
Abstract
Acute subdural hematoma (ASDH) is associated with high morbidity and mortality. Whether the volume effect of the hematoma and increase of intracranial pressure (ICP) or the local effect of blood are responsible for this severe pathophysiology is unclear. Therefore, we compared subdural infusion of autologous blood and paraffin oil in a rat model of ASDH. In a histological study, we investigated the effects on acute ICP, cerebral perfusion pressure (CPP), cerebral blood flow (CBF), tissue oxygen changes, and brain damage at 2, 24, and 96 h post-infusion. Inflammatory reaction was analyzed by immuno-staining for microglia (ionized calcium binding adaptor molecule 1 [Iba1]) and activated astrocytes (glial fibrillary acidic protein [GFAP]). Besides acute ICP and CBF changes, we investigated the development of behavior (neuroscore and beamwalk test) for up to 4 days after injury in a behavioral study. Despite comparably increased ICP, there was a more pronounced lesion growth in the blood infusion group during the first 96 h. Further, there was an increased peri-lesional immunoreactive area of Iba1 and GFAP 96 h post-infusion, primarily in the blood infusion group, whereas hippocampal damage was comparable in both infusion groups. In the behavioral evaluation, paraffin-infused animals showed a better recovery, compared with the blood infusion group. In conclusion, comparable acute time-course of ICP, CPP, and CBF clearly indicates that the differences in lesion size, inflammatory reaction, and behavioral deficits after blood- and paraffin oil–induced ASDH are partially due to blood constituents. Therefore, current data suggest that subdural hematomas should be completely removed as quickly as possible; decompression alone may not be sufficient to prevent secondary brain damage.
Introduction
A
When compared with subdural hematoma, epidural hematoma appears to be a more benign lesion, with lower morbidity and mortality rates. In epidural hematoma, dura mater may serve as a barrier preventing direct contact with blood constituents. In a rat model, epidural hematoma did not result in significant biochemical changes. 5 Thus, a main factor for worse outcome in epidural hematoma is markedly increased ICP. 6,7
On the contrary, even small ASDH can cause massive brain swelling. Consequently, additional factors appear to influence lesion growth after ASDH. It therefore is suggested that harmful mediators diffuse from a hemorrhage into the parenchyma. 8 In order to test this hypothesis, we and others replaced the subdural blood volume by an inert mass, such as silicon gel (a mixture of silicone grease and oil), 9 –11 paraffin oil, 12,13 or a balloon. 14 Elimination of all blood-derived factors caused a much smaller lesion in all experiments, suggesting a strong involvement of blood-derived factors. However, possible candidates have not been investigated thoroughly in the model of acute subdural hemorrhage. A previous study found differences in glycolysis using silicone gel 9 in the peri-ischemic area, compared with blood infusion. Using paraffin oil as subdural mass, less hypoglycolysis was detected at the border of ischemia and no difference in hyperglycolysis was induced. 12 In both studies, the resulting brain lesion was significantly larger with subdural blood infusion, for which additional pathophysiological processes must be responsible. Placement of blood on the cortical surface seems not sufficient to induce massive brain damage, 15 whereas 3-h superfusion with a mixture of potassium and hemoglobin produces a large ischemic lesion, even in the absence of high ICP. 16
Comparison of effects induced by subdural blood or a substituting mass is difficult since ICP, a key factor for injury severity in humans, has not been well-controlled in animal studies. For instance, ICP was either not measured 9,11 or was significantly different between whole blood and silicone gel infusion. 10 In the latter study, no difference was found for local CBF and electroencephalography. Thus, the smaller lesion with silicone gel might be heavily influenced by the reduced ICP and not by blood constituents. In order to distinguish between mass effects and effects of blood components, ICP should be identical during the initial subdural infusion. Eliminating ICP as a confounding factor, developing differences in other parameters could then clearly be related to blood-derived factors. Therefore, we compared subdural infusion of autologous venous blood and paraffin oil on their effect on acute ICP, cerebral perfusion pressure (CPP), CBF, tissue oxygen changes, and development of behavioral, histological and inflammatory changes for up to 4 days after injury.
Methods
Animals
A total of 85 male Sprague-Dawley rats were used in all experiments (Charles-River, Germany). All experimental procedures were approved by the local ethic committee and were in accordance with the German guidelines for animal use and care.
Surgical preparation
Animals were anesthetized with chloral hydrate (36 mg/mL) using 1 mL/100 g body weight as an initial dose (intraperitoneally) and 1 mL/h to maintain anesthesia via intraperitoneal catheter. Each animal was injected with 1 mg atropine. Body temperature was maintained constant at 37°C with a feedback-controlled homeothermic blanket (Harvard, South Natick, MA). The rats were intubated and mechanically ventilated. For arterial blood pressure monitoring and blood gas analysis, a polyethylene tube was inserted into the tail artery. The volume needed for each blood gas analysis was 210 μL (ABL615/EML105; Radiometer, Copenhagen, DK). A jugular vein was cannulated to obtain blood for subdural infusion. Rats were fixed in a stereotaxic frame. After craniotomy 2 mm right of and 2 mm posterior to the bregma, a 23-gauge L-shaped blunted needle (Sterican, B. Braun, Melsungen, Germany) was inserted under the dura mater, and the craniotomy was sealed again with tissue glue (Histoacryl, B. Braun). 2,3,12,17
Anterior to the infusion needle and bregma suture, a skull area of 2 × 2 mm was thinned out. A Laser-Doppler probe (Vasamedics, St. Paul, MN) was placed over this window to measure CBF. A contralateral second burr hole was made for ICP measurement (Neurovent-P 3F, Raumedic, Münchberg). In a subset of animals (all from 2 h-survival group, see below) a burr hole was drilled posterior to the craniotomy for subdural hematoma infusion to insert a brain tissue oxygen (PtiO2) oxygen probe (Licox Medical Systems GmbH, Kiel, Deutschland). After an equilibration period with stable physiological values, a 15-min baseline period was employed. Next, 300 μL of autologous venous blood were infused at a rate of 50 μL/min subdurally. 18 Animals were monitored for at least 40 min from that time onward.
Experimental series and groups
In combination with ICP, CPP, CBF, and PtiO2 monitoring, we sought to investigate the temporal profile of histological, immunohistochemical, and behavioral changes after subdural blood or paraffin oil infusion. Thus, animals were randomly assigned to the following experimental series and groups: 1. Histological series (n = 57) a) acute ICP, CPP, and CBF changes and histological/immunohistochemical analysis after 24 h and 96 h survival (n = 42) b) acute PtiO2 changes and histological/immunohistochemical analysis after 2 h survival (n = 15) 2. Behavioral series (n = 28) c) acute ICP, CPP, and CBF changes and behavioral testing for 96 h.
Evaluation of behavioral and neurological deficits
Testing was performed in a quiet room in dim light. Before operation, all animals were trained daily to walk along a beam (diameter 1.8 cm, length 1 m, 2.5 cm between bars, 50 cm above ground) while their missteps were counted and the time it took to reach a black box was measured. 19 Sensory and motor integrity were tested with a neuroscore by evaluating motor activity, orientation, and reaction to tactile, visual, and auditory stimuli. The points obtained in the individual tests were then added together to yield an overall score ranging from 0 points (no deficit) to 100 points (most severe deficit). These tests were performed on Days -1 (baseline), 1, 2, 3, and 4 post-injury. 2,19,20
Neurohistological evaluation
Animals of the histological series were transcardially perfused with 4% buffered paraformaldehyde (pH 7.4). Their brains were carefully removed and post-fixed for 24 h before embedding in paraffin. Coronal sections (3 μm) spaced 250 μm apart were made through the paraffin-embedded brains and stained with hematoxylin and eosin to delineate the injury. The damaged area on each section was photographed with a CCD camera (SSC-C370P; Sony) connected to a light microscope (Zeiss; Oberkochen, Germany). The areas of ischemic brain damage underlying the subdural hematoma and immuno-reactive areas were surveyed with image analyzing software (Optimas 6.51; VSG, UK). 17
Immunohistochemistry
Sections were immunohistologically stained for glial fibrillary acidic protein (GFAP; astrocytes) and for ionized calcium binding adapter molecule 1 (Iba1; microglia).
Sections were incubated with 1% Triton X-100, treated with 10% hydrogen peroxide and blocked with non-immune horse serum. They were than either incubated with rabbit polyclonal antibody to GFAP (astrocytes; 1:500; Dako, Carpinteria, CA) or Iba1 (microglia; 1:1000; Wako Chemicals, Richmond, VA) overnight at room temperature. Sections were rinsed and incubated with biotinylated goat anti-rabbit secondary antibody for 30 min and rinsed in phosphate-buffered saline. The reaction product was visualized with diaminobenzidine tetrahydrochloride (Vector Labs). Omission of the primary antibody was used as a negative control (Supplementary Fig. 1; see online supplementary material at
Histological study.

Representative histological sections.
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). Comparison of the different groups was performed with one-way analysis of variance with post hoc comparisons for individual differences (Student-Newman-Keuls test) or by analysis of variance for repeated measures with Bonferroni correction for multiple comparisons. Data that were not distributed normally were analyzed by analysis of variance on ranks followed by the Dunn post hoc test (Sigma- Plot 11.0; Systat, CA). Differences were considered statistically significant at p < 0.05.
Results
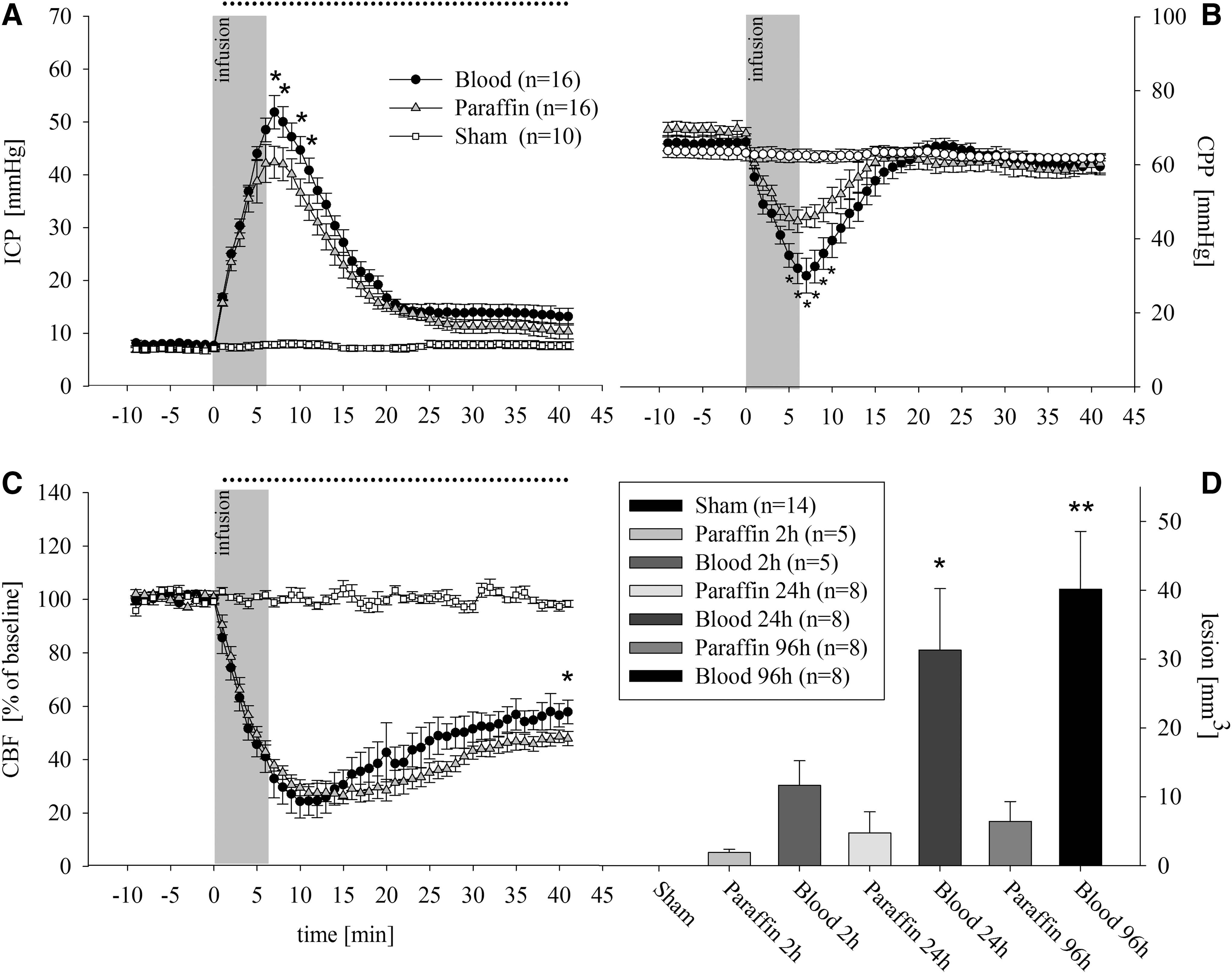
Acute effects of subdural blood and paraffin oil infusion on ICP, CPP, and CBF ICP reached its maximum values at the end of subdural infusion (Sham: 9 ± 2 mm Hg vs. Blood 52 ± 6 mm Hg vs. Paraffin 41 ± 6 mm Hg; p < 0.05 vs. Sham). Only in minutes 6 to 10 after subdural infusion the ICP was higher in the blood infusion group, compared with the paraffin oil infusion group. ICP remained elevated in both infusion groups until the end of measurement (Sham 9 ± 1 mm Hg vs. Blood 14 ± 1 mm Hg and Paraffin 11 ± 1 mm Hg; p < 0.05 vs. Sham; Fig. 1A). CPP had a reversed course. Starting at normal perfusion pressures CPP was critically low at the end of the blood injection and recovered (Fig. 1B). Meanwhile, CBF dropped in both infusion groups to 25% of baseline values and recovered incompletely (Sham 98 ± 3 LDU vs. Blood 54 ± 3 LDU and Paraffin 50 ± 3 LDU; p < 0.05 vs. Sham; Fig. 1C).
Acute effects of subdural blood and paraffin oil infusion on PtiO2
In a subgroup comprising 15 rats (n = 5/group), we additionally measured the effect of subdural infusion PtiO2. During baseline, there were normal PtiO2 values without differences between groups (Sham 30 ± 1 mm Hg vs. Blood 34 ± 4 mm Hg vs. Paraffin 27 ± 1 mm Hg; nonsignificant). At the end of the infusion, PtiO2 values dropped in infusion groups, compared with baseline values and Sham group (Sham 31 ± 1 vs. Blood 16 ± 3 mm Hg vs. Paraffin 16 ± 4 mm Hg; p < 0.05 vs. Sham). The values remained reduced in both groups during the whole measurement period, while the PtiO2 values of the Sham group remained constant.
Lesion growth and inflammatory reaction
After 2 h, the highest lesion volume was seen in the Blood group (Blood 11.6 ± 3.5 mm3 vs. Paraffin 1.9 ± 0.4 mm3; p < 0.05). This was more pronounced after 24 h (Blood 31.0 ± 9.1 mm3 vs. Paraffin 4.7 ± 3.7 mm3; p < 0.05). the most extreme differences with additional lesion growth were seen after 96 h (Blood 40.2 ± 8.4 mm3 vs. Paraffin 6.4 ± 2.8 mm3; p < 0.05; Fig. 1D and Fig. 2).
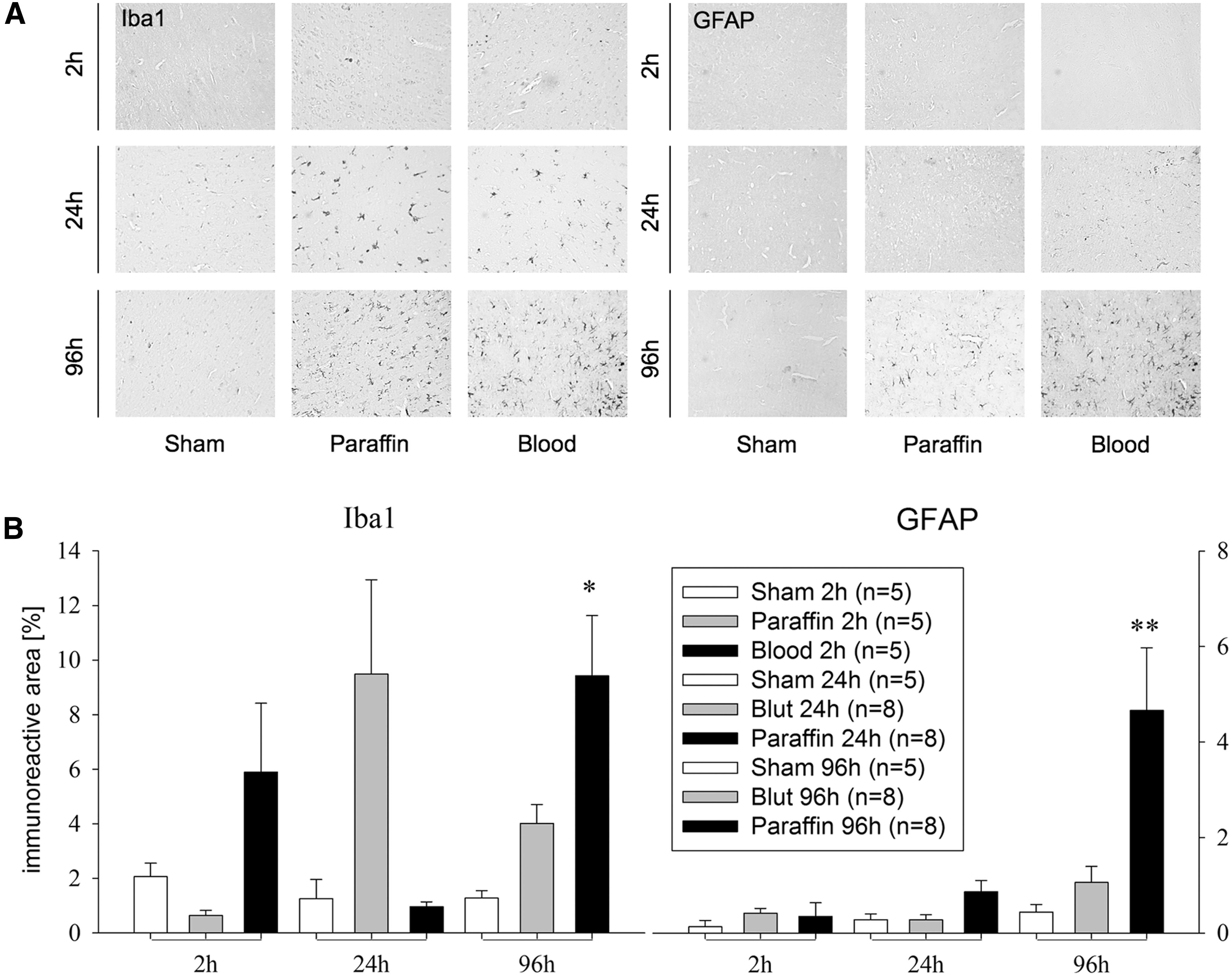
There was an increased Iba1- and GFAP- immunoreactive area on the ipsilateral hemispheres in the Blood group, compared with the Paraffin and Sham groups on Day 3, indicating microglial and astrocyte activation (Iba1: Blood 9.4 ± 2.2% vs. Paraffin 4 ± 0.7% vs. Sham 1.3 ± 0.3%; GFAP: Blood 4.7 ± 1.3% vs. Paraffin 1.1 ± 0.3% and Sham 0.4 ± 0.2%; p < 0.05; Fig. 3). No significant differences between groups were found on the contralateral hemisphere. Hippocampal damage was seen in both infusion groups. We found similarly reduced neuron counts in ipsilateral CA1 region 96 h after infusion (Sham 1688 ± 85 neurons/mm2 vs. Paraffin 1155 ± 147 neurons/mm2 and Blood 1055 ± 68 neurons/mm2; p < 0.05 vs. Sham).
Acute inflammatory reaction.
Effects of subdural blood and paraffin oil infusion on neurological and motor deficits
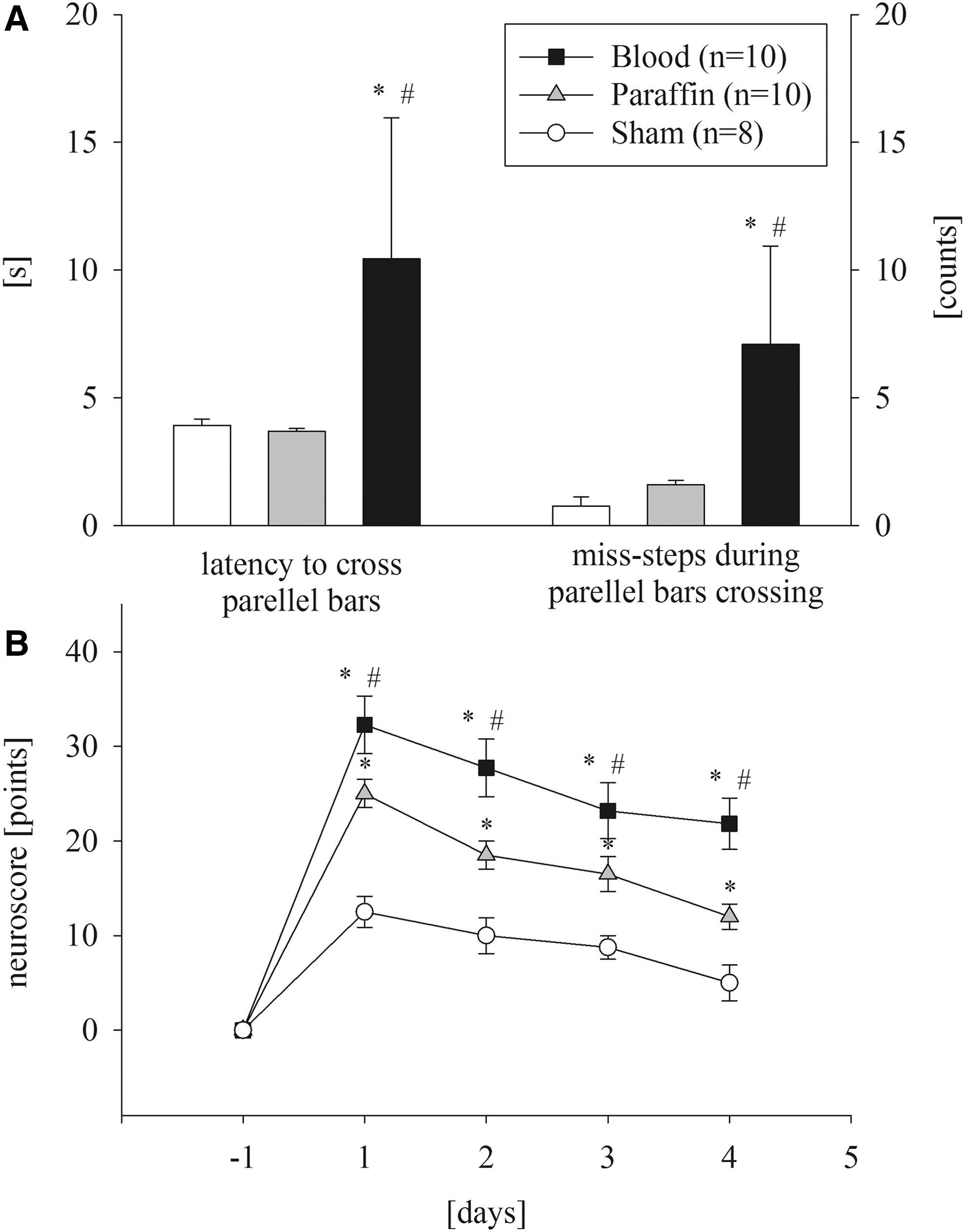
Acute changes in physiological parameters (mean arterial pressure, ICP, CBF) were similar to acute experimental series with histological/immunohistochemical endpoints (Fig. 1). After stable baseline conditions ICP reached its maximum values at the end of subdural infusion (Sham 6 ± 1 mm Hg; Blood 49 ± 3 mm Hg; Paraffin 47 ± 1 mm Hg; p < 0.05 vs. Sham). ICP remained elevated in both infusion groups until the end of measurement (Sham 6 ± 1 mm Hg vs. Blood 15 ± 1 mm Hg and Paraffin 15 ± 1 mm Hg; p < 0.001). In all infusion groups, the neurologic deficit, as expressed by the neuroscore, was increased as early as the first day after operation (Blood: 34 ± 11 points vs. Paraffin: 25 ± 5 points vs. Sham: 13 ± 5 points, p < 0.05 vs. Paraffin and Sham) but improved throughout the observation period, but with persistent differences between all groups (Fig. 4B). Rats receiving subdural blood infusion had the worst motor and neurological performance even at Day 4 post-injury. The beam-walk test showed similar results. After 3 days of training, rats crossed the bars in 3.8 ± 1 sec and with 1.6 ± 1 missteps on average. On Day 4 after ASDH, rats receiving subdural blood infusion performed worse than the two other groups (latency: Sham 3.9 ± 0.2 sec and Paraffin 3.7 ± 0.1 sec vs. Blood 10.4 ± 5.5 sec; missteps: Sham 0.8 ± 0.4 and Paraffin 1.6 ± 0.2 vs. Blood 7.1 ± 3.9; p < 0.05; Fig. 4A).
Behavioral study.
Discussion
In our study of ASDH, blood constituents seem to play a major role in secondary brain damage when compared with infusion of paraffin. Despite comparable ICP, CPP, and CBF, there was a time-dependent lesion growth and more severe neurological deficits in the blood infusion groups. ASDH is often associated with poor outcome. A crucial factor seems to be the early evacuation of blood in order to improve the prognosis of patients. 1 Besides ICP-related reduction of CPP and CBF, there are additional factors causing secondary damage. From clinical experience, the prognosis of patients with subdural hematomas is worse, compared with patients with epidural hematomas, ruling out ICP as sole injury mechanism. The different outcome might be explained by the prevention of harmful blood-derived factors to cross the separating dura mater freely. 6
Since ICP is only one damaging factor, other processes seem to influence the differences in lesion development. Data from an intracerebral hematoma model suggest that both tissue pressure and vasoactive substances are components influencing cerebral blood flow. 8 This study monitored ICP during the first minute of blood, silicon gel, and CSF infusion, but did not elaborate on additional factors. In our study, there were comparable ICP, CPP, and CBF courses in the Blood and Paraffin groups during the acute stage. The level of PtiO2 seems to be predominately determined by regional CBF. 22 Since there were no differences between groups in CBF in our study, we also could not detect PtiO2 changes. After ASDH in rats, decreased hippocampal blood flow and delayed damage are described. 23 Despite varying peri-lesional inflammation, we could not detect differences in hippocampal damage between infusion groups, suggesting that a decrease in CBF—and thus PtiO2—during the acute ICP peak led to hippocampal damage. At 2, 24, and 96 h, we found a progressive lesion growth and peri-lesional inflammation, which was considerably lower in the Paraffin groups.
The local effect of blood seems to be a major factor contributing to the peri-lesional damage. Among others, there seems to be microvascular disturbances after ASDH leading to excitotoxicity and edema formation. 24 A number of mechanisms appear to be involved in edema formation—for instance, red blood cell (RBC), lysed RBC, and hemoglobin-induced neuronal toxicity. Following intracerebral hemorrhage in rats, brain water content increased significantly after 1 day using lysed RBC and after 3 days using whole blood. 25 This and other studies suggest that a number of components of blood are capable of inducing brain injury and brain edema. 26 A cascade of events caused by RBC constituents increases blood–brain barrier (BBB) permeability. 12,27 To name just one, carbonic anhydrase-I (CA-1) derived from erythrocyte cytoplasm increases BBB permeability promoting vasogenic edema. 28 Subsequently, BBB breakdown leads to increased levels of intracerebral thrombin. 29 Thrombin itself is able to induce neuronal apoptosis in in vitro experiments 30 and to promote further delayed brain edema formation in an animal intracerebral hemorrhage (ICH) model. 31 Further, direct injection of thrombin activates the complement system and beyond that increases brain edema. 32
Independent of the effects of blood constituents on brain swelling and perfusion, direct contact of thrombin with brain cells is able to induce cell death. 33,34 This is in agreement with the fact that thrombin, produced by the coagulation cascade, is elevated in ischemic tissues and exerts cytotoxic effects in a dose-dependent manner. 35 A recent review describes the role of thrombin in brain injury. 36 Moreover, there are reports of thrombin-enhancing glutamate efflux from swelling astrocytes and subsequently inducing excitotoxicity. 37 Likewise, this might trigger spreading ischemia and play a role in lesion growth. 16 Thrombin increases neuronal excitability. 38 This pattern presumes that cortical spreading depolarization (CSD) occurs early after ASDH and therefore may contribute to the rapid lesion development. 39 This is supported by the fact that the subdural volume significantly reduces glucose metabolism early (90–120 min) after infusion in the ischemic areas, but was already more widespread with blood, compared with silicon gel and paraffin oil. 9,12 In addition, blood, but not silicon gel, was found to cause hippocampal hypermetabolism and led to secondary ischemic infarction due to a mismatch between glucose supply and demand. 9 In line with this, we found an early lesion growth only in the blood infusion group. This also was mirrored by the more severe neurological deficits in neuroscore and motor deficits in beam walk test.
Neuroinflammation is likewise a key factor causing additional cellular damage following brain injury. Activation of microglia was examined using immunostaining with Iba1 antibody and the formation of glial scars with GFAP antibody. By this time, few inflammatory factors have been suspected to cause microglial migration, but the mechanisms are not fully understood. 40 Microglia plays a crucial role in neuroinflammatory response and a major factor of pro-inflammatory molecule production (cytokines, chemokines, nitric oxide, growth factors). 40,41
For example, thrombin injection induces microglial activation, leading to significant motor deficits in a mouse model of ICH. This can be attenuated by use of thrombin inhibition or anti-inflammatory drugs. 42 As mentioned above, CA-1 from lysed erythrocytes induces edema and moreover leads to microglial activation and neuronal death. 43 Further, heme and its degradation products activate microglia and astrocytes, leading to leukocyte invasion and cytokine production potentiating injury. 44 On the other hand, hemoglobin degradation products such as heme and iron seem to be factors triggering oxidative brain injury, 45 but currently, the exact mechanisms remain unclear.
Finally, methemoglobin in the subarachnoid space induces microglial activation and tumor necrosis factor (TNF) α upregulation via Toll-like receptor 4, promoting widespread neuroinflammation. 46 Proportional to injury severity, astrocytes become activated, causing a scar with potentially beneficial and detrimental consequences. 47 Suppression of microglia activation seems to have consequences on behavioral deficits. In a hippocampal lesion model in mice, experimental microglia elimination improves functional recovery. 48 There seems to be a temporal split up of cytokine production and delayed microglial activation. In a rat model of ICH, there was an early interleukin (IL)-1ß, IL6, IL10 and TNF-α upregulation after 6 h and delayed microglial activation with a maximum from Day 3 to 7. 49 Congruently, other studies have found a peak of microglia activation after 72 h. 50,51 In our study, we found a peak of microglia activation 96 h post-injury, while there was increased lesion volume already after 24 h. There seems to be an early production of pro-inflammatory cytokines before microglial activation can be measured, which might lead to early injury. Modulation of pro-inflammatory factors utilizing immunotherapies may promote functional recovery. However, the key players, which according to our study may be a part of blood constituents, have yet to be identified after acute subdural hematoma.
Conclusion
Our data show that subdural blood itself aggravates lesion development, which is not due to increased ICP only. In this study with well-controlled physiological parameters, blood constituent–triggered pathomechanisms led to pronounced histological and functional damage despite comparable ICP, CPP, and CBF changes. With distinct peri-lesional inflammation, primarily in the blood infusion groups, and similar hippocampal damage in both infusion groups, our data suggest the local effect of blood as a major factor contributing to histological and functional damage. Therefore, current data suggests that subdural hematomas should be completely removed as quickly as possible. Further research is warranted to identify the blood constituents such as thrombin, CA-I, hemoglobin, or CSD, most responsible for secondary lesion growth after acute subdural hematoma.
Footnotes
Acknowledgments
The study was supported by Deutsche Forschungsgesellschaft (DFG) grant KE338/6-1. Data are part of doctoral thesis of CP and SE. We thank A. Ehlert, B. Koehler, and M. Malzahn for their excellent technical assistance.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.