
Abstract
The purpose of this study was to examine neuronal activity levels in the hindlimb area of motor cortex following spinal cord injury (SCI) in rats and compare the results with measurements in normal rats. Fifteen male Fischer-344 rats received a 200 Kdyn contusion injury in the thoracic cord at level T9–T10. After a minimum of 4 weeks following SCI, intracortical microstimulation (ICMS) and single-unit recording techniques were used in both the forelimb and hindlimb motor areas (FLA, HLA) under ketamine anesthesia. Although movements could be evoked using ICMS in the forelimb area with relatively low current levels, no movements or electromyographical responses could be evoked from ICMS in the HLA in any of the injured rats. During the same procedure, electrophysiological recordings were obtained with a single-shank, 16-channel Michigan probe (Neuronexus) to monitor activity. Neural spikes were discriminated using principle component analysis. Neural activity (action potentials) was collected and digitized for a duration of 5 min. Despite the inability to evoke movement from stimulation of cortex, robust single-unit activity could be recorded reliably from hindlimb motor cortex in SCI rats. Activity in the motor cortex of SCI rats was significantly higher compared with uninjured rats, and increased in hindlimb and forelimb motor cortex by similar amounts. These results demonstrate that in a rat model of thoracic SCI, an increase in single-unit cortical activity can be reliably recorded for several weeks post-injury.
Introduction
S
Further, scaled cortical activity encoding temporal intervals has been shown to occur even after spinal cord transection in rats. 9 Should a closed-loop, brain-computer spinal cord interface device be feasible in SCI patients, activity levels in the intact or the de-efferented cortex might be used to bypass the disruption of the corticospinal system. Thus, information regarding changes in spontaneous firing rates as a result of SCI will need to be incorporated in decoding algorithms.
Clinical studies confirm that brain networks involved in motor control (both M1 and secondary motor and somatosensory areas) remain active and responsive, even in chronic paralysis. 10 This preserved brain activity is associated with attempts to move, or motor imagery 11 and has been harnessed in therapeutic strategies for restoring motor function using brain-computer interfaces. 2
Rodent models of SCI differ substantially from clinical injuries due to differences between rodents and primates in the anatomy of descending spinal cord pathways as well as the degree of overlap in somatosensory and motor representations. 12 –14 Thus, it is important to determine how neuronal activity changes in rodent models after SCI. Previous studies have shown that after neonatal spinal cord transections, spontaneous, as well as stimulus-evoked single unit activity is significantly increased in the cortical somatomotor hindlimb area. 15 However, no studies have examined spontaneous activity levels of single neurons in cerebral cortex after SCI in adult animals.
The present study is novel in that it is the first to examine cortical neuronal action potentials in an adult rat model of spinal cord contusion injury. After at least 4 weeks post-injury, and under ketamine anesthesia, spontaneous action potentials (spikes) were recorded in the hindlimb and forelimb areas of motor cortex. Results in SCI rats were compared with those obtained in normal (uninjured) rats.
Methods
General experimental design
Twenty adult male, Fischer-344 inbred rats (Harlan Laboratories, Indianapolis, IN) were selected for this study. This strain of rat has been used extensively as a model subject in experiments examining SCI resulting in impairment of hindlimb movements. 16 –23 A total of 15 rats received a contusion injury in the thoracic cord at level T9–T10 (See details below; Fig. 1), whereas 5 rats were randomly selected as uninjured controls. Behavioral performance was assessed in 11 of the SCI rats using the Basso, Beattie, and Bresnahan (BBB) scale. 24 After a minimum of 4 weeks following SCI, ICMS and single-unit recording techniques were used in both the forelimb and hindlimb motor areas (FLA and HLA, respectively) of all 15 SCI and 5 normal rats to determine movements evoked by ICMS, current thresholds, and spike rates.
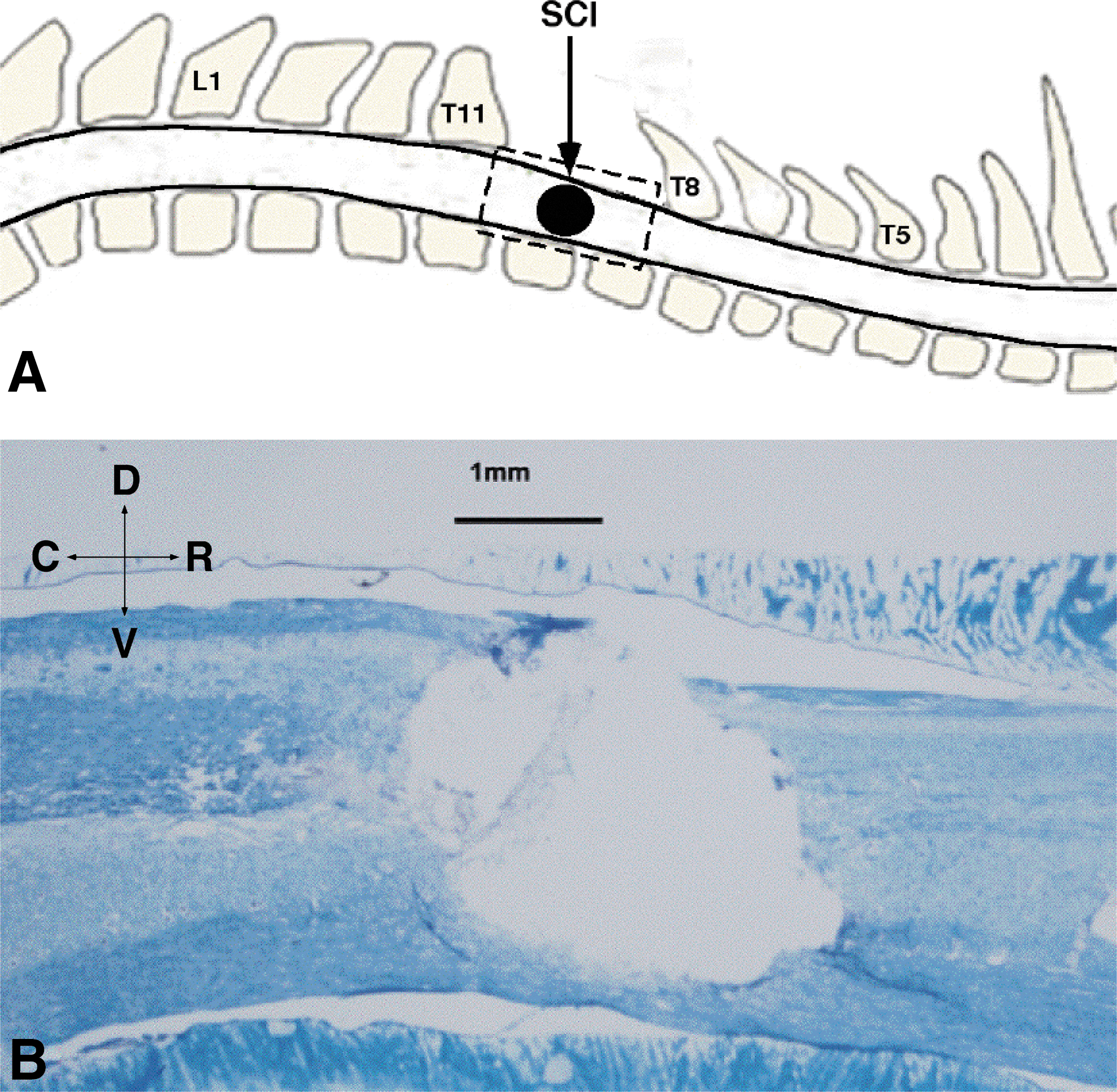
Spinal cord contusion injury in the thoracic spinal cord.
This study was performed in accordance with standards detailed in the Guide for the Care and Use of Laboratory Animals (Institute for Laboratory Animal Research, National Research Council, Washington, DC: National Academy Press, 2011). The protocol was approved by the University of Kansas Medical Center Institutional Animal Care and Use Committee.
Spinal cord injury
At the time of the spinal cord surgery, ages of the rats ranged from 100–110 days. Surgeries were performed under ketamine hydrochloride (80 mg/kg)/xylazine (7 mg/kg) anesthesia and aseptic conditions. Each of the SCI rats underwent a T9–T10 laminectomy and contusion injury using an Infinite Horizon spinal cord impactor (Precision Systems and Instrumentation, LLC, Fairfax Station, VA) with a 200 Kdyn impact (Fig.1). Displacement distance reported by the impactor software for each contusion was recorded. At the conclusion of surgery, 0.25% bupivacaine hydrochloride was applied locally to the incision site. Buprenex™ (0.01mg/kg, SC) was injected immediately after surgery and 1 day later. All animals were monitored daily until the end of the experiment. On the first week after surgery, the rats received daily injections of 30,000 U penicillin (Combi-Pen 48™, SC) in 5 mL saline to prevent infections and dehydration. The rats' bladders were expressed twice daily until the animals recovered urinary reflexes. From the second week onward, the animals were supplemented with vitamin C pellets (BioServ, Frenchtown, NJ) to avert urinary tract infection.
Post-SCI behavioral assessment
BBB scoring was performed on 11 rats 4 weeks after SCI as described by other invesigators. 24,25 This time-point, and the time of the neurophysiological procedures, was chosen because scores typically plateau by4 weeks and behavior is relatively stable. 23,26 A straight alley with a darkened goal box was utilized for the testing. For 1 week prior to BBB scoring, rats were habituated to the apparatus. Left- and right-side scores were averaged and rounded to the nearest integer.
Neurophysiological procedures
Cortical exposure
Surgical and neurophysiological procedures were conducted under aseptic conditions at various time-points up to 18 weeks post-SCI. At the time of these procedures, ages ranged from 4.5 to 7.5 months and body weights ranged from 250 g to 476 g (mean = 368 ± 57 g standard deviation [SD]). After an initial, stable anesthetic state was reached using isoflurane anesthesia, isoflurane was withdrawn and the first dose of ketamine hydrochloride (100 mg/kg)/xylazine (5 mg/kg) was administered intraperitoneal (IP). The rats were placed in a Kopf small-animal stereotaxic instrument (David Kopf Instruments®, Tujunga, CA) and the incisor bar was adjusted until the heights of lambda and bregma skull points were equal (flat skull position). The cisterna magna was punctured at the base of the skull to reduce edema during mapping. A craniectomy was performed over the motor cortex. The general location of the craniectomy was based on previous motor mapping studies in the rat (Fig. 2A). 27,28 This stereotaxic location matches that reported by others for the somatomotor hindlimb area. 13,29 The dura over the cranial opening was incised and the opening was filled with warm, medical grade, sterile silicone oil (50% Medical Silicone Fluid 12,500, 50% MDM Silicone Fluid 1000, Applied Silicone Corp., Santa Paula, CA).

ICMS methods.
ICMS mapping procedures
For each rat, standard ICMS techniques for mapping the representations of movements in motor cortex were used (Fig. 2), as reported in previous publications. 27,28,30,31 Throughout the experimental procedure, core temperature and vital signs were monitored. Core temperature was maintained within normal physiological limits using a feedback-controlled heating pad during the entire procedure. Care was taken to maintain a stable anesthetic state. The anesthetic level was assessed by monitoring the respiratory and heart rate, and reflexes to noxious stimuli. The pinch withdrawal reflex and corneal reflex was conducted every 15–30 min. Additionally, a sudden increase in respiratory rate or an increase in heart rate (>285 beats/min) indicated the need to test for pinch and corneal reflexes. The presence of the withdrawal reflex to a pinch of the forelimb or hindlimb or the presence of a blink response to corneal stimulation indicated the need for an additional dose of ketamine. ICMS mapping procedures were conducted only during periods of stable anesthesia and were halted during occasional periods of shallow (excessive muscle tone in the limbs and rapid heart rate) or deeper anesthesia (marked by unusually high ICMS thresholds). A single bolus of ketamine hydrochloride (10 mg intramuscularly) was administered when an increased heart and respiration rate, and positive reflexes to noxious stimuli warranted a need for a deeper anesthetic state. Each injection was counted as an additional anesthetic dose.
A magnified digital photograph of the cortical surface was obtained through a surgical microscope and displayed on a computer monitor. A 250 μm grid pattern was placed over the image to indicate intended locations for microelectrode penetrations. The precise location of the microelectrode placement in the cortex was based on stereotaxic coordinates relative to bregma, derived in normal rats in a previous study to define the location of the hindlimb area in this strain (Fig. 2A). 28 The center of the hindlimb region in this rat strain is located at 2 mm posterior and 2.64 mm lateral to bregma. 28 ICMS was conducted at this and surrounding locations in each rat. The stimulating microelectrode consisted of a platinum wire inserted into a pulled glass micropipette filled with 3.5 M NaCl solution and sharply beveled to 15–20 μm at the tip. Microelectrode impedance ranged from 750 kΩ to 1 MΩ. Microelectrodes were advanced perpendicular to the cortical surface to a depth of ∼1700 μm (the approximate location of layer 5 in this species), using a hydraulic microdrive (David Kopf Instruments®, Tujunga, CA). The stimulus consisted of 13, 200 μs cathodal pulses delivered at 350 Hz repeated at 1/sec from an electrically isolated, charge-balanced (capacitively coupled) stimulation circuit. The electrode was introduced sequentially at interpenetration distances of 250–500 μm using the grid pattern sited with reference to the surface vasculature.
Movement fields were defined by movements evoked by ICMS at near-threshold current levels (60 μA maximum). Forelimb and hindlimb movements were evoked almost exclusively in the contralateral side of the body. The HLA was explored as completely as possible. Microelectrode penetrations (avoiding blood vessels) were made at intersections on the grid pattern until the HLA was circumscribed by sites evoking either forelimb movements, trunk movements, or no visible response. To limit the duration of the procedures, ICMS mapping in FLA was confined to the caudal portion.
Spike recording
In each rat, neuronal recordings were begun at ∼3 h after initiation of the procedure to control for effects of anesthetic duration on subsequent spike recordings. Anesthetic levels were maintained using the same procedure described above. Neuronal action potentials (spikes) were recorded with a single-shank, 16-channel Michigan-style linear microelectrode probe (Neuronexus, Ann Arbor, MI; Fig. 3). The same probe was used for each rat. Each of the 16 sites on the probe was15 μm in diameter and 50 μm apart, with an impedance of ∼0.9 MΩ. One site (site 7) was defective and was not used in the analysis of the results. To analyze mean spike rate per penetration as a function of cortical depth, five recording sites each were combined for superficial (SF), middle, and deep recordings, respectively. Sites 9, 8, 10, 13, and 4 were defined as superficial sites, sites 12, 5, 15, 2, and 16 were defined as middle sites, and sites 1, 14, 3, 11, and 6 were defined as deep sites (Fig. 3B). The tip of the probe was lowered to a depth of ∼1720 μm below the cortical surface and recordings began after a 1 min resting period. Activity was recorded at penetration locations in both HLA and FLA. The mean number of penetrations per rat used in the analysis of results was 9.5 (SD = 1.8); the mean number of penetrations was 6 (SD = 0.9) in the HLA and 4 (SD = 1.3) in the FLA.

Activity recording.
Because no hindlimb responses were evoked using ICMS in SCI rats, the location of HLA in SCI rats was determined by the stereotaxic coordinates derived in normal rats in a previous study (centered at 2 mm posterior and 2.64 mm lateral to bregma), 28 as well as the expected location based on the position of the ICMS-derived FLA. At each cortical location, electrical activity was collected and digitized for 5 min from each of the 15 active sites using neurophysiological recording and analysis equipment (Tucker Davis Technologies, Alachua, FL). Neural spikes were discriminated using principle component analysis. 32 Sample waveforms (1.3 msec in duration) were collected that passed −5.5 × SD below root mean square (RMS). After each experiment, the probe was cleaned with Opti-Free solution (Alcon Laboratories, Fort Worth, TX), followed by ethanol and then rinsed thoroughly in distilled water. The electrode impedance of each site remained at ∼0.9 MΩ for each experiment. At the end of the recording session, rats were humanely euthanized with an overdose of sodium pentobarbital (Beuthanasia®-D, Schering- Plough Animal Health Corp., NJ).
Statistical analysis
The main a priori hypothesis to be tested was that spike rates would be higher in M1 after SCI. Spike rates over the 5 min sampling period for each of the 15 probe sites were averaged for each of the penetration locations. Then spike rates per penetration were calculated for each cortical area and used in the test of the main hypothesis using a two-way repeated measures analysis of variance (ANOVA), with Group (normal vs. SCI) and Area (HLA, FLA) as the main factors (alpha = 0.05). Additional secondary analyses were performed to determine whether spike rates were affected by time after SCI, and whether there were laminar differences in spike rates.
Results
Histology
As reported in a previous study, histology shows that only 9% of the spinal white matter is spared at the injury epicenter following this severe contusion injury. 23 BBB scores indicate that the impact at this level of the spinal cord left intact central pattern generators purportedly contributing to recovery.
Both the dorsal corticospinal and the rubrospinal tracts were damaged in all SCI cases, showing significant demyelination. However, there was some sparing of the small ventro-medial part of the corticospinal tract near the sulcus, which is the location of the ventral, uncrossed component of the rat corticospinal tract. 33 Damage to spinal cord tracts relevant to propriospinal projections as well as inputs to central pattern generators is evident. 34,35
BBB scores
The mean BBB score for 11 SCI rats at 4 weeks post-injury was 10.5 (standard error [SE] = 0.4), significantly different from normal BBB scores of 21 (t = 15.2, p < 0.0001). This level of severity at 4 weeks post-injury has been described as severe in a previous report. 23 Although BBB subscores were not derived, rats had persistent deficits in paw position, trunk stability, toe drag, and tail position.
Cortical representations of HLA and FLA
In uninjured (control) rats, ICMS evoked appropriate joint movements in both HLA and FLA (Fig. 2B). In SCI rats, movements evoked by ICMS in FLA were similar to those evoked in control rats. Forelimb movement thresholds averaged 33.8 μA (SE = 1.2) in SCI rats and 38.1 μA (SE = 2.6) in control rats (p > 0.05). However, hindlimb movements were not evoked in any of the SCI rats at the maximum current of 60 μA (Fig 2C).
Single-unit recordings
Robust single-unit activity was recorded reliably from both HLA and FLA in each rat (Fig. 4 and Fig. 5). Mean spike rates in M1 (combined HLA and FLA) of normal and SCI rats are shown in Figure 6. A wide variation in spike rates was observed among individual rats. Rates ranged from 0.40 (SE = 0.28) to 3.5 (SE = 1.0) spikes per second in normal rats and 0.32 (SE = 0.18) to 8.0 (SE = 2.9) spikes per second in SCI rats. Although normal rats tended to require more ketamine doses to maintain a surgical level of anesthesia (1.35 ± 0.65 SE doses per h on average for normal rats vs. 1.16 ± 0.67 SE for SCI rats), this difference was not significant (unpaired t test, p = 0.1291).
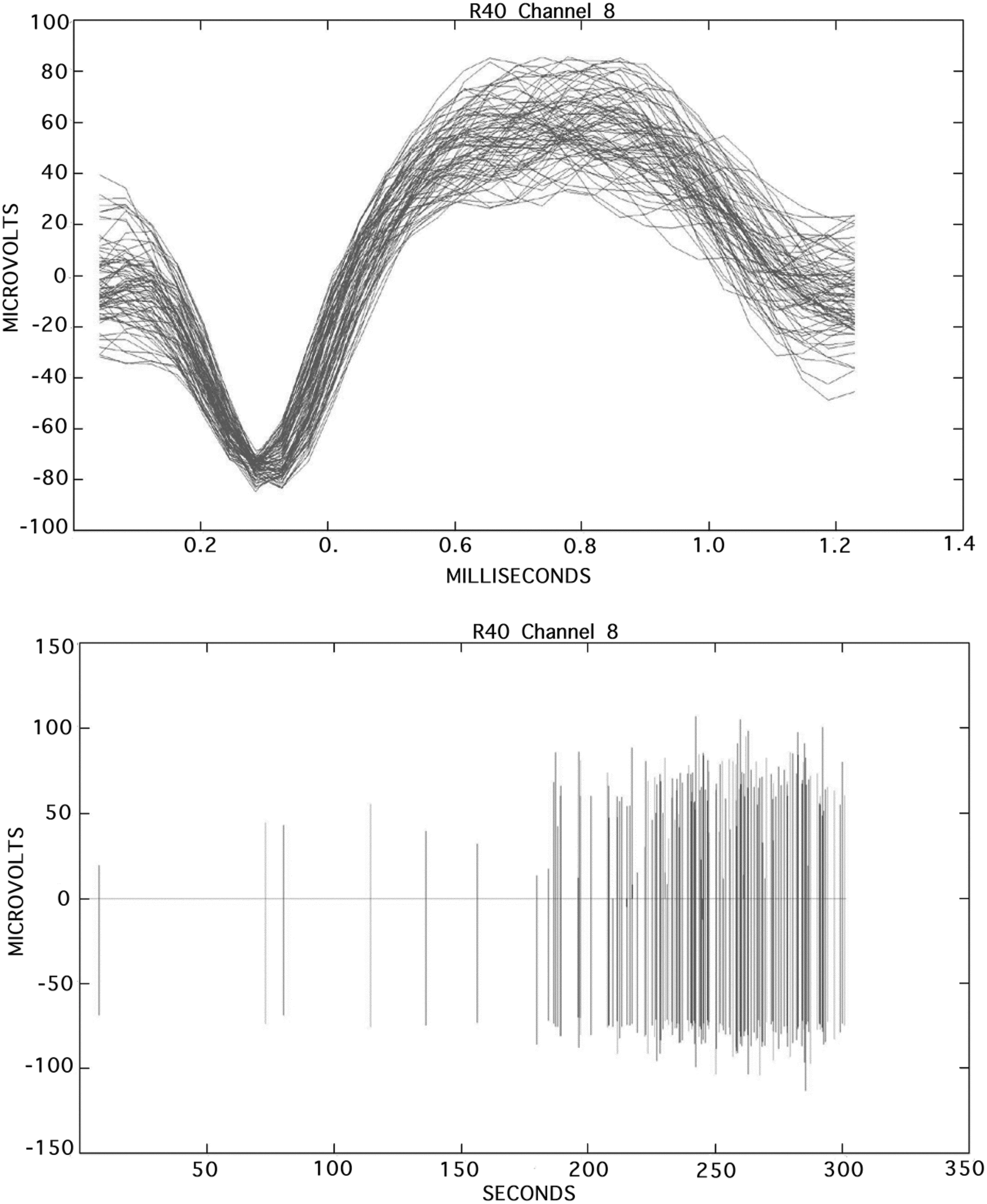
Action potentials (spikes) measured in the hindlimb motor area of a ketamine-anesthetized spinal cord-injured rat. Top: Overlaid subset of spiking activity profiles recorded from a single-channel dataset. Bottom: Spikes recorded from the same channel displayed over the entire time period of 300 sec.
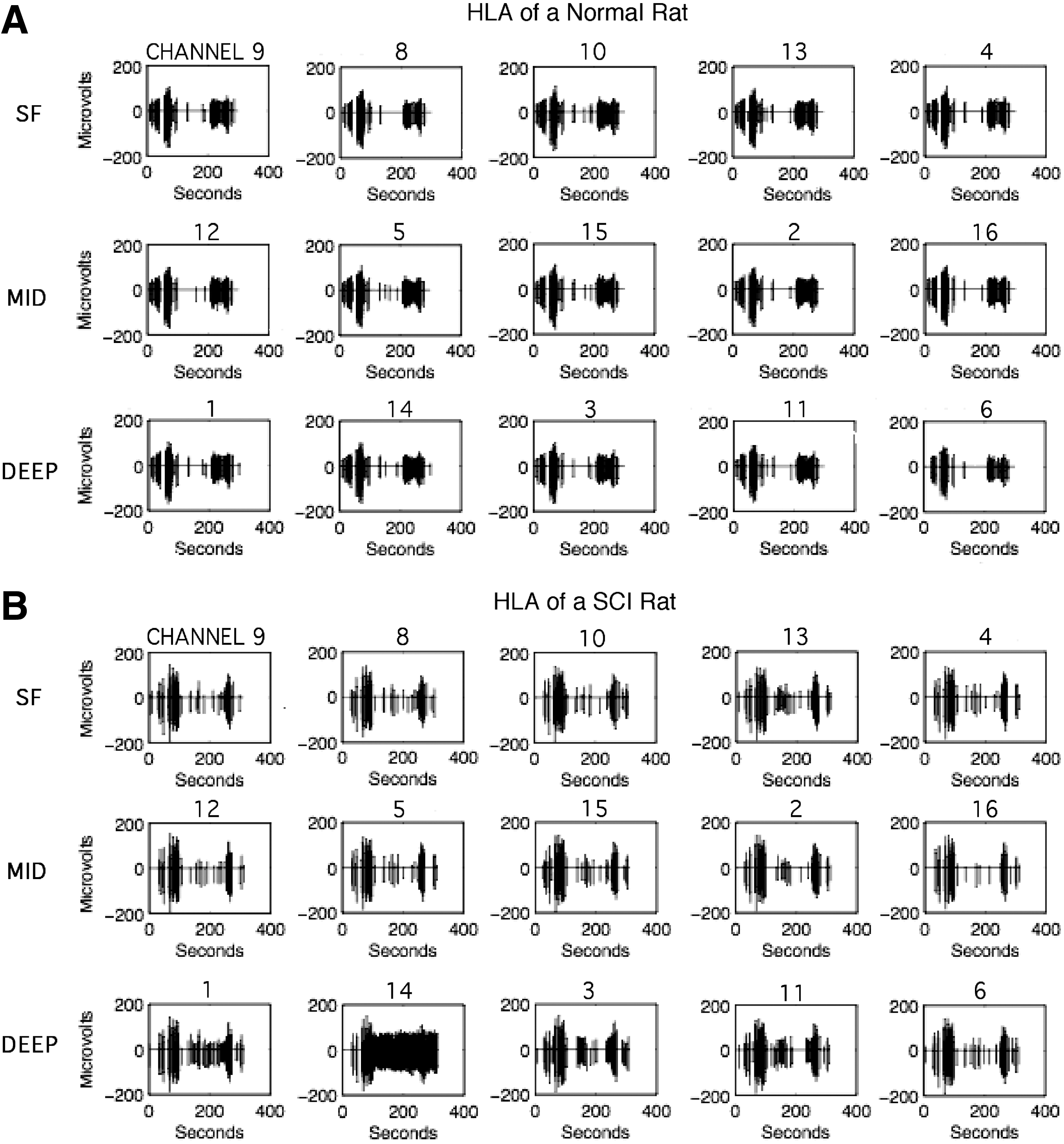
Typical spike recordings from each of the 16 channels in a single penetration in the HLA of a
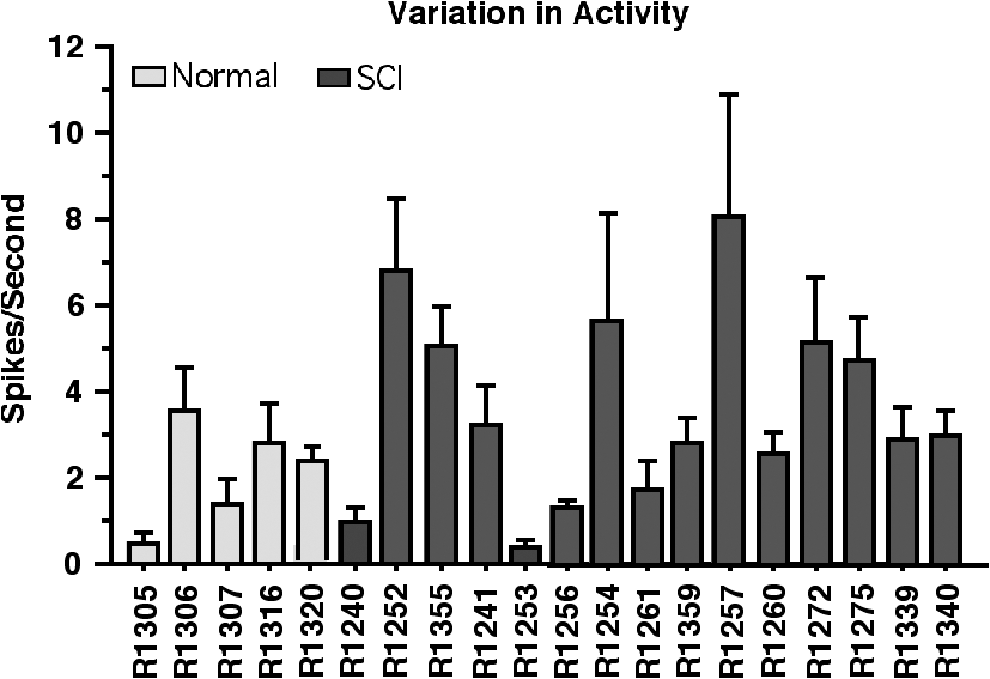
Average spontaneous spike activity in M1 (combined HLA and FLA) for the entire dataset of 16 channels at all penetration sites in each rat. Error bars are ± SE. FLA, forelimb motor area; HLA, hindlimb motor area; M1, primary motor cortex; SE, standard error.
Mean spike rates in HLA and FLA of normal and SCI rats are shown in Figure 7A. A two-way ANOVA revealed that there was a significant effect of SCI on spike rate across M1 (HLA + FLA) (F = 6.067; p = 0.024), with rates 75% higher in SCI rats compared with normal rats (M = 3.68, SE = 0.32 in SCI rats; M = 2.10, SE = 0.56 in normal rats). There was no significant effect of cortical area, though rates in FLA tended to be a bit higher than rates in HLA in both normal and SCI rats. Spike rates were significantly higher in the FLA of SCI rats than in the HLA of normal rats (p < 0.05). There was no significant Group X Area interaction in spike rates (F1,18 = 0.334; p = 0.571). In a post hoc comparison of change within areas, spike rates tended to increase in both HLA and FLA by similar amounts: spike rates in HLA of SCI rats were 71% higher than in the HLA of normal rats; rates in FLA of SCI rats were 78% higher than in the FLA of normal rats. Thus, the HLA/FLA ratios were similar (M = 1.20, SE = 0.63 in normal rats; M = 1.15, SE = 0.39 in SCI rats; Fig. 7B). Because effects were so similar in HLA and FLA, subsequent secondary analyses were performed after pooling data from FLA and HLA.
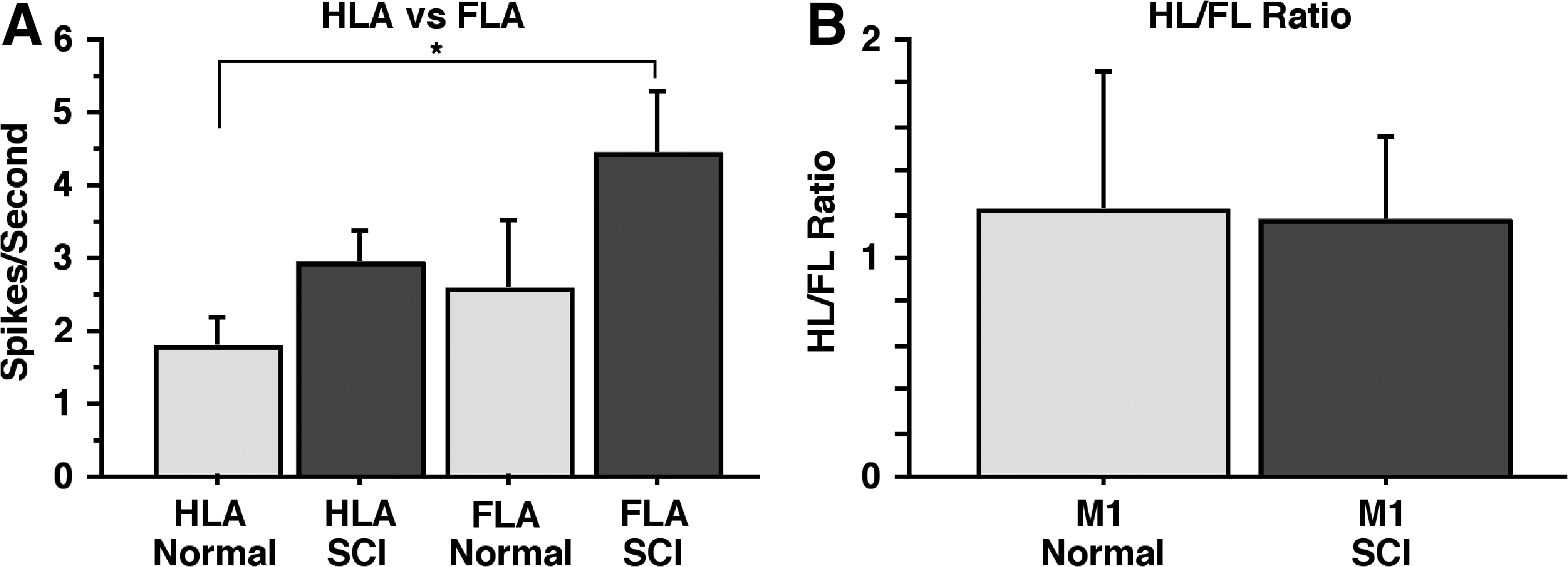
Average spontaneous spike activity in M1 in normal and SCI rats.
Secondary analyses revealed no significant difference in spike rates between rats studied at weeks 4–6 versus weeks 7–18 (F1,13 =1.232; p = 0.2872). Spike rates in M1 were 3.039 (SE = 0.763) spikes per sec at 4–6 weeks post-injury and 4.328 (SE = 0.934 spikes per sec at 7–18 weeks post-injury; Fig. 8).
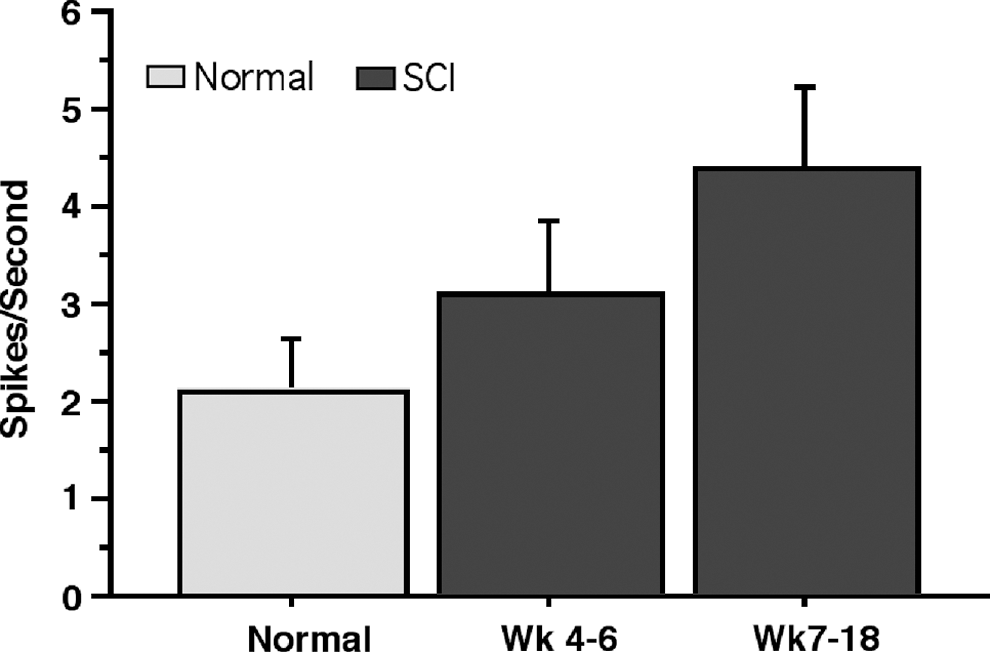
Average spontaneous spike activity in normal and SCI rats at 4–6 and 7–18 weeks post-injury. Normal: n = 5; week 4–6: n = 9; week 7–18: n = 6. Error bars are ± SE. SCI spinal cord injury; SE, standard error.
Finally, a two-way ANOVA revealed a significant difference in spike rate as a function of cortical depth (F2,36 = 18.204, p < 0.0001; Fig. 9). Tukey's honest significant difference (HSD) test showed that spike rates were consistently highest in deep layers in both normal and SCI rats (p < 0.05). Although not specifically tested statistically, differences in spike rates as a function of depth were similar in HLA and FLA.
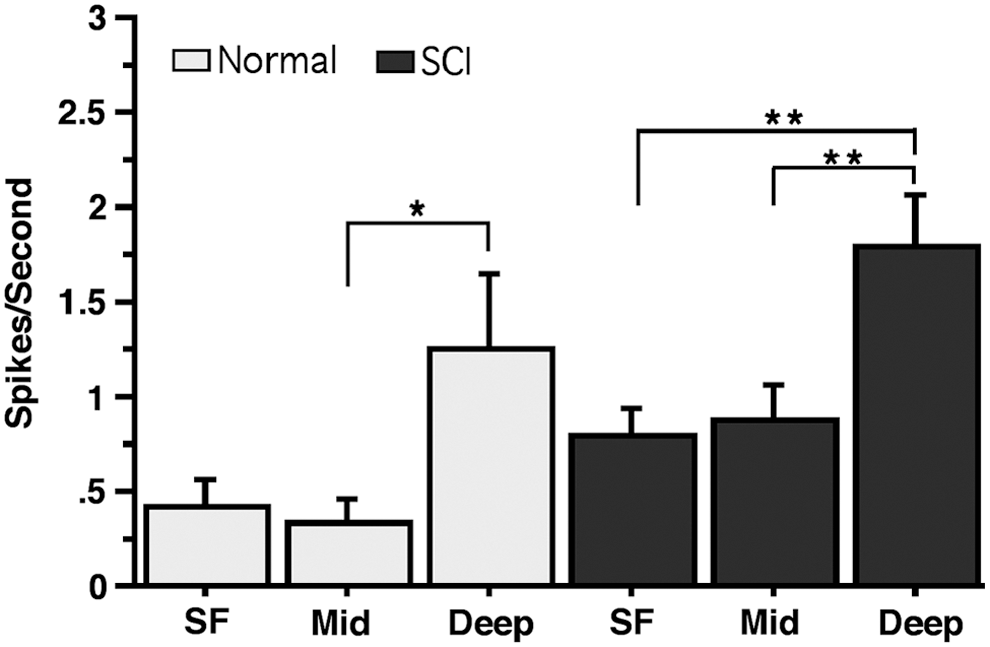
Average spontaneous spike activity in superficial (SF), middle (Mid), and deep electrode sites in normal and SCI rats. Error bars are ± SE. * = p < 0.05; ** = p < 0.0001. SCI spinal cord injury; SE, standard error.
Discussion
It is evident from the histology of the damaged spinal cord that the majority of the descending corticospinal projections have been damaged. A small sparing of the ventral, uncrossed component of the rat corticospinal tract is possible. Although some of the propriospinal tract may possibly be functioning, coordination is lacking, suggesting that the ascending propriospinal tract coordinating hindlimb/forelimb coordination is non-functional.
ICMS results showed normal FLA maps in SCI rats but a complete inability to evoke movement of the hindlimb at 60μA in the typical location for hindlimb motor cortex. These results show that in this model of SCI, the FLA cortex retains the efferent connections mediating voluntary forelimb movements in the cervical spinal cord but the hindlimb area is disconnected from motor neurons in the lumbar cord mediating hindlimb movements. In spite of this difference in the integrity of efferent connections, spontaneous single-unit activity could still be recorded in both FLA and HLA. Further, the ratio of spike activity in the HLA and FLA of SCI and normal rats was the same (Fig. 7B). This suggests that neuronal activity in the FLA increases to the same degree as activity in the HLA after thoracic SCI. Thus, greater activity in M1 (both HLA and FLA) of SCI rats is a more global result of SCI than just de-efferentation of localized regions.
The greater amount of activity in the FLA is reminiscent of the enhancement of responses in the forelimb somatosensory area evoked by forepaw stimulation immediately after thoracic spinal transection. 36 Whereas responses to stimuli were enhanced in the somatosensory area in the transection model, an overall more silent cortical spontaneous activity was reported. 36 The authors conclude that within minutes of SCI there is a change in the state of large cortical networks and that this change plays a critical role in early cortical reorganization after SCI. It is evident from the results reported here that activity in the FLA also changes after injury in the thoracic cord, in this case a general increase in spontaneous spiking weeks after injury, even though the efferent and afferent connections to the forelimbs remain intact.
A large amount of individual variation in activity levels was observed in both SCI and normal rats, a result that may reflect individual differences in level of anesthesia during the recording of activity. In spite of the attempt to limit variation in anesthesia levels during cortical activity recordings by matching the duration of anesthesia and the observable indicators of anesthetic level (heart rate, respiration rate, responses to noxious stimuli), individual variations in the response to ketamine are expected. It has been established that SCI rats require less urethane to maintain a stable level of anesthesia than normal rats 7–12 h after spinal cord transection. 37 Although they tended to need slightly fewer doses, SCI rats 4–18 weeks post-contusion injury in this study did not require significantly fewer doses of ketamine to maintain a stable level of anesthesia. Any difference in anesthetic dosing of individual rats may contribute to higher cortical activity during the recording session and could contribute to individual variation in recorded activity.
The increased levels of spontaneous cortical neuronal activity in deep (infragranular) layers of somatomotor cortex seen here in adult rats following spinal cord contusion injury is similar to that previously described in rats following neonatal transection of the spinal cord. 15 Although higher activity levels in deeper layers may be at least partially related to sampling bias due to the larger neuron size in layers 5 and 6 of cortex, 12 this enhanced activity may also suggest a mechanism for plasticity in the cortical networks altered by injury in the spinal cord.
Clinical studies examining brain activity levels in the cortex of SCI patients have found activations in cortical areas involved in motor control (primary and secondary motor cortex) during attempts to move, and during mental imagery of movement tasks. 11 Several studies of brain activation levels in SCI patients have reported an increase in activity in cortical motor areas of SCI patients above controls, as well as activation of subcortical areas not active in controls. 10 However, the results appear to be related to time after injury. In the early subacute phase after SCI, patients have reduced activity levels in M1 and greater activity levels in secondary motor areas when compared with controls. 38 In the later chronic phase, there is a progressive increase in M1 activity levels and decrease in secondary motor area activity levels in injured patients until activation is similar to that of controls. 38 In the results reported here, rats with SCI tended to have higher levels of activity in M1 at different time-points after SCI compared with normal rats; with 7–18 weeks post-injury levels tending to be higher than 4–6 weeks post-injury, although these results were not significantly different (Fig. 8). Although comparison of the human results with those presented here in anesthetized rats is not straightforward, it is clear that SCI results in a significant and time-dependent change in excitability in the motor cortex. The increase in spontaneous activity across M1 following thoracic SCI may demonstrate a measurement of plasticity and change in cortical networks in the chronic SCI condition that could play a role in cortical reorganization after SCI.
Footnotes
Acknowledgments
This work was supported by a generous gift from the Ronald D. Deffenbaugh Family Foundation and NIH/NINDS R37 NS030853. Core facility support was provided by the Kansas Intellectual and Developmental Disabilities Center (NICHD HD002528). The authors thank Eugene Gregory, Judith Meriwether, DVM, and Linda Eggimann for expert technical support.
Author Disclosure Statement
No competing financial interests exist.