
Abstract
Cardiovascular disease is the leading cause of death for individuals with spinal cord injury (SCI). Because of a lack of a standardized and accessible animal model for cardiovascular disease after SCI, few laboratories have conducted pre-clinical trials aimed at reinstating descending cardiovascular control. Here, we utilized common contusion methodology applied to the midline of the upper-thoracic cord of adult Wistar rats accompanied with telemetric blood pressure monitoring and FluoroGold retrograde neuronal tracing, as well as lesion site and lumbrosacral afferent immunohistochemistry. We demonstrate widespread cardiovascular (i.e., impaired resting hemodynamics, autonomic dysreflexia) and hindlimb dysfunction at 1 month post-injury. Further, we provide a description of the neuroanatomical changes that accompany cardiovascular abnormalities. Specifically, we describe 1) the injury site including white matter sparing as well as lesion volume, and their correlations to cardiovascular as well as motor outcomes; 2) the severity of injury-dependent changes in sympathoexcitatory medullary neuron spinal connectivity, as measured using FluoroGold tracing; and 3) the extent of aberrant afferent plasticity within the lumbosacral region of the spinal cord, which has been linked to the development of autonomic dysreflexia. We believe that this model, which utilizes equipment common to numerous SCI laboratories, can serve as a research standard for studies specifically aimed at investigating autonomic neuroprotective and regenerative strategies following SCI.
Introduction
S
The major mechanisms purported to underlie cardiovascular dysfunction after SCI broadly relate to changes within the medullary cardiovascular control center (i.e., rostral ventrolateral medulla [RVLM]) and the extent of neural damage at the site of injury. The disconnection of descending sympathoexcitatory neurons from the RVLM to sympathetic pre-ganglionic neurons is associated with loss of descending sympathetic control, 7 a series of plastic changes including the transient atrophy of pre-ganglionic cell bodies, 8 as well as the development of intraspinal plasticity. 9 –11 Therefore, therapies aimed at restoring or preserving maximal amounts of descending sympathetic axons through the lesion site may prove fruitful in maintaining cardiovascular control after SCI. This may be achieved by enhancing supraspinal control and preventing aberrant plasticity in the lumbosacral spinal cord, ultimately decreasing this population's dramatic abnormalities in cardiovascular control. 12
Driven by the advancement in pre-clinical models, the re-establishment of functional behavior following experimental SCI has seen significant advancement in recent years. 12 –14 Because of the high proportion of blunt injuries in the human population, 15 most investigations into restorative treatment following experimental SCI have utilized contusion models. 13,16,17 The development of these models in rodents and higher mammals has aided the translational process and allowed for testing of potential treatments through pre-clinical studies. Despite these advances, the implementation of promising therapies still has significant barriers, as there is often conflicting evidence, insufficient mechanistic insight surrounding therapeutic agents, or little success upon clinical translation. 13,18,19 Therefore, the development of models that achieve widespread functional impairments (i.e., motor/sensory/autonomic) and are aligned with the common clinical pattern (i.e., high thoracic/cervical) are critical to synthesizing findings across laboratories. Further to this, the functional behavioral outcomes typically studied (i.e., motor/sensory behavior) represent the most complex and evolutionarily mammalian functions. Conversely, autonomic function is an evolutionarily old system, governed by a few medullary and pontine centers, and requires an exponentially lower number of descending projections necessary for normal control opposed to the motor/sensory system. 7,20,21 This raises the possibility that the cardiotherapeutic benefit of neuroprotective or regenerative therapies previously studied in the SCI population may have been missed as a result of only examining motor/sensory function.
Given that recovery of autonomic function is rated among the highest health priorities for individuals with SCI, 22 there continues to be a paucity of research investigating therapies to improve these functions. 12 This is in large part because of a lack of a standardized model across laboratories to study autonomic dysfunction after SCI. Previous experiments investigating the mechanisms underlying autonomic dysfunction following SCI have typically used high thoracic complete transections, 5,6,10,23 –27 clip compression, 28 –32 or contusion models. 33 Moreover, although much of the pre-clinical SCI field is moving toward cervical contusions, these models spare a significant amount of white matter because they are unilateral 34 –39 or because of their lesser degree of severity, 40 –44 which is difficult to increase without causing dramatic mortality. Therefore, these models are either unlikely to result in autonomic impairment or ethically problematic. Although transection models have been critical in providing numerous insights into the development of AD, cardiac impairments, cerebrovascular abnormalities, and other autonomic complications, they are limited in their applicability for neuroprotective strategies, as they do not provide a mechanism of injury congruent with such strategies. Further, clip compression models are difficult to apply across a wide array of laboratories because of the need for specialized equipment. Lastly, although a previous T3 contusion model examined hemodynamics, no data were included to characterize the AD response to this injury, 33 which is important as AD is likely one of the most life-threatening autonomic dysfunctions that occur and, therefore, is often the autonomic clinical target. Therefore, we sought to develop a clinically relevant high thoracic contusion model to study cardiovascular dysfunction after SCI, including AD, using the Infinite Horizon (IH) impactor in order to provide a model that can easily be accessed by numerous laboratories.
Herein, we describe the attributes of a consistent model of high thoracic contusion SCI that resulted in cardiovascular (i.e., impaired resting hemodynamics, AD) and hindlimb dysfunction. Further, we describe a subset of histological changes that may contribute to the observed cardiovascular abnormalities. Specifically, we describe 1) the lesion volume, white matter sparing, and locomotor functional outcomes; 2) the severity-dependent difference in sympathoexcitatory medullary neurons; and 3) the extent of aberrant afferent plasticity within the lumbosacral spinal cord, which has previously been linked to the development of AD.
Methods
Ethical approval
All procedures were conducted in accordance with the Canadian Council for Animal Care. Ethical approval was obtained from the University of British Columbia.
Experimental design
Twenty-seven male adult Wistar rats (age = 9 weeks; weight = 300–450 g; Harlan Laboratories, Indianapolis, IN) were used in this study, as we and others have described the reliable development of AD in this species, in contrast to Sprague–Dawley rats. 45 –48 Rats were randomly assigned to one of three groups: sham-injured control (SHAM; n = 5), moderate SCI (MODERATE; n = 5), or severe SCI (SEVERE; n = 5) (details provided subsequently). Following SCI surgery, animals underwent behavioral assessment according to the Basso, Beattie, and Bresnahan (BBB) method at day 7 and day 30. On day 31, animals were implanted with a telemetric blood pressure monitor in the carotid artery. Basal hemodynamics and the severity of AD in response to colorectal distension (CRD) were assessed. All animals were then immediately perfused and used for spinal cord lesion characterization and colonic afferent sprouting quantification, and to determine the number of remaining adrenergic-positive cell bodies within the RVLM.
In order to confirm the immunohistochemistry findings in the RVLM, an additional 12 animals were used to perform retrograde axonal tracing to the RVLM (SHAM, n = 2; MODERATE, n = 5; SEVERE, n = 5). At day 31, these animals underwent a spinal cord re-transection at the TVII vertebral level in order to place FluoroGold (FG) for retrograde sympathoexcitatory neuron tracing and subsequent cell-body quantification in the RVLM (details provided subsequently). No other behavioral or histological analyses were conducted on these animals.
Spinal cord surgery and animal care
Surgery and animal care procedures were conducted according to the Autonomic Research Laboratory's standard operating procedures. 23 Three days prior to surgery, animals were provided a prophylactic dose of enrofloxacin (Baytril; 10 mg kg−1, s.c., Associated Veterinary Purchasing [AVP], Langley, Canada). On the day of spinal cord contusion, animals were anesthetized (initial chamber induction at 4% isoflurane with 2l min−1 oxygen, followed by maintenance on a Bain system at 1.5–2% isoflurane with 1.5–2l min−1 oxygen), and preoperatively administered enrofloxacin, buprenorphine (Temgesic; 0.02 mg kg−1, s.c., McGill University), and warmed lactated Ringer's solution (5 mL, s.c.) while the surgical site was prepared. A dorsal midline incision was made in the superficial muscle overlying the C8-T3 vertebrae. A T3 laminectomy was performed to expose the T3 spinal segment. The spinal column was then stabilized by clamping the TII and TIV spinous bodies with modified Allis forceps. Each forceps was secured in place by locking universal joints, which were mounted on a secure metal stage that the animal was placed upon. The animal was then placed such that the impactor tip (2.5 mm) of an IH Impactor (Precision Systems and Instrumentation, LLC, Fairfax Station, VA 49 ) was directly over the exposed T3 dura. Care was taken to ensure the tension and alignment of the vertebral column was consistent. Further, the alignment of the impactor tip in relation to the T3 segment was confirmed as midline by two separate experimenters according to the surface anatomy. In addition, a surgical microscope was positioned from the side such that both experimenters could more easily visualize the position of the impactor tip. Once properly aligned, the impactor tip was lowered until contact with the dura was made, and then retracted 2 mm above the dura. It was at this point that SHAM animals were removed from the surgical stage and subsequently sutured. For spinal cord-injured animals, injuries were induced by driving the impactor tip downward with either 200 kdyn (MODERATE) or 400 kdyn (SEVERE) of predefined force. The IH impactor was set to dwell on the injury site for 5 sec prior to retracting to enhance injury severity. The wound was subsequently irrigated with saline, and the muscle and skin were closed with 4-0 Monocryl (Ethicon, USA) and 4-0 Prolene sutures (Ethicon, USA), respectively. Animals recovered in a temperature-controlled environment (Animal Intensive Care Unit, Los Angeles, CA). Animals were then administered enrofloxacin (10 mg kg−1, s.c..), buprenorphine (0.02 mg kg−1, s.c.) and ketoprofen (5 mg kg−1, s.c.) every 12 h for 3 days postoperatively. Bladders were manually expressed three times per day until spontaneous bladder voiding was restored (∼1 week post-injury in SEVERE animals).
Determination of injury parameters (pilot work)
Based on previous work, 28 we know that autonomic dysfunction is often accompanied by low BBB scores (i.e. <4). Therefore, in order to establish the impact severity necessary to elicit autonomic dysfunction, we sought an injury severity that would result in little hindlimb motor recovery (i.e., no weight-supported stepping). At 300 kdyn, we observed near immediate recovery of paw placement after injury. Based on previous incomplete models of autonomic dysfunction, it was expected that this level of function was likely to result in little to no autonomic dysfunction. 28 As such, we increased the severity of injury by adding 5 sec of dwell time in order to obviate further increases in force. Despite this, we still observed hindlimb recovery and paw placement within 2 days post-injury. Lastly, we increased the level of force to 400 kdyn and included the 5 sec of dwell time (SEVERE). At 1 week post-injury, we observed only uncoordinated slight hindlimb movements (i.e., BBB score <4). We therefore opted to move forward with this injury severity and included an equivalent injury, with half the force (i.e., 200 kdyn) as our MODERATE injury.
BBB functional behavior assessment
Hindlimb locomotor function was assessed using an open field test according to the BBB method. 50 Briefly, rats were acclimatized to the testing area for three sessions of 4 min 1 week prior to baseline testing. Rats had their bladders expressed prior to the session and were then placed in a 1.5 × 1.5 m box 53 for 4 min while two experienced, blinded experimenters scored hindlimb locomotor function. Animals were tested 1 day prior to contusion, and then 7 days and 31 days post-contusion (prior to telemetry implantation), always at the same time of day. Scores and subscores were averaged across the left and right hindlimbs.
Implantation of telemetry device
At day 31, animals were implanted with a telemetric monitoring device. Prior to surgery, the telemetry devices were prepared for implantation, according to the manufacturer's guidelines (Telemetry Research, USA). Animals were anesthetized using isoflurane (initial chamber induction at 4% isoflurane with 2l min−1 oxygen, followed by maintenance on a Bain system at 1.5–2% isoflurane with 1.5–2l min−1 oxygen), the hair on the neck was shaved, and the surgical site was cleaned with alcohol and iodine. 5 The left common carotid artery was exposed and isolated from the internal jugular vein using blunt dissection. The carotid artery was then permanently occluded rostral, and temporarily occluded caudal to the implantation site using 4-0 silk sutures. A small incision was made in the artery wall using a bent tip 20 gauge needle, and the telemetry device was guided into the lumen and advanced caudally to ∼5–6 mm rostral to the aortic arch. The catheter was secured with a small amount of tissue adhesive and two 4-0 silk sutures, and the body of the transducer was placed between the scapulae. The muscle and skin were closed using 4-0 Monocryl (Ethicon, USA) and 4-0 Prolene (Ethicon, USA) sutures, respectively. Animals were then transferred to a temperature-controlled environment (Animal Intensive Care Unit, Los Angeles, CA) for 90 min for the effects of isoflurane to dissipate. Surgeries were completed at approximately the same time of day for all animals.
Resting hemodynamics and autonomic dysreflexia
Ninety minutes following telemetry implantation on day 31, and at a similar time of day for all animals, animals were transferred to a SmartPad (TR181; Millar Telemetry Research, TX), where resting beat-by-beat arterial pressure and heart rate (HR) were recorded for 10 min, converted from analog to digital (PowerLab; AD Instruments, CO), and sampled at 1000 Hz (LabChart; AD Instruments, CO). Ninety minutes was used to allow sufficient time (half-life = ∼5 min) for isoflurane to be metabolized out of the circulation, because of its known effects on blood pressure. 33,52 Prior to baseline recordings, bladders were expressed to avoid any confounding effects of distension. Systolic, diastolic, and mean arterial pressure (MAP) were then derived on a beat-by-beat basis over 10 min to establish resting hemodynamics. Following resting hemodynamic recordings, an experienced researcher who was blinded to injury severity initiated CRD (i.e., induction of AD) according to previously described methods. 53 Briefly, a latex 2 mm balloon-tipped catheter (AA6110-COLOPLAST Folysil “Foley” pediatric two way indwelling catheter, straight, 10 French, 3 mL, 35 cm, latex free) was inserted 2 cm into the rectum and secured to the tail with tape. This procedure was performed without restraint, even for animals with some hindlimb function. Animals were covered with a towel to reduce any unneeded stimulation and to reduce any large movements, which can affect the blood pressure recording. Following a baseline recording (10 min), the balloon was slowly inflated with 2 mL of air over 10 sec, and maintained for 60 sec. Following 60 sec of distension, the air was released and the animal rested for 5 min. This procedure was repeated a second time. Hemodynamic parameters were recorded before (baseline; 10 min), during (distension; 60 sec), and after (recovery; 60 sec) balloon inflation. For each animal, the difference in systolic blood pressure (SBP), MAP, diastolic blood pressure (DBP), and HR from baseline and the peak value during distension was subsequently calculated for each trial, and averaged across the two trials. Data were averaged every 1 sec using a rolling average, and the peak was calculated as the highest value during the distension phase. 46 Finally, trials were time locked according to the beginning of distension, and averaged across animals, to produce a representative trace. However, this trace was not used to calculate any peak or delta values, because of potential underestimation of such indices arising from between-animal differences in the temporal AD response.
FG placement
Animals (SHAM, n = 2; MODERATE, n = 5; SEVERE, n = 5) underwent a transection of the spinal cord at T7 at day 31 post-injury. 54 Pre- and postoperative care were identical to what had been provided for the original contusion injury (described previously). A TVII laminectomy was performed following a dorsal midline incision and blunt dissection of superficial musculature. The T8/9 segment of the spinal cord was completely transected using microscissors, and completeness was confirmed by two separate experimenters. Following hemostasis, Gelfoam soaked in 10 μL of 4% FG (Fluorochromes Inc., CO) in sterile saline was placed against the rostral stump of the transected spinal cord. The muscle and skin were closed using 4-0 Monocryl and 4-0 Prolene sutures, respectively. Animals were euthanized and perfused 8 days following FG injection to allow sufficient time for retrograde transport to the RVLM.
Tissue processing, histology, and immunohistochemistry
Immediately following resting hemodynamic and AD assessments, or on day 8 post-FG injection, animals were overdosed with chloral hydrate and perfused transcardially with 500 mL of 0.1 M phosphate-buffered saline (PBS), followed by 500 mL of 4% formaldehyde in PBS. Brains, lesion sites (± 4 mm of epicenter), and lumbar enlargements were immediately dissected and post-fixed for 24 h and then transferred to 20% sucrose for cryoprotection. 46,55 A segment of medulla oblongata was dissected from the brain beginning 1 mm caudal to the obex and spanning rostral 5 mm in order to isolate the RVLM. 54 Each tissue was embedded in cryomatrix, frozen in liquid nitrogen, and stored in −80°C for histological and immunohistochemical investigation. Medulla oblongatae were cross-sectioned at 30 μm in the cryostat, with sections mounted on alternating slides (Fisherbrand Superfrost; six sections/slide) to create paired slide sets (total of 15 sets per medulla). Thus, the inter-section distance was 60 μm, preventing the possibility of double-counted RVLM cell-bodies, which span only ∼30 μm. 54 Injury site sections were cross-sectioned at 20 μm and mounted on sets of 10 slides, with eight sections per slide (total 8 sets of slides) and an inter-section distance of 200 μm. 51 L6/S1 (i.e., innervation to the rectum) was identified using the appropriate spinal ganglia prior to dissection, and sections were serially cut at 20 μm. 5
All sections were prepared for immunohistochemistry according to the following methods. 51 Sections were thawed at room temperature for 1 h, at which time a hydrophobic barrier was drawn. Sections were then rehydrated in 0.1 M PBS for 10 min then incubated with 10% normal donkey serum for 30 min, followed by incubation with primary antibodies overnight at room temperature. The following morning, sections were washed with 0.1 M PBS three times (10 min each), incubated with secondary antibodies for 2 h, and then washed in 0.1 M PBS three times (10 min each). Lastly, sections were cover-slipped using Immunomount and stored at 4°C. The following primary antibodies were used: chicken anti-myelin basic protein (MBP) (1:200, Aves, MBP), goat anti-glial fibrillary acidic protein (GFAP) (1:1000, Dako), guinea pig anti-neuronal nuclei (NeuN) (1:500, Millipore, ABN90P), mouse anti-dopamine β-hydroxylase (DBH) (1:500, Millipore, MAB308), and goat anti-calcitonin gene-related peptide (CGRP) (1:200, Abcam, ab36001). Secondary antibodies included: Alexa Fluor 647 Donkey Anti Goat (1:200; Jackson ImmunoResearch; 705-606-147), Alexa Fluor 488 Donkey Anti Mouse (1:200; Jackson ImmunoResearch; 715-546-151), Alexa Fluor 647 Donkey Anti Guinea Pig (1:200; Jackson ImmunoResearch; 706-606-148), Alexa Fluor 488 Donkey Anti Chicken (1:200; Jackson ImmunoResearch; 703-546-155), and Alexa Fluor 488 Donkey Anti Goat (1:200, Invitrogen, A11055).
Immunofluorescence was imaged digitally using an Axioplan 2 microscope (Zeiss) using ZEN 2 Blue software (Zeiss). Images were digitally processed using Photoshop (Photoshop CC) or ImageJ (ImageJ National Institutes of Health [NIH]) software.
Image analysis and histological quantifications
Lesion site analyses
Images were taken in quadrants at 5 × , and prepared for analyses using Photoshop CC. Cord size and lesion area were quantified (GFAP channel) every 400 μm from 4.0 mm rostral to 4.0 mm caudal to the injury epicenter. The injury epicenter section was identified by a blinded rater, based on the section with the least intact GFAP signal. Lesion area was manually outline based on the following definition: GFAP-negative or GFAP-positive area with disrupted or abnormal cytoarchitecture.
56
Care was taken to avoid inclusion of any artifacts. Myelin preservation (i.e., white matter sparing) was estimated by manually outlining based on the presence of an MBP-positive area with normal or near-normal cytoarchitecture. The total number of pixels within each section was measured using Photoshop CC. Lesion volume was then calculated according to the following formula:
Neuronal preservation and astrogliosis were qualitatively evaluated using the NeuN and GFAP signals, respectively. All individuals conducting analyses were blinded to grouping details.
RVLM localization and neuronal counts
The general location of the RVLM was first determined using a cresyl violet stain. Briefly, sections were immersed in cresyl violet for 20 min, and dehydrated through successive alcohol washes before being placed in xylene and cover-slipped. The nucleus ambiguous and hypoglossal nucleus were then identified and used as landmarks for the identification of the RVLM (Fig. 1). Paired slides of the identified sections were then prepared for immunohistochemistry according to established methodology. 54 Immunofluorescent images were imaged at 5 × over the ventrolateral column where the RVLM is localized. Neurons labeled with DBH were counted manually for the entirety of the RVLM (15 sections) for each animal.
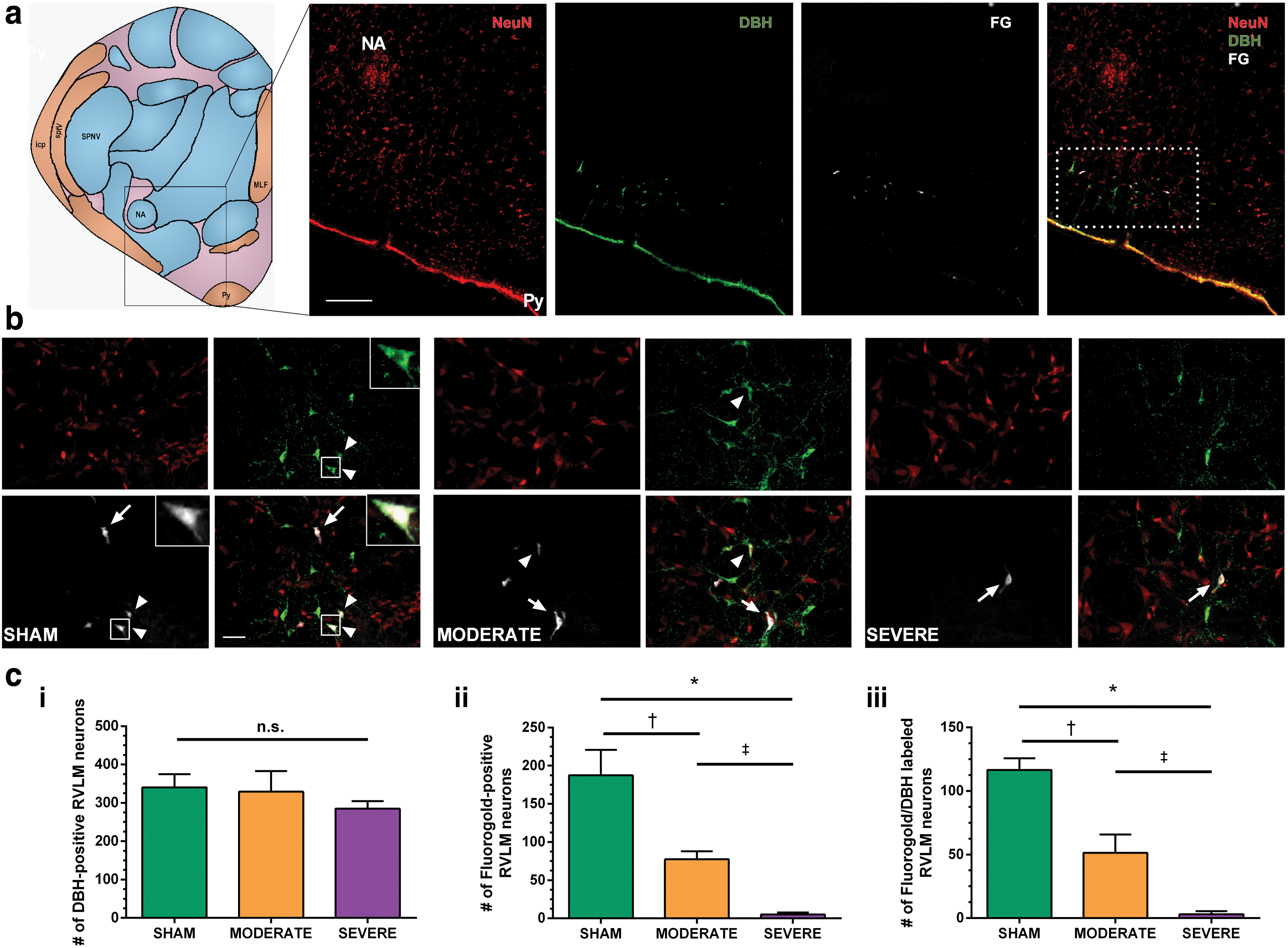
Anatomical localization of the rostral ventrolateral medulla (RVLM) and examination of the severity dependent difference in descending sympathoexcitatory axons of FluoroGold (FG) traced animals.
For FG animals, an identical procedure was completed for the DBH analyses; however, FG-labelled neurons were also individually counted. Further, co-labelled neurons (DBH, FG) were identified by examining both channels, and counted on ZEN 2 Blue software (Zeiss) to estimate the number of descending sympathoexcitatory neurons crossing the injury site. For all neuronal counts, the total number of counted neurons on both the left and right side was tabulated.
CGRP+ axon density quantifications
Five images per animal were taken on five individual sections, with randomization of left and right side of the cord, and 20 μm inter-section distance. Images were taken at 20 × over laminae III-V (identified beginning at the deep border of lamina II and stopping at the central canal) at the L6/S1 spinal level and analyzed using ImageJ. Images were converted to eight bit and a threshold (Default Dark) was set. Particle analysis was then used to determine the percent area labelled as CGRP+. 5 The average across all images per animal was then used.
Statistical analysis
All statistical analyses were conducted using SPSS 19/20 (IBM, Armonk, NY) and all graphical representations of data were made using Prism 6 (GraphPad, La Jolla, CA) in combination with Photoshop CC 2014 release (Adobe, San Jose, CA). All behavioral and cardiovascular data were statistically analyzed using independent-samples t tests, one way analysis of variance (ANOVA) or repeated-measures ANOVA, as appropriate, and are presented in text as mean ± standard deviation and visualized using mean ± standard error. Significant main or interaction ANOVA effects, as appropriate, were further probed using Tukey's honest significant difference (HSD) post-hoc testing. Between-group differences in the severity of AD were analyzed using an independent samples t test (MODERATE/SEVERE). Correlational analyses were conducted using parametric statistics (Pearson's correlation coefficient [r]). The significance level for all statistical tests was set at p < 0.05.
Results
IH Impactor produces consistent biomechanical output in T3 spinal cord contusion
We report that a spinal cord contusion can be performed at the T3 spinal segment using the IH impactor. The biomechanical outputs from the IH were consistent across all metrics (i.e., force, displacement, velocity) for both the SEVERE and MODERATE groups. As expected, the SEVERE injury parameters were more extreme than those for the MODERATE injury in terms of force (400 ± 14 vs. 210 ± 2.5; p < 0.001) and displacement (1692 ± 69 vs. 1120 ± 59; p < 0.001); however, the velocity of impact was similar (121 ± 2.2 vs. 120 ± 1.4; p = 0.469).
T3 spinal cord contusion impairs resting hemodynamics
SBP was significantly reduced at rest compared with in SHAM animals (Fig. 2a; Table 1) in both MODERATE (p = 0.015 vs. SHAM) and SEVERE (p = 0.040 vs. SHAM) injured animals. MAP was also significantly lower after MODERATE SCI (p = 0.016 vs. SHAM) and SEVERE SCI (p = 0.046 vs. SHAM), consistent with previous reports using transection models. 5,24,58 In addition, we observed a concomitant increase in HR compared with SHAM following MODERATE SCI (p = 0.017) and SEVERE SCI (p = 0.005). In contrast, we did not observe any significant reductions in resting DBP after either MODERATE SCI (p = 0.095 vs. SHAM) or SEVERE SCI (p = 0.111 vs. SHAM). No significant differences between MODERATE and SEVERE SCI were observed for any basal hemodynamic measures (all p > 0.05).
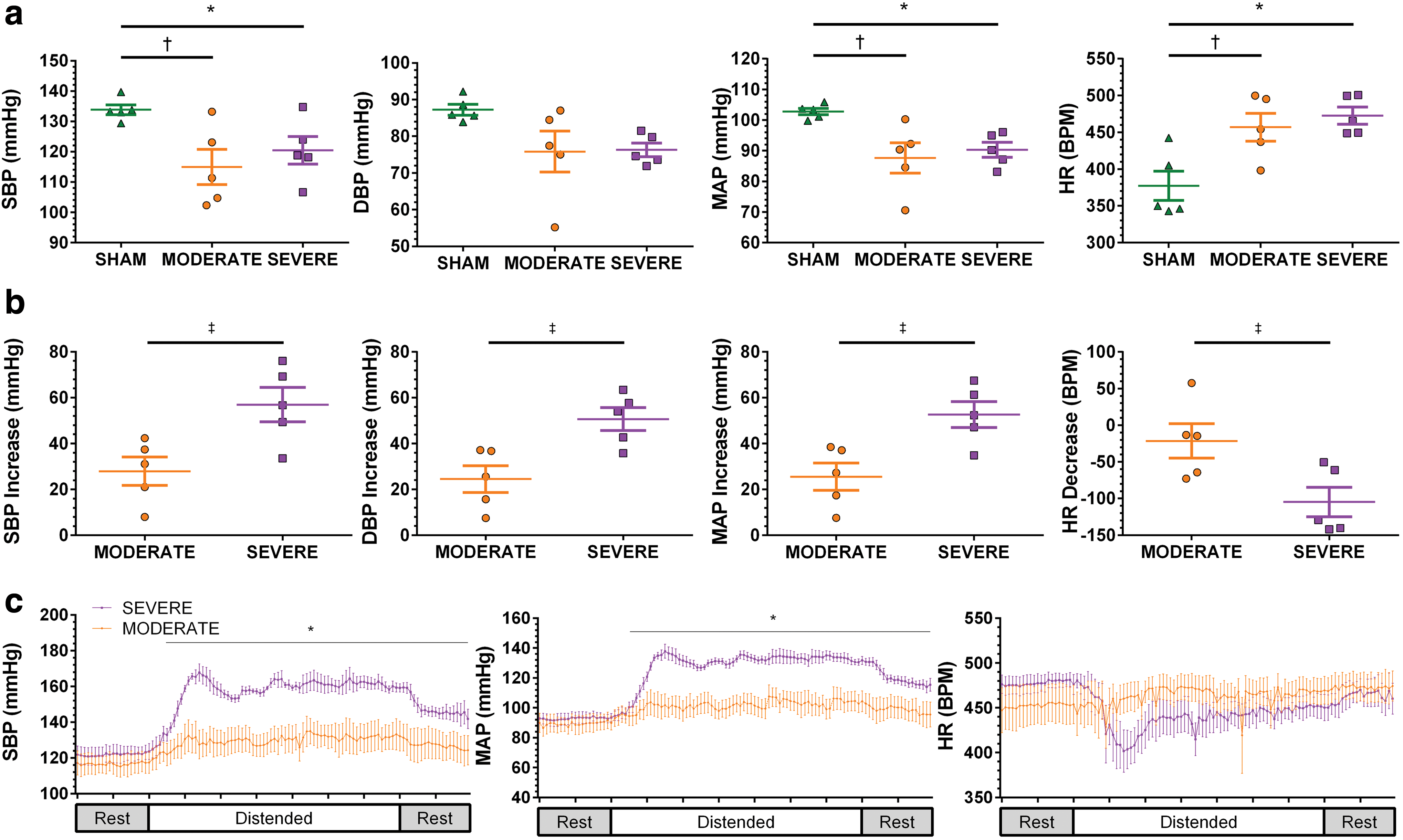
Resting hemodynamic and autonomic dysreflexia responses.
Data are presented as mean ± standard deviation.
p < 0.05 SHAM versus SEVERE; † p < 0.05 SHAM versus MODERATE; ‡ p < 0.05 MODERATE versus SEVERE.
SBP, systolic blood pressure; DBP, diastolic blood pressure; MAP, mean arterial pressure; HR, heart rate. Delta reflects change from baseline.
T3 spinal cord contusion induces AD
In all cases, SEVERE SCI induced a greater hypertensive and bradycardic response to CRD than did MODERATE SCI (Fig. 2b; Table 1). SBP maximally increased from baseline in the SEVERE SCI animals and was significantly greater than the response in MODERATE SCI animals (p = 0.017). This finding was similar for MAP (p = 0.011) and DBP (p = 0.009). Further, SEVERE SCI animals demonstrated a larger decrease in HR than did MODERATE SCI animals (p = 0.027). Time-locked group average SBP data revealed a significant interaction effect (p ≤ 0.001) whereby SEVERE injured animals' SBP rose significantly above that of MODERATE injured animals immediately after distension (all p < 0.01; Fig. 2c). We observed similar results for MAP (p < 0.01 following distension) but not for HR (ANOVA p < 0.001; Tukey HSD p > 0.05).
Lesion size, myelin preservation, neuronal preservation, and astrogliosis are severity dependent
MODERATE and SEVERE T3 contusion induced marked damage at the spinal cord lesion epicenter, which extended in both rostral and caudal directions (Fig. 3b, c). Qualitative analyses of both GFAP and NeuN immunoreactivity suggest that SEVERE SCI resulted in enhanced astrogliosis extending both rostrally and caudally. Further, few neuron cell bodies were observed near the epicenter. Conversely, MODERATE SCI animals revealed increased GFAP density at the epicenter (Fig. 3a), and neuronal cell bodies appeared as closer to the lesion epicenter. We observed significant decreases in spinal cord area compared with SHAM animals at 5 weeks following both MODERATE and SEVERE SCI (ANOVA p < 0.001), with MODERATE SCI animals showing recovered cord area ∼3.5 mm from the lesion epicenter (Fig. 3b). Lesion area was quantified every 400 μm, and a significant main effect was observed between MODERATE and SEVERE SCI, whereby SEVERE animals demonstrated significantly more lesion area (ANOVA p < 0.001). Lesion volume was then quantified and was significantly greater following SEVERE than following MODERATE SCI (16.0 ± 2.6 vs. 8.6 ± 0.97; p < 0.001; Fig. 3d). SEVERE SCI animals demonstrated significantly less white matter sparing (ANOVA p = 0.022; Fig. 3e). Individual animal lesion volumes were significantly correlated with impact force (R2
= 0.804; p < 0.001; Fig. 3g), displacement (R2
= 0.667; p = 0.007; Fig. 3j), In addition, AD severity was correlated to white matter sparing (R2
= 0.567; p = 0.012; Fig. 3f; Table S1) (see online supplementary material at
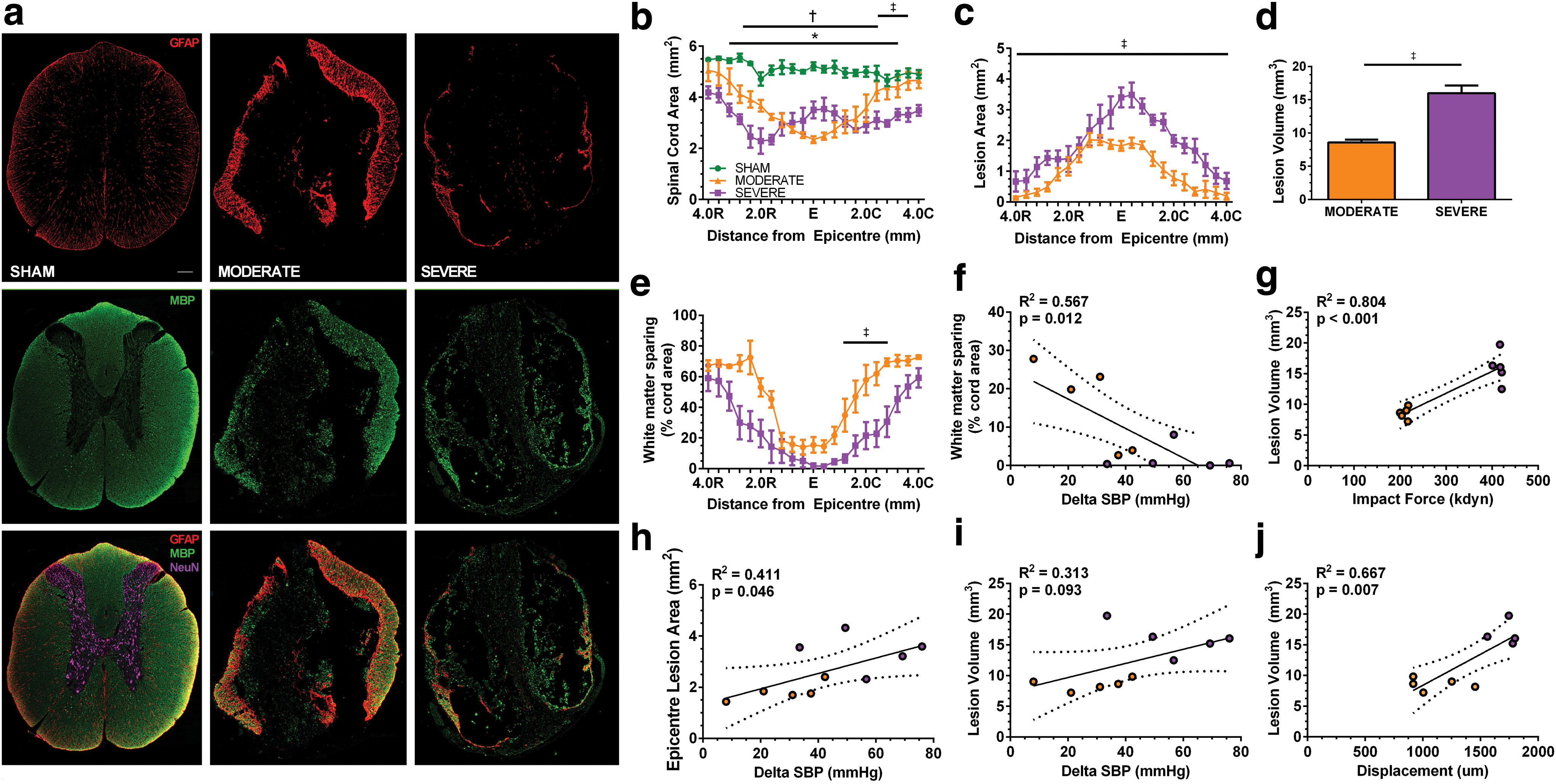
Lesion site characterization.
Sympathoexcitatory neurons projecting from the RVLM are impaired after T3 spinal cord contusion
The RVLM was located immediately anterior and medial to the nucleus ambiguous, caudal to the inferior olivary nucleus, and immediately rostral to the facial nucleus (Fig. 1a). Consistent with previous reports, 54,59 –62 the RVLM extended for ∼1000 μm and, therefore, fifteen 30 μm sections were taken for all analyses to estimate the number of neurons within the RVLM. For non-FG animals, there was a significantly lower number of DBH-positive neurons in the SEVERE SCI than in the SHAM animals (286 ± 32 vs. 379 ± 8; p = 0.011). In animals with FG placed below the level of injury, the number of DBH-positive neurons was not different between any groups (p = 0.149; Fig. 1ci ). In contrast, the number of FG-labelled neurons was significantly reduced in the SEVERE SCI animals (5 ± 2.8) compared with both MODERATE SCI (77 ± 10.7; p < 0.001) and SHAM (188 ± 33; p < 0.001; Fig. 1cii ) animals. Further, the the number of FG-labelled neurons in the MODERATE group was also significantly reduced compared with in the SHAM animals (p = 0.001). When examining DBH/FG co-labelled neurons there was a similar finding, in which SEVERE SCI animals (3 ± 2.5) had the fewest co-labelled neurons compared with both MODERATE SCI (51 ± 14.3; p < 0.001) and SHAM (117 ± 9.2; p < 0.001; Fig. 1ciii ) animals. MODERATE SCI animals also showed a significant reduction compared with SHAM (p < 0.001).
T3 spinal cord contusion induces dorsal horn afferent plasticity
Following SEVERE SCI, we observed a significant increase in the positive proportional area of CGRP+ axons within deep laminae (III-V) of the L6/S1 dorsal horns compared with both MODERATE SCI (1.56 ± 0.69 vs. 0.70 ± 0.08; p = 0.043) and SHAM (1.56 ± 0.69 vs. 0.58 ± 0.22; p = 0.023). No significant difference was observed between MODERATE SCI and SHAM (p = 0.913; Fig. 4).
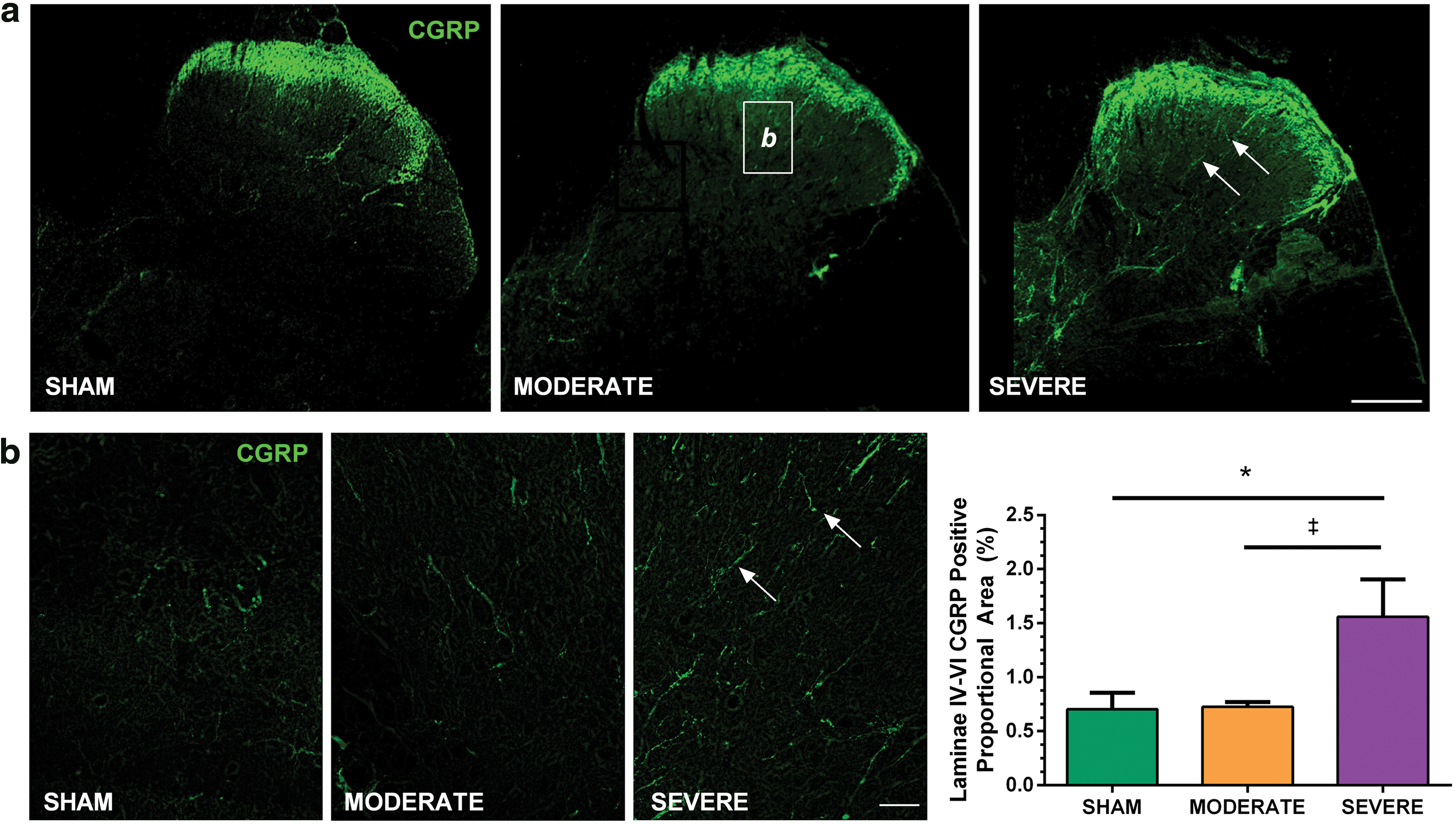
Calcitonin gene-related peptide (CGRP) immunohistochemistry in the L6/S1 dorsal horn following SHAM (n = 5), MODERATE (n = 5), and SEVERE (n = 4) SCI. One SEVERE SCI animal was excluded because of damaged tissue.
T3 spinal cord contusion induces severity-dependent decrease in functional motor behavior
Locomotor behavior was impaired after MODERATE and SEVERE SCI compared with SHAM. At 1 week post-injury, there were significant differences in total BBB score (maximal score = 21; Fig. 5a) between SEVERE SCI and SHAM (1.6 ± 1.1 vs. 18.9 ± 3.5; p < 0.001), and MODERATE SCI and SHAM (5.8 ± 1.3 vs. SHAM; p < 0.001), as well as between SEVERE SCI and MODERATE SCI (p < 0.001). Similar results were observed at 5 weeks, whereby there were significant differences in total BBB Score between SEVERE SCI (2.3 ± 1.9), MODERATE SCI (10.1 ± 1.0), and SHAM (19.3 ± 1.6; all p < 0.001). BBB subscore indicated that SEVERE SCI (0 ± 0) animals did not regain any weight-bearing ability on their hindlimbs at either 1 week or 5 weeks post-injury (Fig. 5b). At 1 week post-injury, MODERATE SCI animals displayed no weight-bearing ability (0 ± 0), but did regain small amounts by 5 weeks (1.6 ± 2.3). Both MODERATE and SEVERE SCI animlas were significantly different from SHAM (11.4 ± 1.5; all p < 0.001). Lastly, there was a significant correlation between the lesion volume and BBB score for MODERATE and SEVERE animals at the 5 week time point (R2 = 0.694; p = 0.003; Fig. 5c), as well as between delta SBP and BBB score (R2 = 0.555; p = 0.013; Fig. 5d).
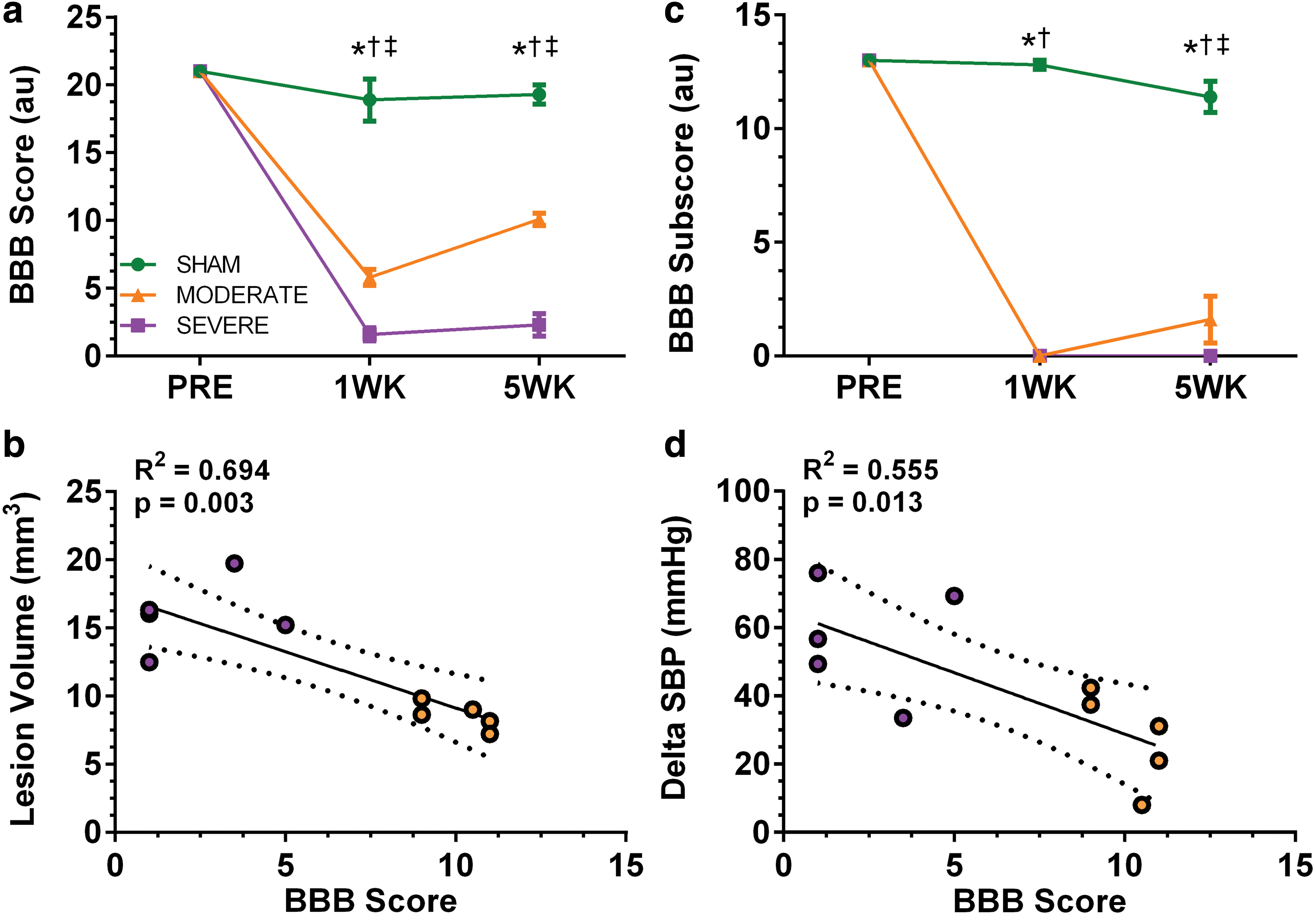
Overground locomotion of the animals was scored using the Basso, Beattie, and Bresnahan (BBB) scale (SHAM n = 5; MODERATE n = 5; SEVERE n = 5). There was a significant decline in locomotor function in both the MODERATE and SEVERE injury groups compared with SHAM, with the SEVERE group demonstrating even greater dysfunction in both the total score (
Discussion
Here, we present our findings that a T3 severe contusion SCI provided by an IH impactor produces a severity-dependent decline in autonomic and motor function in the rodent. Further, we provide neuroanatomical histological data underlying autonomic dysfunction after T3 contusion, and demonstrate novel relationships between injury site metrics and AD. The ability to model higher and more severe SCIs is of high importance within the pre-clinical SCI field. This new model may serve as a research standard to test different potential neuroprotective/neuroregenerative strategies and look at a variety of outcome measures including autonomic, locomotor, and sensory changes.
T3 spinal cord contusion impairs resting hemodynamics
A reduction in resting SBP and MAP has been demonstrated in a variety of animal models of SCI, 5,24,55,63,64 in addition to its long established impact in the clinical SCI population. 65,66 Resting hypotension reflects the decentralized state of the sympathetic nervous system after SCI and, therefore, is a critical outcome measure, as it has been linked to cardiac dysfunction 55 and cerebrovascular dysfunction, 6,67,68 and likely contributes to the SCI population's dramatic risk for heart disease and ischemic stroke. 3,69 We demonstrate here that both moderate and severe T3 spinal cord contusion induces decreases in both SBP and MAP. Although the absolute BP is difficult to compare across studies, here we demonstrate a 15 mm Hg reduction in SBP compared with sham animals. These decreases are in line with previous work utilizing both T3 transection models and clip compressions. 5,24,63 The mechanism of this decrease has been previously established to result from a decrease in resting sympathoexcitatory influence on sympathetic pre-ganglionic neurons responsible for vasomotor tone (i.e., T6-T12 spinal segments that provide major sympathetic output to the splanchnic vasculature). 70 This vasculature comprises the major branches of the celiac trunk, superior mesenteric artery, and inferior mesenteric artery, and is responsible for holding approximately one third of the resting blood volume in both rodents and humans. 70 Therefore, a disconnection between sympathoexcitatory neurons in the RVLM and pre-ganglionic sympathetic neurons responsible for controlling the vasomotor tone results in a decreased resting SBP and MAP. This mechanism is further supported by our finding of reduced counts of sympathoexcitatory RVLM (i.e., DBH-positive) neurons, with preserved axons extending below the injury (Fig. 1). That we observed a significant decrease in resting hemodynamics in both the MODERATE and SEVERE groups suggests that partial loss of descending sympathoexcitatory control over the splanchnic and lower limb vasculature beds may be sufficient to reduce systemic arterial tone. Moreover, the larger variability in the MODERATE SCI group may indicate differential loss of axons within the dorsolateral funiculus, leading to variability in descending control, in contrast to the more consistent SEVERE SCI group where there was near complete loss of all white matter. Our data also demonstrate that rodents with a T3 SCI of both moderate and severe severity exhibit an increase in resting HR compared with sham animals. An increase in resting HR following high thoracic SCI has previously been reported by us and others, 55,71 and has been purported to be a result of increased sympathetic activity to the myocardium, 71 which, importantly, leaves animals at higher risk for reperfusion-induced arrhythmias. 71
T3 spinal cord contusion results in AD
Clinically, AD is one of the most dangerous secondary complications that individuals with SCI experience. 2,65 Further, AD has long been the target of neuroprotective and regenerative therapies aiming to restore cardiovascular function. 12 Therefore, it is critical that SCI models aiming to investigate cardiovascular dysfunction exhibit clear and consistent AD. The current model demonstrated consistency in the AD response within both the MODERATE and SEVERE SCI groups, and provides a methodology accessible to many laboratories, the lack of which was a major limitation of previous models. 29 –32,72,73
In the current experiment, the SEVERE group demonstrated responses in line with previous work utilizing transection models 5,74 and exhibited blood pressure increases well above physiologically safe limits, 2 a critical determinant for a clinically relevant model. For example, clinically, SBP increases >180 mm Hg have been associated with cerebrovascular events (i.e., hemorrhagic stroke), and SBP values >200 mm Hg are associated with fatal outcomes. 2 Therefore, the AD responses evoked in response to CRD in this model represent a strong starting point for investigating further mechanisms and therapeutic options.
Lesion size, cavity size, myelin preservation, neuronal preservation, and reactive astrogliosis are severity-dependent
Following severe T3 spinal cord contusion, a lesion was formed that extended >4 mm in both caudal and rostral directions. This was accompanied by near complete abolishment of any spared white matter at the lesion epicenter, similar to a complete transection but with a critical difference in mechanism of injury. These findings are in line with previous studies utilizing severe clip compression models, in which the authors observed similar levels of tissue destruction and distance in injury site. 54,75 Although the white matter sparing was consistent in the SEVERE SCI group, we did see two outliers in the MODERATE SCI group, which demonstrated white matter sparing more in line with the SEVERE SCI groups. Further, these two animals demonstrated significant AD compared with others in the group, despite having BBB scores in line with the rest of the MODERATE group (i.e., 9 and 11). Moreover, one of the animals did demonstrate relatively low DBH neuron counts in the RVLM (274), similar to the SEVERE SCI group. However, this ultimately highlights our critical finding that the severity of AD appears to be significantly related to white matter sparing at the injury epicenter (Fig. 3f), which is a crucial piece of evidence when considering potential therapeutic options.
We observed only minimal GFAP-positive expression at the lesion epicenter in the SEVERE group. Conversely, there were relatively large of amounts of GFAP production at the lesion epicenter in the MODERATE injury group. Moving away from the epicenter, SEVERE SCI animals tended to exhibit increased levels of GFAP expression relative to SHAM animals. The overall increase in GFAP is a well-characterized phenomenon after SCI, and likely reflects both the dramatic hypertrophy and excess production of intermediate filaments (i.e., GFAP), 76 and the influx of neural progenitor cells from the surrounding spinal cord into the injury site, which then proliferate into astrocytes. 77 –80 These two mechanisms therefore account for what is known as the glial scar. This scar prevents axonal regeneration through the production of substances such as chondroitin sulfate proteoglycans. 76,81 In addition to the current findings surrounding GFAP, NeuN staining revealed that severely injured animals have no sparing of neuronal cell bodies at the lesion epicenter. This lack of neuronal cell bodies extended in both directions, reflecting the extensive damage and glial scar formation. Overall, severe T3 spinal cord contusion resulted in near complete ablation of all MBP and NeuN positive cells. This damage extended both rostral and caudal, and the full extent of the lesion volume was intimately related to the force of impact, the displacement of the cord, and the hindlimb function, as well as the severity of AD.
Sympathoexcitatory neurons projecting from the RVLM are impaired following T3 spinal cord contusion
Our main finding was that T3 spinal cord contusion resulted in a severity-dependent decline in both the number of FG-positive neurons within the RVLM and the number of co-labelled FG/DBH-positive neurons, similar to our previous report. 54 The RVLM has been extensively characterized for its impact on cardiovascular function and primary composition of C1 adrenaline-synthesizing neurons. 24,54,59 –62,82,83 For example, electrical and pharmaceutical stimulation of C1 adrenergic neurons within the RVLM increases blood pressure, 84,85 whereas pharmaceutical blockade of this area induces sympathetic hypoactivation. 86 –89 Adrenergic neurons within the RVLM project their axons to the spinal cord, which travel within the dorsolateral funiculus to stimulate sympathetic pre-ganglionic neurons. 7,59 As such, the maintenance of these axons around the site of injury is of critical importance in maintaining appropriate inputs to sympathetic pre-ganglionic neurons and preserving cardiovascular function after injury.
Although the raw number of RVLM neurons is difficult to compare across studies because of the variance of animal models, the methods of counting of neurons and the number of sections examined, as well as the chosen phenotypic marker, the relative decrease in number of positively traced neurons we report here is consistent with previous findings. 54,83,90 –92 For example, following moderate SCI, there was a 56% decrease in the number of co-labelled FG/DBH neurons within the RVLM. Moreover, severe SCI resulted in a 97% decline in the number of co-labelled neurons. 54 As the RVLM axons project down the spinal cord within the dorsolateral funiculus, 7 this dramatic decrease in both groups likely reflects the extensive destruction of white matter both at the injury epicenter and the extension of that injury rostrally and caudally. Accordingly, both moderate and severe SCI induces a dramatic reduction in the number of sympathoexcitatory axons maintaining their connection with sympathetic pre-ganglionic neurons.
The number of FG/DBH-positive neurons was consistently lower than the number of DBH-positive neurons across all groups. At ∼65%, this proportion of co-labeled neurons most likely reflects the level at which the tracer was inserted. For example, the remaining ∼35% of the DBH-positive cell bodies likely project to sympathetic pre-ganglionic neurons above the level of tracer placement and, therefore, would not be co-labelled. This proportion of co-labelled neurons has previous been reported for similar levels of tracer placement. 54,83,90 –92
Our final finding was that the total number of DBH-positive neurons was significantly decreased following severe SCI in animals on which we did not place a tracer. It is possible that the axons extending from the RVLM to the injury site underwent retrograde degeneration (i.e., axonal dieback), which is a well-established consequence of traumatic axotomy. 93 –96 This contention is further supported by our observation of less DBH expressing neurons in the SHAM group following spinal cord re-transection for FG placement, which may be mediated by similar mechanisms. However, although there was some reduction in the number of RVLM DBH-positive neurons, we demonstrated that the vast majority (75%) of these neurons continued to express DBH, and therefore likely remained functional. This suggests that neuroprotective/neuroregenerative therapies may be useful in restoring function of these neurons. A recent study demonstrated an improvement in both resting hemodynamics and the severity of AD following neural stem cell grafting and subsequent regeneration. 24 Moreover, previous evidence implicating serotonergic fibers in the development of AD 32 suggests that the maintenance of other descending bulbospinal pathways may aid in the resolution of AD.
T3 spinal cord contusion induces dorsal horn afferent plasticity that scales with injury severity
Extensive animal and human research has demonstrated both a severity-dependent relationship and a time-course relationship between the extent of CGRP+ immunoreactivity within the dorsal horn and the severity of AD. 9,25,74,75,97 CGRP is expressed primarily by nociceptive unmyelinated C fibers as well as Aδ fibers within the dorsal root ganglia and superficial laminae. 9 However, following neuronal injury, these axons extend deep to laminae III-V where they interact with intraspinal networks and activate an inappropriately large number of sympathetic pre-ganglionic neurons, which ultimately results in the exaggerated vasoconstriction response (i.e., AD) to stimuli below the level of injury. 9,10,75,98
We found that T3 spinal cord contusion results in an increase in the expression of CGRP within the deep laminae. The increased expression of CGRP and influx of axons into the deep laminae following severe T3 contusion is likely a direct result of increased neural growth factor (NGF) influx from the dorsal root ganglia. The concentration of NGF within the cord is intimately related to the influx of these axons, 74 and manipulation of the levels of NGF has been shown to reduce or amplify the severity of AD. 25 Our observed increase in CGRP axons within the dorsal horn may act in concert with other previously established consequences of aberrant plasticity after SCI. Namely, previous electrophysiological and morphological work has highlighted that ascending propriospinal axons undergo aberrant plasticity, which may further aid the rostrocaudal spread of pre-ganglionic neuron stimulation. 10,11 However, as we did not assess these outcomes in the current study, further work would need to substantiate this hypothesis.
Although several studies have examined methods to reduce the severity of CGRP+ afferent sprouting, many have utilized a transection model, 25,74 further mechanistic data are needed to determine the utility of these axons. Our data provide similar findings to those of previous models, and provides a clinically relevant model with established mechanisms to further examine this phenomenon.
T3 spinal cord contusion induces a severity-dependent decrease in functional motor behavior
T3 spinal cord contusion resulted in a severity-dependent difference in functional hindlimb behavior, as assessed using the BBB open field locomotion assessment. BBB scores were significantly correlated with lesion volume, indicating that our data replicate the well-established relationship between the sparing of descending pathways and hindlimb function. 99 Similar to previous studies, MODERATE SCI animals regained more function by the 5 week time point, whereas SEVERE SCI animals regained very little function and were not able to weight bear. 28,72 However, longer-term studies may be needed to establish if this behavior is sustained over time.
Conclusion
Here, we provide cardiovascular, histological, and locomotor data that demonstrate the reproducibility of this new midline T3 contusion model. This T3 contusion model may, therefore, be used to investigate the mechanisms and the therapeutic potential of various neuroprotective/neuroregenerative agents to improve cardiovascular control after SCI. We believe this model is of critical importance, as cardiovascular dysfunction is the leading cause of death for individuals with SCI, and is rated among the top health priorities of these individuals. Further, we have utilized the IH contusion apparatus that is common to numerous SCI laboratories, in the hopes that this will aid this model's widespread use.
Footnotes
Acknowledgments
The laboratory of A.V. Krassioukov is supported by the Canadian Foundation for Innovation, BC Knowledge Translation Foundation, the Canadian Institute for Health Research, and the Craig H. Neilsen Foundation. J.W. Squair is supported by a Frederick Banting and Charles Best Canada Graduate Scholarship from the Canadian Institutes of Health Research and a 4 year fellowship from the University of British Columbia. P. Assinck is supported by a Frederick Banting and Charles Best Canada Graduate Scholarship from the Canadian Institutes of Health Research and a 4 year fellowship from the University of British Columbia. W. Tetzlaff holds the John and Penny Ryan British Columbia Leadership Chair in Spinal Cord Research. C.R. West is supported by a Scholar award from the Michael Smith Foundation for Health Research and the Rick Hansen Institute.
Author Disclosure Statment
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.