
Abstract
Task-specific rehabilitation has been shown to promote functional recovery after acute spinal cord injury (SCI). Recently, the omega-3 polyunsaturated fatty acid, docosahexaenoic acid (DHA), has been shown to promote neuroplasticity after SCI. Here, we investigated whether the combination of a single bolus of DHA with rehabilitation can enhance the effect of DHA or rehabilitation therapy in adult injured spinal cord. We found enhanced functional improvement with DHA in combination with rehabilitation compared with either treatment alone in a rat cervical lateral hemisection SCI model. This behavioral improvement correlated with a significant sprouting of uninjured corticospinal and serotonergic fibers. We also observed that the greatest increase in the synaptic vesicle protein, synaptophysin, and the synaptic active zone protein, Bassoon, occurred in animals that received both DHA and rehabilitation. In summary, the functional, anatomical, and synaptic plasticity induced by task-specific rehabilitation can be further enhanced by DHA treatment. This study shows the potential beneficial effects of DHA combined with rehabilitation for the treatment of patients with SCI.
Introduction
A
Evaluation of the effects of task-specific training after cervical SCI has focused primarily on the recovery of fine forelimb function. Previous studies have demonstrated that specific task forelimb training, a voluntary rehabilitative strategy, enhances functional recovery through spinal tract modification in cervical SCI animal models. 8,9 Several studies have indicated that a greater effect may be achieved when a combination of interventions is implemented. For example, in a thoracic SCI animal model, treadmill training combined with olfactory ensheathing glial cell transplantation 10 or radiation therapy 11 demonstrated significant improvement compared with individual therapy. A number of studies also showed that voluntary forepaw motor rehabilitation shows a strong synergistic effect when given together with chondroitin sulfate proteoglycan digestion 8 or neurotrophic factor treatment 12 in a cervical SCI model.
Docosahexaenoic acid (DHA) is an omega-3 polyunsaturated fatty acid that is essential in brain development and has structural and signaling roles. DHA supplementation can also improve learning by enhancing synaptic plasticity in the brain. 13,14 We have recently documented that acute DHA administration improves neurological functional recovery following injury in rodent pyramidotomy and cervical SCI animal models through promoting neuroplasticity. 15 Recent data revealed that a combination of behavioral training and dietary DHA promoted a significant enhancement in neurological and functional recovery after traumatic brain injury compared with either intervention alone. 16 The present study was aimed at assessing the effectiveness of DHA paired with task-specific rehabilitation in cervical SCI.
Methods
Animals
All animal research was carried out under the United Kingdom Animals (Scientific Procedures) Act of 1986. Surgery was carried out under anesthesia and pain relief was provided appropriately during post-operative care. The experimental time line in Figure 1 offers an overview of the study design.
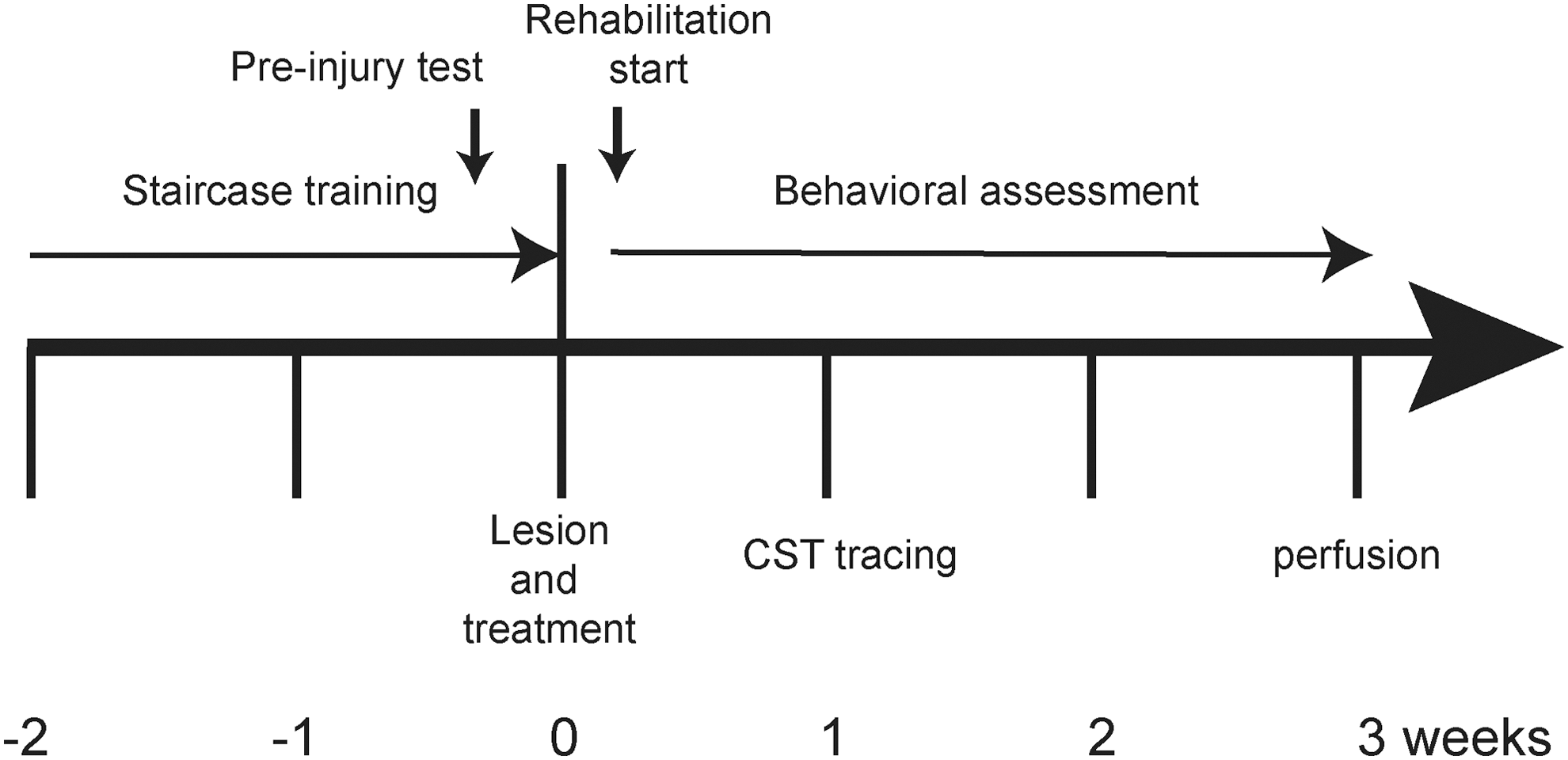
Time line of the experiment. Two weeks prior to cervical hemisection, animals were trained on the Montoya staircase every day. Task-specific rehabilitation training began 2 days after cervical hemisection. An anterograde tracer to label the intact corticospinal tract (CST) was injected one week following cervical lesion. All rats underwent the Montoya staircase test (every day in week 1 after injury and then every other day from weeks 2 to 3 after injury) and the grid exploration test (every week) and were perfused under anesthesia for histological analysis 3 weeks after injury.
Cervical hemisection in the rat
Cervical lateral hemisection of the spinal cord was performed in adult female Sprague-Dawley rats (250–300 g) using methods previously described. 15,17 Briefly, animals were anesthetized with isoflurane and a dorsal midline incision was made at cervical level, to expose the cervical vertebrae. A left hemilaminectomy was carried out at C4 and C5, and then a lateral hemisection was made at the C4–5 level using a microblade. Sham animals received only a hemilaminectomy, to expose the spinal cord without injury. After surgery, the muscles and skin layers were sutured and animals were returned to a warm incubator for recovery. Thirty minutes after hemisection, animals received a tail vein injection of either vehicle (0.2% ethanol in saline) or DHA (Sigma D2534, 250 nmol/kg) in a volume of 2.5 mL/kg. The DHA dose chosen was based on previous studies from our laboratory with this fatty acid, showing functional improvements after SCI. 15,17,18 Post-operative care involved subcutaneous injection of analgesic (buprenorphine, 0.01 mg/kg) and saline twice daily for 3 days following surgery.
Anterograde tracing
To evaluate whether the treatment had promoted axonal sprouting, we performed anterograde tracing of the intact corticospinal tracts (CST) as previously described. 19,20 Tracing occurred one week after the hemisection in rats. Animals were placed in a stereotaxic frame and burr holes were made into the skull ipsilaterally to the hemisection. In the hemisected rats, burr holes were made at the following coordinates, defined as anteroposterior (AP) and mediolateral (ML): (i) AP: −1.5 mm, ML: 2.5 mm; (ii) AP: −0.5 mm, ML: 3.5 mm; (iii) AP: −0.5 mm, ML: 2.5 mm; (iv) AP: +0.5 mm, ML: 3.5 mm; (v) AP: +1.0 mm, ML: 1.5 mm; and (vi) AP: 1.5 mm, ML: 2.5 mm; and (vii) AP: 2.0 mm, ML: 3.5 mm, relative to bregma. At each site, injections of biotinylated dextran amine (BDA; 10%; 10,000 MW, 1 μL/site) were delivered using a glass micropipette attached to a Hamilton syringe via water-filled polyethylene tubing. The micropipette was inserted 2 mm deep below the skull surface and BDA was delivered at a rate of 0.2 μL/min using a microinfusion pump. Animals were subsequently maintained for 2 weeks before tissue was collected for histology.
Immunocytochemistry
At the end of the study, animals were deeply anesthetized with sodium pentobarbital (120 mg/kg, i.p.) and flushed with saline transcardially followed by 4% paraformaldehyde. Once spinal cord tissues had been cryoprotected in 20% sucrose, spinal cord segments containing the hemisected region were cut horizontally at 20 μm using a cryostat and organized as slide series. Slides of spinal cord sections were randomly chosen and washed with gentle agitation in phosphate-buffered saline (PBS) (3 × 5 min). Then, the sections were incubated in 10% normal donkey or goat serum for 30 min followed by an overnight incubation with primary antibodies (rabbit anti-serotonin, 1:3000, ImmunoStar; rabbit anti-synaptophysin, 1:1000, Cell Signaling Technology; mouse anti-Bassoon, 1:3000, Enzo Life Sciences). The next day, sections were washed three times (5 min each) in PBS before being incubated for 2 h in the appropriate secondary antibodies conjugated to Alexa Fluor 488 or 594 (1:1000). After another three 5-min washes in PBS, sections were then counterstained with the fluorescent nuclear dye Hoechst 33342 (2 μg/mL; Sigma, UK) for 5 min to facilitate detection of cell nuclei. Slides were mounted in ProLong® Gold antifade reagent.
The tyramide signal amplification technique was carried out for the detection of BDA-labeled CST fibers. Sections were washed three times (5 min each) in PBS and then incubated with 0.3% hydrogen peroxide for 30 min. After further 3 × 5-min washes with PBS, the sections were incubated in avidin-biotin-peroxidase complex (Vectastain ABC Elite Kit, Vector Laboratories) for 30 min at room temperature. Following 3 × 5-min washes in PBS, sections were incubated with biotinyl tyramide (1:75, NEN Life Sciences) for 10 min. After a further 3 × 5-min washes in PBS, the sections were incubated with ExtrAvidin® FITC (1:400) for 2 h before the sections were further immunostained with another antibody for double-labeling or washed and coverslipped in mounting medium.
Behavioral testing
All behavioral testing and data analysis were carried out blind by the experimenter. In accordance with our animal use regulations, the minimal number of animals were used for repeated behavioral measures with the Montoya test (n = 8–10) and grid exploratory test (n = 6). A randomly selected group of animals was chosen for the grid exploratory test.
Skilled forelimb test
The Montoya staircase test is a well-established measure of forelimb motor function in rats and has been demonstrated to provide a reproducible and sensitive assessment of motor impairment. 21 In our study, animals were trained on the Montoya staircase test daily for 14 days prior to SCI. We then performed the test every day for the first 10 days after injury and then every other day until the end of the experiment. Two parameters were assessed in this task. One was the number of food pellets retrieved and eaten by animals (skilled forelimb control) and the other one was the number of food pellets displaced or retrieved (gross forelimb control). One food pellet was placed per step, and the number displaced and the number eaten were recorded.
Skilled locomotion test
Deficits in voluntary descending motor control can be examined by assessing the ability of mice and rats to precisely control and place their fore- and hindlimbs onto grid wires as they roam around. 20,22 Animals were placed on a 20-cm raised grid platform (40 cm × 60 cm containing 5 cm × 5 cm square grids) every week post-operatively, to assess the misplaced steps made by the injured left forelimb. A misplacement was recorded when the limb protruded entirely through the grid and extended below the wire surface. All animals were tested for their baseline performance by subjecting them to the grid exploration test three times prior to surgery.
Task-specific rehabilitation
Staircase training has been shown to improve food pellet grasping after cervical SCI 8 and brain ischemic lesions. 23 Recovery of reaching and pellet retrieval is associated with plasticity of the corticospinal and rubrospinal tracts and motor cortex, and requires integration of segmental, intersegmental, and supraspinal input to propriospinal interneurons and motor neurons, over many spinal levels. 24,25
During the 2-week period before injury, all animals were trained to grasp and eat food pellets from the Montoya-type staircase device. Two days after cervical spinal cord lateral hemisection, the rehabilitation group of animals was trained on a task-specific rehabilitation procedure. One food pellet was put on the right side wells (non-lesion side), and a surplus of food pellets (8–10 pellets per well) was placed on the left side so the animals were encouraged to grasp the food pellets with their affected forelimbs. The training was performed for 30 min twice daily with the first daily training session starting in the morning and the second session in the late afternoon. The staircase testing was performed one hour before the afternoon rehabilitation training session. In one group, rats received DHA 250 nmol/kg intravenously 30 min after cervical hemisection, and in another group rats received a vehicle injection, as described in our previous study design. 15 After 3 weeks of behavioral testing, the animals were perfused and the spinal cord tissue was harvested for further histological examination.
Image analysis
Image analysis and quantification were carried out with the observer blinded to the group assignment. At least four sections per staining of the corresponding horizontal plane containing the region of interest (e.g., motor neurons) were viewed and captured on a Leica epifluorescence microscope at 20 × magnification. The depth and distance from the lesion site of the images taken and analyzed were consistent for all animals. Quantitative analysis of the number of serotonin-labeled axons, BDA-labeled corticospinal axons, Bassoon-labeled boutons, and synaptophysin-labeled boutons was conducted by randomly capturing images of the spinal cord 5 mm caudal to the lesion site. The immunoreactivity in captured images was then quantified in a 500 μm × 500 μm measuring frame by using the ImageJ analysis program (ImageJ 1.46u, National Institutes of Health). Images were converted to a set threshold and then counted automatically using customized macros. The level of immunoreactivity was expressed as the percentage of the area of the measuring frame that contained immunoreactivity.
Statistical analysis
The behavioral assessment and histological analysis were performed blind. All statistical analyses were performed using GraphPad Prism Version 6 (GraphPad Software Inc., San Diego, CA). The data were presented as means and standard error of the means. One-way, or two-way repeated measures analysis of variance (ANOVA) were used to compare experimental groups, with Tukey's post hoc comparisons for equal sample sizes or Bonferroni's post hoc test for unequal sample sizes. Differences were considered significant when p < 0.05.
Results
Combined therapy induced greater skilled forelimb functional recovery than single treatment
To test whether rehabilitation can support functional improvement after cervical lateral hemisection, animals with and without rehabilitation were compared using the Montoya staircase test. Our results showed that 3 weeks of rehabilitative training increased the functional neurological recovery of both vehicle- and DHA-treated rats (Fig. 2 and 3). In the vehicle-treated rehabilitation group, trained rats significantly displaced (Fig. 2A) and ate (Fig. 3A) more food pellets after 9 to 10 days post-injury. A similar significant increase was observed in the number of food pellets eaten by rehabilitation-trained rats receiving DHA treatment, after 12 days post-injury (Fig. 3B). Interestingly, the rehabilitation training did not significantly increase the ability of rats receiving DHA treatment to displace more food pellets (Fig. 2B). The data suggest that the rehabilitation training can increase the accuracy and success of food retrieval from 9 to 12 days following cervical lateral hemisection in rats. At the 3-week time-point after cervical hemisection, animals that received the combined therapy of DHA and rehabilitation were able to significantly retrieve more food pellets than the other three groups, that is, the DHA treatment only, rehabilitation training only, and the control vehicle-treated only group (Fig. 3C). The order of functional improvement (highest number of food pellets eaten first) was: DHA and rehabilitation (2.7 ± 0.6) > DHA only (1.4 ± 0.7) = vehicle and rehabilitation (1.2 ± 0.3) > vehicle only (0 ± 0.1). This shows that rehabilitation or DHA treatment alone enabled animals to eat more food pellets compared with the vehicle group, but the combination of DHA and rehabilitation enabled animals to grasp and eat more food pellets than each treatment alone.
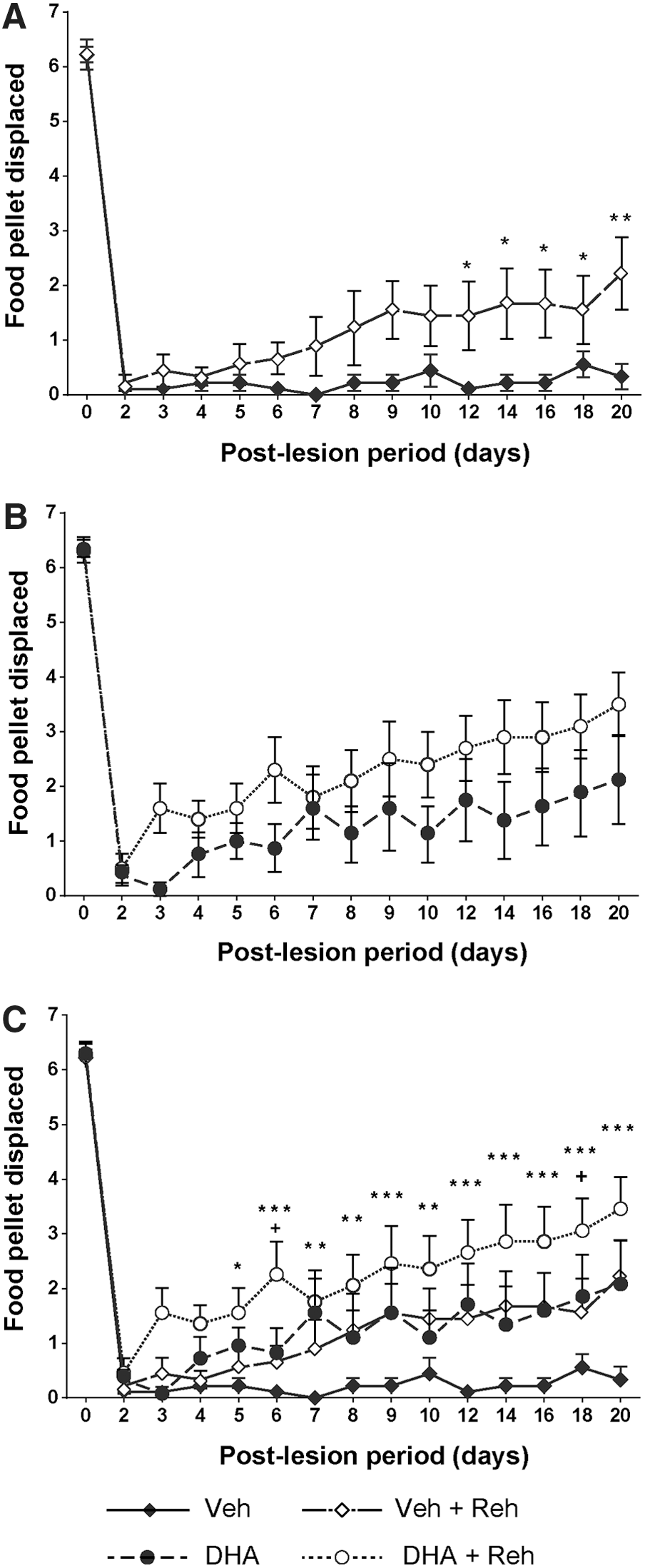
Effect of rehabilitation training on gross forelimb function. After spinal cord injury (SCI), all rats lost gross forelimb function in the Montoya staircase test. Starting from 10 days post-injury, rats that had received rehabilitation training began to displace significantly more pellets in the vehicle groups (
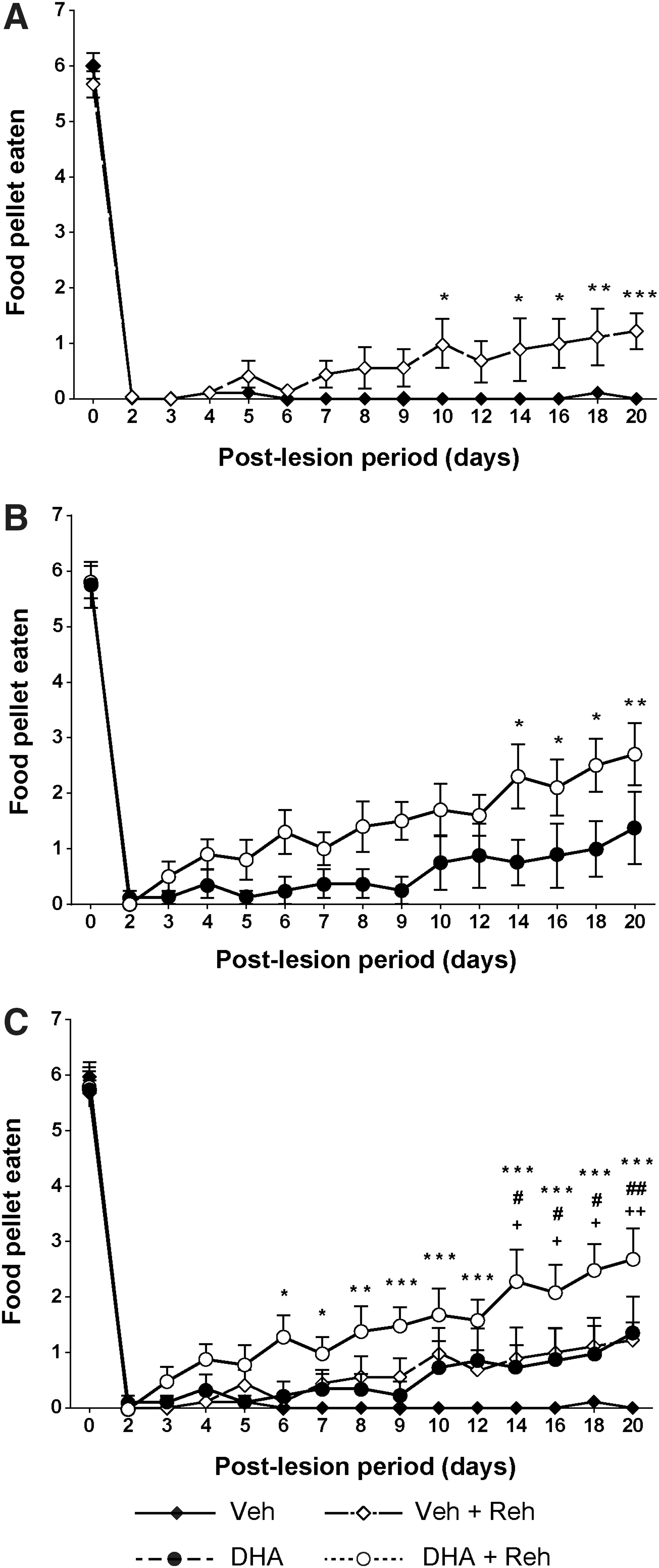
Effect of combined therapy on skilled forelimb function. For the first 2 days after surgery, no rats showed any ability to retrieve food pellets either vehicle-treated with/without rehabilitation (
Rehabilitation training has no effect on skilled locomotion recovery
The grid exploration test can also assess fine motor control of the forelimb, including the wrist and paw. The use of the forelimbs to walk over rungs is fundamentally a different motor skill than the reaching for pellets assessed in the Montoya staircase test. After injury, throughout 3 weeks, there was no significant difference in the misplacement of the left forelimb between the rehabilitation and non-rehabilitation group (Fig. 4A, 4B). However, in the vehicle-treated group without rehabilitation, there was significantly more misplacement than either after DHA alone or DHA with rehabilitation (Fig. 4C). The data suggest that rehabilitation cannot provide functional improvement in the skilled locomotion test, and this is also not enhanced further with DHA treatment compared with DHA treatment alone. However, the rehabilitation did not deteriorate the performance of non-trained skilled locomotor movement.

Effect of combinatorial treatment on non-trained skilled locomotion. In the vehicle-treated group, overall, the rats receiving rehabilitative training exhibited fewer forelimb misplacements during the test period; p value = 0.09 in the first week, p value = 0.37 in the second week, and p value = 0.16 in the third week (
Combined therapy promotes axonal sprouting
Corticospinal axons and collaterals
The anatomical changes in the contralateral spared CST were studied following ipsilateral BDA injection into the forelimb region of the sensorimotor cortex. Contralateral CST sprouting across the midline above and below the lesion site was examined. In the vehicle-treated group, the rehabilitation training promoted significant CST fiber sprouting above and below the lesion site (Fig. 5A). In the DHA-treated group, there was also a similar significant increase in CST fiber sprouting in the rehabilitation group compared with the DHA treatment alone group (Fig. 5B). However, regardless of DHA treatment or not, rehabilitation training showed increased axon sprouting across the midline compared with the vehicle group (Fig. 5C). These data suggest that combined therapy has a more marked effect on CST sprouting than DHA treatment alone.

The effect of rehabilitation training on corticospinal tract (CST) axon sprouting. Representative images of CST labeling in the dorsal horns 5 mm caudal to the injury site, in rats receiving docosahexaenoic acid (DHA) or vehicle treatment, with or without rehabilitation training. Arrows indicate the sprouting CST fibers from the contralateral side. The CST sprouting fibers in the spinal cord rostral and caudal to the lesion site were quantified. The data revealed large numbers of sprouting axonal fibers in rats receiving rehabilitative training in the vehicle group (*p < 0.05) (
Combined therapy and serotonergic fiber sprouting
The sprouting of other CNS axons was also examined, to reveal whether or not sprouting was restricted to corticospinal fibers. Utilizing 5-HT immunohistochemical staining, serotonergic fibers were identified. To investigate the sprouting axon fibers below the lesion, the number of serotonin fibers was quantified 5 mm caudal to the lesion site. The quantification showed that the animals that received combined therapy displayed very marked sprouting and had significantly more serotonergic fibers caudal to the lesion in comparison with the three other groups (Fig. 6).
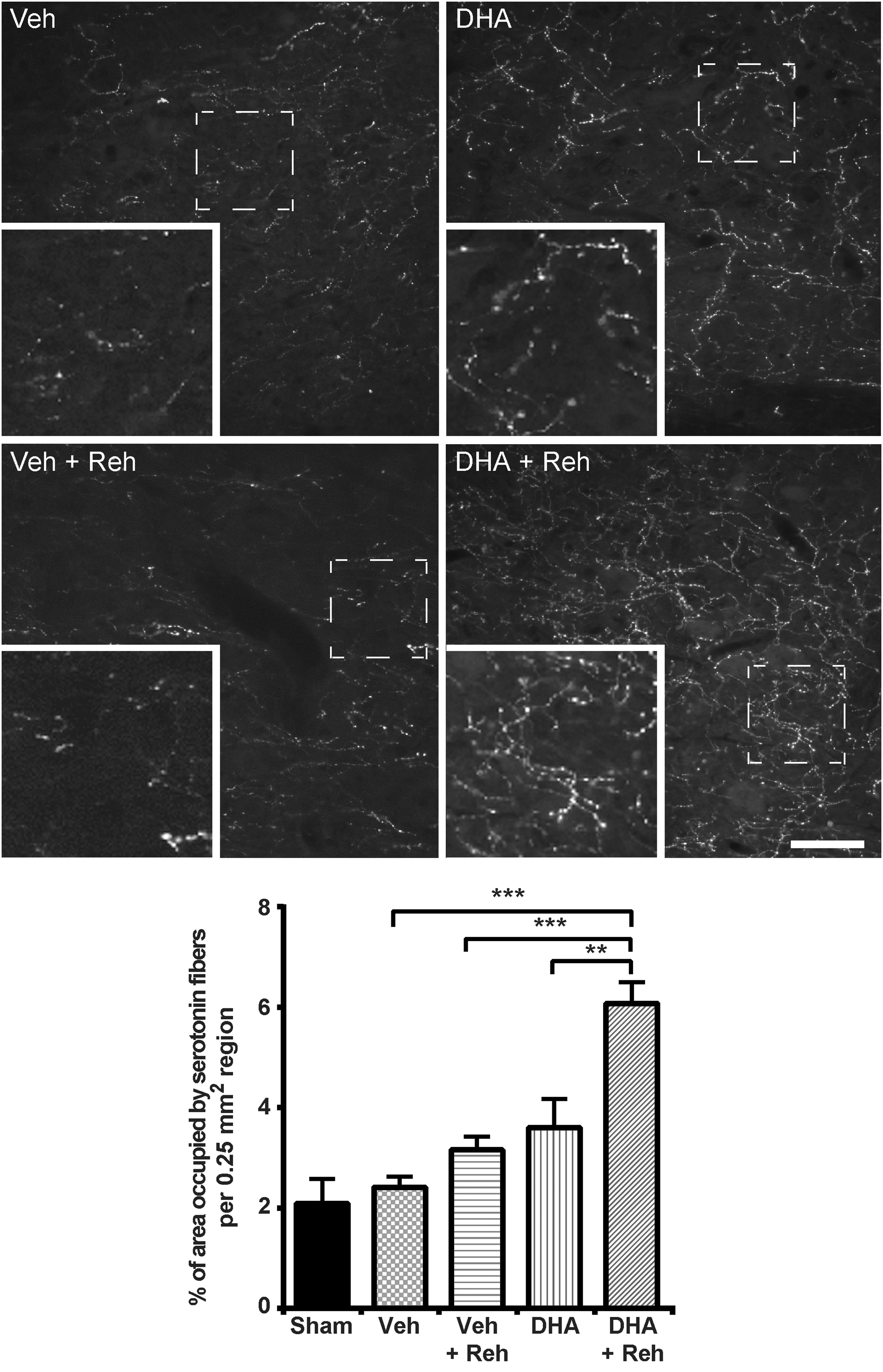
Effect of combined therapy on serotonin fibers. Representative images of 5-HT labeling in the dorsal horns 5 mm caudal to the injury site in rats receiving docosahexaenoic acid (DHA) or vehicle treatment with or without rehabilitation training. The quantification showed that combined therapy significantly increased the density of serotonin fibers compared with other groups (***p < 0.001 represents Vehicle versus DHA + Rehabilitation; ***p < 0.001 represents Rehabilitation versus DHA + Rehabilitation therapy; **p = 0.003 represents DHA versus DHA + Rehabilitation therapy). Results represent mean ± standard error of the mean (SEM). Data obtained from n = 5 animals in each group. Scale bar = 100 μm.
The effect of combined therapy on synaptic plasticity
Synaptophysin immunohistochemistry
Synaptic terminals around motor neurons can be identified by synaptophysin immunostaining. Recently, we have demonstrated a loss of synaptophysin immunostaining after cervical SCI. 15 Given that we showed in a previous study that DHA alone administered after injury promoted synaptic plasticity after cervical SCI, we asked whether combined therapy could maximize the expression of synaptophysin post-injury. Using an immunohistochemical staining method, the synaptophysin expression around ventral horn neurons located in the spinal cord 5 mm caudal to the lesion site was examined in both vehicle- and DHA-treatment groups. The rehabilitation training did not significantly increase the level of synaptophysin (Fig. 7). The group receiving combined DHA treatment with rehabilitation expressed the highest level of synaptophysin of all the injured groups (Fig. 7). The data suggest that there was no significant additional synergistic effect of rehabilitation compared with DHA treatment alone, as far as pre-synaptic plasticity was concerned.

Effect of combination treatment on synaptophysin. Representative images in the cervical ventral horn taken 5 mm caudal to the epicenter, in docosahexaenoic acid (DHA)- or vehicle-treated rats, with or without rehabilitative training. Sections were immunostained for synaptophysin at 3 weeks after spinal cord injury (SCI). Quantitative analysis revealed a significant increase in the DHA-treatment group receiving training versus the vehicle group with/without training (*p < 0.05). Results represent mean ± standard error of the mean (SEM). Data obtained from n = 5 animals in each group. Scale bar = 100 μm.
Bassoon immunohistochemistry
Synapse formation requires the organization of pre-synaptic active zones, the synaptic vesicle release sites, in precise apposition to post-synaptic neurotransmitter receptor clusters. Bassoon has been previously demonstrated using immuno-electron microscopy to be localized in the pre-synaptic active zone of synapses in the central nervous system. 26 We demonstrated that the immunoreactivity of Bassoon was significantly upregulated following rehabilitation training in the injured rats compared with vehicle or DHA treatment alone (Fig. 8). The highest level of Bassoon was seen in the group with combined treatment. The data suggest that rehabilitation promotes pre-synaptic formation via increased pre-synaptic active zones, and this can be enhanced with DHA treatment.
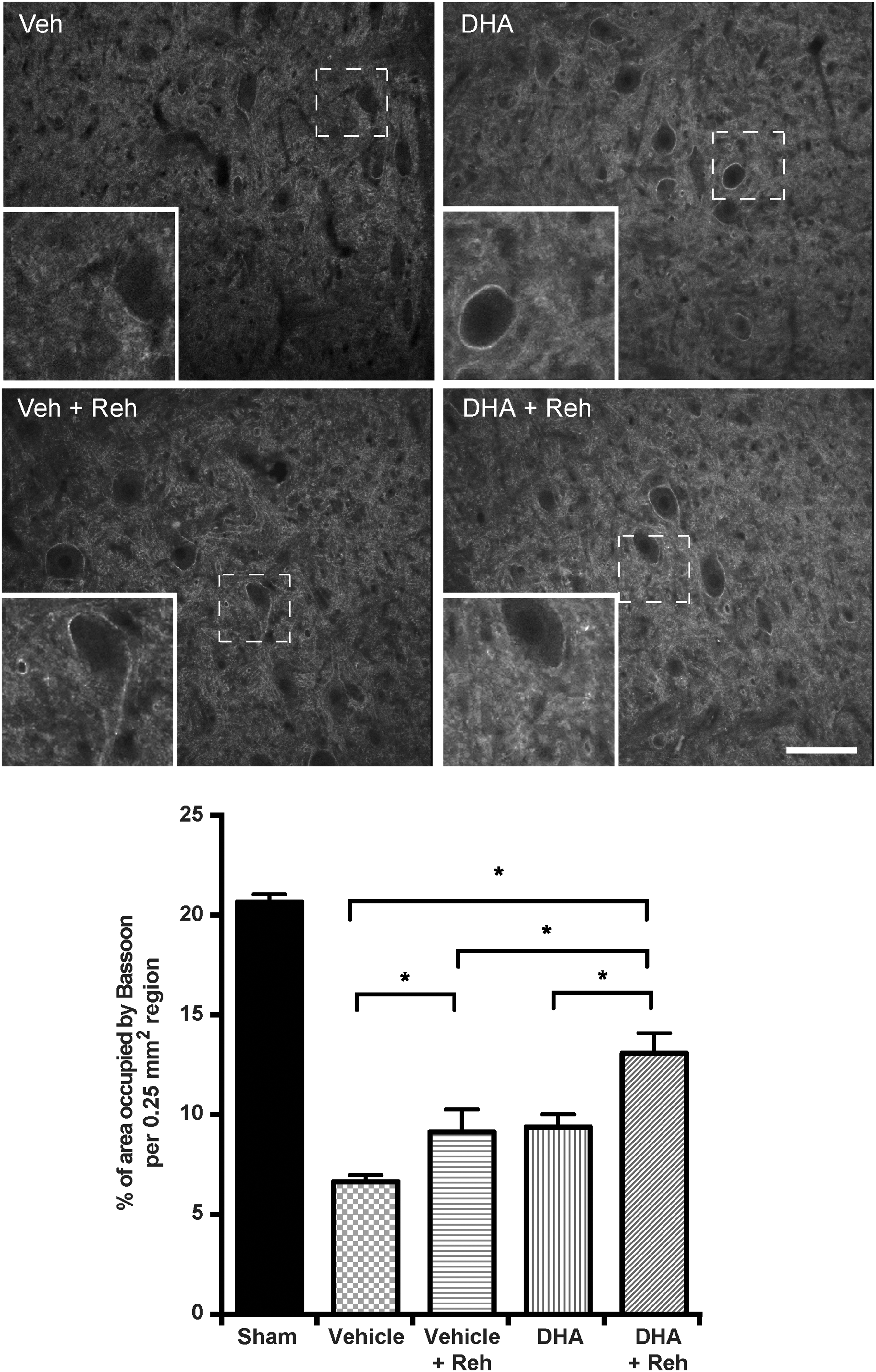
Effect of combination treatment on Bassoon. Representative images in the cervical ventral horn taken 5 mm caudal to the epicenter, in docosahexaenoic acid (DHA)- or vehicle-treated rats, with or without rehabilitative training. Sections were immunostained for Bassoon at 3 weeks after spinal cord injury (SCI). Quantitative analysis revealed a significant increase in the DHA-treatment group receiving training versus the vehicle group with/without training (*p < 0.05). Results represent mean ± standard error of the mean (SEM). Data obtained from n = 5 animals in each group. Scale bar = 100 μm.
Discussion
Several publications have demonstrated that DHA promotes beneficial functional effects in SCI. 17,18,27,28 However, the combined effect of rehabilitation training and DHA treatment following SCI has not been studied. The results presented in our study show that the neuroplasticity following cervical hemisection, in terms of both functional neurological manifestations and histological outcome, is significantly improved after combined therapy, compared with DHA treatment or rehabilitation alone. In this study, specific forelimb behavioral assessments (staircase test) showed better functional recovery in rats receiving intensive rehabilitation training combined with DHA treatment. However, in the skilled locomotion test (grid exploration test), rehabilitation did not significantly enhance the effect of DHA. Our data demonstrate that acute administration of DHA post-injury combined with early rehabilitation can greatly increase axonal sprouting and synaptic plasticity in the spinal cord proximal to the lesion site after cervical lateral hemisection of the spinal cord.
Anatomical neuroplasticity following combined therapy
Following cervical SCI, deficits in voluntary and involuntary motor control have been attributed to direct damage to the corticospinal and bulbospinal tracts and the motor neurons within the gray matter of the ventral horn. Recently, it has been recognized that spontaneous and treatment-induced functional recovery correlates with sprouting of lesioned and spared descending axons after SCI. 29 –31 Rehabilitative training in a reaching task has been shown to increase spontaneous collateral sprouting of CST axons after a central nervous system (CNS) lesion. 9,32 In our experiment, we found that contralateral sprouting of CST axons was enhanced in animals that underwent rehabilitative training with or without acute DHA treatment. However, in our study the maximum effect on CST axonal sprouting was seen in the combined therapy group, that is, rehabilitative training and acute DHA treatment.
Our laboratory and others have previously reported that rapidly diminished spinal levels of serotonin occur ipsilateral to a spinal lesion following thoracic SCI and cervical SCI, with levels of serotonin returning by 4 weeks after injury. 15,33 –35 These dynamic changes can be used as an indicator of injury severity 36 and provide a substrate for changes in somatosensory and locomotor behaviors. Interestingly, the analysis of serotonin fibers 5 mm below the lesion site revealed that rehabilitation can lead to an increase in serotonin fiber density, which is consistent with previous published studies. 37 However, the most robust and significant sprouting of serotonergic axons was detected in the group with combined DHA and rehabilitation treatment.
Our recent study found that acute DHA treatment after cervical SCI could induce robust sprouting of uninjured corticospinal and serotonergic fibers into the denervated side of the cord and form contacts with interneurons. 15 One possible explanation for the benefit of combined therapy is that once new neuronal circuits have been formed, the rehabilitation training may favor appropriate projections by modulating and stabilizing the neuronal pathways, thus boosting functional improvement.
Synaptic functional upregulation following combined therapy
Krajacic and colleagues have previously demonstrated that rats trained to grasp food pellets following cervical SCI have a significantly higher success rate than untrained rats even after pyramidotomy. 38 These data indicate that CST sprouting is not the only mechanism to promote functional recovery following rehabilitation training. Another possible contributor to this improvement in a specific task after training is the upregulation of synaptic function. This enhancement can be promoted by the increased activity of existing receptors and/or by increasing the number of receptors at functional synapses. A role for DHA in synaptic plasticity is well established in both in vivo and in vitro studies. DHA has also been shown to promote the synthesis of synaptic membranes and specific presynaptic proteins (synapsin-1 and syntaxin-3) and post-synaptic proteins (post-synaptic density protein 95). 39,40 Upregulation of these synaptic proteins after dietary consumption of DHA has also been linked to improvement in brain learning 14 and spinal cord sensorimotor learning following SCI. 41 To evaluate a possible functional role for synaptic upregulation, we measured the level of a pre-synaptic protein (synaptophysin) in the cervical spinal cord. The levels of synaptophysin were significantly elevated in the spinal cord of rats that had the combined DHA and rehabilitation treatment. In contrast, the modest effect of rehabilitation was not a reflection of the significant difference seen in functional outcome between the vehicle and rehabilitation only groups. This suggests that other factors than merely the levels of synaptophysin are important in rehabilitation-triggered processes, such as the generation of new circuitry or the modulation of various types of synapses (excitatory or inhibitory).
Synapse formation requires the organization of pre-synaptic active zones, the synaptic vesicle release sites, in precise apposition to post-synaptic neurotransmitter receptor clusters. The recent use of Bassoon, a pre-synaptic cytosolic scaffolding protein essential for the active zone assembly in synapses, has enabled its use for the detection of active zones in synapses of the CNS and the peripheral nervous system (PNS). 42 Previous studies revealed that training increased Bassoon immunoreactivity at the neuromuscular junction. 43 In our study, we analyzed the levels of Bassoon immunoreactivity in different treatment groups within the CNS. Our data are in line with the previous study carried out in the PNS, and further consolidates the hypothesis that rehabilitation training can increase Bassoon immunoreactivity, which in turn contributes to functional recovery—and this effect is enhanced by the fatty acid.
Competing effects after rehabilitation training
In our behavioral test, the ability of food pellet retrieval was significantly improved after DHA and rehabilitation therapy compared with the DHA alone group. However, there was no significant difference in food pellet displacement between these two groups. This lack of improvement in the displacement task could be interpreted as lack of skill transfer in rehabilitation training.
Some studies reported that the training-induced improvement in one particular task may be at the cost of an untrained task. 8,9 This supports the hypothesis that there is a limited resource for new circuit formation following injury, and different behaviors may compete for that resource. 44 To address this question, we tested for skilled locomotion, to examine if task-specific training interfered, positively or negatively, with another untrained skill. In the DHA- and vehicle-treatment group, there was no significant difference. Our data are in agreement with other published studies, in which after cervical lesion, rats receiving forelimb skilled grasping training did not develop deficits in running along a ladder or beam. 37,38
Mechanism underlying the effects of the combined therapy
Several studies have proposed that brain-derived neurotrophic factor (BDNF) plays a key role in the synergistic effect of DHA and exercise training. Previous results have shown that training can upregulate expression of BDNF in the nervous system and that this is pivotal for recovery after SCI. 12,45 The level of BDNF is also elevated after DHA treatment and a combinatory effect with exercise has been demonstrated in different animal models. 14,16
BDNF is associated with structural changes including increased sprouting of serotonergic axons in the lumbar spinal cord of mice after moderate contusion injury. 46 Another study showed that the reorganization of CST circuits is guided by BDNF secretion from target interneurons. Further, knockdown of BDNF in spinal neurons diminished the formation of new CST pathways. 47 Therefore, some of the functional effects seen in this study might be mediated by an increase in overall activity in motor systems modulated by increased availability of BDNF.
One strategy to promote axonal growth is to overcome the inhibitory environment. Glial scar formation is a reactive cellular process involving astrogliosis that inhibits axonal growth following SCI. Our laboratory has recently documented that DHA-enriched dietary supplementation modified the biochemical composition of the scar border. 48 The level of chondroitin sulfate proteoglycans and gliosis activity is diminished after treatment with DHA or DHA-enriched preparations. 48,49 Our recent study showed that acutely administered DHA can promote neuroplasticity by modulating miR-21 expression following cervical SCI, and downregulating phosphatase and tensin homolog (PTEN). 15 miR-21 has been demonstrated as a potential regulator of astrogliosis after SCI by using transgenic mice that overexpress in astrocytes either miR-21 or a miRNA sponge designed to inhibit MiR-21 function. 50 Our results indicated that DHA treatment can provide favorable conditions that can promote axonal regrowth following rehabilitation, which in turn contributes to the increased effects of combined therapy.
One limitation of our study is a short-term rehabilitation regime in the early acute phase following SCI. Continuous improvement was observed toward the end of experiment, so the full potential of combined therapy was not comprehensively demonstrated, because of the termination of the experiment. Another concern that should be considered is the timing of initiation of rehabilitation, because the timing of rehabilitation initiation remains controversial. Several studies have documented that rehabilitation is more efficacious when initiated within a few days after injury. 51,52 In our study, the rehabilitation program began 2 days after SCI, so to support the expression of plasticity promoting genes and growth factors, which are upregulated immediately, but decline over time following SCI. 53 –55 In future studies, we could explore the temporal aspect of intervention with rehabilitation training, to maximize this combined treatment strategy.
Conclusion
Our study demonstrated that the ability of rehabilitation to promote neurological functional recovery after cervical SCI can be greatly enhanced by acute neuroprotective DHA treatment. Most importantly, we demonstrated that rehabilitation can interact with DHA treatment in terms of axonal sprouting and synaptic upregulation. This study further supports the potential of combining drug therapy with rehabilitation for the treatment of patients with SCI and illustrates the importance of combining an intervention implemented very early after injury with an appropriate rehabilitation regime offered to the patient in the recovery phase.
Footnotes
Acknowledgments
We acknowledge the generous support of National Science Council, Chang Gung Memorial Hospital, Taiwan (CMRPG3F1001-2) (ZHL), CMDRP and Barts, and the London Charity (PKY and AMT).
ZHL and PKY carried out the surgery. ZHL carried out the behavioral testing, histology, and analysis. ZHL, PKY, JVP, and AMT designed the experiments and wrote the manuscript.
Author Disclosure Statement
No competing financial interests exist.