
Abstract
Traumatic brain injury (TBI) is a leading cause of death and morbidity in industrialized countries with considerable associated health care costs. The cost and time associated with pre-clinical development of TBI therapeutics is lengthy and expensive with a poor track record of successful translation to the clinic. The zebrafish is an emerging model organism in research with unique technical and genomic strengths in the study of disease and development. Its high degree of genetic homology and cell signaling pathways relative to mammalian species and amenability to high and medium throughput assays has potential to accelerate the rate of therapeutic drug identification. Accordingly, we developed a novel closed-head model of TBI in adult zebrafish using a targeted, pulsed, high-intensity focused ultrasound (pHIFU) to induce mechanical injury of the brain. Western blot results indicated altered microtubule and neurofilament expression as well as increased expression of cleaved caspase-3 and beta APP (β-APP; p < 0.05). We used automated behavioral tracking software to evaluate locomotor deficits 24 and 48 h post-injury. Significant behavioral impairment included decreased swim distance and velocity (p < 0.05), as well as heightened anxiety and altered group social dynamics. Responses to injury were pHIFU dose-dependent and modifiable with MK-801, MDL-28170, or temperature modulation. Together, results indicate that the zebrafish exhibits responses to injury and intervention similar to mammalian TBI pathophysiology and suggest the potential for use to rapidly evaluate therapeutic compounds with high efficiency.
Introduction
I
Numerous animal models have been developed to understand the pathophysiology of TBI, and to aid in the identification of potential therapeutic strategies. These animal models have been used to elucidate cellular properties of trauma, as well as to investigate long-term consequences and neurological susceptibility to neurodegenerative disorders. 3 –9 The most commonly used model organisms include rats, mice, and swine with a variety of assays developed to quantify outcome. 5 However, the evaluation of therapeutic compounds using these models remains a laborious and time-consuming process. The lengthy pre-clinical development phase remains a significant hurdle in the timely identification of chemical entities or signaling pathways to treat TBI. The evaluation of a single pharmacological entity can take several years for comprehensive outcome analysis to be completed. In vitro models, although amenable to some degree of high throughput evaluation or therapeutics, are limited in the scope of anatomical and cellular complexities associated with whole animal preparations. Although in vitro models have been useful to parse out specific mechanisms of cellular injury after trauma and offer some degree of anatomical maintenance (e.g., organotypic cultures), 10 they do not fully encapsulate the cell-cell interactions that occur after TBI. In this regard, the zebrafish has potential as a bridging model organism with significant advantages to address the technical and time-dependent obstacles associated with pre-clinical drug validation.
Specifically, zebrafish are suited to the study of neurological diseases, boasting well-developed central nervous systems with brain structures analogous to mammalian anatomy. 11 –13 There are also significant advantages in live imaging and documentation of injury progression as transgenic lines allow for the development of transparent fish or fish with specific and readily identifiable fluorescent reporters with relevance to neurotrauma. 12,14,15 The adult zebrafish demonstrates homology in the expression and function of key receptors involved in synaptic neurotransmission, including 2-amino-3-(3-hydroxy-5-methyl-isoxazol-4-yl) propanoic acid (AMPA) and N-Methyl-D-aspartic acid (NMDA) receptors, 16,17 known to be important contributors to the excitotoxic pathways associated with TBI. Zebrafish also display remarkable similarity to mammals in sequence homology and susceptibility of cytoskeletal proteins such as the heavy neurofilament chain and alpha II-spectrin. 18 Further, there are numerous cognitive and behavioral outcome measures that can be rapidly quantified through automated image capture and analysis systems to correlate with molecular findings. 15 Thus, there is high relevance and potential for the use of zebrafish as a bridging model organism to increase the efficiency of translational studies of TBI.
Ultrasound is an energy-based modality with a number of diagnostic and therapeutic applications in medicine and biology. High-intensity focused ultrasound (HIFU) is a relatively new therapeutic ultrasound modality with a number of promising attributes in focalized, non- or minimally invasive treatment procedures. 19,20 HIFU beams can be generated extracorporeally by means of special ultrasound transducers and sent to, and focused at a target region inside the body. HIFU energy, depending on its exposure parameters, can induce thermal and/or mechanical bioeffects in tissue. Pulsed HIFU (pHIFU) is a special HIFU modality in which short pulses of HIFU energy are used to primarily induce mechanical effects in tissue such as stress forces and cavitation. 21
Currently, the only described models of brain injury in zebrafish involve penetrating injuries into brain tissues. 22,23 These models do not adequately recapitulate closed-head TBI. In this study we developed and characterized an adult zebrafish model of closed-head TBI. We further validated the responsiveness of the injury model to intervention using previously validated neuroprotective compounds as well as modulation of temperature to demonstrate the modifiable nature of the trauma.
Methods
Zebrafish care
All work was conducted in accordance with guidelines established by the Animal Care Committee at St. Michael's Hospital in accordance with the standards set by the Canadian Council on Animal Care.
Male and female wild-type adult zebrafish (6–12 months) of short-fin wild-type phenotype (AB, short fin) from heterogeneous stock were used in all experiments. Exclusion criteria included age greater than 1 year and any physical deformities, including crooked vertebrae associated with age or trauma. Fish were kept in tanks with pH 6.8–7.0 and maintained at 25°C with a light/dark cycle of 12/12 h. Normothermic conditions for AB zebrafish are considered to be between 25 and 28°C. 24
Ultrasound injury
A closed-head brain injury was induced using an image-guided 1-MHz pHIFU system (Fig. 1A.). The system was specifically developed for investigating effects of high-pressure focused ultrasound on soft tissue. The enclosure that houses the HIFU transducer and specimen holder is filled with room temperature water. The inner dimensions of this section are 23.5 cm in length and width with a depth of 19 cm. The HIFU transducer used in this study is a single-element spherical focused transducer with the aperture diameter of 125 mm and focal length of 100 mm. The axial focal length and the lateral focal width at the full width at half maximum (FWHM) are 8 mm and 2 mm, respectively. It employs an advanced 1–3 piezo-composite high-power ultrasound transducer technology (Imasonic SAS, Voray sur l'Ognon, France) toward an accurate delivery of ultrasound energy in tissue. The high-power wideband transducer technology used in this system enables generating either long (a few seconds) intensive bursts of ultrasound for thermal therapy or short (a few microseconds) pulses of high-pressure ultrasound for generating acoustic shockwave. The incorporation of a 7–9 MHz high-resolution ultrasound imaging phased array probe and its associated scanner (Sonix RP Scanner, BK Ultrasound, Richmond, BC, Canada) into the system gives it the required real-time confocal imaging capability that is critical for target tissue/organ localization and treatment monitoring. The distance from the base of the transducer housing to the specimen holder is approximately 12 cm. The distance from the tip of the ultrasound imager to the specimen holder is approximately 8 cm; however, the precise value is adjusted according to the ultrasound image targeting acquisition.

Ultrasound set-up. Targeted 1-MHz pulsed high-intensity focused ultrasound (pHIFU) system.
Highly focused and high-pressure ultrasonic shockwave beam and high-resolution imaging capability lead to a well-controlled and precise deposition of the acoustic energy in a millimeter-sized focal spot in the tissue target region, that is, the adult zebrafish brain. The acoustic focal pressure and dimensions of the focal zone of the pHIFU transducer used in this study were obtained using calibrated hydrophone measurements and computer numerical simulations from a previous study. 25 The pHIFU focal zone can be modeled as an ellipsoid of −6 dB axial and lateral dimensions of 7.5 and 1.2 mm, respectively (Fig. 1B). The alignment of the head and the HIFU transducer was performed using the integrated ultrasound imaging module of the system. Alignment of the system was set such that maximum acoustic pressure was achieved at the location of the zebrafish head (Fig. 1C). The focal zone of the pHIFU pulse encompasses a large portion of the adult zebrafish brain. The adult brain has approximate dimensions of length of 3 mm (measured from the olfactory lobe to the posterior end of the brainstem) and width of 2.5 mm measured from the tectum opticum and thickness of 2 mm. Focal peak positive acoustic pressure with the maximum output power setting of the system can reach to 20 MPa.
Fish were anesthetized using 100 ppm clove oil before immediately being placed into a custom holder that was then placed into the ultrasound apparatus. Fish were positioned on their left side within the holder and secured with surgical tape such that the head was positioned above a circular target window lined with a clear membrane to allow passage of ultrasound waves from below (Fig. 1A, inset). The holder was partially filled with water containing clove oil to maintain anesthesia. The ultrasound apparatus was filled with filtered room temperature water to allow propagation of the ultrasound waves from the transducer, up through the holder window, and terminating at the target zone (zebrafish head). The parameters of the pHIFU (i.e., amplitude and duration) were input to the arbitrary function generator. Once the fish and holder were positioned within the apparatus, the pHIFU pulse was delivered by pressing the activator switch on the function generator. Control fish were anesthetized and inserted into the set-up and allowed to recover in identical conditions with no pHIFU exposure (pHIFU system off).
Behavioral monitoring
Measures of recovery from pHIFU exposure were manually timed with a stopwatch and recorded by two observers with one blinded to the level of pHIFU exposure. Recovery times were recorded immediately following pHIFU exposure and return to recovery tanks, indicated as time 0 in Figure 2. Automated behavioral tracking tests were initiated at approximately 30 min following recovery of proper swim ability, but also were indicated as time 0 in Figures 3 and 4. The novel tank test (NTT) and shoaling behavioral tests were recorded using a digital video camera (Canon Vixia HF R500) and video files analyzed using EthovisionXT (Noldus, Netherlands). Injury levels used in behavioral studies were performed at 11 MPa and 50,000 cycles (50 msp HIFU duration), unless varying intensities were specified as in titration tests (1, 5, and 11 MPa).
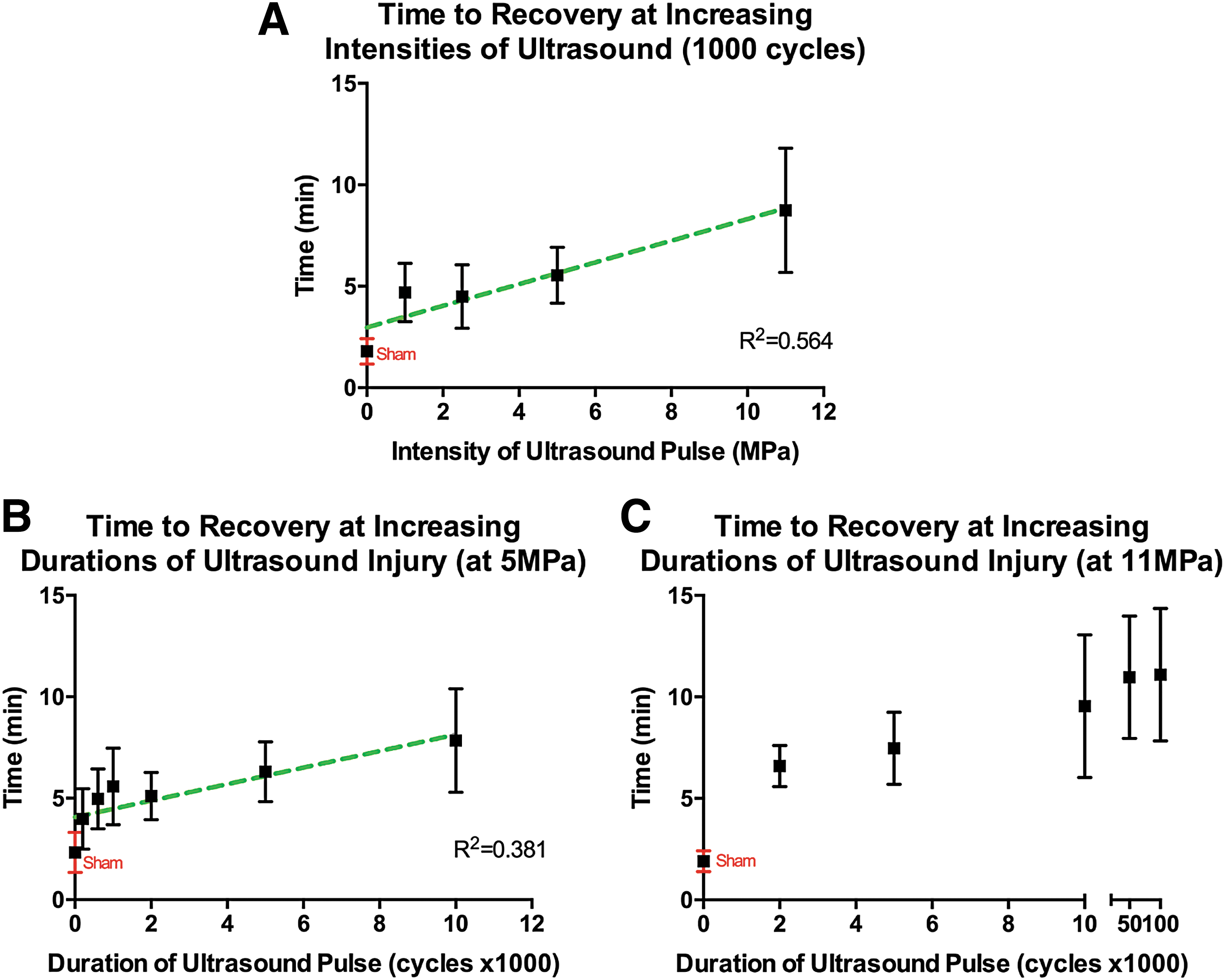
Recovery times. Data represent means ± SEM for n = 12 per time group.
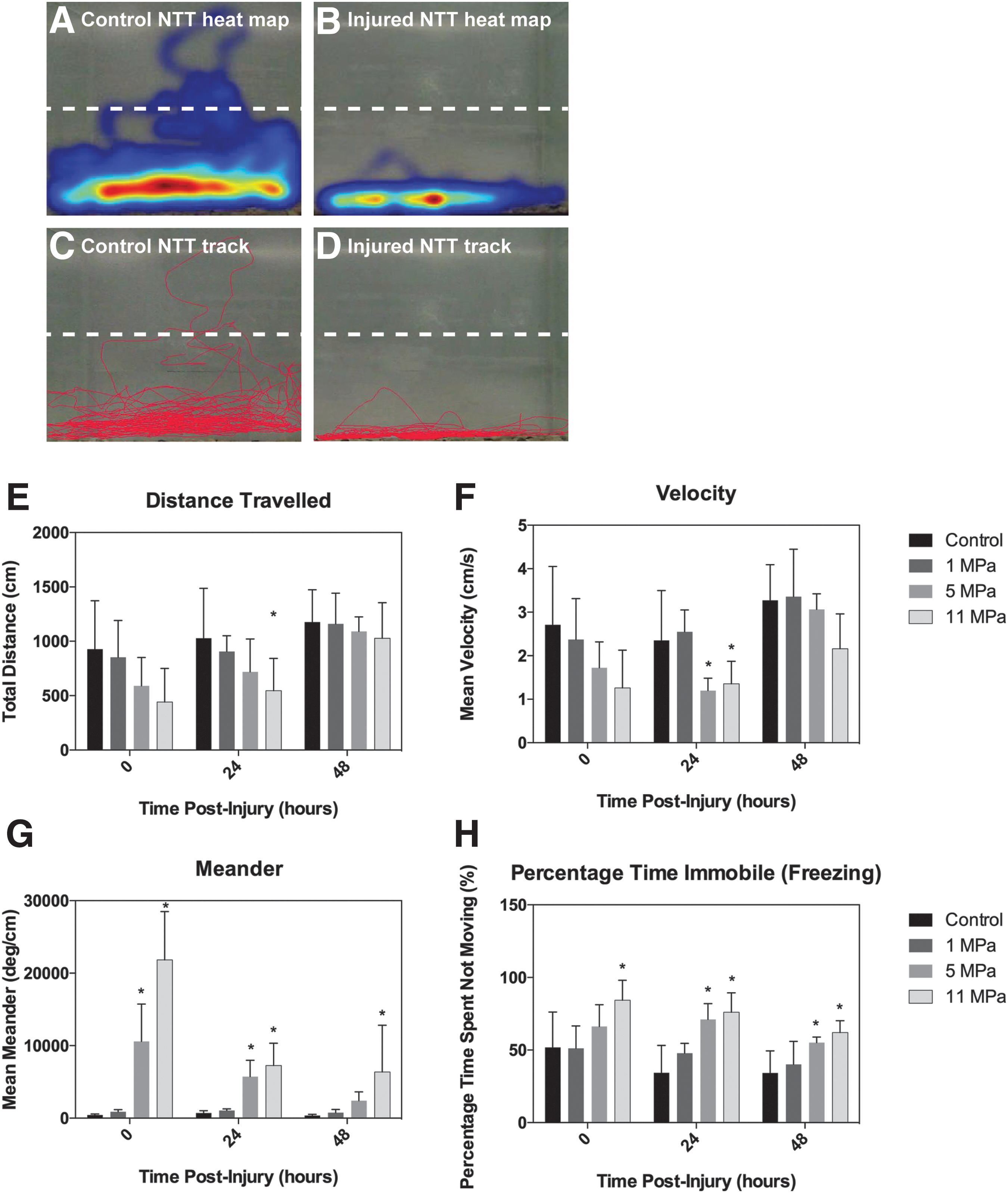
Novel tank test (NTT): locomotor activity.

Effects of pHIFU on anxiety and social behavior. Summary of behavioral data for n = 10. Data represent means ± SEM.
Novel tank test
The complex behavioral repertoire of zebrafish has been evaluated in tests such as the NTT. 26 –28 In general, this task evaluates locomotor and vertical exploratory activity in a novel environment and has become the standard for measuring stress and anxiety in zebrafish as well as exploration, 26,29 where individual fish are introduced into a new tank and recorded by side-view camera for 6 min. Zebrafish initially dive to the bottom and explore horizontally before gradually swimming to the upper levels of the tank. 30 Numerous studies have pharmacologically validated the NTT as an assessment able to evaluate behavioral changes in zebrafish, such as anxiogenic and anxiolytic manipulations. 31 –33
The NTT was performed as previously described 15,34 (Fig. 2) with n = 10 per treatment group. Briefly, a rectangular tank was made opaque on three sides (back and sides of tank) and filled with 25°C conditioned water maintained at pH 6.8–7.0. Tests were run on a stable surface with all environmental interferences kept to a minimum. For the testing procedure each fish was removed from its communal holding tank and released into the testing tank and individually recorded for 6 min indicated as 0, 24, and 48 h post-injury recovery intervals. The 0 h post-injury time-point refers to the starting point of automated behavioral data acquisition, which corresponds to an actual measurement time of 30 min post-injury following recovery from anesthesia and proper swim behaviur. The behavioral measures were captured at a video frame rate of 30 frames/sec. Control fish were included and analyzed with each testing group. The locomotor activity of the zebrafish was assessed by measuring the total distance traveled, average velocity (not including stationary time), total time immobile (freezing behavior), and meandering (absolute turn angle—representative of the variations in direction of the center point of the animal divided by the total distance traveled). Latency to the top zone was examined to evaluate vertical exploration activity. Increased latencies to explore the upper region have been shown to correlate with levels of anxiety. 35
Shoaling test
Shoaling is a typical, robust group-forming behavior and closer shoaling distances have been linked to increases in predatory stimuli and anxiety. 36,37 The shoaling test measures the average distance between fish in groups, a paradigm used extensively to study psychiatric disorders as well as anxiety. 27,31 At 0, 24, and 48 h post-injury, adult zebrafish from the same treatment group were placed in a rectangular test tank (10 gal) in groups of 5 for a total of n = 10 per treatment group. Behavior was recorded for 10 min and analyzed using EthovisionXT (Noldus). Automatic target tracking determines a numerical value for shoaling by calculating the mean distance between subjects, a shoaling-specific parameter.
Western blotting
Whole brains (n = 4 per time-point) were dissected from adult zebrafish anesthetized in 100 ppm clove oil. Post-injury time-points assessed were 6, 12, and 24 h post-injury. Entire brains were homogenized in lysis buffer containing protease inhibitors (50 mM Tris-HCl, 1% NP-40, 150 mM NaCl, 1 mM EDTA, 1 mM PMSF, 1 mM NaF, 1 lg/mL apro-tinin, 1 lg/mL leupeptin, and 1 lg/mL pepstatin). Protein quantification was determined by the modified Lowry method. 38 Samples were normalized for equal loading (40 lg/lane), and were electrophoresed on 7.5% or 12% SDS-PAGE gels, and transferred overnight to nitrocellulose membranes. Blocking of membranes was performed in 5% non-fat milk blocking solution for 1 h at room temperature. Immunoblots were probed for Neurofilament 160 (NF 160; Sigma-Aldrich, Oakville, Ontario), β-III tubulin (Sigma-Aldrich), the activated form of caspase-3 (Abcam, Toronto, Ontario), and beta APP (β-APP; Sigma-Aldrich).
These protein markers represent key pathways of secondary injury following TBI. NF 160, a cytoskeletal medium neurofilament and β-III tubulin, a microtubule protein expressed in neuronal cytoskeletons, are components of axonal structure and transport that are disrupted following injury. They accumulate and are found in axonal swellings and varicosities. 39,40 Caspase-3 is a calcium-activated cysteine protease that results in delayed, apoptotic cell death. 41 –43 Beta-APP immunoreactivity is used as a marker for axonal damage and impaired axonal transport post-TBI. 44 Primary incubations were performed overnight at 4°C at a dilution of 1:500 (Caspase-3), 1:3000 (β-III tubulin), 1:200 (βAPP), and 1:1000 (NF 160) in blocking solution. Secondary antibody incubation was performed for 1 h with horseradish peroxidase (HRP)-conjugated goat anti-rabbit or goat anti-mouse secondary antibodies at 1:3000 dilution. Rabbit polyclonal anti-GAPDH antibody (Sigma-Aldrich) was used as a loading control at 1:1000 dilution, and visualized with HRP-conjugated goat-anti-rabbit secondary antibodies at 1:3000 dilutions. Washes in three changes of TBST were performed between incubations. Bands were visualized using a chemiluminescence kit (Perkin Elmer, Waltham, MA) and images were acquired on a BioRadVersaDoc Imaging system (BioRad, Mississauga, Ontario; Fig. 5A). Densitometry analysis was performed with NIS-Elements BR Microscope Imaging Software (Nikon Instruments Inc., Melville, NY). Protein expression density values were normalized as changes relative to control levels of expression (Fig. 5 B–E). Western blot samples were run in triplicate to ensure consistency of results.
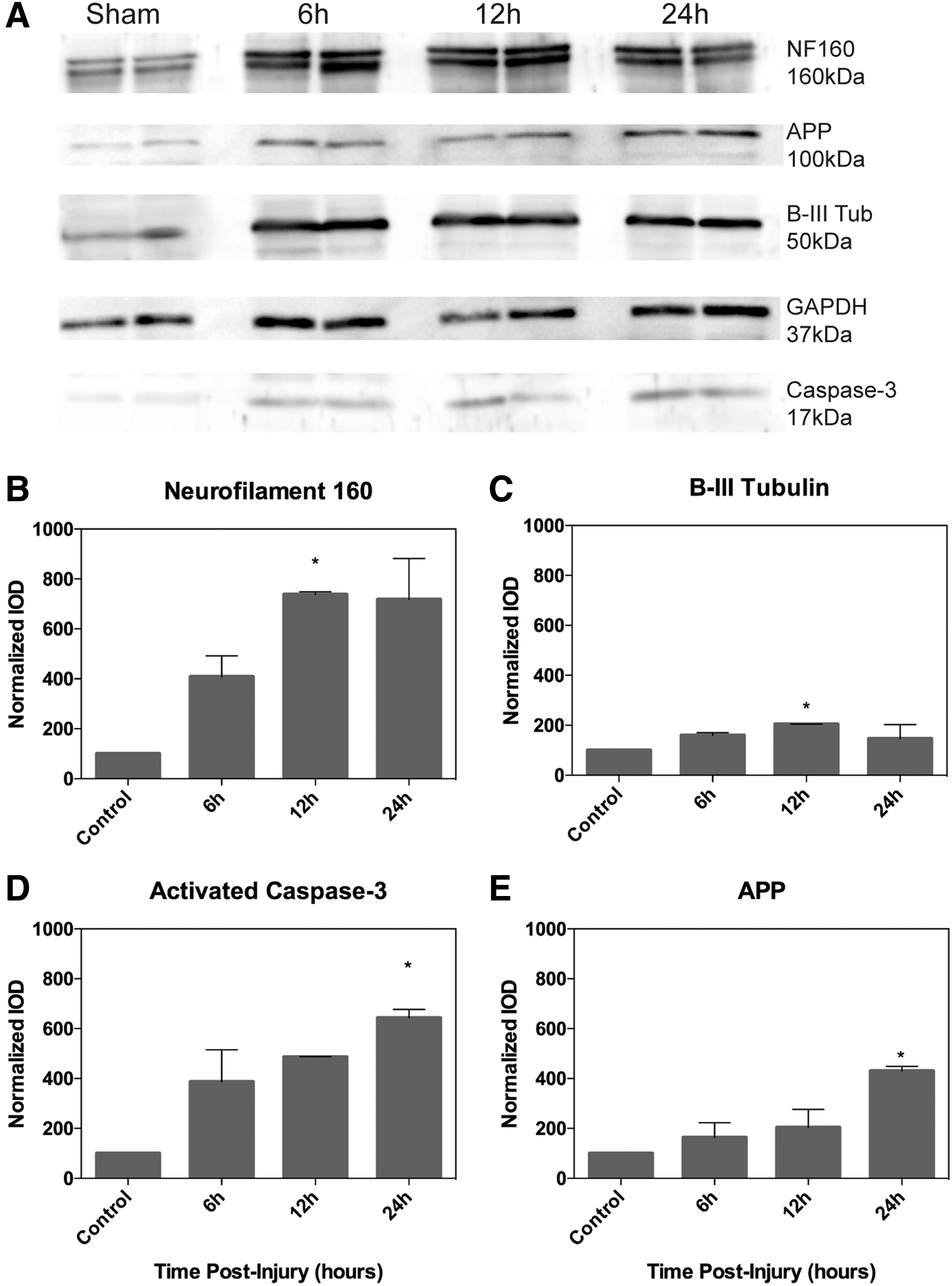
Protein analysis. Data represent means ± SEM for n = 4 per time-point.
Temperature intervention
Lowering core temperature by varying methods has been used clinically to treat TBI patients for over 100 years and many experimental studies have confirmed that moderate hypothermia confers protection against ischemic and non-ischemic brain hypoxia, TBI, and other anoxic injury. 45,46 We used decreased temperature to demonstrate the sensitivity of our model to a clinically relevant intervention. Temperature experiments were performed as described above (100 ppm clove oil anesthesia and pHIFU application) with recovery tanks set 3°C below normothermic conditions (22°C versus 25°C; n = 10 per treatment group). Post-injury survival was determined by monitoring fish in recovery tanks at 22, 25, and 28°C. Behavioral testing was performed 24 h post-injury comparing injured fish recovering at 25 and 22°C.
Drug injections
The NMDA glutamate receptor inhibitor MK-801 targets the upstream initiator of the excitoxicity cascade and has been previously demonstrated to be neuroprotective in models of TBI. 47 We applied MK-801 intervention as proof-of-concept of a model organism responsive to pharmacological interventions with relevance to mammalian TBI. Doses (2 μl and 4 μl) of 12.5 μM MK-801 (Sigma-Aldrich) were injected into adult zebrafish via retro-orbital (RO) injection 48 1 h prior to 11 MPa ultrasound insult (n = 6 per treatment group). RO injection has a lower mortality rate than traditional intra-cardiac or intra-peritoneal injections while reducing risk of injury to a major organ as well as delivering injected samples directly into the bloodstream of the adult fish. 48 Behavioral tracking was performed as described above. Injections of 4 μl saline were administered in sham fish. Injections of MK-801 (4 μl) in fish not exposed to pHIFU served as inhibitor controls.
Statistical analysis
Dose-stratified recovery times were analyzed using linear regression.Western blot densitometry analysis was performed using a one-way analysis of variance (ANOVA) and Tukey's post hoc test. Dose-stratified behavioral end-points were analyzed by two-way repeated measures ANOVA with Tukey's post hoc test for multiple comparisons. Survival curve comparison was performed using a log rank test. Hypothermia and inhibitor intervention behavioral end-points were analyzed by one-way ANOVA with Tukey's post hoc test for multiple comparisons. Data were expressed as means ± standard error (SEM) and the significance was set at p < 0.05.
Results
The effect of pHIFU injury on recovery time
Acute TBI was induced by applying high-intensity, targeted ultrasound to adult zebrafish at varying intensities and durations (n = 12 per treatment group). We observed 0% mortality in the first 48 h following 5 and 11 MPa injury at all pHIFU durations. Fish subjected to pHIFU injury had impaired acute locomotor activity characterized by prolonged recovery from anesthesia followed by obvious impaired swimming capabilities. No subjects were excluded in data analysis. Observed obvious locomotor impairments included abnormal swim patterns characterized by corkscrew swim paths and an inability to maintain a proper horizontal position in all subjects injured at 11 MPa. Recovery time post-pHIFU insult was measured from time of insult until fish were upright, horizontal, and swimming without obvious motor impairment. We first examined the effect of increasing intensity (amplitude) of ultrasound on recovery. Overall, there was a strong positive linear correlation between injury intensity and recovery. As intensity increased, recovery times of injured fish were also prolonged (Fig. 2A; R2 = 0.564). Next, intensity was kept constant as duration of ultrasound pulse was increased. At 5 MPa, recovery times increased as the pHIFU pulse duration increased to 50 msec corresponding to 50,000 cycles (Fig. 2B; R2 = 0.381). At 11 MPa, recovery times increased until reaching a plateau at 50,000 cycles (Fig. 2C).
The effect of pHIFU insult on behavior
Locomotion in the novel tank test (NTT)
EthovisionXT software was used to analyze the effects of ultrasound injury on locomotor and vertical exploration in the NTT (Figs. 3,4, n = 10 per treatment group). Analysis of total distance traveled following insult at 0, 24, and 48 h time-points revealed significant differences between treatment groups (p = 0.003) using a repeated measures two-way ANOVA (Fig. 3E). At 24 h post-insult the 11 MPa injured groups demonstrated significantly less distance traveled compared with control fish (*p = 0.046). Recovery of swim distance in the 11 MPa group was significantly improved by 48 h relative to the 24 h time-point (p = 0.030). Control as well as 1 and 5 MPa insults did not have significant differences in distance traveled over time. Mean calculated velocities were found to be significantly different between treatment groups (p = 0.0004; Fig. 3F). Post hoc analysis indicated both 5 and 11 MPa insults resulted in significantly decreased velocities at 24 h (both *p = 0.043) relative to controls. Injuries in the 1 MPa group were not significantly different from controls at 48 h. In the 5 MPa group there was a significant improvement in swim velocity from 24 to 48 h post-injury (p = 0.005). There were no significant differences at 48 h post-injury between controls and the three injury intensities.
Meander outcome measures (Fig. 3G) demonstrated significant differences over 48 h between treatment groups (p < 0.0001). Post hoc analysis indicated significant increases at the 0 h time-point after ultrasound insult (30 min post-injury and recovery) as well as at 24 and 48 h later as compared with controls. Specifically, at 0 and 24 h post-injury, both 5 and 11 MPa injury levels resulted in significant increases in meander (*p = 0.034 and *p = 0.012 at 5 MPa, and *p < 0.0001 and *p = 0.0001 at 11 MPa), whereas only the higher intensity of 11 MPa had a lasting impairment at 48 h (*p = 0.036). The 11 MPa injury level also significantly affected zebrafish freezing behavior, reported as time spent immobile during the test (Fig. 3H). The percentage of time spent immobile was significantly different between treatment groups (p = 0.035). In the 11 MPa injury group, freezing time was significantly higher than controls at all three time-points (0, 24, 48 h; *p = 0.035, *p = 0.043, and *p = 0.006, respectively). At 5 MPa zebrafish exhibited significant increases in freezing behavior at 24 and 48 h post-injury as compared with controls (*p = 0.043 and *p = 0.044, respectively).
Anxiety and vertical exploration (NTT) and social interaction (shoaling test)
Latency to the upper zone of the tank was significantly different between groups (Fig. 4A; p = 0.0005). Latency was significantly higher at 0 h after 11 MPa injury compared with controls (*p = 0.036). The mean time to vertical exploration was 179.50 ± 42.17 sec in controls and 42.20 ± 28.08 sec at 11 MPa. Both 5 MPa and 11 MPa injuries resulted in significant increases in latencies at 24 h (*p = 0.031 and p = 0.035, respectively) and 48 h post-insult (*p = 0.032 and *p = 0.042, respectively) compared with controls. Latencies in the 11 MPa injury group decreased significantly at 24 and 48 h as compared with the 0 h time-point (p = 0.044 and p = 0.048, respectively).
The effects of injury on shoaling behavior were evident immediately after injury in all insult groups (1, 5, 11 MPa; Fig. 4 B). Overall, treatment groups were significantly different (p < 0.0001). Post hoc analysis indicated decreases in shoaling distance compared with controls in all injury treatment groups at 0 h (*p = 0.0009, *p = 0.002, and *p = 0.002). Shoaling distances were still significantly lower than controls at 24 and 48 h in 5 and 11 MPa injury groups (*p < 0.0001).
The effect of pHIFU insult on protein expression
Protein expression changes were examined at three time-points after 11 MPa ultrasound injury (6, 12, and 24 h, n = 4 per time-point). NF 160, an abundantly expressed protein in myelinated axons, had a 7.4-fold increase in levels of expression 12 h post-injury relative to control fish (Fig. 5B; p = 0.029). The same time-point revealed significant (2-fold) increase in the expression of β-III tubulin, a microtubule protein (Fig. 5C; p = 0.047). Activated caspase-3, a downstream marker of activated apoptotic pathways, as well as β-APP, a marker of axonal injury, were not found to be significantly increased until 24 h post-injury (Fig. 5D,E; p = 0.039 and p = 0.042, respectively).
Hypothermia modifies survival and impaired behavior following injury
Post-injury survival was significantly affected with varying recovery temperatures (Fig. 6; p < 0.0001, n = 10 per treatment group). Recovery carried out in normothermic conditions (25°C) resulted in a median survival of 8 days, whereas injured subjects that recovered at 28°C died earlier (median survival of 5 days). When the temperature was decreased to 22°C however, all subjects survived. Control subjects maintained at the hyperthermic conditions (28°C) had less than 1% mortality.

Hypothermia: effect on survival following 11 MPa pHIFU. Data represent percent survival for n = 10 (log rank test for comparing survival curves). Survival proportions of adult fish injured at 11 MPa and recovering in decreasing temperatures indicate significantly less mortality at both 25 and 22°C compared with injured fish kept at 28°C (p < 0.0001). pHIFU, pulsed high-intensity focused ultrasound.
In the NTT behavioral test (n = 10 per treatment group), injured fish that recovered under hypothermic conditions had significantly increased distance and velocity measures at 24 h post-injury (Fig. 7A,B; p = 0.0499 and p = 0.0374, respectively). Both meander and percentage of time spent immobile were significantly decreased at 24 h in the hypothermic group compared with injured fish in normothermic conditions (Fig. 7C,D; p = 0.019 and p = 0.002, respectively). Latency to upper zone was also decreased in hypothermic groups relative to hyperthermic fish (Fig. 7E; p = 0.0438). Social interaction was also significantly altered at 22°C recovery as shown by significantly increased distances between subjects at 24 h (Fig. 7F; p < 0.0001).
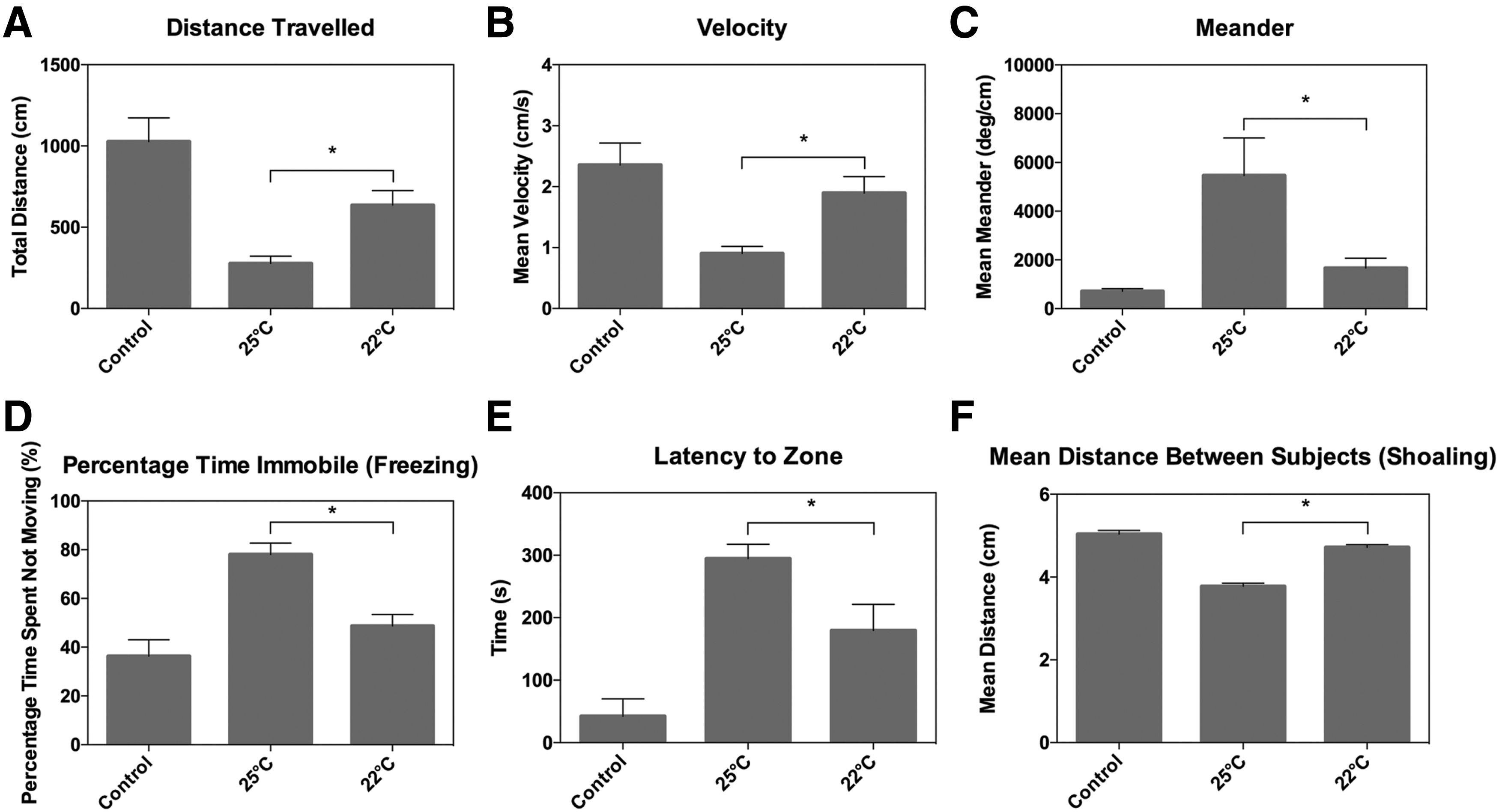
Hypothermia: NTT. Summary of behavioral data for n = 10. Data represent means ± SEM. Effects of decreased temperature on locomotor activity, vertical exploration, and social interaction 24 h post-injury.
MK-801 improves outcome following pHIFU injury
The NMDA receptor antagonist, MK-801, administered retro-orbitally 1 h pre-ultrasound insult resulted in decreased locomotor deficits and increased vertical exploration (Fig. 8, n = 6 per treatment group). Behavioral analysis showed improved outcome in all measured parameters at 4 μl of MK-801 at 24 h post-injury. Panels A–D of Figure 8 show significantly improved locomotor outcome (p = 0.003 distance traveled, p = 0.004 velocity, and p = 0.024 freezing). Statistically significant (p = 0.049) decreases in latencies to upper zone in subjects pre-treated with 4 μl MK-801 demonstrated increased vertical exploration (Fig. 8E). Finally, significant increases in mean distances between subjects in the shoaling test (Fig. 8F; p < 0.0001) were observed with the 4 μl injection volume of MK-801.
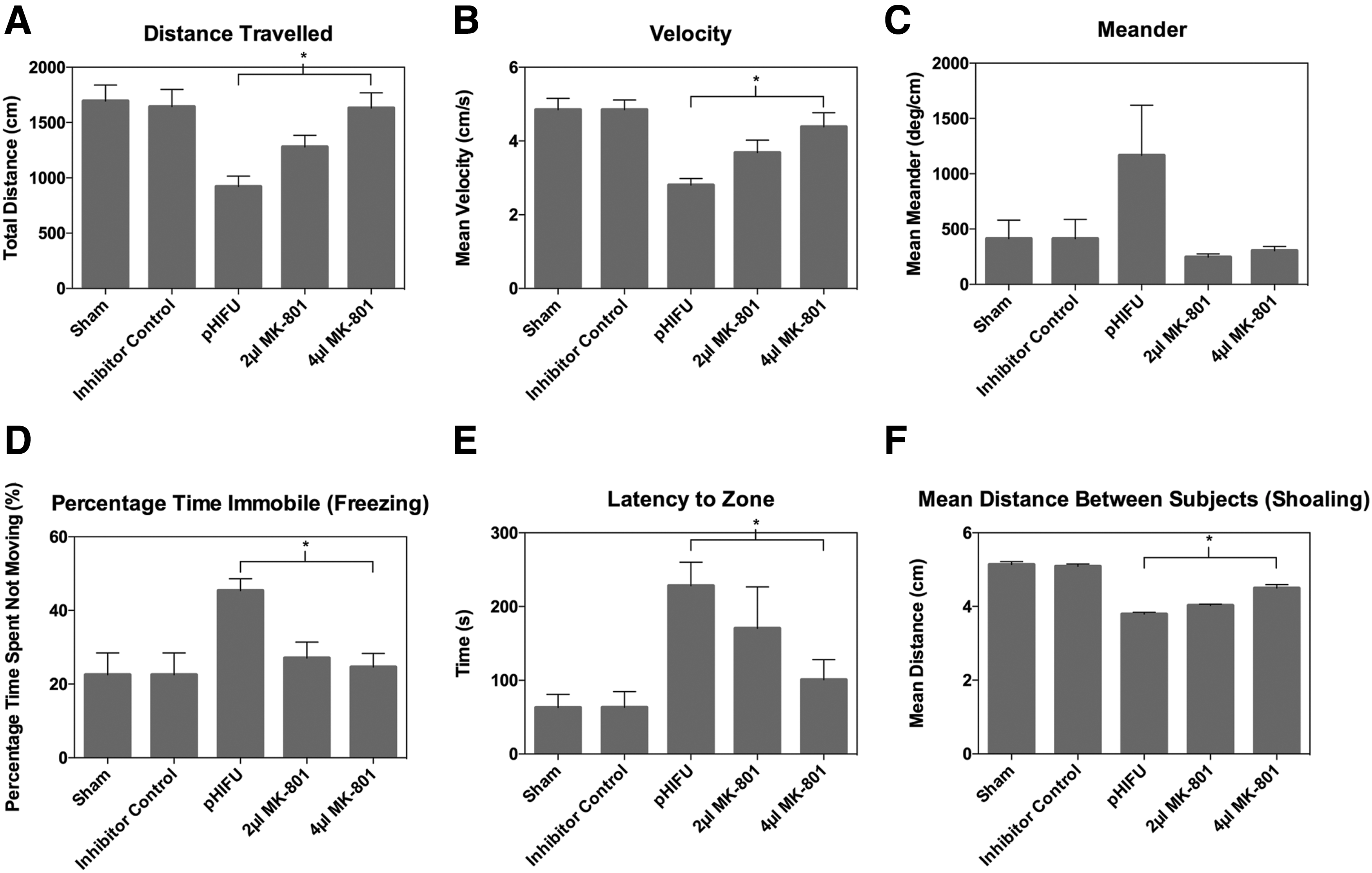
Inhibitor injection: NTT. Summary of behavioral data for n = 6. Data represent means ± SEM. Effects of MK-801 pre-treatment on behavior following pHIFU.
Discussion
In this characterization study we evaluated the potential of the adult zebrafish as a model to study TBI and evaluate therapeutic interventions. We considered several criteria in our evaluation of whether this model would add value in the study of TBI. These criteria were: 1. The ability to measure functional readouts that demonstrated reliable changes after trauma. 2. These measures of injury co-varied with the severity of trauma. 3. The model demonstrated responsiveness of injury readouts to known positive controls of validated interventions in mammalian models of TBI.
The zebrafish injury model provides additional advantages over rodent models by introducing time and cost efficiencies in modeling and outcome testing. These advantages were achieved by means of a no-touch injury model, eliminating the need for invasive surgery to administer the trauma, and automated behavioral end-points, which could be quickly analyzed with video tracking software. Video recording and data analysis allows for rapid and extensive data collection using established software templates. The ability to record multiple arenas at once increases throughput efficiency and capitalizes on batch calculations to analyze data. In these ways the zebrafish model is highly amenable to high-throughput evaluation and is significantly less time- and cost-consuming than other animal model systems that require testing each subject individually. In terms of housing and rearing, zebrafish can be stocked at higher density and bred at higher frequency (on a weekly basis) compared with rodents, thereby reducing costs associated with animal husbandry.
TBI is not a single pathophysiological event but a complex disease process. 49,50 The construct, face, and predictive validity of a TBI model are of importance when implementing a model system that will have objective translational value. 14,51 Generally, face validity is the phenomenological analogy with the modeled condition, whereas construct validity indicates the model has a sound theoretical rationale. 52 Predictive validity dictates that the animal model must depict sufficient similarity to the human condition it is intended to simulate. In simple terms, predictive validity can be calculated in terms of reliability and relevance. 53 Traditionally, an animal model is considered to be valid, if it “resembles the human condition in aetiology pathophysiology, symptomatology and response to therapeutic interventions.” 54 By extension in our zebrafish model, we demonstrate that the adult zebrafish injury model shares similarities with the mammalian rodent system that are widely accepted as having construct, face, and predictive validity.
Modeling the entire spectrum of TBI pathophysiology in an animal is not possible given the heterogeneity of the injury itself. As such, select aspects of TBI have successfully been reproduced in a range of animal models. 3,5,7,9 The targeted ultrasound insult was used to produce a form of closed-head, brain-specific injury. This mode of injury was able to reproduce molecular changes and associated behavioral impairments with a high degree of similarity to mammalian models of TBI. This adult zebrafish TBI model thus exhibits predictive validity—the key ability of animal models to accurately and reliably depict the human condition. Our adult injury system demonstrated reproducibility in outcome as well as higher-throughput capacity than rodent systems. As a whole, the model demonstrates key relevance to mammalian pathophysiology through well-defined cellular and behavioral outcome measures. Presumably, this model can be used to identify potential therapeutic compounds as well as targets associated with TBI.
Despite the advantages of this system, it is not a replacement for larger mammalian studies. Localized injury with pHIFU ultrasound method is not currently technically feasible due to size limitations of both the pHIFU wave and the size of the zebrafish brain. Also, tissue samples from specific brain regions are too small to allow region-specific analysis. As such, changes in protein expression may suffer from signal to noise artifacts, masking the actual magnitude of changes in protein expression within the traumatized regions. Further, in the current study we only examined the acute effects of injury on outcome. Whether zebrafish exhibit persistent neurological deficits is currently unknown, and how these time lines would correspond to mammalian pathophysiological events also remains to be determined. Moreover, the observed delayed mortality in a portion of the normothermic injury group at 5 and 10 days post-injury suggests that secondary injury mechanisms continue to exacerbate the primary injury response.
This suggests that zebrafish share similar pathophysiological traits in response to injury as seen in mammalian system in regards to secondary injury progression. It is unknown at this point, however, how the temporal sequence of events differ between species or whether the thresholds for survival differ between zebrafish and mammals undergoing comparable levels of brain trauma. The current model serves as a quick and cost-efficient means of generating empirical evidence about the efficacy of treatment prior to investment in a full mammalian study. Ease of drug delivery whether through water application or targeted microinjections makes this model a valuable translational bridge between in vitro and in vivo systems by simplifying the evaluation process but maintaining a degree of anatomical complexity through use of a whole organism. Despite these limitations, the ever growing library of transgenic reporter lines lends the model to numerous potential applications to study TBI pathophysiology.
Post-injury recovery times increased in a dose-dependent manner with increasing focal pressure as well as duration. These results suggest that our model is capable of reproducing aspects of mild TBI. Classification of injury severity in animal models, however, remains somewhat ambiguous and unstandardized across injury platforms. There remains no common scoring system for injury severity based on brief neurological examination widely adopted in animal models. 5 Here we rely on survival, functional outcome, and recovery to note similarities between our results and a general categorization of mild TBI previously described in animal and human cases.
The adult zebrafish pHIFU injury model demonstrates changes in white matter protein expression. Expression of axonal cytoskeletal proteins and activated caspase-3 were consistent with the response to trauma in rodent models of non-penetrating TBI. NF 160, a medium neurofilament that provides structural support and tensile strength to axons, 40 along with heavy and light neurofilaments, has been shown to accumulate in the axonal swellings that result from the axonal pathology of TBI. 40,55 Microtubule β-III tubulin is expressed almost exclusively in neuronal cytoskeletons and is essential for axonal growth and transport. 40 Microtubule disruption during secondary injury results in disrupted intra-axonal transport as well as axonal swelling and detachment. 44 We have previously reported increases in NF 200 in milder injury paradigms. 56,57 This change in expression is also supported in our zebrafish injury model. It is interesting to note that heavy neurofilament and microtubule expression preceded expression of caspase-3 activation and β-APP accumulation. These results indicate a progressive evolution of mild injury mechanisms in the zebrafish. More work remains to be done to corroborate these events in rodent models.
The zebrafish provided useful behavioral outcome measures that are difficult to assess in rodent models. Specifically, the evaluation of shoaling (grouping) interactions have high relevance for the evaluation of mild brain injury deficits. Social dysfunction is often apparent in psychiatric disorders as well as trauma and can be influenced by genetics, chemicals, environment, and disease. 58,59 Zebrafish are highly social animals that display well-documented characteristic social behaviors. 36,60,61 In addition to the automated analysis systems available, and the sensitivity of the outcome measure to TBI and intervention, zebrafish are well suited for mild TBI studies. Importantly, these outcomes are analyzed in our model with minimal user intervention. Our results are consistent with reports indicating that anxiety as well as predatory stress increases grouping activities of zebrafish. 59,62 The use of the NTT and the shoaling test can combine as an effective strategy to evaluate zebrafish injury and behavior post-insult.
The adult zebrafish injury model was responsive to interventions that have previously been validated in mammalian systems. Zebrafish are poikilothermic, meaning their internal temperature varies according to external environment changes and the species exhibits a wide tolerance to water temperature ranges. 63 By lowering water temperature in our model by 3°C we were able to rescue behavioral deficits post-injury. These results indicate zebrafish physiology, despite differences in their temperature maintenance ability in relation to homeothermic species, is responsive to temperature intervention in a manner that is consistent with mammals in terms of improving outcome after neurotrauma.
In addition to the proof-of-concept of thermoregulation intervention following TBI, the efficacy of MK-801 treatment further demonstrates the model's similarity in pathophysiological response to injury compared with mammalian models. Despite being clinically ineffective, the targeting of NMDA-receptors with the use of MK-801 demonstrates the similarity in secondary cellular injury pathways between the zebrafish and rodent models.
Conclusion
Our results indicate that the zebrafish response to targeted pHIFU brain trauma exhibits mechanisms of secondary injury similar to mammalian pathophysiology after TBI. We report the development of a novel vertebrate animal model of TBI in the zebrafish with high relevance and applicability for TBI studies. This is an important novel tool for the implementation of medium-throughput assays to reduce the time and cost associated with pre-clinical drug evaluation for TBI.
Footnotes
Acknowledgments
We would like to thank Arthur Worthington from the Advanced Biomedical Ultrasound Imaging and Therapy Laboratory, Department of Physics, Ryerson University, for his technical help in developing and testing of the pHIFU equipment. Partial funding for pHIFU equipment development was provided by an Ontario Research Fund-Research Excellence (ORF-RE) grant that was awarded to J. Tavakkoli. The authors acknowledge funding from the Brain Canada Foundation (Dr. Wen), Ontario Centres of Excellence (OCE)-CQDM (Dr. Wen), and the Canada Foundation for Innovation (Dr. Wen).
Author Disclosure Statement
No competing financial interests exist.