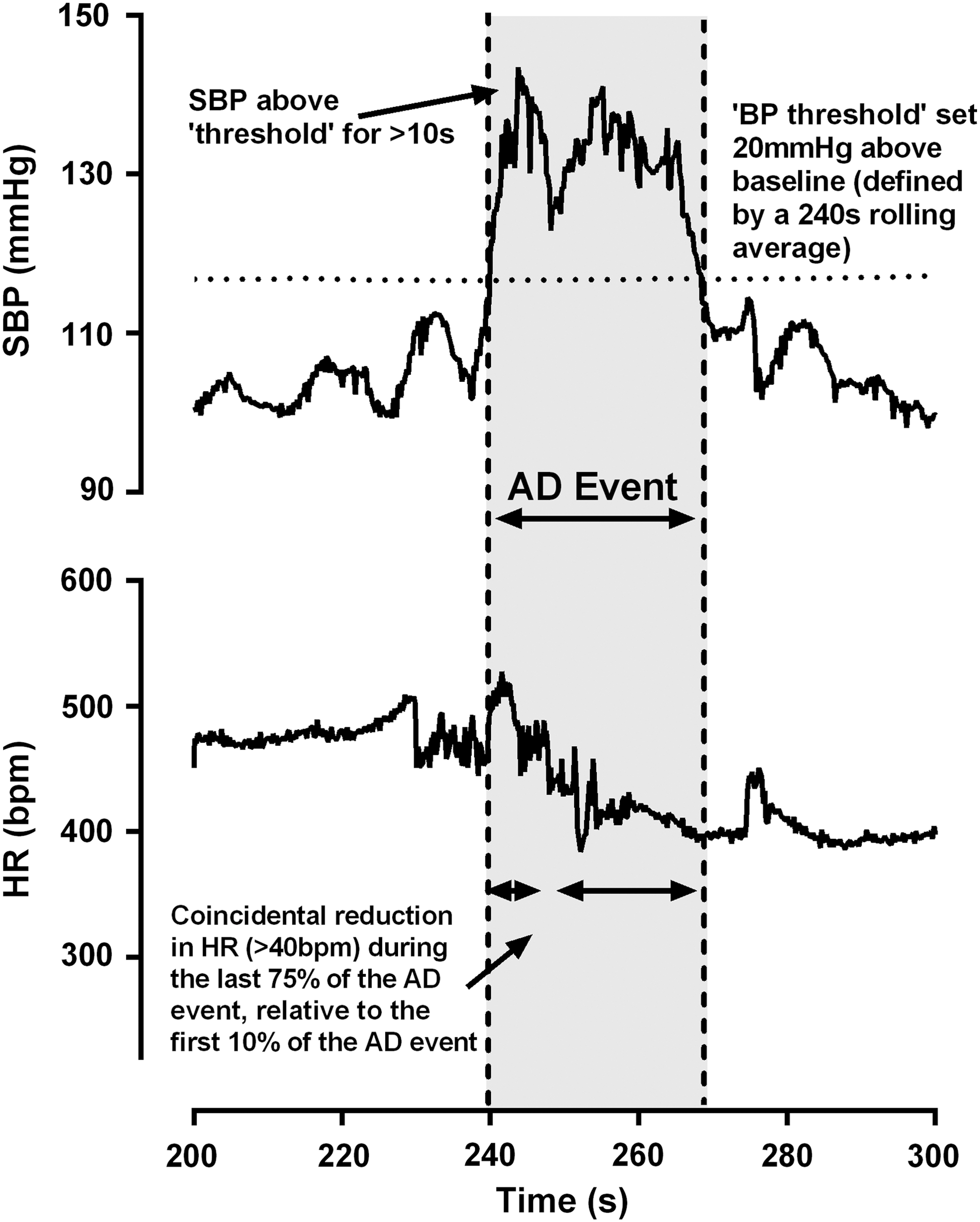
Example of detection of an autonomic dysreflexia (AD) event using our algorithm. First, a 240 sec rolling average baseline is fitted to the systolic blood pressure (SBP) and heart rate (HR) data. Next, a SBP and HR “threshold” is transposed 20 mm Hg above the moving baseline or 40 bpm below the moving baseline, respectively. To be considered an AD event, the SBP must rise above the threshold and remain elevated above threshold for at least 10 sec. During this time, there must also be a coincidental reduction in HR below the threshold.
Induced AD
Severity of induced AD was assessed on a weekly basis on days when 24/7 blood pressure monitoring was not conducted (days 7, 13, 21, and 29 post-SCI). AD was induced via two bouts of CRD, a procedure that is well characterized in our laboratory. 9,18,19 Briefly, a small deflated plastic balloon (the balloon tip of a Swan–Ganz catheter; 10 mm in length) was inserted rectally for a distance of 1.5 cm. A custom-built tube was placed around the outside of the catheter to prevent rats from biting/pulling out the catheter. After insertion, rats were given 10 min to freely move in their home cages. When BP and HR had stabilized, the balloon was infused with 2 mL of air over 10 sec, and distension was maintained for 1 min. CRD was performed twice with a minimum interval of 10 min. Beat-by-beat data were averaged over 1 sec intervals, and the maximum increase in SBP and maximum decrease in HR was averaged over the two trials.
Statistical analysis
Dependent variables were assessed for assumptions of parametric testing. Alterations in hemodynamics across time were assessed using a one way repeated measures ANOVA with Bonferroni-corrected post-hoc comparisons. The presence or absence of a nocturnal dip in hemodynamics along with the incidence of AD during day and night cycles were assessed using a two factor repeated measures ANOVA, with one factor for time of day and one factor for time post-injury. Statistical analyses were conducted using STATA v12.1. Statistical significance was accepted at p<0.05.
Results
SCI is associated with time-dependent changes in BP, HR, and core body temperature
SBP was reduced at day 2 post-SCI compared with pre-SCI (p=0.001; Fig. 2). There was a transient recovery of SBP at days 4 and 6 post-injury, such that SBP was not different at these time points from pre-SCI (both p>0.092). From day 8 onwards, there was a chronic reduction in SBP compared with pre-SCI (all p<0.002). Core body temperature was reduced at day 2 post-SCI compared with pre-SCI (p=0.001), but recovered by day 4. There was a spurious increase in core body temperature at day 14 post-SCI compared with all other days (all p<0.047). There was a trend toward an increase in HR at days 2 and 4 post-SCI compared with pre-SCI, but this just failed to reach statistical significance (both p=0.056).

Temporal changes in heart rate (HR) (panel
Loss of rhythmicity in cardiovascular function in response to SCI is partially reversible with time
Prior to SCI surgery, all rats exhibited typical diurnal rhythmicity in cardiovascular control, as characterized by the presence of nocturnal dip (all p<0.012: Fig. 3). During the first 14 days post-SCI, there was no difference in SBP or core body temperature between day and night (all p>0.323), implying the normal diurnal rhythmicity in BP and core body temperature was disrupted. In particular, animals struggled to increase core body temperature during the day, and the peak-to-trough value was substantially higher at days 4 (3.84±1.87°), 7 (2.09±0.75°) and 14 (2.21±0.52°) post-SCI compared with pre-SCI (1.26±0.23°; all p<0.038). Conversely, the nocturnal dip in HR remained stable after SCI, as evidenced by a significantly lower HR during night compared with day (all p<0.019). At 3 and 4 weeks post-SCI, rats regained some degree of cardiovascular control, as evidenced by the recurrence of a nocturnal dip in SBP and core body temperature (all p<0.024). Similarly, the peak-to-trough value for core body temperature was no longer statistically different at day 28 post-SCI (1.68±0.39°) compared with pre-SCI.

Diurnal rhythms for heart rate (HR), systolic blood pressure (SBP), and core body temperature stratified by time after spinal cord injury (SCI). Black and white rectangles represent night and day cycles, respectively. Note that there was substantial disruption in typical diurnal rhythms during the first 2 weeks post-SCI (panels
SCI is associated with time-dependent development of AD
We used a custom-built algorithm to detect the incidence, severity, and duration of spontaneously occurring AD, along with the time of day at which AD occurred (Fig. 1). During the three weeks of data collection pre-SCI, our algorithm picked up an average of one “AD event” in each of the animals (data not shown). It should be noted that these patterns of BP that resembled an AD event pre-SCI never exceeded 12 sec in duration. Post-SCI, we found that animals exhibited AD as early as days 2 and 4 post-SCI (Fig. 4). During this acute period, animals exhibited up to 14 AD events per day, with an average pressor response of 28.5±11.1 mm Hg. Between days 6 and 12 post-SCI, AD incidence was lower than at day 4 and day 14 onwards (all p<0.035). Both the maximal SBP and magnitude of the AD pressor response was lower between days 6 and 12 compared with day 14 onwards (all p<0.006). From day 22 onwards, the AD pressor response exceeded all previous time points (all p<0.0022), except for day 4, when there were no statistical differences. There were no differences across time for the degree of AD-induced bradycardia, the duration of AD events, or whether AD occurred predominantly during the light or dark period. By day 28 post-SCI, rodents exhibited up to 39 AD events per day with a maximum pressor response of 68 mm Hg.

Data collected from each of the spontaneous autonomic dysreflexia (AD) events stratified by time post-injury. During the first 4 days post-injury there was a high daily incidence of spontaneously detected AD (panel
Further to investigating spontaneously occurring AD, we also conducted weekly evaluations of AD severity by assessing the hemodynamic responses to CRD (Table 1 and Fig 5). We found that the pressor response to CRD was minimal at day 7 post-SCI, was increased at day 14 (p=0.004 vs. day 7) and day 21 (p=0.003 vs. day 7), but was most pronounced at day 28 post-SCI (p<0.034 vs. days 7, 14, and 21 post-SCI; see also Fig 5).
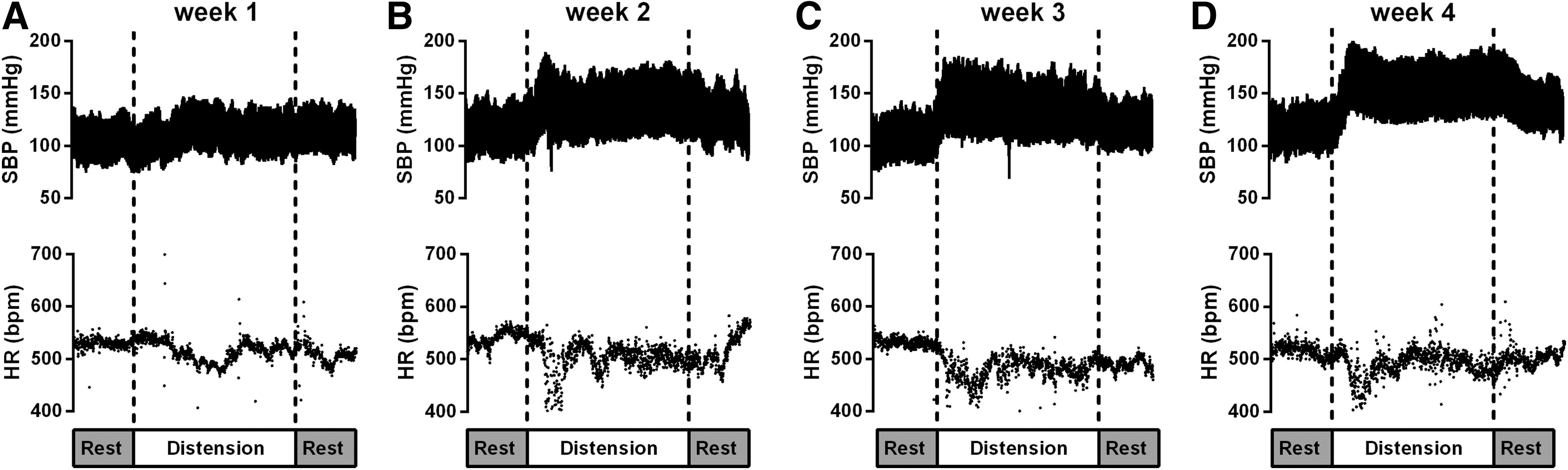
Example of systolic blood pressure (SBP) and heart rate (HR) response to colorectal distension in one rodent with spinal cord injury (SCI). Note that autonomic dysreflexia was not present during the 1st week post-SCI (panel
Hemodynamic Responses to Colorectal Distension by Time Since Spinal Cord Injury
p<0.05 vs. week 1; † p<0.05 vs. week 2, ‡ p<0.05 vs. week 3.
SBP, systolic blood pressure; MAP, mean arterial blood pressure; HR, heart rate.
Discussion
Our study is the first to concurrently examine resting hemodynamics, thermoregulation, CRD-evoked AD, and spontaneously occurring AD events across time pre- and post-SCI. We found that acute SCI is associated with rapid alterations in cardiovascular control that manifests as hypotension, reduced core body temperature, and loss of typical diurnal rhythms in hemodynamics. Using our novel AD detection algorithm, we found that spontaneous episodes of AD were present in acute SCI, but were significantly reduced during the subacute period (i.e., days 6–10 post-SCI). From day 14 post-SCI onwards, diurnal rhythmicity started to return, albeit to an altered state, but hypotension persisted. Day 14 onwards was also the point at which the daily incidence (up to 39 events) and severity of spontaneous AD (up to 68 mm Hg) was markedly increased compared with the subacute period, and the day at which rodents generated a significant pressor and bradycardic response to colorectal distension.
Basal hemodynamics and core body temperature
The temporal BP response to complete T3 SCI was characterized by an acute reduction in arterial BP, a brief recovery to pre-SCI levels, and a subsequent persistent hypotension. Acute hypotension is in agreement with previous studies in animal models of high-thoracic SCI, 3,4 and is likely secondary to neurogenic shock and a consequent depression of sympathetic activity and spinal reflexes. The persistent hypotension we found in the chronic phase post-injury, however, cannot be explained by the same phenomenon, because neurogenic shock typically subsides rapidly within the first few days in rodent models of SCI. 3 Instead, chronic hypotension is likely caused by loss of suparspinal control over the majority of spinal sympathetic preganglionic neurones, especially those in the splanchnic bed, which are critical for the maintenance of arterial BP. 20 Chronic hypotension in our rodent model of SCI is in agreement with previous studies that have utilized a complete high-thoracic transection model, 4 but differs from that found in studies that utilized a mid-thoracic (T5) 50 g clip compression incomplete model of SCI, whereby BP recovers to pre-SCI levels within 5–10 days. 3,21 This difference in responses may be attributed to a partial preservation of bulbospinal sympathoexcitory inputs to sympathetic preganglionic neurons in a clip compression model, which are absent in a complete transection model. In this respect, animals and humans with the greatest loss of descending sympathetic control exhibit the greatest impairments in cardiovascular function. 21 –23 To compensate for the acute hypotension, animals exhibited tachycardia that is likely mediated by both vagal withdrawal and an increase in supraspinal sympathetic drive to the intact T1 and T2 spinal sympathetic neurones that ultimately innervate the heart. In this respect, mid-thoracic experimental SCI has been shown to produce a pronounced increase in left-ventricular sympathetic innervation and robust arborization of cardiac sympathetic preganglionic neurones. 24 That there was no chronic tachycardia in the face of persistent hypotension is likely the result of a chronic reduction in cardiac output, which we have previously reported with this injury model. 25
During the first 4 days post-injury, rodents exhibited a substantial fall in core body temperature. This acute hypothermia is in agreement with previous reports in animal models with high-lesion SCI. 4 Unfortunately, no study has examined core temperature in humans during acute SCI, but there are numerous reports of chronic SCI individuals, especially those with injury above T6, exhibiting poikilothermia. 26,27 Impaired thermoregulation during acute SCI is attributed both to a lack of an afferent pathway from skin receptors to temperature regulation centers, 28 and to sympathetic decentralization, which limits the ability to vasoconstrict, vasodilate, and sweat in the insensate portion of the body. 29 The consequent loss of tone in cutaneous vascular beds likely induces vasodilation and a consequent increase in skin blood flow and heat loss through dissipation. In this respect, Laird and coworkers 4 reported that the large reduction in core body temperature during the first few days post-injury was accompanied by an increase in surface (mid-tail and hindpaw) temperature at the same time points. By day 4 post-SCI, daily averages for core body temperature returned to pre-injury values. Interestingly, this is the same time at which SBP also recovered to pre-injury levels, suggesting the recovery is caused, at least in part, by the subsidence of spinal shock.
Diurnal rhythms
Diurnal oscillations in BP were disrupted during the first 2 weeks post-SCI, but recovered by 3 weeks post-SCI. Clinical studies that have investigated diurnal control of BP in chronic SCI individuals reveal that tetraplegics with complete loss of descending sympathetic input to the sympathetic preganglionic neurons exhibit disrupted circadian control, whereas paraplegics exhibit relatively normal diurnal rhythmicity. 15 We attribute the initial impairment of diurnal BP control to neurogenic shock, which depresses sympathetic activity. From 2 weeks post-SCI onwards, neurogenic shock subsides and the nocturnal dip in BP returns. This partial recovery of the nocturnal dip was surprising, given the majority of the spinal sympathetic neurons that are crucial for the generation of sympathetic rhythmic activity (i.e., the T3–T12 region) remain disconnected from supraspinal regulation with our injury model. This leads us to speculate that either there is an overcompensation of sympathetic activity through rostral spinal sympathetic circuits, or that unopposed spontaneous firing of sublesional sympathetic circuitry is sufficient to recover some diurnal rhythmicity. Unlike BP, diurnal HR rhythms remained relatively well preserved post-SCI, most likely because of the heart retaining partial supraspinal sympathetic control and full vagal control with our T3 injury model. Perhaps the most remarkable alterations in diurnal rhythms were seen with core body temperature. Although daily averages for core body temperature appeared to return to normal from day 4 post-SCI onwards, we revealed substantial disruptions to normal diurnal control that were characterized by a “slow” kinetic response and a substantial increase in the peak-to-trough variation. By 2 weeks post-SCI onwards, the nocturnal dip in core body temperature returned, but the peak-to-trough variation was still substantially higher than pre-SCI. Increased peak-to-trough variation in chronic SCI is consistent with that in the clinical tetraplegic population. 17 Although the mechanisms underlying this phenomenon remain unclear, it is possible that an SCI-induced alteration in nocturnal melatonin release, 30 which is known to closely resemble that of core body temperature, 31 may contribute to impaired diurnal rhythmicity in core body temperature.
AD
Using our novel pattern recognition algorithm to examine spontaneously occurring AD, we extend the findings of Rabchevsky and coworkers 5 and Zhang and coworkers 11 by investigating the pressor response, maximum SBP, average SBP, minimum HR, average HR, duration of the AD events, and time of day at which the spontaneous AD events occurred. In agreement with previous reports, we found that rodents with high-thoracic SCI develop AD as early as 2–4 days after SCI. During this acute period, the average SBP response is relatively weak, but does exceed 20 mm Hg, which is the clinical definition of AD. 7 Although AD is typically considered a chronic effect, it has been reported clinically in the acute period, and our data corroborated these findings. 6,9 It is proposed that loss of descending sympathetic inhibition is the primary mechanism responsible for AD during the acute period. 9
Between days 6 and 12 post-SCI, animals exhibited a gradual reduction in the frequency of spontaneously occurring AD events and a significantly reduced pressor response during AD. We believe that this reduction in AD incidence and severity during the subacute phase was the result of transient plastic changes within the sympathetic preganglionic neurones, including atrophy and diminished dendritic arborization, that form a crucial part of the AD circuitry. 32 From day 14 onwards, the frequency of AD significantly increased compared with the acute and subacute phases; however, it was not until 3–4 weeks post SCI that the AD pressor response fully developed. This time course for the development of AD is largely in agreement with previous studies that have examined spontaneously occurring AD events in mouse and rat models of SCI. 5,11 Interestingly, day 14 onwards is also the time at which we had previously found that sympathetic preganglionic neurons began to re-establish their dendritic arbor after an initial atrophy, and the point at which local, potentially aberrant, cord reorganization took place. 33 Such arborization/cord reorganization is presumably a gradual ongoing process; however, our data suggest that at 14 days post-SCI there is a sudden (rather than gradual) increase in the incidence and severity of AD. We believe this may reflect a situation in which arborization/reorganization has to reach a “threshold” level before the AD reflex can function. This sudden increase in the incidence and severity of AD may also reflect an increased sensitivity of the peripheral vasculature, which has been previously reported at day 14 post-SCI. 34
By 1 month post-injury, rodents exhibited frequent and severe episodes of spontaneously occurring AD, a finding that is broadly in agreement with the only other study to investigate spontaneously occurring AD in experimental SCI. 5 Finally, an increased incidence and severity of AD during the chronic post-SCI period may be in part caused by the discontinuous of manual bladder expression. From 10–12 days post-SCI onwards, rats regained reflexive micturition, and hence no longer required manual bladder expression. Given that a full bladder is one of the most potent stimulants of the AD reflex in humans, 35 it is possible that regular bladder filling and emptying was at least partially responsible for the increased severity and incidence of spontaneous AD in chronic SCI. To the authors' knowledge, a similar study has yet to be conducted in humans, but is urgently required to assist in the management of AD among patients with SCI. The sheer number of AD events we found in animals with SCI is particularly worrying given the life-threatening nature of AD. 35,36 Further, these frequent AD events in the face of persistent hypotension could cause potentially deleterious swinging of arterial BP that could damage the cardiovascular organs, particularly the peripheral and cerebral vasculature and the heart.
The spontaneous AD detection algorithm used in the current study has important differences compared with precedent algorithms. The main difference is that the precedent algorithms averaged MAP and HR indices into 2 sec or 5 sec epochs, whereas we used beat-by-beat BP values. We believe our approach to be superior, because averaging hemodynamic data will likely underestimate the true physiological severity of the pressor response during AD. The absolute value of SBP reached during AD has been shown to be clinically important, because episodes of AD that have an accompanying SBP that exceeds 180 mm Hg are associated with life-threatening events such as hemorrhage and cerebral ischemia/infarction.
35
Further differences between the current and precedent algorithms include a minimum AD duration clause of 10 sec and an HR drop of ≥40 bpm from baseline during the final 75% of the duration of the AD event (see
The spontaneously occurring AD profile at all time points post-injury appeared visually different from that during CRD-induced AD episodes. For example, spontaneous episodes of AD tended to exhibit a slower kinetic response that peaked and then began to recover almost immediately (Fig 1). This is in contrast to CRD-induced AD whereby there is a rapid and persistent increase in BP that is accompanied by bradycardia (Fig. 5). Interestingly, this is similar to the observations of Rabchevsky and coworkers, 5 who also were unable to find any spontaneously occurring AD events that were associated with prolonged increases in BP and bradycardia. Although it is unclear what stimulus was responsible for the spontaneously occurring AD events that we recorded, it is likely that these occurred during spontaneous bladder or bowel emptying. Following experimental SCI, reflex bladder emptying returns at ∼10 days post-SCI. 37 During reflex emptying, it is likely that any AD stimulus will pass as soon as the bladder/bowel is empty. This is in contrast to CRD-induced AD in which pressure is maintained in the colon until the balloon is deflated. Such an induced stimulus is likely to be more potent, and may explain the difference in appearance of spontaneous and induced AD events. Despite this observation, we found a remarkable similarity between the maximal pressor to spontaneously occurring and induced AD at each time point post-SCI, suggesting that CRD (at least with 2 mL of air inflation) is a relevant stimulus for examining AD severity in rodents with SCI when the assessment of spontaneous AD events via telemetry is not available.
Conclusion
Our study is the first to concurrently examine resting hemodynamics, thermoregulation, CRD-evoked AD, and spontaneously occurring AD events across time post-SCI. We found that the acute period post-SCI is characterized by hypotension, hypothermia, and pronounced disruption to diurnal rhythms. Using a novel “AD detection algorithm,” we also found that AD is present as early as 2 days post-injury, although this seems to subside from days 6–12 post SCI. Day 14 post-injury appears to represent a major changing point for cardiovascular function. Diurnal rhythms have recovered and temperature control is improved, albeit to an “altered” phenotype compared with pre-SCI. Perhaps the most remarkable change at day 14 post-SCI, however, is that the number and severity of spontaneously occurring AD events is significantly increased almost twofold, and the pressor response to CRD-evoked AD is 100% higher than during acute SCI.
Footnotes
Acknowledgments
This work was supported by grants from the Canadian Institute for Health Research and the Craig H. Neilsen foundation. Research in the laboratory of A.V. K. is supported by the Canadian Foundation for Innovation, BC Knowledge Translation Foundation, Canadian Institute for Health Research, and Craig Neilsen Foundation. C. R. W. is the recipient of Craig Neilsen Foundation and Michael Smith Foundation postdoctoral fellowships.
Author Disclosure Statement
No competing financial interests exist.