
Abstract
In the military environment, injured soldiers undergoing medical evacuation via helicopter or mine-resistant ambush-protected vehicle (MRAP) are subjected to vibration and shock inherent to the transport vehicle. We conducted the present study to assess the consequences of such vibration on the acutely injured spinal cord. We used a porcine model of spinal cord injury (SCI). After a T10 contusion-compression injury, animals were subjected to 1) no vibration (n = 7–8), 2) whole body vibration at frequencies and amplitudes simulating helicopter transport (n = 8), or 3) whole body vibration simulating ground transportation in an MRAP ambulance (n = 7). Hindlimb locomotor function (using Porcine Thoracic Injury Behavior Scale [PTIBS]), Eriochrome Cyanine histochemistry and biochemical analysis of inflammatory and neural damage markers were analyzed. Cerebrospinal fluid (CSF) expression levels for monocyte chemoattractant protein-1 (MCP-1), interleukin (IL)-6, IL-8, and glial fibrillary acidic protein (GFAP) were similar between the helicopter or MRAP group and the unvibrated controls. Spared white/gray matter tended to be lower in the MRAP-vibrated animals than in the unvibrated controls, especially rostral to the epicenter. However, spared white/gray matter in the helicopter-vibrated group appeared normal. Although there was a relationship between the extent of sparing and the extent of locomotor recovery, no significant differences were found in PTIBS scores between the groups. In summary, exposures to vibration in the context of ground (MRAP) or aeromedical (helicopter) transportation did not significantly impair functional outcome in our large animal model of SCI. However, MRAP vibration was associated with increased tissue damage around the injury site, warranting caution around exposure to vehicle vibration acutely after SCI.
Introduction
I
It has been reported that vibration induces a variety of effects at the tissue and cellular level. Vibration has been linked to structural damage in various internal organs and tissues. 3 –10 For example, whole body vertical sinusoidal vibration (100 Hz, 2 h/day for 10 days) resulted in damage to cellular organelles in the liver, including enlarged and elongated mitochondria with reduced cristae, enlarged cisternae in the sarcoplasmic reticulum, and large areas of intracellular glycogen. 9 A single 4 h session of local vibration at 30, 120, or 800 Hz in a rat tail model evoked endothelial and nerve cell disruption (including swollen and disrupted myelin, intraneural edema, and dilated arterioles within the ventral tail nerve) and elevated nitrotyrosine immunoreactivity indicative of free-radical damage. 11 –13 Several studies conducted in vitro indicated that vibratory loading induced a variety of abnormalities such as changes in gene expression, protein metabolism in intervertebral disc cells, and water content. 14 –19 Research has also shown that whole body vibration of rats (1–20 and 120 Hz) influences brain monoamines such as noradrenaline and 5-HT, well known neurotransmitters in the central nervous system (CNS). 20,21 Clearly there is compelling evidence from these in vitro and in vivo studies to suggest that vibration can induce a myriad of biological responses and pathological changes at the cellular and tissue level.
Typically, transportation-related vibration occurs within a range of frequencies between 0.5 and 80 Hz and causes cyclical motion of the body at its interface(s) with the vibrating structure. 22 The frequency of vibration will depend on many factors, including: the vehicle type (ground or airborne), patient suspension, support and immobilization systems, and the terrain or external forces encountered by the vehicle. Some investigators have shown that newborns are exposed to considerable mechanical vibration during ambulance transport, which might be a potential risk factor for cerebral bleeding among very-low-birth-weight infants. 23 –25 Grosek and coworkers 26 found an association between ground transportation and increased heart rate and peripheral blood leukocyte counts in neonates. Although the factors leading to these phenomena are complex, mechanical vibration during ambulance transport may conceivably play a role given its known effects on biologic tissues.
Recently, we investigated the effect of subjecting the traumatically injured spinal cord in a porcine model to resonance frequency vibration. 27 Surprisingly, this study revealed that exposure to resonance vibration after traumatic SCI did not result in significant behavioral or histological worsening of the injury, even though resonance frequency vibration induced a rather dramatic degree of motion of the spinal column (between 8 and 18 mm of vertical motion) that occurred five times per second. The experimental setup that we developed to evaluate resonance vibration allowed us to apply vibratory stimuli with controlled frequency, amplitude, and duration. In this study, we aimed to extend this line of investigation to the setting of helicopter and ground transport. Specifically, using our porcine model of SCI, we examined the effects of two different types of transportation-induced vibration signatures: 1) a highly impulsive vibration profile with frequencies and amplitudes that simulated those measured in a mine-resistant ambush-protected vehicle (MRAP) as transportation, and 2) a less amplitude-varying vibration signal with frequencies and amplitudes that simulated those measured in a military helicopter.
Methods
All animal procedures were performed in accordance with the guidelines of the Canadian Council for Animal Care and approved by the University of British Columbia Animal Care Committee and the United States Army Medical Research and Materiel Command (USAMRMC) Animal Care and Use Review Office (ACURO).
Animals and experimental design
Female miniature Yucatan pigs (Sinclair Bio-resources, Columbia, MO) weighing 20–30 kg were group housed at our large animal facility for 5 weeks before surgery. Experimental animals were subjected to: 1) a 3 h custom vibration profile after SCI with frequencies and amplitudes that simulated those measured in a military helicopter (n = 8, with the dominant frequency being 17 Hz); or 2) a 3.5 h custom vibration profile with frequencies and amplitudes that simulated those measured in an MRAP (n = 10, with the dominant frequency being 2 Hz). Vibration exposure signals were recorded by our collaborators at The United States Army Aeromedical Research Laboratory (USAARL) and processed to extract MRAP and helicopter vibration signatures, which were then applied to the animals. Two SCI control groups were included consisting of 7 and 8 animals, which received no vibration; therefore, each experimental group had an appropriate time-matched control. There were some additional differences in the methods between the groups, and those will be described in the appropriate sections subsequently. Figure 1A summarizes the methodology and cerebrospinal fluid (CSF) collection times across the experimental groups.

Schematic of timing of spinal cord injury (SCI), vibration duration, and cerebrospinal fluid (CSF) collection for the vibration and control animals.
Porcine model of traumatic SCI
Surgical procedures for SCI and postoperative care were performed as previously described. 27 Using anatomic landmarks, the T9, T10, and T11 pedicles were cannulated and instrumented with screws (Select™ Multi Axial Screw, Medtronic, Minneapolis, MN). After the T10 laminectomy, the weight-drop device was rigidly secured to the pedicle screws and positioned so that the impactor (mass: 50 g) would fall directly on the exposed dura and spinal cord at T10. The tip of the impactor (diameter: 9.53 mm) was instrumented with a load cell (LLB215, Futek Advanced Sensor Technology, Irvine, CA) to record the force at impact. For the MRAP portion of the study a displacement transducer was added to the weight-drop device so that we were able to measure displacement and velocity as well as force during the impact sequence. Displacement was determined from the position where the force sensor detects values greater than the background (“noise”) levels to the point where maximum force was encountered. Immediately following the contusion injury (drop height: 20 cm), compression was applied by placing a 100 g mass on top of the impactor for 5 min. Subsequently, the weight-drop apparatus was removed, and a curved plastic spacer (∼50 mm2) was inserted between the dura and the laminae to simulate ongoing extradural compression. After exposure to vibration had ceased (described in detail subsequently), the spacer was removed.
CSF collection
The technique of thoracic puncture for the collection of CSF is the same as that used in the study by Streijger and coworkers. 27 In the helicopter study, a CSF catheter was inserted into the intrathecal space so that the catheter tip would rest ∼2 cm caudal from the injury epicenter. For the MRAP study, we placed the catheters 8 cm caudal to the injury site for easier CSF collection over time. A total of 500 μl of CSF was collected 15 min before injury and right before the onset of the vibration procedure (Helicopter: 30 min post-injury; MRAP: 1 h post-injury). For both the helicopter and MRAP group, additional CSF samples were collected at 1, 2, 3, and 5.5 h after onset of the vibration protocol (Fig. 1A). For the MRAP portion of the study, two additional collection time points were added (12 and 24 h post-injury). These were added because the CSF catheter was positioned further away from the impact site (8 cm) compared with a catheter tip position in the helicopter study (2 cm), and we wanted to account for a potential ‘delay’ in monitoring changes in CSF concentration. Animals were euthanized at 12 weeks post-injury at which point the last CSF sample was collected. CSF samples were collected at the same time frame for the unvibrated control groups. Immediately after collection, samples were spun at 1000g for 10 min at room temperature. The supernatant was immediately frozen on dry ice and stored at −80°C until examined.
Specific methods for helicopter vibration (Experiment 1)
Animals in the helicopter group (n = 8) were vibrated in the operating room, as the test equipment was portable. Each pig was placed in a prone position on a long spine board (50-0013, NSN# 6530-01-490-2487, North American Rescue LLC, Greer, SC). Three triaxial accelerometers (CXL04GP3, Crossbow Inc., San Jose, CA) were mounted to the T9, T10, and T11 pedicle screws. Two straps (50-0027, North American Rescue LLC, Greer, SC) were positioned over the shoulder and hip area of the animal to secure the animal and spine board to a standard military litter (100047, Arizona Industries for the Blind, Phoenix, AZ). Initial tension in the seatbelts was maintained across animals. Vibration was controlled by an electromechanical shaker (LDS V455, Brüel & Kjær, Nærum, Denmark). Briefly, two rectangular frames made from steel Unistrut were mounted on top of each other. The lower frame supported the weight of the upper frame, pig, spine board, and litter. The upper frame was attached to the shaker through a threaded rod and rested on four springs located at each corner of the lower frame. The litter was attached to the upper frame. One single axis accelerometer (786A Wilcoxon, Meggitt Sensing Systems, Germantown, MD) was attached to the upper frame as feedback to the shaker. Four accelerometers (786A Wilcoxon, Meggitt Sensing Systems, Germantown, MD) were attached to the upper frame to verify that the direction of the vibration was mostly vertical. One accelerometer (786A Wilcoxon, Meggitt Sensing Systems, Germantown, MD) was attached to the bottom of the spine board to assess the vibration that was delivered to the litter. A custom vibration profile based on helicopter field measurements was used to vibrate the animals for 1.5 or 3 h, beginning 30 min after SCI (Fig. 1B). The control group (n = 8) went through the exact same surgical and handling procedures, but was not subjected to any vibration.
Specific methods for MRAP vibration (Experiment 2)
Unlike the helicopter vibration signature, the MRAP vibration profile contained low dominant frequency signals around the 2 Hz range. Because the shaker used for the helicopter vibration experiment (LDS V455, Brüel & Kjær, Nærum, Denmark) did not have the capacity to generate vibration in this low frequency range with sufficient amplitude, we had to acquire a different shaker system for the MRAP experiment (V850-440-SPA32K, Brüel & Kjær, Nærum, Denmark). This shaker was considerably larger than the shaker used in the helicopter vibration experiment and required a distinct power supply, making it impossible to house it in our standard operating room within the veterinary facility. We therefore set up the shaker in a separate vibration testing room ∼150 m away from the operating room. The animals were transported slowly on a wheeled operating table to the vibration testing room after the contusion SCI and insertion of the epidural cord compression spacer within the standard operating room. Portable breathing and anesthesia equipment was transported with the animal. Once inside the testing room, the animal was wheeled into a BioBubble (bioBUBBLE, Fort Collins, CO) to provide increased sterility and lifted onto the litter. The litter was bolted to a solid magnesium plate which was bolted to the electrodynamic shaker. The animal was shifted on the spine board such that the T10 accelerometer was vertically aligned with the center of the shaker. Seatbelts positioned over the pig's shoulders and hips, in a manner consistent with Experiment 1, secured the pig and spine board to the litter. A compression disc load cell (LAU200-50 lb, Futek, Irvine, CA) was used to measure the force between the belt and posterior-most aspect of the pig at the shoulder and hip to maintain consistent belt tension across animals. The spine-mounted accelerometers were repositioned (leveled) with the animal on the shaker. Four accelerometers were secured to the magnesium plate in locations comparable with those on the upper frame for the helicopter setup. A single axis feedback accelerometer (Brüel & Kjær) attached to the magnesium plate was used to control the shaker through custom software (Shaker Controller, Brüel & Kjær). The system amplifier was turned on and the gain increased to 25%, which caused an abrupt vertical motion of the animal as a result of the magnesium plate moving to the “zero” position for the system. The accelerations caused by this abrupt motion were recorded. Similarly, prior to removing the pig from the shaker, and in accordance with the manufacturers' (Brüel & Kjær) recommendations, the gain and amplifier were turned off and the accelerations were recorded.
The entire procedure from SCI until the animals were prepared for vibration recording took 60–70 min. For each animal (n = 10), the vibration was applied at 75 min post-injury. The custom vibration profile, based on field data collected on an MRAP, was 68 sec and was looped for 3 h, ∼157 times (Fig. 1C). The looping algorithm resulted in an ∼8–10 sec pause between loops. The animals were vibrated for 3 h, although the actual test lasted for ∼3 h and 30 min, to account for the pauses. The profile was a scaled version of the acceleration captured by the sponsor during an MRAP field test. The original signal was amplitude scaled to 69% in Excel. Then, the scaled signal was further modified using custom software for the electrodynamic shaker (Waveform Editor, Brüel & Kjær); the signal was low pass filtered with a cutoff frequency of 75 Hz and high-pass filtered with a cutoff frequency of 0.5 Hz. The DC offset was also removed in the software. These adjustments were made to the signal so that the displacement (twice integrated acceleration profile) was within the ±2.54 cm travel limits of the shaker.
The control animals (n = 7) were transported to the BioBubble and lifted onto the litter. The system was turned on and the animals experienced the abrupt motion from the magnesium head plate moving to the zero position. After 1 min, the system was turned off and the animals were returned to the wheeled operating table and positioned in the BioBubble for 3 h. Finally, the animals were transported back to the operating room for closing the wound. The only difference between the vibration and control animals was the applied vibration.
Functional behavior analysis
The stage of recovery of hindlimb locomotor function was assigned using the 10 point classification score (Porcine Thoracic Injury Behavior Scale [PTIBS]) as described in detail previously. 27,28 This scale ranges from no active hindlimb movements, score 1, to normal ambulation, score 10. PTIBS scores of 1–3 are characterized by “hindlimb dragging,” scores of 4–6 reflect varying degrees of “stepping” ability, and scores of 7–10 reflect varying degrees of “walking” ability.
CSF biochemical analysis (with enzyme-linked immunosorbent assay [ELISA])
Interleukin (IL)-6 (Cat. No. P6000, R&D Systems, Inc., Minneapolis, MN), IL-8 (Cat. No. P8000, R&D Systems, Inc., Minneapolis, MN), monocyte chemoattractant protein-1 (MCP-1) (Cat. No. E101-800, Bethyl Laboratories Inc., Montgomery, TX) and glial fibrillary acidic protein (GFAP) (Cat. No. RD192072200R, BioVendor, Candler, NC) was performed with a quantitative and commercially available ELISA according to the manufacturer's instructions. All CSF samples were tested in duplicate.
Spinal cord immunohistochemistry
At the end of the experiment (12 weeks post-injury), animals were euthanized, and the spinal cord was harvested, post-fixed, and cryoprotected as described previously. 27 Subsequently, spinal cords were frozen on dry ice, cut into 1 cm segments centered on the injury site before being cut into 20 μm thick cross cryosections. Sections were serially mounted onto adjacent silane-coated SuperFrost-Plus slides (Fisher Scientific, Pittsburgh, PA) such that sections on the same slide were obtained from tissue 400 micrometer apart and stored at −80°C. For differentiating gray and white matter, Eriochrome Cyanine R (ECR) histochemistry was performed. EC-stained sections were examined, and pictures (5x objective) were taken of sections at 800 μm intervals throughout the lesion site (Zeiss AxioImager M2 microscope, Carl Zeiss Canada Ltd., Toronto, ON, Canada). Images were analyzed using Zen Imaging Software (Carl Zeiss Canada Ltd., Toronto, ON, Canada), by manually tracing the spinal cord perimeter and spared tissue for each image captured. The spared white matter was defined as the areas that were stained for ECR, whereas gray matter was considered spared when it was a stereotypic light gray color with a consistent neuropil texture containing neuronal and glial cell bodies. The percentages of white matter and gray matter were calculated by dividing the spared white or gray matter by the total area of the spinal cord on a given section.
Statistical analysis
Parameters collected were body weight; behavioral recovery (using PTIBS) over time after SCI; % gray and white matter sparing over distance relative to epicenter at 12 weeks post-injury; IL-6, IL-8, MCP-1, and GFAP expression levels over time after SCI; and injury parameters (force, displacement, velocity). For the histological data, data were binned into 0.8 mm categories before statistical data analysis. Missing values were imputed using grand means for the given variable. Next, the data were analyzed using repeated measures analysis of variance (ANOVA), controlling for baseline expression level (biochemistry data), body weight, and force. MRAP data were also controlled for displacement and velocity (in the helicopter data, there were no observations on displacement and velocity). The results of the univariate tests were reported where the overall multivariate analysis of variance (MANOVA) was significant. Gray/white tissue sparing and PTIBS correlations were evaluated using the Spearman's rank correlation test. All data are reported as mean ± SEM. Values of p ≤ 0.05 were considered significant.
Results
For all animals, the urinary catheter was removed 6–8 days after SCI surgery. In the MRAP study, of the 17 SCI operated animals, 1 animal was excluded because of a low impact force (1834 kdynes), which fell more then four standard deviations away from the mean. In addition, two MRAP vibrated SCI animals were euthanized respectively 12 h and 3 days postoperatively because of pulmonary complications. A total of 8 helicopter unvibrated SCI controls, 8 helicopter-vibrated SCI animals, 7 MRAP unvibrated SCI controls, and 7 MRAP-vibrated SCI animals were included in the analysis.
Injury parameters
All animals received a contusion injury by dropping a 50 g mass from a 20 cm height at the T10 level of the spinal cord. For the helicopter vibration portion of the study, the maximum impact force applied to the exposed spinal cord measured at the tip of the impactor was 3353 ± 115 kdynes and 2846 ± 269 kdynes for the unvibrated SCI control and helicopter-vibrated SCI groups, respectively (Table S1) (see online supplementary material at
For the MRAP-vibrated SCI group and the corresponding unvibrated SCI controls, displacement was also measured (Table S2), defined as the distance the impactor tip travels from initial contact with the exposed spinal cord. A 20 cm weight drop resulted in 8.20 ± 0.11 mm and 8.32 ± 0.16 mm displacements for the unvibrated SCI control and MRAP-vibrated SCI groups, respectively. The mean impact velocity (displacement divided by time) was 1890 ± 22 and 1830 ± 21 mm/sec for the unvibrated SCI control and MRAP-vibrated SCI groups, respectively. There were no significant differences in displacement or impact velocity between the two groups.
Experiment 1: Helicopter vibration
Supplementary Figure 1 illustrates the effect of helicopter-simulated vibration on IL-6, IL-8, MCP-1, and GFAP levels in the CSF of SCI pigs (see online supplementary material at
To investigate the effect of helicopter vibration on hindlimb locomotor recovery, motor function was assessed weekly using the PTIBS: a test for recovery of locomotor function after SCI in pigs. 28 Figure 2A shows average PTIBS scores over a 12 week period for unvibrated SCI-controls and helicopter-vibrated SCI animals. After injury, unvibrated SCI animals showed major neurological deficits, with the majority of the animals (seven out of eight) demonstrating varying degrees of weight-bearing hindlimb movements with a complete lack of stepping (PTIBS ≤4). Although there was no statistical difference in the PTIBS scores between the two groups (p = 0.478), it was notable that half of the helicopter-vibrated SCI animals (four out of eight) recovered to occasional weight supported stepping (PTIBS ≥5) during the first 10 weeks post-SCI, compared with only one out of eight animals in the unvibrated SCI group. It became apparent that after 10 weeks, certain animals showed inconsistent performance (week 11–12 post-SCI). For that reason, we also determined the maximum PTIBS (Max PTIBS) score by taking each animal's highest PTIBS score obtained during the 12 week period (Fig. 2B). The average maximal PTIBS score trended to being higher (p = 0.058) in the helicopter-vibrated SCI group compared with the unvibrated SCI controls, respectively 4.75 ± 0.37 and 3.75 ± 0.31.
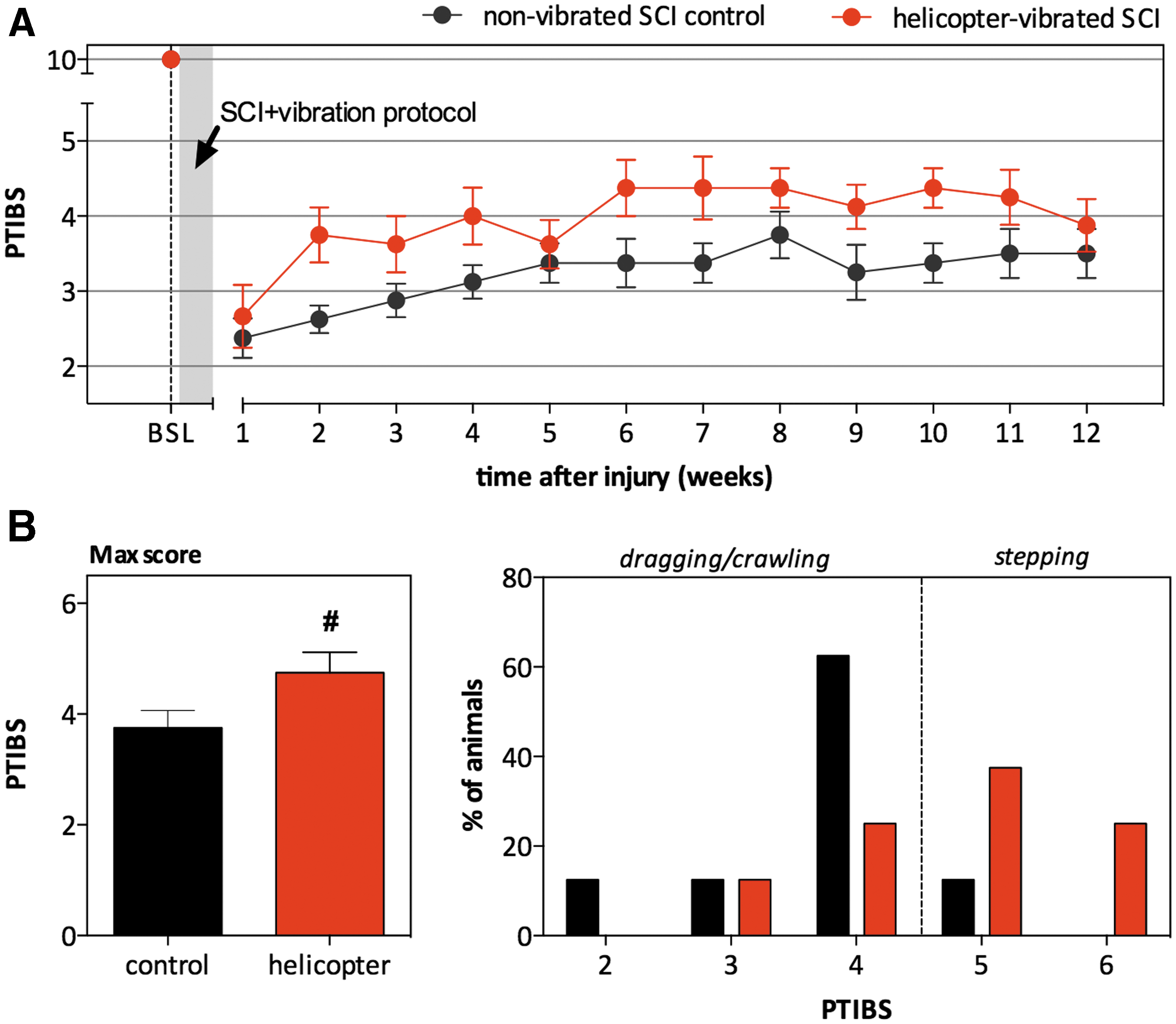
Effect of helicopter-simulated vibration on locomotor recovery (Porcine Thoracic Injury Behavior Scale [PTIBS] score) after spinal cord injury [SCI].
In order to quantify the extent of gray (Fig. 3A) and white matter damage (Fig. 3B) at the lesion epicenter as well as the rostrocaudal spread of the injury, analysis was performed on serial sections stained with Eriochrome Cyanine. The lesion epicenter was defined as the tissue section with the least percentage of tissue sparing. In the undamaged porcine spinal cord at the thoracic level, the gray matter represents ∼15% of the surface area of the entire spinal cord, whereas the white matter accounts for the remaining 85%. At 12 weeks following SCI, no preservation of gray and white matter was observed at the epicenter, whereas at 4 mm rostrocaudal distances from the epicenter, ∼10% of the gray and 60% of the white matter remained intact. No differences in spared gray or white matter were observed between the unvibrated controls and the helicopter-vibrated SCI animals. Lesion extension in both groups was ∼14.4 mm along the rostrocaudal axis of the spinal cord.

Effect of helicopter-simulated vibration on tissue sparing 12 weeks after spinal cord injury (SCI).
Experiment 2: MRAP vibration
Supplementary Figure 2 clearly shows a continual increase in IL-6, IL-8, MCP-1, and GFAP levels within both groups over time (see online supplementary material at
Similar to what was observed for the helicopter unvibrated SCI control group, the majority of the MRAP unvibrated control SCI animals demonstrated varying degrees of hindlimb movements with or without weight support and no stepping (PTIBS 2–4) (Fig. 4A). Whereas the MANOVA showed a trend for between-group differences for PTIBS scores (p = 0.065), with lower scores in the MRAP-vibrated SCI group, the univariate analysis showed no difference at any of the time points between the two groups. No significant differences were observed for the max PTIBS score between the helicopter-vibrated SCI group and unvibrated SCI controls (respectively 3.71 ± 0.27 and 3.57 ± 0.45 (Fig. 4B).
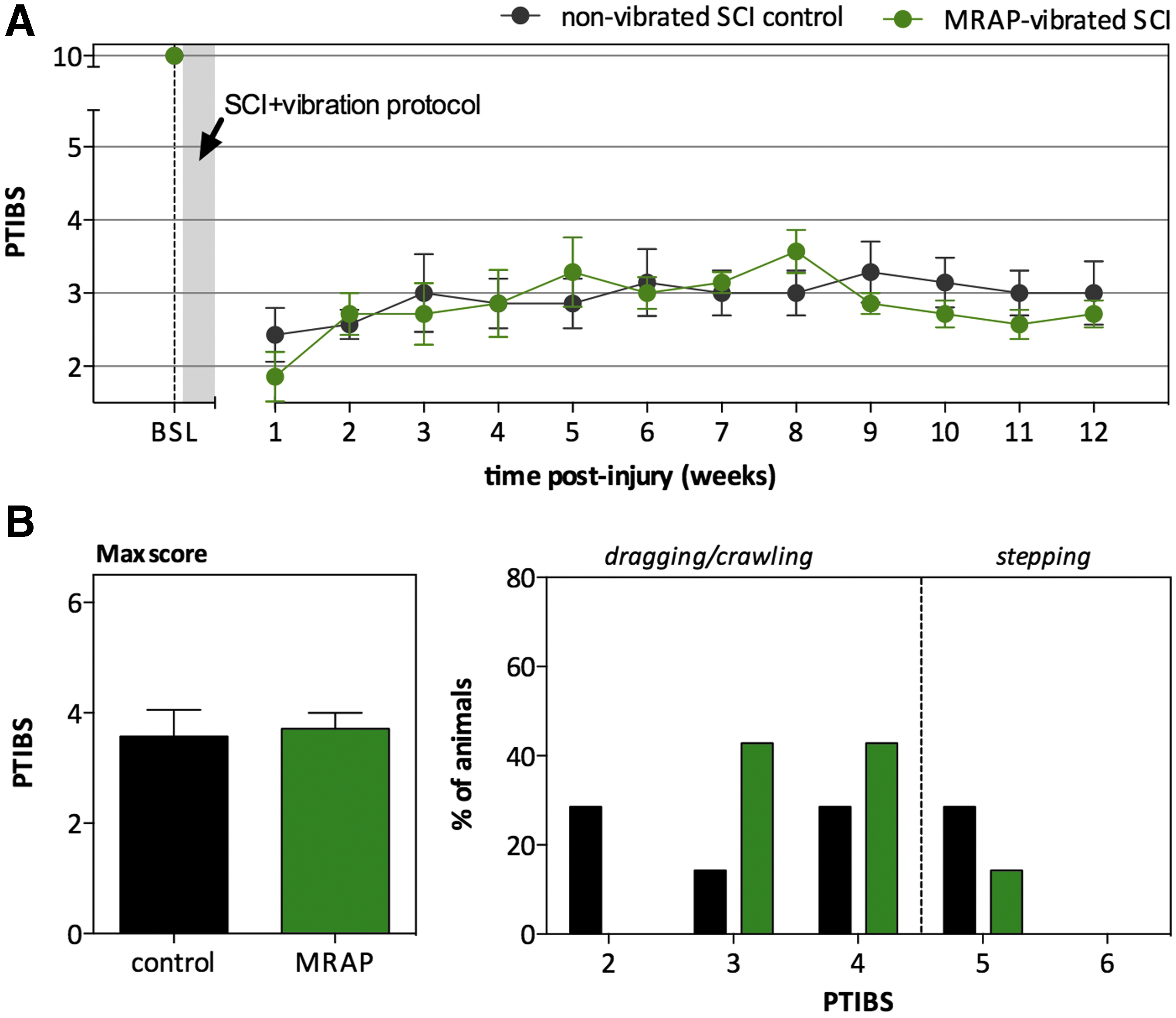
Effect of mine-resistant ambush-protected vehicle (MRAP)-simulated vibration on locomotor recovery (Porcine Thoracic Injury Behavior Scale [PTIBS] score) after spinal cord injury (SCI).
Whereas both groups demonstrated considerable spinal cord damage, white and gray matter loss appeared much greater in spinal cords of MRAP-vibrated SCI animals than in those of unvibrated SCI controls (Fig. 5). Although MANOVA failed to reveal overall group effects, the univariate ANOVA revealed significantly decreased gray matter sparing from 5.6 mm up to 8.0 mm rostral to the epicenter in the MRAP-vibrated SCI group (p = 0.010) (Fig. 5A). Notably, MRAP vibration caused a complete loss of gray matter up to 12 mm along the length of the spinal cord, which is almost twofold the extent observed in the unvibrated SCI controls (∼ 6.2 mm). Whereas there was also an apparent trend toward reduced white matter sparing following MRAP vibration, particularly rostral to the epicenter (4.8 mm as far as 9.6 mm), neither MANOVA nor univariate ANOVA revealed a statistical difference (Fig. 5B).

Effect of mine-resistant ambush-protected vehicle (MRAP)-simulated vibration on tissue sparing 12 weeks after spinal cord injury (SCI). Percentage spared gray and white matter to the whole spinal cord were measured using Eriochrome Cyanine stained sections.
Correlation between spared tissue and behavioral recovery
To evaluate a relationship with locomotor recovery as determined by PTIBS, cumulative gray or white matter sparing was used, obtained by summing the percentage of spared gray/white matter evaluated rostral and caudal to the epicenter (14.4 mm in each direction) as has been done previously (Fig. S3) (see online supplementary material at
At the individual animal level, the variability in cumulative spared white matter significantly correlated with binned PTIBS scores; however, cumulative spared gray matter did not. The analysis revealed a positive correlation, indicating that animals with lower values of spared white tissue tended to have lower PTIBS values. This correlation became stronger over time, with the highest correlation coefficient observed at the 9–12 week time period (r = 0.5).
Discussion
In this research project, we investigated the impact of MRAP and helicopter vibration profiles on the biochemical, histological, and behavioral outcomes after SCI in a porcine model. The present study revealed evidence of worsened tissue damage in the spinal cord of animals exposed to MRAP vibration acutely after traumatic SCI. Despite the worsened pathological damage, functional deficits and locomotor impairment were not significantly different from those seen in unvibrated SCI controls. When exposed to continuous helicopter vibration, half of the helicopter-vibrated SCI animals (four out of eight) recovered to occasional weight supported stepping during the first 10 weeks post-SCI, compared with only one out of eight animals in the unvibrated control group. However, when considering the group as a whole, there was no statistically significant difference. We additionally reported a positive correlation between white matter tissue sparing and functional improvement. However, the positive correlation was rather moderate, suggesting that tissue damage may only partially explain the variability in functional recovery. It would, therefore, be interesting to know whether a change in density of synaptic terminals in the lumbar spinal cord may better explain the inter-animal variability. Overall CSF expression levels for MCP-1, IL-6, IL-8, and GFAP were similar between the helicopter or MRAP group and the unvibrated SCI controls.
In a previous study using the same porcine model of SCI, we demonstrated that 3 h of whole body vibration at resonance frequency 5–7 Hz after SCI led to similar, (albeit not identical), improvements in locomotor recovery acutely after SCI as observed in the current helicopter study. 27 This was despite the fact that resonance frequency vibration resulted in a rather dramatic degree of motion of the spinal column (between 8 to 18 mm of vertical motion that occurred five times per second). 27 It has to be taken into consideration that the helicopter-simulated group displayed high variability in its performance, and that the positive effect on locomotor ability was only observed in four out of eight animals. It stands to reason that the relatively small sample size may have precluded us from obtaining significant p values (type II error). Nonetheless, it is also possible that the absence of a significant improvement on the PTIBS with helicopter-simulated vibration was partially the result of a “ceiling” effect in PTIBS scoring, with an improvement from a PTIBS score of 4 (hindlimb movements without stepping) to 5 (some degree of stepping ability) being too large of an incremental improvement to achieve. Notwithstanding, our data and the current literature 29 –36 do emphasize that certain combinations of frequency, intensity, and exposure times relevant to helicopter transport may not lead to exacerbation of the secondary damage after acute traumatic SCI.
There is compelling evidence from in vitro and in vivo studies to suggest that vibration can also induce a myriad of biological responses and pathological changes at the cellular and tissue level. 37 –43 However, many of these animal studies employed very intense vibration stimuli for prolonged periods of time, and whether such responses would occur in a medical transportation vehicular setting is unknown. In regard to the biological effects of high-frequency vibration and shock, little is understood about the underlying pathophysiologic mechanisms. In vitro and in vivo studies have shown that exposure to a wide range of vibration frequencies results in cellular effects such as decreased protein synthesis, aggrecan expression, water content, metalloproteinase expression, 15,16,19 disruption of microtubule organization, 44 endothelial and nerve cell disruption, and elevated nitrotyrosine immunoreactivity indicative of free-radical damage in rats. 11 –13 Further, acceleration-deceleration forces during aeromedical and maritime transport 45,46 have been associated with a potential for increases in CSF pressure, which if it persists, may lead to hypoxic injury. Extrapolated to our research study, it is unclear which of these mechanisms (if any) are applicable. Given that spinal immobilization is sought in the acute transport of SCI victims, we had intuitively expected that vibration with its resultant movement of the spine would have a gross mechanical impact of the injured spinal cord tissue. This was not so evident within the helicopter vibration group compared with the MRAP vibration animals. One intriguing question is whether vibration influences spinal cord pressure or perfusion. As we have experience with such measurements, we acknowledge that these would be highly technically challenging to measure in real time with intraparenchymal monitoring because of the motion artefact from the vibration.
Although the pig is more similar in size and anatomy to the human than the typical rodent model, we acknowledge that there are limitations to the use of this animal model in our investigations. It is recognized that human medical evacuation typically occurs with the patient in a supine position, and in our pig model it was technically impossible to perform the experiment with the pig in anything but the prone position. Changes in body position (lying on stomach versus back) may have a considerable impact on the magnitude of the transmission of vibrations, as soft tissue, fluids, joints, muscles, and bones are capable of absorbing and damping vibration before reaching the spinal cord. If the vibration amplitude is adequately damped down during the vibration wave propagation then only a small part of the vibration may actually reach the spinal cord. It was not possible to determine the differential effect of vibration on the animal in prone versus supine positioning in our experimental model. Additionally, although the pig spinal cord anatomy and size are much more similar to those of humans than to those of rodents, there are still obvious differences between the pig and the human that should be noted. For example, there are differences in size and structural anatomy (skin thickness, fat deposit, number of bones of the vertebral column, number of ribs) between humans and pigs, factors that might also influence the propagation of the vibration. Differences in vertebral body and canal size, subarachnoid space dimensions, spinal cord caliber, and gravitational load in a quadruped pig compared with a bipedal human should also be acknowledged.
As in any experimental study utilizing a rodent or large animal model of SCI, differences between the animal species and the human condition are important to consider when interpreting and extrapolating the data to the human condition. Nonetheless, the results of our study draw attention to the potential aggravation of secondary damage following traumatic SCI caused by excessive vibration arising from transportation vehicles. Further, in the present study, the MRAP acceleration amplitudes applied were merely 69% of full scale and, therefore, we do not know the effect of applying a full-scale MRAP vibration profile with larger displacements to SCI animals. It is also important to recognize that human SCIs are very heterogeneous and occur with many different mechanisms resulting in varying degrees of primary cord damage, spinal instability, and ongoing spinal compression. Our SCI model simulates an injury with sustained compression, but without significant spinal instability. Such instability would be associated with abnormal intervertebral motion at the injury site, and hence the effects of MRAP and helicopter-simulated vibration as observed in our study might be an underestimation of that what would occur clinically.
Individuals who sustain an acute SCI are typically immobilized on a spine board when transported to a medical facility.
2
This spine board (or “litter”) is in contact with the transport vehicle, either by being placed on the floor or by being secured to the vehicle frame. The approaches to isolate the litter from vibration or shock include introducing vibration isolation (i.e., rubber pads, shock absorbers, or other elastomeric materials) at the litter–vehicle interface or optimizing the vehicle suspension. It is also possible to introduce vibration isolation elements at the interface between the fabric bed of the litter and the side support structure of the litter. Another approach would be to fit dynamic vibration absorbers to the litter or to a separate support structure that could be designed to support the litter. Dynamic vibration absorbers incorporate an oscillating mass that vibrates in opposition to the vibration to be damped out and in that way “absorb” the unwanted vibration and lower the amplitude of the vibration. Vibration absorbers are tuned to a particular frequency; therefore, they are most effective against vibration at a single frequency. In this sense, vibration absorbers are a poor match to the variable frequency vibration and shock that occurs during medical evacuation in helicopters and MRAPs. However, active vibration isolators have been developed for various applications, and these devices can absorb vibration and various frequencies and, therefore, this technology also holds some potential to reduce the amount of vibration that individuals being evacuated after SCI are exposed to. Both vibration isolation and vibration absorber systems can be tuned to dampen or absorb particular frequencies and amplitudes of vibration by changing stiffness, damping, and mass properties, and, therefore, there are many approaches to controlling vibration in a medical evacuation context that have not yet been exhaustively evaluated to our knowledge. It is noteworthy that at conferences such as the annual Medical Health Systems Research Symposium (MHSRS) (
Conclusion
In summary, exposures to vibration in the context of ground (MRAP) or aeromedical (Helicopter) transportation did not compromise functional outcome. It is intriguing to note that the helicopter-simulated vibration conferred a modest positive effect on motor function in some animals, although this observation certainly does not substantiate it as a beneficial intervention. The helicopter field signal contained frequencies that we incorporated in a simplified sinusoid signal; however real-world vibrations are not sinusoidal. Part of the usefulness of sine testing is its simplicity, and we feel it is a good starting point into the study of vibration exposure acutely after SCI. Conversely, MRAP shock-type vibration was associated with increased grey tissue damage around the spinal cord injury site and an apparent trend for increased white matter damage. Therefore caution is warranted with regards to exposure to vehicle vibration and shock after acute SCI. These findings may stimulate further research into how such effects of vibration and shock may be mitigated in road vehicular transport, particularly in a military environment for soldiers injured in the field.
Footnotes
Acknowledgments
The authors thank Dr. Eric C. Sayre for his ongoing statistical support and the veterinary medical team and technicians at the University of British Columbia Centre for Comparative Medicine (Vancouver, BC, Canada) for their expertise with and dedication to the animals. Funding for this study was received from the United States Defense Medical Research & Development Program. Dr. Kwon holds the Canada Research Chair in SCI and the Dvorak Chair in Spinal Trauma from the VGH & UBC Hospital Foundation.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.