
Abstract
Although the deleterious effects of primary blast on gas-filled organs are well accepted, the effect of blast-induced shock waves on the brain is less clear because of factors that complicate the interpretation of clinical and experimental data. Brain cell aggregate cultures are comprised of multiple differentiated brain cell types and were used to examine the effects of underwater blast. Suspensions of these cultures encased in dialysis tubing were exposed to explosive-generated underwater blasts of low (∼300 kPa), medium (∼2,700 kPa), or high (∼14,000 kPa) intensities and harvested at 1–28 days post-exposure. No changes in gross morphology were noted immediately or weeks after blast wave exposure, and no increases in either apoptotic (caspase-3) or necrotic (lactate dehydrogenase) cell death were observed. Changes in neuronal (neurofilament H, acetylcholinesterase, and choline acetyltransferase) and glial (glial fibrillary acidic protein, glutamine synthetase) endpoints did not occur. However, significant time- and pressure-related increases in Akt (protein kinase B) phosphorylation were noted, as well as declines in vascular endothelial growth factor levels, implicating pathways involved in cellular survival mechanisms. The free-floating nature of the aggregates during blast wave exposure, coupled with their highly hydrolyzed dialysis tubing containment, results in minimized boundary effects, thus enabling accurate assessment of brain cell response to a simplified shock-induced stress wave. This work shows that, at its simplest, blast-induced shock waves produce subtle changes in brain tissue. This study has mechanistic implications for the study of primary blast-induced traumatic brain injury and supports the thesis that underwater blast may cause subtle changes in the brains of submerged individuals.
Introduction
B
The contribution of primary blast to overall bTBI is controversial. 5 –11 Although this type of injury to gas-filled organs is well accepted, with auditory damage, blast lung, and gastrointestinal injuries being well documented as being caused by blast waves in unprotected individuals, 12,13 their effect on solid organs such as the brain is much less clear. The identification of well-documented clinical cases is difficult and rare, 14 –16 whereas the technical difficulties associated with simulating and then isolating primary blast in the laboratory are considerable, resulting in an inconsistent literature that is difficult to interpret. 17 –19
The mechanisms by which primary blast incurs damage to the brain are complex and require multi-disciplinary efforts to fully address. However, a basic question to resolve is the tolerance of brain cells to the mechanical stress relevant to blast exposures to the head. A simplified approach to this question has been utilized by several laboratories that have developed cell and tissue culture models in efforts to ascertain the effects of shock or blast wave exposure. 20 –33 Invariably, these studies utilize surface culture models, imposing undesirable boundary conditions that cause spurious shock reflections and stresses, thus rendering the conditions imparted by the shock insult not only unrealistic, but also extremely difficult to measure or model.
Brain cell aggregate cultures are composed of the major brain cell types in ratios that are similar to that of whole brain. 34 –36 These cultures can be maintained for periods of several weeks, gain differentiative function, and, importantly, are suspension cultures that can be exposed to blast waves under conditions free of nearby reflective boundaries. Using this cell culture model, studies have been designed to subject brain cells to underwater shock of intensities beyond levels possible in conventional shock tubes. The underwater shock test scenario was chosen not only because a fluid-borne stress wave is ultimately imparted to brain cells in an air-blast exposure of the head, but also because there are negligible global acceleration effects, which can confound injury interpretations in certain air-blast experiments. This is a major difficulty, especially where small animal models are used without consideration of scaling. 17,37 In addition, underwater shock exposure has relevance to the assessment of TBI risk of divers from underwater explosive (UNDEX) events. There is considerable clinical evidence for central nervous system (CNS) disruption attributed to underwater shock of the head dating from World War II (WWII), of exposures of shipwrecked sailors left in the water after a sinking. 38 –41 More recent studies have considered the problem of military divers subjected to explosive attack, recreational or commercial divers accidentally exposed to UNDEX, as well as UNDEX effects on marine mammals. 42,43 Of particular interest in the clinical studies from WWII are consistent references to “psychogenic changes” 38 as well as “…persistent headaches…and definite departure from stable personality” 40 symptomatic of airblast mild TBI problems.
This report describes work that has been deliberately simplified in approach and utilizes a brain cell suspension culture model that is free of boundary reflections to examine the effect of fluid-borne shock waves. The experimental setup used in these studies, that of high-pressure shock-induced stress on isolated brain tissue, does not exist in practical scenarios. In reality, the diffraction of an air- or water-borne shock wave over the head will impart complex three-dimensional (3D) principal and shear stresses in the brain. However, a core question to resolve is the response of brain tissue to a simple, well-defined principal stress wave as induced by a shock wave exposure. The results of this study have mechanistic implications for the study of primary blast-induced TBI, as well as for the effects of underwater blast on submerged individuals.
Methods
Materials
Basal medium Eagle's (BME), fetal calf serum (FCS), glutamine, penicillin-streptomycin, and phosphate-buffered saline (PBS) were purchased from Invitrogen (Burlington, Ontario, Canada). Routine culture medium consisted of BME supplemented to 10% FCS (v/v), penicillin-streptomycin (10 IU/mL–10 μg/mL),
Rat brain cell aggregate culture
In conducting this research, the authors adhered to the “Guide to the Care and Use of Experimental Animals” and “The Ethics of Animal Experimentation” published by the Canadian Council on Animal Care. Timed pregnant Sprague-Dawley rats were acquired from Charles River Laboratories (St. Constant, Quebec, Canada). At 18 days of gestation, animals were anesthetized in a chamber containing isoflurane and the embryos isolated. Brains were removed into a culture dish containing ice-cold dissection buffer. Brain aggregate cultures were initiated using cerebral hemispheres with the meninges removed. Culture preparation followed established methods. 34 –36 Briefly, the dissociation process consisted of using a Pasteur pipette to gently triturate pooled brain hemispheres in ice-cold culture medium (10% FCS), followed by sieving through 70-μm nylon mesh. Cell suspensions were centrifuged at 300g for 15 min at 4°C and the resultant pellets combined into one 50-mL centrifuge tube. Cells were resuspended using trituration in cold culture medium, and an aliquot was taken for counting viable cells using trypan blue exclusion. Cells were again centrifuged, as before, and the resultant cell pellet gently resuspended in ice-cold culture medium supplemented up to 15% FCS. Spinner flasks (125-mL) were routinely inoculated with cells isolated from the embryos from 1 animal ∼2.0–2.5 × 108 in 80-mL of culture medium (15% FCS). The spinner flasks were immediately placed on stirrers (60 rpm) in a humidified 37°C incubator at 7% CO2. At 2 h in culture, 20 mL of culture medium (5% FCS) were added and the rotational speed increased to 70 rpm. At 24 h in culture, an additional 25 mL of culture medium (5% FCS) were added and the rotational speed increased to 80 rpm. Cultures were thereafter checked daily, and the rotational speeds of the impellers adjusted to ensure a gradual increase in aggregate size to a target diameter of ∼300–450 μm. Cultures were initially refed with 60 mL of fresh medium (10% FCS) three times per week, which increased to every second day at around 10 days in culture, when up to 90 mL of medium were replaced at each refeeding. This was required because of the increased cell proliferation that took place for the next 7–10 days. The three/week refeeding schedule was recommenced for the last week before and after blast treatment. Aggregate cultures were exposed to underwater blast at 27–28 days in vitro.
Blast exposure
On the day of exposure, aggregate cultures were transported to the experimental facilities next to the Underwater Explosive (UNDEX) pond (Fig. 1) and placed back into routine culture conditions in an incubator set at 7% CO2 and 37°C. This artificial pond is designed for the study of underwater explosion effects and has a diameter of 50 m and a maximum depth of 8 m at the center. It is shaped as an inverted, truncated cone, and holds over 7 million L of fresh water. Immediately before blast exposure, brain cell aggregates were isolated from the spinner flasks and transferred into dialysis tubing (BioDesign Dialysis Tubing, 1.13 inch wet diameter, 6.45 mL/cm volume; BioDesign Inc. of New York, Carmel, NY) that had previously been cut into ∼20-cm lengths and tied off with surgical thread at one end. These were autoclaved in PBS 24 h before use and remained in the PBS until utilized. After transfer of culture medium containing the aggregates, the tubing was tied off to form a sausage ∼10 cm long (Fig. 2A,B). This was placed into medium to prevent desiccation of the dialysis tubing and hand carried to the UNDEX pond where it was mounted to nylon fishing line using clips. The sausage was lowered into the water to a depth of ∼2 m in an upside-down position. After nonrequired personnel were removed to a defined safe distance, individuals trained in the safe use of explosives placed the explosive charge in the water at a distance and depth designed to provide the blast wave at the desired pressure. Once all personnel were located in safe zones, the countdown was initiated from 15 sec. At 7 sec, a solenoid was activated that released the bottom of the sausage, allowing it to return to an upright position before the explosive detonation at time zero. This caused the aggregates to be in suspension at the moment of blast wave arrival. After a standard wait time (∼2–4 min) to assess the safety of the explosive site, the sausage containing the brain cell aggregates was retrieved from the water and returned to the tissue culture facilities. The aggregates were transferred back into their spinner flasks and then transported back to the main laboratory facilities, where they were placed back into routine culture conditions until harvest and subsequent analyses.

UNDEX pond for underwater explosion research. The underwater explosive (UNDEX) pond has a diameter of 50 m and a maximum depth of 8 m at the center. It is shaped as an inverted, truncated cone and holds over 7 million L of fresh water. The building in the background (

Exposure of aggregate cultures to underwater blast. The aggregates are loaded into ∼10-cm-long cylinders of dialysis tubing secured on either end with surgical thread and the sausage is clipped to fishing line (
Explosive charge configurations and pressure measurements
Three charge configurations were used to obtain blast exposures at peak pressures of ∼300 kPa (low), ∼2700 kPa (medium), and ∼14,000 kPa (high). For the low-pressure configuration, a RP-83 Electric Bridgewire detonator containing ∼1.1 g of high explosive (primarily RDX) was used (Fig. 2C). For the medium- and high-pressure configurations, ∼100 g charges of C4 explosive (91% RDX) were used (Fig. 2D). The standoff distances between the charge and the gauges are shown in Table 1. The explosive charges were submerged to a depth of ∼1 m, whereas all pressure gauges and the brain cell aggregate sausage were submerged to a depth of ∼2 m.
The pressure of the underwater blast waves was measured using PCB Piezotronics model #138A01 sensors. These gauges have a usable measurement range of ∼0–6.895 MPa, a maximum pressure rating of ∼344.750 MPa, and a rise time of less than ∼1.5 μsec. They consist of a piezoelectric tourmaline crystal immersed in a nonconductive silicon-based fluid, housed inside a clear, flexible polyvinyl chloride plastic boot. A source follower amplifier is integrated into the gauge body. Before deploying the gauge, a coaxial microdot cable was connected to the gauge and the entire assembly was protected from water, electrical noise, and shock with Tygon tubing.
A schematic diagram of the experimental layout of the explosive charges, pressure gauges, and target brain cell aggregate culture samples is shown in Figure 3 (not to scale). The movable dock (6.57 × 3.99 m) is stationed at the center of the pond and is represented with the access hatch (2.89 × 2.15 m) in its midsection. Charges and gauges (PF1–4) were placed at pre-determined distances from the sausage containing the target brain cell aggregate cultures as defined in Table 1. Gauges PF-3and PF-4 were placed 10 cm to either side of the target at the same distance from the explosive charge in order to record the pressure at the target position. Gauges PF1–3 were deployed in their normal boot, whereas the fourth gauge (PF-4) was deployed inside the same dialysis tubing used to contain the brain cell aggregates, to confirm that the pressure was not affected by its presence (as compared to the pressure measured by gauge PF-3).
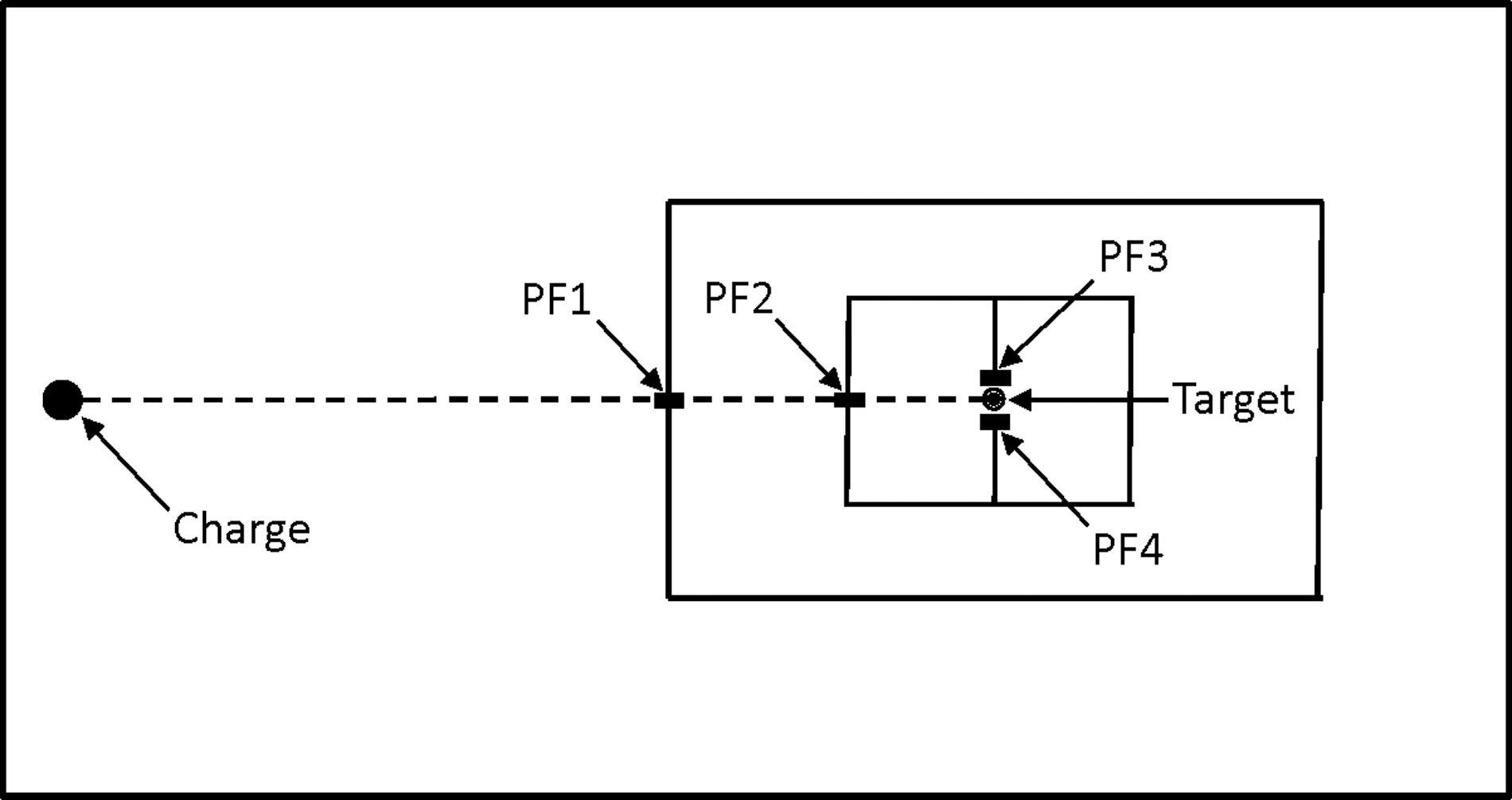
Schematic diagram of the experimental layout of the explosive charges, pressure gauges, and target brain cell aggregate culture samples (not to scale). The movable dock (6.57 × 3.9 m) is represented with the access hatch (2.89 × 2.15 m) in its midsection. Charges and gauges (PF1–4) were placed at pre-determined distances from the sausage containing the target brain cell aggregate cultures as defined in Table 1. Gauges PF-3 and PF-4 were placed 10 cm to either side of the target at the same distance from the explosive charge in order to record the pressure at the target position. Gauges PF1–3 were deployed in their normal boot, whereas the fourth gauge (PF-4) was deployed inside the same dialysis tubing used to contain the brain cell aggregates, to confirm that the pressure was not affected by its presence (as compared to the pressure measured by gauge PF-3).
Western blot analysis
Rat brain homogenate protein lysates (10–15 μg of protein) were separated on a 4–20% gradient pre-cast gel (Bio-Rad, Mississauga, Ontario, Canada) and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with 5% skim milk and 0.1% Tween 20 in PBS and then incubated with antibodies against glial fibrillary acidic protein (GFAP; 1:2000; Millipore, Etobicoke, Ontario, Canada), neurofilament H (NFH; 1:1000; Millipore), phospho-S473 protein kinase B (Akt) or total Akt (1:500; Cell Signaling Technology, Danvers, MA) overnight at 4°C. Membranes were then washed with PBS plus 0.1% Tween 20 three times at 10 min each. Primary antibodies were probed by incubating membranes with a secondary antibody, donkey anti-rabbit or mouse immunoglobulin G/horseradish peroxidase (GE Health Care Biosciences, Quebec, Canada) diluted 1:5000. Detection was carried out by using enhanced chemiluminescent advanced detection reagents (GE Health Care Biosciences) and imaged using a Molecular Imager VersaDoc MP 4000 system (Bio-Rad). To confirm equal protein loading, blots were reprobed with anti-pan-actin antibody (1:1000; Cell Signaling Technology). Band intensities were quantified using Quantity One software and expressed as a percentage of control. At least 5 samples were analyzed from each group and results are represented as mean ± standard error of the mean.
Biochemistry
Rat brain cell aggregate lysates were assessed for glutamine synthetase (GS), choline acetyltransferase (ChAT), and 2′,3′-cyclic nucleotide 3′-phosphohydrolase (CNPase) activity using spectrophotometric titerplate assays. GS activity was measured using a modification of the method of Gawronski and Benson, 44 whereas the method of Chao and Wolfgram 45 was adapted to measure ChAT. Acetylcholinesterase (AChE) activity was measured using the radiometric assay of Johnson and Russell. 46 Rat brain homogenate lysates were assayed for CNPase activity kinetically using a modification of the assay developed by Dreiling and Mattson. 47 Lactate dehydrogenase (LDH) was determined using an LDH kit (Sigma-Aldrich). Vascular endothelial growth factor (VEGF) levels were measured according to manufacturer instructions using components contained in the Bio-Plex Pro Rat Cytokine Assay 23-Plex, Group 1 kit (Bio-Rad).
Statistical analysis
Statistical differences were analyzed using two-way analysis of variance and Sidak's multiple comparisons test.
Results
Typical incident pressure histories for the three blast insult levels are shown in Figure 4A–C. The arrival of the blast wave produces a sharp rise in pressure that is immediately followed by an exponentially decaying tail. The onset of cavitation, which is local to the water surface, is shown as a negative phase spike downstream from the decaying tail. However, this spike is a gauge artifact, given that underwater piezo-electric gauges are not designed to measure the sudden pressure relief because of cavitation. Figure 4A–C also depicts the comparison of the pressure traces obtained from gauges mounted either in their normal boot configuration, or in the same type of dialysis tubing that the aggregates are contained within. With the exception of small differences between the pressure histories obtained by these gauge configurations at the low pressure exposure level, the traces are virtually identical, indicating that the dialysis tubing produces few boundary effects as the pressure wave progresses through the membrane. Figure 5 shows the bubble pulse histories that are encountered far downstream of the shock wave peak. Figure 5A highlights the detail of both the shock wave and the bubble pulse from the pressure history of a typical high (∼14,000 kPa) blast profile. Figure 5B–D shows typical bubble pulse pressure changes obtained from the low, medium, and high blasts used in these studies. The slow rise indicates that these pressure changes are not shock waves, with only the high blast producing an appreciable pressure increase in the bubble pulse, compared to the shock wave. The details of the pressure histories from the three target pressures (low, ∼300 kPa; medium, ∼2700 kPa; and high, ∼14,000 kPa) are shown in Table 2. Peak overpressures varied closely around the targeted pressures, with durations ranging from 117 to 427 μsec (Table 2). The calculated impulses for the three test overpressures ranged from 6.2 to 878.2 kPa-ms. The durations and impulses were determined based on an integration interval of 6 times the characteristic time scale of the exponential decay. 48

Representative blast wave pressure profiles from low (

Representative pressure histories of the bubble collapse from underwater blast exposures. The delayed bubble collapse pulse from a high blast exposure is shown in (
SD, standard deviation.
The aggregate cell cultures were exposed to underwater blast on day 27 or 28 of culture and then returned to routine culture conditions. Sampling from the spinner flasks was carried out on days 1, 3, 7, 14, and 28 post-exposure, when samples were either fixed for imaging, or cellular lysates prepared and frozen at −80°C until biochemical/molecular analysis. On the day of treatment, the aggregates were spherical with smooth surfaces. No debris and few or no single cells were present in the cultures. Preparation for, and exposure to, underwater blast exposure, during which time the cultures were outside of routine culture conditions, was generally less than 20 min. No obvious adverse effects attributed to this interval outside of optimal culture conditions, or of exposure to underwater blast, were observed microscopically. Exposure of the aggregates to underwater blast pressures up to and including ∼14,000 kPa did not have any obvious effects on morphology immediately after exposure or during the next 28 days. The aggregates retained their spherical shape, and no or little debris was noted (Fig. 6).

Light micrograph images of aggregates exposed to underwater blast. Aggregate cultures were exposed to either sham or high (∼14,000 kPa) underwater blast and then placed back into routine culture conditions. Images show aggregates at 14 days post-exposure. No visible differences are obvious between the two treatments. Color image is available online at
No signs of either apoptotic or necrotic cell death were noted at any time point (Fig. 7A,B). Sham control caspase-3 enzyme activity declined with time in culture, so that 28 days after blast exposure, the values were approximately one quarter of those at 1 and 3 days. However, exposure of the aggregates to underwater blast did not induce apoptotic cell death at any pressure, with no significant changes in caspase-3 activity compared to sham controls. Similar trends were observed with LDH release into the medium, with no pressure-related changes in LDH activity compared to sham controls. Although this enzyme activity is best used as an indicator of cell death when it can be standardized against cell number, total protein, DNA, or medium, this particular cell culture system does not allow this because of its suspension nature and the frequent medium changes. However, as a measurement of potential toxicity against similarly treated samples, it does provide a rough assessment by observing the change in absorbance at 350 nm compared to control samples. The lower LDH activities noted on day 1 are most likely attributed to the introduction of fresh culture medium after aggregate underwater blast exposure.
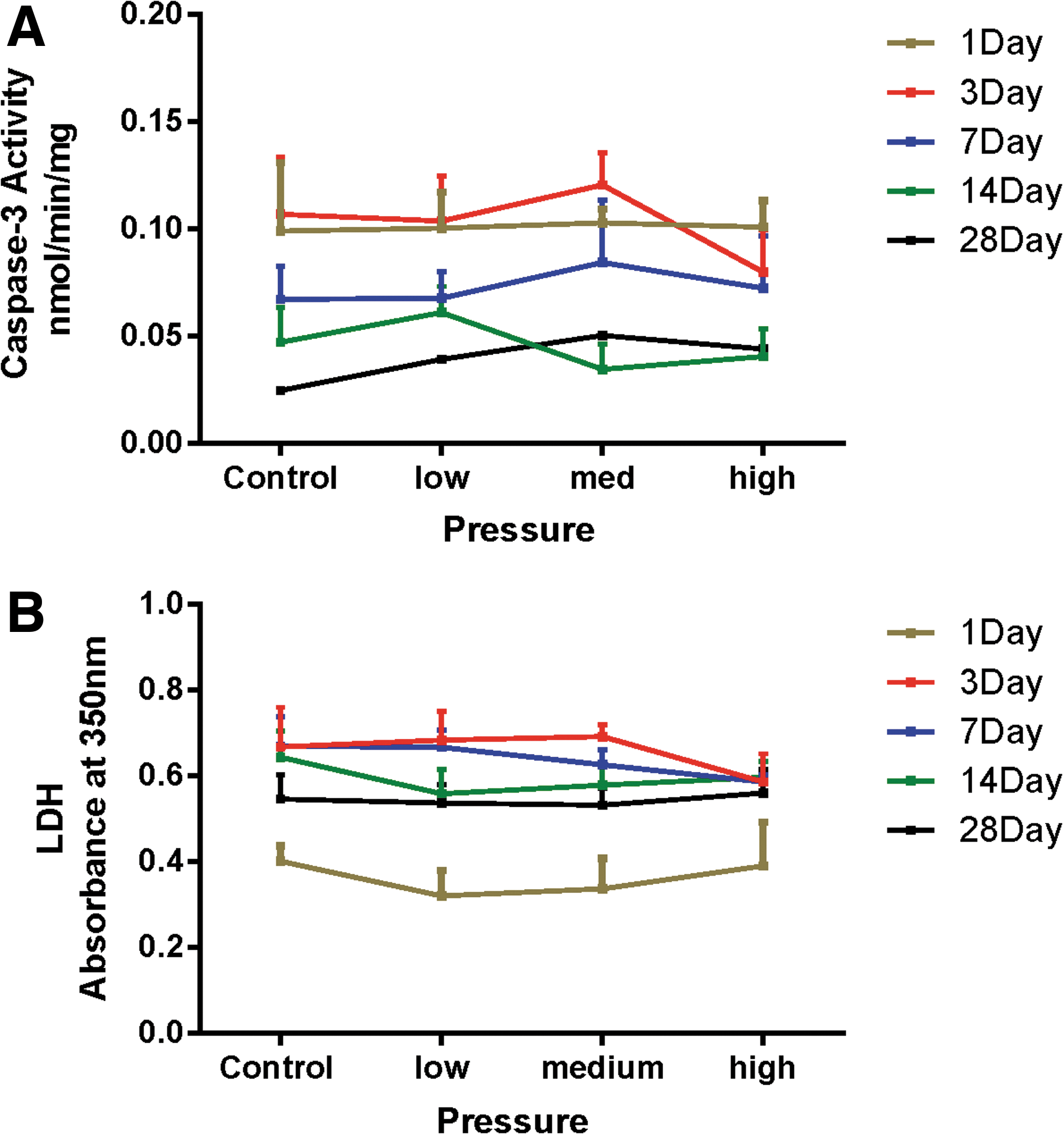
Effect of underwater blast on the viability of rat brain aggregates. Aggregate cultures were exposed to low, medium, or high levels of underwater blast and then placed back into culture. Cultures were assessed for either apoptotic cell death using caspase-3 activity (
No statistically significant changes in either the astrocytic marker, GFAP (Figs. 8A, B, D), or the neuronal marker, NFH (Figs. 8B–D), as a function of pressure or time were observed and levels remained at or near baseline. The effect of underwater blast on the activity of the neuronal enzyme markers, AChE and ChAT, are depicted in Figure 9 A,B. Sham control AChE activities decreased from day 1 to day 28 after blast (4.57 ± 0.43 vs. 2.88 ± 0.46 nmol/mg/min, mean ± standard deviation [SD]; n = 4). However, no significant pressure-related changes were observed at any time point, although there appeared to be a slight, but statistically insignificant, trend toward inhibition at the higher pressures. No appreciable change in sham control ChAT values were observed (9.78 ± 0.56 vs. 8.90 ± 0.85 nmol/mg/min; mean ± SD; n = 4) during the time course. Underwater blast produced no change at any time point, and all ChAT enzyme levels were closely associated with baseline values. Similar findings were obtained with the glial enzyme marker, GS (Fig. 9C). Sham control values were 48.7 ± 5.0 nmol/mg/min (day 1) compared to 38.6 ± 2.9 nmol/mg/min on day 28 (x ± SD; n = 4), whereas shock wave exposure did not induce any changes at any time points after exposure. The effects of underwater blast on the oligodendrocyte enzyme marker, CNPase, are depicted in Figure 9D. Sham values were 203.4 ± 67.0 nmol/mg/min on day 1 compared to 260.6 ± 87.8 nmol/mg/min on day 28 (x ± SD; n = 4). Underwater blast caused an increase in enzyme activity at 7 days, with high-pressure exposure causing a statistically significant elevation of ∼150% compared to sham control values.

Effect of underwater blast on brain aggregate GFAP and NFH levels. Aggregate cultures were exposed to low, medium, or high levels of underwater blast and then placed back into culture. Cultures were assessed for changes in GFAP (

Effect of underwater blast on brain aggregate AChE, ChAT, GS, and CNPase enzyme activities. Aggregate cultures were exposed to low, medium, or high levels of underwater blast and then placed back into culture. Cultures were assessed for changes in AChE (
Underwater blast provoked a pressure- and time related response of the Akt signal transduction pathway. At 1 day post-exposure, although total Akt was unchanged (Fig. 10B), phosphorylated Akt was elevated at all pressures (Fig. 10A), with statistically significant increases at both medium and high blast exposures. This response was attenuated by 3 days post-exposure, and by 7 days approached baseline. In contrast to the effects on the Akt pathway, blast exposure caused a slightly delayed inhibition of the protein, VEGF (Fig. 10C). Statistically significant decreases in VEGF expression were observed at 3 and 7 days post-exposure. However, at later time points, VEGF levels returned toward baseline levels.

Effect of underwater blast on the phosphorylation of brain aggregate Akt and on VEGF levels. Aggregate cultures were exposed to low, medium, or high levels of underwater blast and then placed back into culture. Cultures were assessed for changes in phosphorylated Akt (
Discussion
The study of PbTBI has been hampered by the technical challenges inherent in simulating primary blast in the laboratory and the necessity of employing a multi-disciplinary approach engaging experts from diverse fields of both the physical and biological sciences. Even once assembled, this expertise cannot properly address this research area without breaking down the problems to their elemental components. Current efforts have tended to focus on research studies where basic issues, such as the validation of the shock insult and the relevance of the shock exposure that the target actually experiences to survivable IED blast, are not adequately addressed. 7,17
A fundamental question that has yet to be answered is the response of brain cells to a simplified wave of principal stress that would be imparted from blast exposure, free of boundary effects and other complications. It is not currently obvious what shock wave properties are of importance in PbTBI, such as the rise time, peak amplitude, decay rate, and duration of the stress wave. Simulation and isolation of primary blast is difficult in the laboratory, and animal models, notably those using small rodents, are subject to scaling issues and resultant artifacts of dynamic pressure. The use of tissue culture models would appear to offer simplified platforms with which to subject brain tissue to defined blast waves and potentially overcome the scaling problems of dynamic pressure. Several laboratories have carried out such studies, either subjecting cultures to air blast 20 –27 or submerging them for underwater shock exposures. 28 –33 However, the use of surface cell and tissue cultures, multi-well culture plates, lidded culture vessels, wraps/bags to protect the cultures from outside contamination, and aquarium walls all impose numerous boundary effects, thus rendering the final pressure insult extremely complex and making its accurate quantitation extremely difficult or impossible. In these scenarios, it is not possible to expose the target cells to a defined shock insult of known parameters, nor is it possible to assess cause-effect relationships.
Aggregate brain cell cultures are dynamic, multicellular cultures that exist in suspension. They are 3D primary cultures comprised of the major cell types of the brain, with the exception of vasculature. Over the space of 3–4 weeks, and under close rotational velocity control of the impeller stirring the cell suspension, the seeded brain cells form clumps of cells that proliferate and differentiate into spherical cell aggregates (300–500 μm) exhibiting organotypic structures and function typical of the intact brain. This culture model system has been well characterized 34 –36 and has been used extensively to investigate diverse aspects of ischemia, 49 –53 inflammation, 54,55 and neurotoxicology. 56 –66 The use of these cultures offers several advantages in the study of shock and blast effects on the brain. Their multi-cell–type composition and differentiated functionality suggests that they will respond in a fashion more similar to that of whole brain, compared to single-cell–type cultures. They can be maintained for periods of 6 weeks or more after exposure, ensuring that potential persistent or delayed effects can be assessed. Last, and perhaps most important from a mechanistic point of view, these brain cell aggregates are suspension cultures, thereby facilitating experiments where the boundary effects imposed by surface culture can be further simplified. However, exposure of aggregates suspended in a solid vessel to air blast will still result in a complex pressure wave signature as the shock interacts with nearby boundaries causing multiple reflections. The experimental regimen was therefore further simplified so that the aggregates were exposed to underwater blast in a pond whose large size placed the target cultures far from reflecting boundaries, allowing for exposures to nearly planar, free-field underwater blast waves. In addition, the highly hydrolyzed dialysis tubing used to contain the aggregates in suspension presented no barrier to the shock front; gauges placed inside or outside of the dialysis tubing and next to one another showed virtually no differences in pressure histories as the shock front passed from the UNDEX pond water to the aqueous environment inside the tubing. Thus, measurement of the blast wave pressures outside of the dialysis tubing enclosing the aggregates was an accurate assessment of what they actually experienced. These experimental parameters enabled the direct exposure of brain cell aggregates to measureable, well-characterized underwater blast waves that exhibited realistic sharp rises and exponential decays free of reflecting boundaries and included a downstream bubble impulse. Although this was indeed the goal of this study, it is important to realize that this type of exposure does not represent a situation that is realistic. The almost total absence of boundary effects produces a shock exposure that produces only principal stress, or a crushing force within the exposed aggregates, and all but eliminates the shear forces and global accelerations that are also thought to be important in blast injury. Experiments using rats have shown that primary blast-induced skull flexure will cause complex 3D stress-fields, including principal and shear stress, 67,68 as well as exaggerated scaling artifacts of head movement. 69 Although studies are currently underway that introduce more complexity and realism into the experimental model system described here, the current tests were specifically designed to isolate the effect of principal stress without shear or global acceleration and investigate its potential biological effects.
The cultures were exposed to three pressures designated low (∼300 kPa), medium (∼2700 kPa), and high (∼14,000 kPa), with the low pressure representing a realistic threshold pressure unlikely to cause injury underwater and the two higher pressures likely representing injurious and lethal exposures, respectively, to a human. 43 These pressure exposures caused little or no change in most of the endpoints utilized to assess overall aggregate viability over the 28 days post-blast exposure. Markers of apoptotic (caspase-3) and necrotic cell death (LDH) were not elevated at any time after exposure, whereas common biomarkers of TBI attributed to impact-acceleration forces, such as glial GFAP and neuronal NFH, also did not change. Enzyme markers of neuronal (ChAT, AChE) and glial (GS) enzyme function did not deviate significantly from baseline sham control values, although CNPase, an oligodendrocyte marker, did show a slight pressure-dependent increase at 7 days that was maximal and statistically significant at the high pressure. However, no other significant effects or trends were noted at any time points for this enzyme.
In contrast to the limited effects that shock wave exposure produced on the commonly used cell-type–specific endpoints described above, blast was noted to cause both pressure- and time-dependent changes in two pathways that have been linked to diverse cellular physiologies. The protein kinase B, or Akt, signaling pathway is widely expressed in mammals and is constitutively expressed in low levels in the adult brain. In response to cellular stress or injury, Akt is activated through phosphorylation in both vascular and neuronal cells and acts as a key regulator in a host of signaling pathways responsible for a wide range of cellular physiology. 70,71 It is known to play an important role in diabetes and cancers and has also been implicated in several diseases of the CNS, a fact that has led to a great deal of interest in this pathway as a therapeutic. 70 –72 Underwater blast caused an early and robust increase in phosphorylated Akt by 1 day at all pressures, that decreased by 3 days, and then returned to baseline by 14 days. Activation of Akt is known to be neuroprotective, 70,73 –76 and the absence of cell death even at very high pressures may be a result of this transient activation. Additional studies in these laboratories have also shown a robust activation of brain Akt in animals exposed in head-only fashion to primary blast (unpublished observations).
Underwater blast also caused a pressure-dependent decline in VEGF levels that were slightly delayed and maximal by 7 days. Expression of this protein occurs not only in the endothelium, but also in neurons, astroglia, and microglia. 77 Reduced levels of VEGF have been implicated in a number of neurodegenerative diseases and in TBI, whereas activation of the VEGF pathway by multiple growth factors is known to be neuroprotective through neurogenic effects and the inhibition of apoptotic cell death. 78 –80 This latter action is mediated, in part, through activation of the Akt signaling pathway. 77,81,82 The opposing actions of blast on Akt and VEGF suggest that blast has separate and independent effects on the two pathways, with early activation of Akt, and a slightly delayed inhibitory effect on VEGF. This delayed VEGF inhibition may be responsible for the rapid decline in Akt levels observed after 1 day. Further work is required to resolve these apparently contradictory findings.
The work presented in this report is part of a comprehensive research effort to understand the effects of primary blast on the biology of the brain. The experimental parameters were intentionally simplified so as to answer the most basic question of whether single-pulse primary blast-induced stress waves cause changes in brain tissue without whole-body systemic involvement, the complicating boundary effects of the skull, or exaggerated global acceleration artifacts. The finding that primary blast impacts at least two cellular pathways known to have multiple roles in cell physiology, TBI, and neurodegenerative diseases has important mechanical and biological mechanistic implications for the study of primary bTBI and also suggests that underwater blast may effect changes in the brains of submerged individuals. Work is currently being carried out that is imposing added complexity and more-realistic shock exposure parameters to this brain cell aggregate model system, so as to further define the factors that determine the outcome of primary blast on the brain.
Footnotes
Acknowledgment
The authors are grateful to Mr. D. Ritzel (Dyn-Fx Consulting) for helpful discussions on several aspects of these studies.
Author Disclosure Statement
No competing financial interests exist.