
Abstract
Traumatic brain injury (TBI) continues to be a serious health care issue while therapies to treat TBI remain elusive. Promising results from the use of endothelial progenitor cells (EPCs) in numerous disease states highlight the pleiotropic capacity of this cell type. We have previously demonstrated that EPC-conditioned media reduces axonal degeneration subsequent to in vitro oxygen-glucose deprivation insult and concurrently improves white matter and microvascular outcome in vivo after mid-line fluid percussion injury. In the current study, we evaluated the effectiveness of allogeneic endothelial cells derived from rat bone marrow on microvascular recovery and neuronal sparing after lateral fluid percussion injury. We observed reduced expression of activated caspase-3 and poly (ADP-ribose) polymerase cleavage at 72 h post-injury. Subacute injury assessments at 30 days post-injury using immunohistochemistry indicated nonsignificant effects on microvascular outcome and neuronal cell density in the hippocampus. Behavioral testing using the Morris Water Maze and rotarod demonstrated significant improvement in locomotor function, as measured by the rotarod task, but no significant differences in spatial memory ability. The data suggest that EPCs contribute to improvements in the early phase of secondary injury through inhibition of apoptosis whereas the effects on longer-term recovery were less clearly defined. There is potential in the use of EPCs to treat secondary injury post-TBI; however, optimization of their effects through increased duration or homing capacities remains to be examined.
Introduction
T
EPCs are recruited from the bone marrow into circulation or reside within the vascular lining 16,17 and play an important role in the maintenance of the vascular system in normal and diseased states. 18 –20 Although the precise definition of EPCs remains controversial because of overlap in expression of surface receptors with other hematopoetic cell types, 17,21 it is evident that EPCs in circulation account for a very small percentage of the total number of cells in blood. 21,22 However, in vitro, EPCs display a marked ability for clonal proliferative potential. 23 Thus, the procurement of an adequate number of EPCs for therapeutic treatment using EPCs expanded in vitro presents a potential therapeutic approach.
We have previously demonstrated diffuse reductions in microvascular density and expression of hypoxia-inducible factor 1 alpha in the brain subsequent to modeled TBI, 24 suggesting impaired cerebral perfusion post-injury. Recently, we demonstrated improvements in cerebral white matter and microvasculature post-EPC treatment in the acute phase after mid-line fluid percussion trauma, 12 suggesting a role for EPCs in mediating vascular and axonal protection and promoting recovery post-trauma. In the current study, we evaluated the effects of allogeneic cultured EPCs on acute and subacute outcome measures after lateral fluid percussion injury (FPI) in rats. We examined the effects of EPC administration on expression of activated caspase-3 and expression of the caspase-mediated poly (ADP-ribose) polymerase (PARP) cleavage product at 48 h post-EPC administration. We also quantified the density of capillary networks and neuronal density in the hippocampus at 30 days post-injury. Behavioral measurements using the rotarod task and Morris Water Maze (MWM) demonstrated some improvement in outcome at 30 days post-injury and highlighted the importance of evaluating outcome at subacute time points to validate the continuity of therapeutic benefit in long-term recovery.
Methods
All animal procedures were performed in accord with guidelines established by the Animal Care Committee at St. Michael's Hospital in accord with the standards set by the Canadian Council on Animal Care. The study was designed in accord with ARRIVE (Animal Research: Reporting and In Vivo Experiments) guidelines.
Lateral fluid percussion injury and endothelial progenitor cell injections
Lateral FPI was performed on adult male Sprague-Dawley rats (275–300 g) as previously described. 24 Rats were housed in pairs in a dedicated animal facility under a 12-h light/dark cycle. Water and rat chow were provided ad libitum. In brief, rats were anesthetized with 2% isoflurane delivered in laboratory-grade air. A thermal heating blanket was used to maintain body temperature at 37°C. A 2-mm-diameter extradural craniotomy was performed approximately 1.5 mm to the right of the mid-line suture, midway between bregma and lambda lines. A polypropylene luer fitting with an inner diameter 1.5 mm was fixed to the opening with cyanoacrylate adhesive and dental acrylic, filled with 0.9% isotonic saline and allowed to cure for 15 min. The fitting was then attached to the FPI device. Rats were subjected to a moderate extradural FPI at a fluid pressure pulse of 2.5 atmospheres with a 15-ms duration. Bone wax was used to seal the hole in the skull, and the scalp was sutured before recovery in a temperature-controlled chamber. Sham rats underwent identical surgical procedures, with the omission of the fluid percussion application. Rats were randomly assigned to receive one of four treatments: FPI with vehicle; FPI with EPC treatment; sham FPI; and sham FPI with EPC treatment.
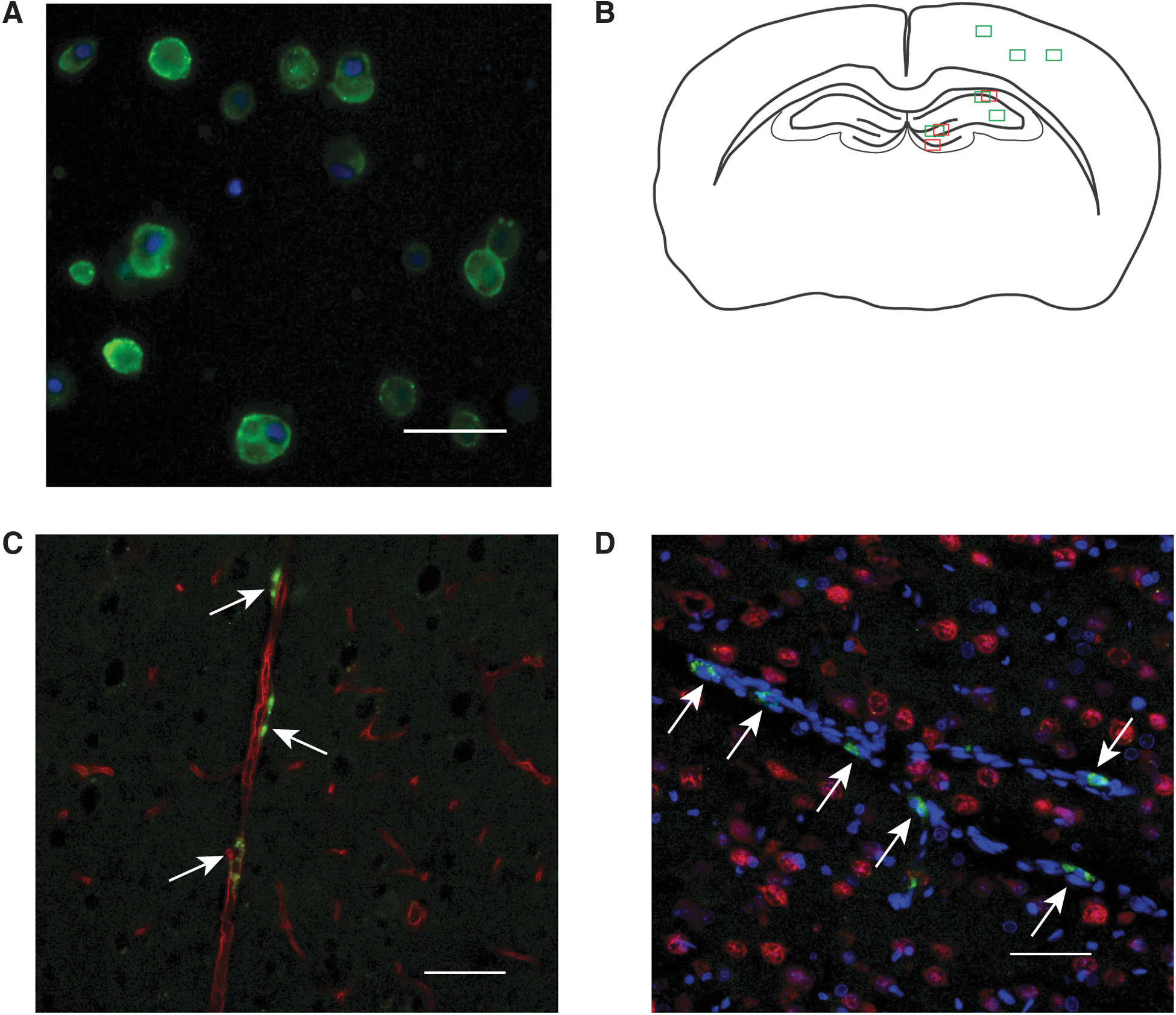
Twenty-four hours post-injury, rats were reanesthetized with 2% isoflurane gas. Body temperature was maintained on a heating pad. Tail vein injections consisted of 500 μL of vehicle Hanks' balanced salt solution (HBSS) or 4- to 5 × 106 EPCs in 500 μL of HBSS. In a subset of rats (n = 4), EPCs were fluorescently labeled using Paul Karl Horan (PKH) 67 (Sigma-Aldrich, Oakville, Ontario, Canada), a lipophyllic fluorescent cell dye, before injection, which allowed visual tracking of EPCs in vivo (Fig. 1A). The PKH67 dye was used at a final concentration of 1 × 10–6 M with a 3-min incubation process for cell labeling.
(
Bone-marrow–derived endothelial progenitor cell cultures
Although the precise definition of EPCs remains somewhat controversial, for simplicity we refer to the bone marrow–derived cultured cells grown in endothelial growth medium-2 (EGM-2) as EPCs. We have previously described methods for procuring and culturing EPCs and have characterized cell-surface expression of markers (VEGF receptor [VEGFR], CD34, and CD133), which are consistent with the EPC phenotype described in the literature. 12,25 Briefly, the femur and tibia were removed from male Sprague-Dawley rats (∼150 g), and all surrounding connective tissue and muscle were dissected. The proximal and distal epiphyses of the femur and tibia were cut, and HBSS was used to flush out the bone marrow. The cell slurry was collected and centrifuged at 400g for 5 min. The pellet was resuspended in EGM-2 (Lonza, Walkersville, MD) and transferred into uncoated T75 flasks. Cells were placed in an incubator maintained at 37°C with 5% CO2. After 24 h of initial plating, the media was replaced with fresh EGM-2. Adherent cells were grown for 8–9 days with media changes every 3–4 days, resulting in a confluent monolayer of cells by day 9. Cells were not passaged and only used from initial plating cultures. Cells were removed from flasks by trypsinization using 0.25% trypsin ehtylenediaminetetraacetic acid (EDTA) solution. Trypsin was inactivated with 15% fetal bovine serum in EGM-2. Cell numbers were quantified with a hemocytometer before injection.
Immunohistochemistry
Rats evaluated in behavioral analysis were sacrificed by transcardial perfusion at 30 days post-injury for immunohistochemistry (IHC) analysis (n = 9 per treatment group). The separate PKH-labeled cohort of rats was sacrificed at 48 h post-injury and 24 hours post-injection. A 1-week survival group was also included in the study, which consisted of n = 6 rats per treatment (sham control, FPI+vehicle, and FPI+EPC). Transcardial perfusion with isotonic saline was followed by perfusion with cooled 4% paraformaldehyde (PFA). Brains were then extracted and post-fixed in 4% PFA for 24 h before cryoprotection in 30% sucrose in phosphate-buffered saline (PBS). Cryoprotected brains were snap frozen in optimal cutting temperature embedding media and sectioned at 20 μm for rat endothelial cell antigen (RECA-1) immunolabeling and at 12 μm for neuronal nuclei (NeuN) labeling.
Nonadjacent sections were sampled from the ipsilateral hemisphere at approximately bregma −1.80 mm and closer to the injury epicenter at approximately bregma −3.60 mm (based on stereotaxic coordinates in Paxinos & Watson, 4th ed.). Two sections per sampling location were used in quantitative analysis of RECA-1 labeling in 7- and 30-day post-injured tissues. Sections were blocked for 1 h in 0.1 M of PBS containing 10% normal goat serum and 0.1% Triton-X. Primary incubation of mouse anti-RECA-1 antibodies (Abcam, Cambridge, MA) was performed at a dilution of 1:500 in blocking solution overnight at 4°C. Secondary incubation was performed with Alexa 488 goat anti-mouse antibodies (Molecular Probes, Burlington, ON, Canada) for 1 h at room temperature (RT) in blocking solution at a dilution of 1:500. Seven-day post-injury tissues were labeled with polyclonal anti-doublecortin (DCX), a marker for immature neurons undergoing neurogenesis (Abcam). 26 The same blocking procedure as previously described was used for DCX labeling. Primary antibody incubation was performed at a dilution of 1:800 overnight in blocking solution. Secondary goat-anti-rabbit Alexa 555 was used for visualization. For PKH67, RECA-1 double-label experiments, Alexa 555 goat-anti-mouse antibodies (Molecular Probes) were used at a dilution of 1:500. Mouse monoclonal anti-NeuN (Millipore, Billerica, MA) labeling for identification of neurons was performed at a dilution of 1:1000 in blocking solution overnight at 4°C. Secondary incubation with Alexa 488 goat-anti-mouse antibodies was performed for 1 h at RT in blocking solution at a dilution of 1:500. Three washes were made in 0.1 M of PBS between each step. Negative controls were run simultaneously with the omission of primary antisera. Hoechst nuclear counterstaining was also applied to each IHC tissue section.
Immunohistochemistry quantification
Sections labeled with NeuN, RECA-1, and DCX were visualized on a Nikon Eclipse 90i epifluorescent microscope (Nikon Corporation, Tokyo, Japan). Image fields were captured using a 20 × objective (Fig. 1B). For RECA-1 quantification, three sampling fields were chosen and values averaged from the cortex of each brain section. In the hippocampus sections, image fields were obtained from the cornu ammonis (CA) 1 region, the stratum radiatum adjacent to the CA2/3 region, and the hilus of the dentate gyrus (DG). Image fields for NeuN quantification in the hippocampus were obtained from the CA1 region, the DG granule cell layer, and the hilus of the DG. The number of neurons quantified in each region was expressed as the number of cells per unit length in order to account for potential biases in total number attributed to differences in section orientation while field sampling. Manual counts of NeuN-labeled cells, which colocalized with an identifiable Hoechst stained nuclei, were performed in the hippocampus sampling regions described above. DCX quantification was performed in the granule cell layer of the DG for comparisons of neurogenic activity between treatment groups. A digital mask was applied using standardized threshold levels for all images to calculate relative density of Alexa 555 labeling. A region of interest, which included only the granule cell layer, was used for quantification of DCX labeling.
NIS Elements software (Nikon Instruments Inc., Melville, NY) was used to quantify the capillary width as well as the capillary density in each sampling field. Capillary density was expressed as the percent area coverage of the total field of view using a 20 × objective. All quantifications were performed by a researcher blinded to the treatment groups.
Western blotting
Brain tissue samples were dissected from the ipsilateral injured hippcampus of sham, injured, and EPC-treated rats at 72 h post-EPC injection (n = 6 per treatment group with the exception of the sham+EPC treatment group; n = 4). Tissue sections were dissected adjacent to the injury epicenter from the region encompassing Bregma −2.0 to −3.0 mm. Samples were homogenized in lysis buffer containing protease inhibitors (50 mM of Tris-HCl, 1% NP-40, 150 mM of NaCl, 1 mM of EDTA, 1 mM of phenylmethylsulfonyl fluoride, 1 mM of NaF, 1 μg/mL of aprotinin, 1 μg/mL of leupeptin, and 1 μg/mL of pepstatin). Protein quantification was determined by the modified Lowry method. 27 Samples were normalized for equal loading (40 μg/lane) and were electrophoresed on 7.5% sodium dodecyl sulfate/polyacrylamide gel electrophoresis gels and transferred to nitrocellulose membranes. Blocking of membranes was performed in 5% nonfat milk solution for 1 h at RT. Expression of activated caspase-3 was examined in sham-, FPI+vehicle-, and FPI+EPC-treated rats (n = 3 per treatment group) using a rabbit polycloncal anti-cleaved caspase-3 antibody (Cell Signaling, Whitby, Ontario, Canada) at a dilution of 1:200 in blocking solution. Expression of calpain-mediated breakdown of spectrin was assessed using a previously described custom antibody directed against the 145-kDa fragment at a dilution of 1:1000 in blocking solution. Primary antibody incubations were performed overnight at 4°C. Anti-GAPDH (glyceraldehyde 3-phosphate dehydrogenase) antibodies (Cell Signaling Technology, Danvers, MA) were used as a loading control at a dilution of 1:6000. Each incubation step was followed with three changes of Tris-buffered saline with Tween 20. Secondary antibody incubations with either goat-anti-rabbit horseradish peroxidase (HRP) or goat-anti-mouse HRP were performed at a dilution of 1:3000 for 1 h at RT. Visualization of signals was performed with an enhanced chemiluminescence kit and detection using a Gel-dock imaging station (Bio-Rad, Mississauga, Ontario, Canada).
Behavioral evaluation
Rats were evaluated for locomotor coordination and memory function up to 30 days post-injury. Rats were tested in an MWM on days 10, 14, 17, 21, and 30 post-EPC or vehicle treatment. A rotarod task was also applied beginning on day 7, with subsequent testing days identical to those used for MWM testing. Treatment groups consisted of n = 12 in sham-, n = 12 FPI+EPC-, and n = 12 FPI+vehicle-treated rats.
The MWM task consisted of a black-tarp–lined circular pool with a diameter of 180 cm. The pool was filled with water to a depth of 30 cm maintained at 22°C with a submersible thermometer. A black circular platform with a diameter of 10 cm was submerged 1 cm below the surface in a pre-determined quadrant for the duration of the study. Environmental cues were placed on the walls for the experiment and were not moved for the duration of the study. Rats were randomly assigned a quadrant for insertion and allowed to explore the maze for 90 sec. Rats that did not locate the platform in the allotted time were placed onto the platform for 15 sec before removal from the pool and towel dried. Each rat was given three trials per session with a minimum 5-min interval between trials. Noldus EthoVison XT video tracking software (Noldus, Leesburg, VA) was used for data analysis to compute the latency to target, velocity, and time spent in each quadrant.
The rotarod task consisted of placing rats on a rotating drum for a maximum of 90 sec. The drum would accelerate over a 45-sec period starting from 1 revolution per minute (rpm) until reaching a maximum of 15 rpm. Automated measurements for time spent on the beam as well as the total distance traveled were recorded. Each rat was pre-trained for three trials on day 6 before data collection on day 7. For each testing session, rats were given three trials with a minimum of 5 min of rest between trials.
Statistical analysis
Quantitative IHC for RECA-1, NeuN, and DCX quantification was analyzed using a one-way analysis of variance (ANOVA) with the post-hoc Holm-Sidak test to compare between sham-, FPI+vehicle-, and FPI+EPC-treated rats. Western blot data were analyzed using a one-way ANOVA with the post-hoc Holm-Sidak test. Behavioral analysis was performed using a two-way repeated measures ANOVA. Statistical significance was assumed at p < 0.05.
Results
Bone-marrow–derived allogeneic cultured endothelial progenitor cells home to the ipsilateral injured brain
We have previously described surface expression of VEGFR2, CD133, and CD34 in bone-marrow–derived progenitor cells. PKH-labeled EPCs were detected in the injured cortex at 24 h post-injury. Localization of EPCs was found to be both within and adjacent to existing microvasculature (Fig. 1C,D) of the ipsilateral injured cortex and cerebellum. We did not detect PKH-labeled cells in 7-day post-injured brain sections.
Endothelial progenitor cell treatment reduces caspase-3 activation and poly (ADP-ribose) polymerase cleavage
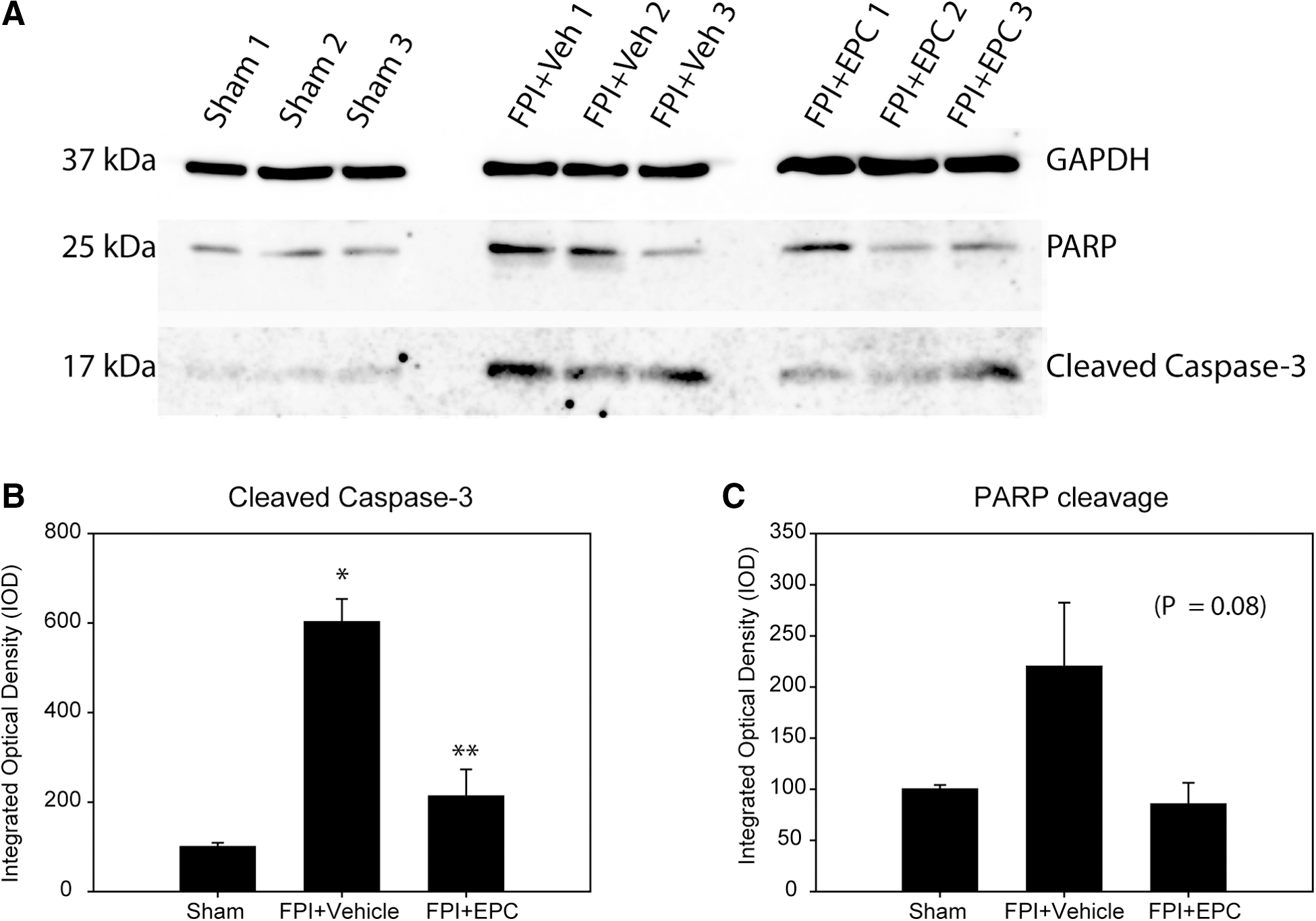
We examined the effects of EPC treatment on signaling pathways resulting in caspase-3 activation and PARP cleavage. Western blot analysis of tissues from 72-h post-injection rats indicated a significant effect of EPC treatment on expression of cleaved caspase-3 in hippocampus tissue homogenates (p < 0.001; Fig. 2A,B). Densitometry analysis indicated an expected significant increase in caspase-3 activation in FPI+vehicle-treated rats (p < 0.001). In the FPI+EPC treatment group, there was a significant reduction in caspase-3 activation relative to FPI+vehicle-treated rats (p < 0.002). There was no significant difference between FPI+EPC-treated rats and sham rats. There was no significant difference in caspase-3 activation in tissue homogenates between sham animals and those sham animals that received EPC injections (data not shown).
(
The 25-kDa cleavage product of PARP, known to be produced through caspase-3 activity, was also examined. PARP expression in treatment groups approached significance (p = 0.08), whereas the data trends were supportive of a similar pattern of reduction in PARP cleavage with EPC treatment (Fig. 2C,D). Similar to caspase-3 activation in sham+EPC-treated rats, we observed no significant effect of EPCs on PARP cleavage products in sham rats treated with EPCs.
Effect of endothelial progenitor cell treatment on neurogenesis and neuronal cell density in the hippocampus
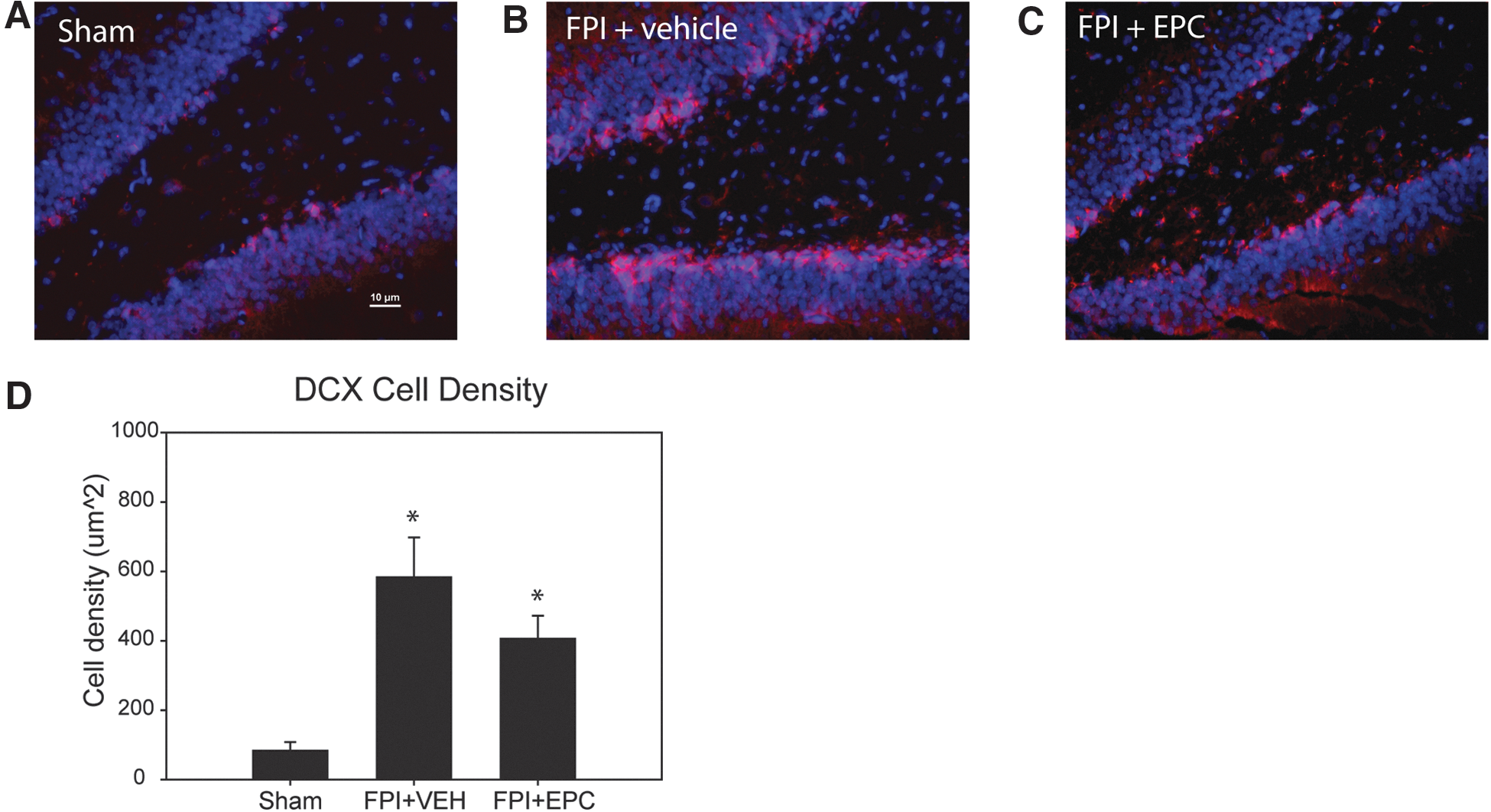
Evaluation of EPC effects on neurogenesis post-injury was performed by quantifying the density of DCX expression at 7 days post-injury. Relatively low levels of DCX expression were observed in the granule cell layer in sham control rats, with a mean density of 83.2 ± 25.0 μm2 (Fig. 3A). In FPI+vehicle-treated rats, there was a dramatic increase in DCX expression at 7 days post-injury, with mean levels of density of 583.3 ± 114.2 μm2 (Fig. 3B). In EPC-treated FPI rats, we observed a mean level of DCX expression of 406.2 ± 66.0 μm2 (Fig. 3C). Mean values between treatment groups were significantly different by one-way ANOVA (p = 0.002). Post-hoc analysis indicated that values from FPI+vehicle compared to sham levels were significantly different (p < 0.001), as were values between FPI+EPC compared to sham (p < 0.001; Fig. 3D). Comparisons between vehicle- and EPC-treated rats indicated no significant difference between treatment groups (p = 0.121), although the data trend suggested less DCX expression in EPC-treated rats.
(
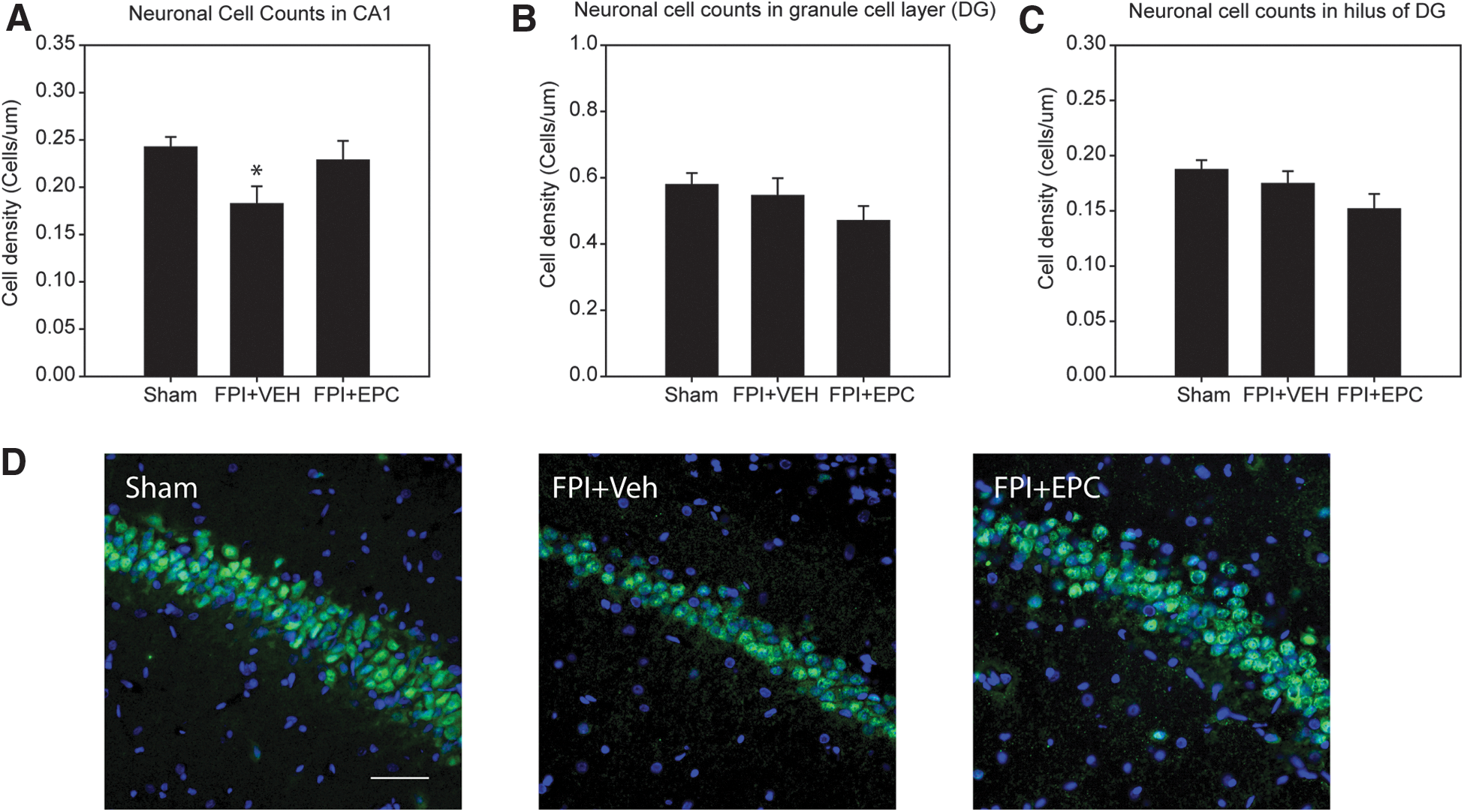
We quantified cells in the hippocampus region from tissues derived from behavioral test rats in order to draw direct comparisons to results from the MWM tests. Quantification of the number of neurons in the cornu ammonis (CA) 1 layer of the hippocampus indicated a significant difference between treatment groups (p = 0.04). The mean quantified cell densities were 0.24 ± 0.01 cells/μm in sham-, 0.18 ± 0.02 cells/μm in FPI+vehicle-, and 0.23 ± 0.02 cells/μm in FPI+EPC-treated rats (Fig. 4A). There were significantly fewer neurons in the FPI+vehicle group relative to sham rats (p = 0.04). However, post-hoc analysis did not detect a significant difference in FPI+EPC-treated rats when compared to either sham or FPI+vehicle rats. Representative images from the CA1 region are shown in Figure 4D.
(
Analysis of neuronal cell counts in the hilus and granule cell layer of the DG indicated no significant difference between treatment groups (Fig. 4B,C). However, there was a trend in the data suggesting that EPC treatment resulted in lower cell density in both regions at 30 days post-injury.
Endothelial progenitor cell treatment improves cornu ammonis 1 microvascular density at 7 days
We examined coronal brain sections for evidence of microvascular recovery post-EPC treatment at 7 and 30 days post-injury. At 7 days post-injury, capillary density in the ipsilateral injured cortex was significantly different between treatment groups (p = 0.014). Capillary density in the FPI+vehicle and FPI+EPC groups were significantly lower than sham density levels (p < 0.05; Fig. 5A). In the CA1 hippocampus, capillary density was significantly lower in the FPI+vehicle group relative to sham (p = 0.002) and significantly lower than FPI+EPC-treated rats (p = 0.003). There was no significant difference between sham and FPI+EPC-treated rats (Fig. 5B,C).

(
At 30 days post-injury, capillary density was analyzed in the same IHC sections used for capillary diameter quantification. CA1 density values were significantly different (p = 0.01) among treatment groups (Fig. 6A,B). Capillary density was significantly lower between FPI+vehicle- and FPI+EPC-treated rats relative to sham rats (p < 0.05). Representative images are shown in Figure 6C. Post-hoc analysis indicated no significant differences in capillary density with EPC treatment relative to vehicle-treated rats. Capillary density in the stratum and DG were not significantly different between treatment groups (data not shown). There was a trend toward statistical difference between treatment groups in the analysis of the cortex (p = 0.09).
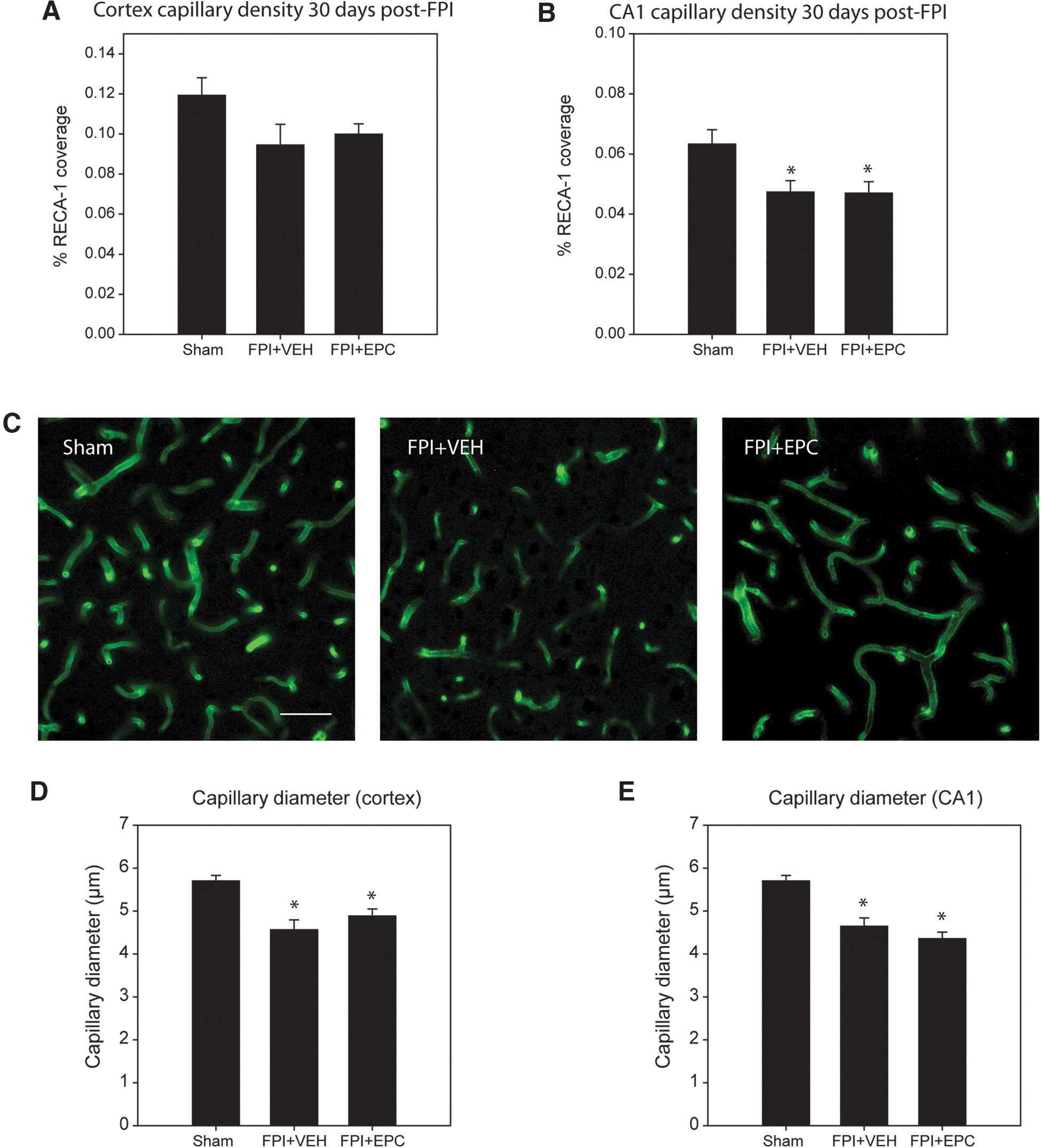
(
Analysis of capillary diameters (vessels with diameters of <10 μm) in the cortex indicated a significant difference between treatment groups (p < 0.001). Post-hoc analysis indicated significantly smaller capillary diameters in FPI+vehicle-treated relative to sham rats (p < 0.001). There was also a significant difference in diameters between sham- and FPI+EPC-treated rats (p = 0.001). There was no significant difference in diameters between vehicle- and EPC-treated rats. Mean capillary diameters in the cortex were 5.87 ± 0.14 μm in sham-, 4.56 ± 0.23 μm in FPI+vehicle-, and 4.88 ± 0.17 μm in FPI+EPC-treated rats (Fig. 6D). Capillary diameters in the CA1 were also significantly different (p < 0.001). Capillary diameters were narrower in vehicle- and EPC-treated groups relative to sham rats (p < 0.001). Mean capillary diameters in the CA1 region were 5.70 ± 0.13 μm in sham-, 4.65 ± 0.19 μm in FPI+vehicle-, and 4.36 ± 0.15 μm in EPC-treated rats (Fig. 6E). Capillary diameters measured from the stratum radiatum and hilus of the DG had similar significant reductions in capillary diameter, as observed in the CA1 region associated with FPI in both vehicle- and EPC-treated rats, but no significant differences between these groups (data not shown).
Effect of endothelial progenitor cell treatment on behavioral outcome measures
We evaluated rats on the rotarod and MWM tests in order to assess locomotor ability and spatial memory recovery, respectively. Rotarod latencies between treatment groups were significantly different by two-way repeated-measures ANOVA (p = 0.001; Fig. 7A). Latency times between sham- and FPI+vehicle-treated rats were significantly different (p < 0.001), as were values between FPI+vehicle and FPI+EPC treatment groups (p = 0.038). Latency times between sham and FPI+EPC treatment groups were not significantly different. There was also a significant effect of treatment dependent on the day of assessment (p = 0.019). On days 17, 21, and 30, results between sham versus FPI+vehicle and FPI+vehicle versus FPI+EPC were significantly different (p < 0.05), with FPI+EPC treatments demonstrating higher latency times spent on the rotarod.
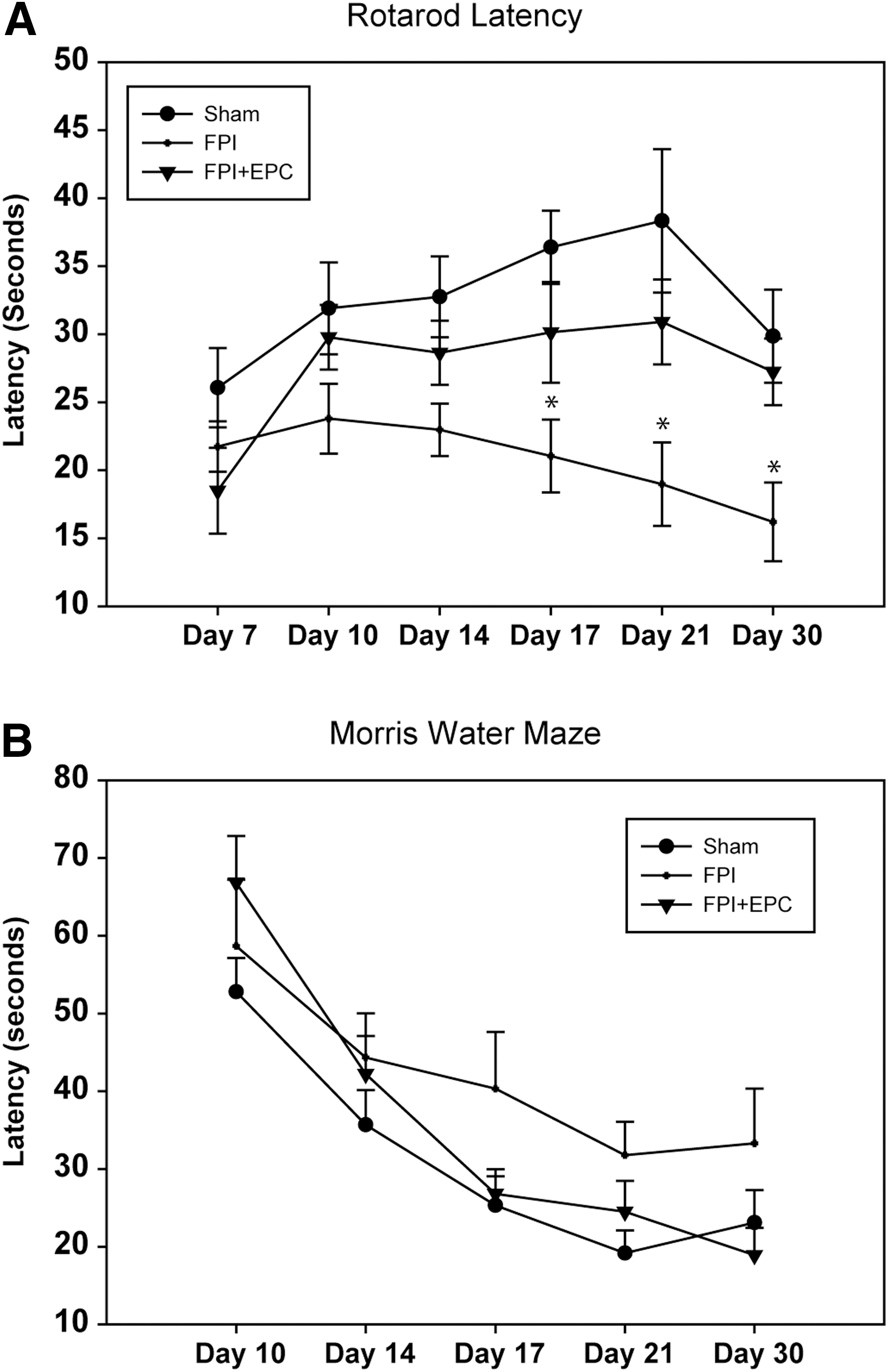
(
Assessment of spatial memory using the MWM failed to demonstrate any significant differences between treatment groups. Overall, each treatment group demonstrated improvements in latency times to the hidden platform over the evaluation period but failed to demonstrate significant intergroup differences (Fig. 7B).
Discussion
The use of endothelial progenitor cells have been increasingly investigated for use as biomarkers or therapeutic strategies for the treatment of numerous disease conditions, including ischemic stroke, 28 TBI, 1 cardiovascular disease, 29 pulmonary hypertension, 30 and diabetes. Similar to mesenchymal stem cells, EPCs can be isolated from bone marrow and in lower numbers from human peripheral circulation. Despite an initially low number derived from host tissues, EPCs demonstrate a high capacity for proliferation in vitro, 23 suggesting the possibility of in vitro expansion as a strategy to increase cell numbers to a level that would augment therapeutic capacity. In the present study, we isolated cells from rat bone marrow and expanded the cells in culture using EGM-2 media, which promoted the differentiation of isolated cells toward an endothelial phenotype. 12 We used these in vitro expanded EPCs to examine the therapeutic effects on outcome after lateral FPI.
Early outcome measures, including expression of activated caspase-3 and PARP cleavage, indicated that EPC treatment reduced induction of apoptotic pathways post-injury. This result was consistent with the therapeutic benefit we observed both in vitro and in vivo when applying cultured EPCs in a mid-line FPI model. 12 The results from subacute outcome measures at 7 and 30 days, including quantitative IHC analysis of capillary density, neuronal cell counts, as well as behavioral measures, were less well defined in regard to the therapeutic benefits of EPC treatment. This highlights the complexity of the pathophysiological response to trauma and the difficulty in treatment, even with a therapeutic approach with presumably pleiotropic neurogenic and angiogenic capacity. Neuronal cell counts from the hippocampus at 30 days post-injury were not statistically significant between injury and treatment, with the exception of the CA1 region, which suggested a potential neuroprotective effect. It is interesting to note that this corresponds to the vascular density improvement observed in the CA1 region in EPC-treated animals at 7 days post-injury. However, by 30 days post-injury, there was heterogeneity in different regions of the brain in regard to the effects of EPC treatment on capillary density and diameter. Qualitative assessments of IHC sections in the cortex suggested some potential improvement in overall capillary density with EPC treatment. However, statistical analysis did not detect a difference between injury and treatment groups. Further, capillary densities in the DG had data trends that suggested worse outcome with EPC treatment, although these differences were not statistically significant. Our sampling sizes were determined a priori based on our previous experience in using RECA-1 and NeuN quantitative IHC as end points for analysis in the lateral FPI model. It is possible that increasing sample size may have clarified the difference attributed to treatment. Our 1-week post-injury data in the granule cell layer of the DG indicates that neurogenesis increases post-trauma. Interestingly, there was no significant difference in DCX labeling between vehicle- and EPC-treated rats. Moreover, the data trend also suggests that EPC treatment may have reduced the degree of neurogenesis in this particular brain region. Inclusion of behavioral data up to 30 days post-injury also highlights the ambiguity of the effects of EPC treatment on longer-term subacute outcome. For instance, the lack of significant effect in the MWM between treatment groups is, in part, consistent with the lack of overall change we observed in neuronal cell counts from the hippocampus regions. Interestingly, analysis of latency times on the rotarod were significantly different with EPC treatment, with results suggesting improved locomotor activity and coordination in EPC-treated rats relative to vehicle controls. These results are also consistent with a previous study published by our group in which virally transfected VEGF-A zinc-finger protein constructs in a lateral FPI model also demonstrated a lack of significant difference between treatment groups in the MWM, but a significant effect in the rotarod task up to 30 days post-injury. 6 This phenomenon of lack of sensitivity using the MW has also been reported in other TBI studies, particularly in instances where repeated measures are performed for behavioral assessments at later post-injury time points. 31,32 This suggests a lack of sensitivity in the MWM for subacute measurements attributed to compensation or spontaneous recovery, or, alternatively, that the rotarod may be a more sensitive measure for evaluating behavioral outcome in later post-injury assessments. Another possibility may be a confounding effect of using the same rats on rotarod testing as were used for MWM testing. Several studies have documented improvements in cognitive function after modeled TBI with the introduction of voluntary exercise. 33,34 More specifically, delayed introduction of exercise has been shown to improve cognitive function. 35 In our study, rats were subject to rotarod testing before MWM testing and may have contributed to a plateau effect, leading to a lack of significant effect with EPC treatment relative to vehicle controls.
The procurement of EPCs for this study did not consist of a homogenous population of cells. As we have previously described, there is a significant portion of CD11b/c-positive cells that are consistent with a macrophage phenotype. Whereas evidence suggests that allogeneic EPCs and their differentiation into mature endothelial cells are immune privileged, 36,37 it is unclear whether the introduction of allogeneic macrophages and their secreted inflammatory products would be beneficial, insignificant, or detrimental in the current model. The fact that we observed EPCs within the injured cortex suggests that the homing capacities of EPCs to injured neuronal tissues played some role in reparative or protective processes, but it is not clear whether these were cells of an EPC phenotype or macrophages. Our results indicating the presence of administered cells in the injury penumbra, but not at later stages, is consistent with other studies of in vivo tracking of EPCs, suggesting that their migration and residence in injured tissues is transient and short-lived. 5 In our current study, we were unable to detect PKH67-labeled cells in the brain at 7 days despite the dye demonstrating effectiveness in vitro for at least 30 days (data not shown), and an in vivo half-life of 10–12 days according to the manufacturer. This would suggest that cells did not incorporate into the damaged tissues or had migrated away at an earlier time point. These findings are consistent with our previous study using exogenous bone-marrow–derived EPCs in a mid-line FPI model where we were unable to detect PKH-labeled cells in vivo at 5 days post-injury. 12 This short duration from a single bolus administration of EPCs post-injury agrees temporally with the observed reduction in caspase-3 and PARP cleavage expression at 72 hours post-injection. However, the lack of more-significant effects in longer-term outcomes may reflect the lack of cellular activity or availability of secreted factors produced by EPCs at later time points. It is also important to consider the timing implications of the various pathophysiological and reparative processes occurring post-trauma. Administration of EPCs may not optimally coincide with various pathways of repair being initiated at the time of cell administration. Mismatched timing between neurogenic and angiogenic pathways occurring post-trauma may not fully benefit from the single application of EPC treatment. Several groups have examined the effects of increasing the homing capacity of EPCs to injured tissues through nanoparticle magnetic coupling. 38,39 These studies suggest that increasing the number of EPCs that home to the site of injury can improve outcome beyond the endogenous homing mechanisms. Whether there would be benefit from increasing the time in which EPCs reside in injured tissues remains to be investigated.
The discrepancy of therapeutic effects between acute reduction of caspase-3 activation and PARP cleavage at 3 days post-injury, relative to subacute microvascular and neuronal outcome at 30 days post-injury, highlights the importance of longer-term outcome measures when evaluating therapeutic strategies. However, this also indicates potential to optimize treatment strategies in the use of EPCs or their secreted products. Sustained duration or availability of EPCs or EPC-derived factors in the injury penumbra may be required to observe longer-term beneficial effects.
Conclusion
We demonstrate that allogeneic in vitro expanded EPCs have therapeutic potential when administered 24 h post-injury. The therapeutic effects are largely limited to acute reduction in caspase-3 activity and some early improvement in CA1 vascular density. Our results also reinforce the notion that the therapeutic effects of EPCs may be short-lived and may require further optimization or repeat administration for optimal subacute effectiveness.
Footnotes
Acknowledgments
This study was funded by the Ontario-China Research Innovation Fund provided through the Government of Ontario, Ministry of Economic Development and Innovation.
Author Disclosure Statement
No competing financial interests exist.