
Abstract
There is a great need to identify potential long-term consequences of contact sport exposure and to identify molecular pathways that may be associated with these changes. We tested the hypothesis that football players with (Ath-mTBI) (n = 25) and without a concussion history (Ath) (n = 24) have altered resting state functional connectivity in regions with previously documented structural changes relative to healthy controls without football or concussion history (HC) (n = 27). As a secondary aim, we tested the hypothesis that group differences in functional connectivity are moderated by the relative ratio of neuroprotective to neurotoxic metabolites of the kynurenine pathway. Ath-mTBI had significantly increased connectivity of motor cortex to the supplementary motor area relative to Ath and HC. In contrast, both Ath-mTBI and Ath had increased connectivity between the left orbital frontal cortex and the right lateral frontal cortex, and between the left cornu ammonis areas 2 and 3/dentate gyrus (CA2-3/DG) of the hippocampus and the middle and posterior cingulate cortices, relative to HC. The relationship between the ratio of plasma concentrations of kynurenic acid to quinolinic acid (KYNA/QUIN) and left pregenual anterior cingulate cortex connectivity to multiple regions as well as KYNA/QUIN and right CA2-3/DG connectivity to multiple regions differed significantly according to football and concussion history. The results suggest that football exposure with and without concussion history can have a significant effect on intrinsic brain connectivity and implicate the kynurenine metabolic pathway as one potential moderator of functional connectivity dependent on football exposure and concussion history.
Introduction
A
Several studies have demonstrated that SRC and mild traumatic brain injury (mTBI) are also associated with disruptions in resting state functional connectivity MRI (fcMRI) at various time points following injury. 11 –15 Few studies, however, have specifically investigated the effects of concussion history on fcMRI or the relationship between fcMRI and structural abnormalities associated with concussion history. A recent study observed thinner cortex in the left anterior temporal lobe in retired football athletes with a history of multiple concussions relative to controls, which was accompanied by an increase in resting state connectivity to the orbital frontal cortex (OFC). 16 A second study found altered default mode network (DMN) connectivity in high school and collegiate-aged athletes. 6 More specifically, asymptomatic hockey players with a concussion history had reduced DMN connectivity in the medial prefrontal cortex (mPFC) and increased DMN connectivity in the precuneus (PCUN) and middle cingulate cortex (MCC) relative to athletes with no concussion history, as well as increased connectivity between the mPFC and the left inferior frontal gyrus. 6 Further, these connectivity abnormalities correlated with white matter integrity in the corpus callosum and anterior coronal radiata in athletes with concussion history. Despite these recent studies, the extent to which structural changes observed in young athletes with varied concussion history affect intrinsic functional connectivity remains unclear.
In addition to characterizing changes in brain structure and function associated with repeated exposure to sport-related brain injury, there is also a great need to identify the molecular mechanisms underlying these structural and functional abnormalities. Glutamate-mediated excitotoxicity caused in part by activation of the kynurenine (KYN) pathway has been suggested as one such mechanism in a variety of neurodegenerative and psychiatric diseases.
17
Specifically, pro-inflammatory cytokines activate the enzyme indoleamine 2,3, dioxygenase (IDO), increasing the production of KYN from tryptophan.
18,19
KYN is in turn metabolized into 3-hydroxykynurenine (3HK) by the enzyme kynurenine monooxygenase (KMO) which also is upregulated under inflammatory conditions, leading ultimately to a relative increase in the neurotoxic metabolites, 3HK and quinolinic acid (QUIN), and a relative decrease in kynurenic acid (KYNA) (see Fig. S1) (see online supplementary material at
We previously reported that the putative “neuroprotective indices” KYNA/3HK and KYNA/QUIN were inversely correlated with hippocampal and amygdalar volumes in patients with major depressive disorder (MDD) and bipolar disorder 24,25 and with cortical thickness in the mPFC in MDD patients. 26 Similarly, with regard to SRC, we recently found that 3HK inversely correlated with hippocampal volume in collegiate football players with a history of concussion, whereas KYNA/QUIN positively correlated with hippocampal volume in football players without a history of concussion. 9 Therefore, alterations in the concentrations of neuroactive kynurenine pathway metabolites may represent one conceivable mechanism behind the structural and functional changes associated with repetitive brain injuries and concussions caused by sport.
The primary aim of this article was to test whether fcMRI abnormalities are present in football athletes with a history of concussion relative to football players without a history of concussion and healthy controls with no football exposure or previous concussion (i.e., to examine the long-term effects of concussion). Second, we tested whether fcMRI abnormalities are present in a group of football athletes without a history of concussion relative to healthy controls, to determine whether football participation is associated with functional changes in the brain (i.e., to examine the long-term effects of repetitive subconcussive brain injuries). We hypothesized that regions with previously documented structural changes in this sample (i.e., CA2-3/DG, vmPFC, and motor cortex) would show abnormal functional connectivity in football athletes with a history of concussion, and to a lesser degree in football athletes without a history of concussion, relative to healthy controls. Finally, we tested the hypothesis that the effects of repetitive brain injuries and concussion history on functional connectivity are moderated by plasma KYNA/QUIN and KYNA/3HK.
Methods
Participants
Male student-athletes from a Division I American-rules football program, noncontact sport collegiate athletes, and nonathletes were recruited for this study. Participants with a current or past history of substance/alcohol abuse or diagnosis of mood/anxiety disorder were excluded. Football players self-reported the number of previous concussions, defined as concussions that were medically diagnosed by a clinician at the time of injury. A total of 49 collegiate football players were enrolled. Twenty-five football players had at least one self-reported incidence of a clinically diagnosed concussion (Ath-mTBI) and 24 self-reported no history of concussion (Ath). One participant was a track and field athlete with a history of concussion and extensive football experience (9 years) and was therefore included in the Ath-mTBI group. All football athletes with a history of prior concussion had complete resolution of symptoms from the most recent concussion, as determined by clinicians trained in sports medicine who functioned independently of the current study. Twenty-seven male noncontact sport athletes and non-athletes were recruited as an additional control group (HC). These participants self-reported no previous concussions and had no football experience. Written informed consent was collected from all participants, and an institutional review board approved this study. Behavioral, demographic, and MR structural data for this sample have been presented in separate articles, and, therefore, are not reproduced here. 9,10
Blood processing
A non-fasting afternoon blood sample was collected in 6 mL K2 EDTA BD Vacutainer™ tubes (Becton, Dickinson and Company, Franklin Lakes, NJ), centrifuged at 1300g for 10 min at room temperature, and plasma was collected. As previously described, plasma was aliquoted into microfuge tubes and stored at −80°C that day or the following morning after being stored overnight at 4°C.
9
Frozen plasma samples were shipped overnight to Brains Online (
Blood was collected within 1 month of the scanning data, on average 0.92 days prior to the scanning session. Not all participants participated in the blood draw (see Table 1 for the number of participants with blood data and with both blood and fcMRI data). In addition, participants with non-brain injuries or infections at the time of blood draw were excluded from analyses. Group differences in KYN metabolites in this sample have been previously investigated and are not reported here. 9
Time since first documented concussion not available for six Ath-mTBI participants.
Not included in blood analyses.
Medication on scanning session, completed blood sample unmedicated,
Reported for recurring headaches.
p < 0.05.
ADHD, attention-deficit/hyperactivity disorder; Ath-mTBI, football players with concussion history; Ath, football players without concussion history, BMI, body mass index; fcMRI, functional connectivity magnetic resonance imaging; HC, healthy control.
fcMRI parameters and preprocessing
MRI was performed using a General Electric Healthcare Discovery MR750 3-Tesla whole body MRI scanner and brain-dedicated receive-only 32-element coil array optimized for parallel imaging (Nova Medical, Inc.). T1-weighted structural images were collected using a parallelized magnetization-prepared rapid gradient-echo sequence with sensitivity encoding with field of view (FOV) = 240 mm, 130 axial slices, slice thickness = 1.1 mm, image matrix = 256 × 256, repetition time/echo time (TR/TE) = 5 ms/1.948 ms, acceleration factor R = 2, flip angle = 8 degrees, TI = 725 ms, sampling bandwidth = 31.25 kHz, and voxel size = 0.9375 × 0.9375 × 1.1 mm. In addition, a 6 min gradient-echo echo-planar image (EPI) sequence was used to collect 180 volumes of resting state data during which participants were instructed to fixate on a cross and think of nothing in particular (FOV = 240 mm, acquisition matrix = 96 × 96, slice thickness = 3 mm, interslice spacing = 0.2 mm, 37 axial slices, TR/TE = 2 sec/30 ms, flip angle = 90 degrees, sampling bandwidth = 250 kHz, acceleration factor R = 2, acquired voxel size = 2.5 × 2.5 × 3.2 mm interpolated to 1.875 × 1.875 × 3.2 mm).
Anatomical images were skull stripped and transformed to standard space using the Advanced Normalization Tools (ANTS) by applying a 12 parameter affine transformation followed by a nonlinear warp. 27 An eroded white matter mask was calculated following tissue segmentation. A bilateral lateral ventricle mask was extracted in native space following automated segmentation via Freesurfer v5.1, 28 transformed to standard space, and eroded by 1 voxel along the x, y, and z directions to ensure exclusion of non-cerebrospinal fluid (CSF) voxels. The AFNI software suite was used to proces EPI data. 29 The first four EPI volumes were removed and AFNI's despike program was used to replace any anomalous time-series data. Following slice-time correction, volumes were registered to a single volume to account for head motion using a 6 degree of freedom transformation, aligned to standard space using both the transformation and nonlinear warp calculated for the anatomical image, and resampled to 1.75 mm isotropic voxel size. A spatial smoothing kernel with full-width at half-maximum (FWHM) of 4 mm was applied prior to nuisance regression. The six rigid-body motion parameters and their derivatives, the average lateral ventricle signal, the averaged local white matter signal, 30 and bandpass filter frequencies (0.01–0.10 Hz) were removed from the time series as signals of no interest. A single measure of head motion was calculated as the Euclidian norm of the six motion parameters. 31 Individual time points with Euclidian norm of motion > 0.30 were censored from analyses along with the preceding time point. Three HCs and one Ath did not have resting state data available. In addition, individual scans with evident artifacts or <128 usable data points following censoring were excluded from imaging analyses, including one Ath, one Ath-mTBI, and two HCs. The final number of subjects with usable fcMRI data can be found in Table 1.
fcMRI seed-based analyses
Seed-regions for connectivity analyses were identified from our previous studies of cortical thickness and hippocampal volume in the same sample. 9,10 Significant clusters identified from our cortical thickness analyses were located in the right motor cortex and left ventromedial prefrontal cortex (vmPFC). The clusters were extracted from the cortical ribbon and transformed to the volume of the standard template. These data were then smoothed using a 4 mm FWHM kernel and resampled to 1.75 isotropic voxels to match the resting state data. Voxels with maximum t values (indicating voxels with the greatest cortical thickness differences from the identified comparison) were selected from each cluster. The vmPFC cluster covered a large area in the volume and, therefore, two separate maximums were chosen in the pregenual anterior cingulate cortex (pgACC) and the OFC. Five mm radius spherical regions of interest (ROIs) were then created at each peak voxel for the left pgACC (MNI coordinates −4, 37, 0), left OFC (−6, 50, −17), and right motor cortex (10, −19, 72). Freesurfer software was used to segment bilateral hippocampi into hippocampal subfields in native space for each participant, as previously reported. 9,32 A binary mask was created using voxels with posterior probability of belonging to either the CA2-3 or DG being ≥95%. The combined CA2-3/DG binary mask was then transformed to standard template space using both the affine transformation and nonlinear warp calculated for the anatomical image and resampled to 1.75 mm isotropic voxels.
Voxelwise correlation was performed for the averaged time-series data within each ROI, and a Fisher's z transformation was applied to approximate a normal distribution. An inclusion mask created from a tissue segmentation of the standard template (gray matter probability [GMP] >0.25) was combined with binary masks of normalized anatomical images from each participant, to limit analyses to gray matter in which all participants had EPI coverage.
Statistical analysis
Statistical analyses were performed in IBM SPSS version 21. One way analyses of variance were used to determine group differences in age, education, body mass index (BMI), and head motion during the scanning session. A Kruskal–Wallis test was used to test for group differences in the number of censored fcMRI time points. A two tailed independent samples t test was used to compare years of tackle football experience in football athletes with and without concussion history. The AFNI program 3dMVM was used to conduct voxelwise one way analyses of covariance (ANCOVA) to determine if connectivity to each seed-region differed between football players with and those without a history of concussion, and non-football players (Aim 1). 33 To determine whether plasma levels of KYN metabolites moderated group differences in functional connectivity, general linear models (GLM) with the effects of group, neuroprotective KYN indices (i.e., KYNA/QUIN and KYNA/3HK), and the interaction between group and neuroprotective KYN indices were tested in the subset of participants with both blood and fcMRI data using 3dMVM. Age was used as a covariate in 3dMVM analyses, consistent with our previous work on this cohort. 9,10 The smoothness of the noise was estimated using a non-Gaussian, spherically symmetric autocorrelation function in AFNI, and 10,000 iterations of a Monte Carlo simulation conducted in native voxel space determined the necessary correction for a familywise error rate of p < 0.05 for voxelwise analyses (smoothness parameters = 0.35, 3.9, and 10.9, effective FWHM = 11.6 mm, voxel p value = 0.005, minimum cluster volume = 1384 μL). Macro-labels of the Eickhoff–Zilles brain atlas distributed in the AFNI software were used for anatomical localization of significant clusters. 34 The online Neurosynth database was used to further identify the respective functional networks to which significant clusters belonged. 35 An ROI approach was adopted to perform follow-up tests within the framework of ANCOVA/GLM using average connectivity values within clusters identified from the 3dMVM analyses following voxelwise multiple comparison correction. An α of 0.05 was used for these comparisons.
Results
Demographics and fcMRI quality assurance
Demographic information has been previously reported for this sample and is only briefly reviewed here.
9,10
The median number of months between the scanning date and the most recent concussion for participants with a concussion history was 5 months (range = 1–42 months, Table 1, Fig. S2) (see online supplementary material at
Group differences in fcMRI
Areas of significant differences in fcMRI between groups are outlined in Table 2, and group connectivity maps for each seed-region can be found in Figure S3 (see online supplementary material at

Group differences in functional connectivity. Regions with a significant main effect of group on functional connectivity to seed-regions in the right motor cortex, left orbital frontal cortex (OFC), and left cornu ammonis areas 2 and 3 (CA2-3) and dentate gyrus of the hippocampus are shown (top). Scatterplots show Fisher's z-scored connectivity (y-axis) for football players with a concussion history (Ath-mTBI), football players without a concussion history (Ath), and non-football players (healthy controls [HC]). Horizontal lines represent group means (bottom). SMA, supplementary motor area; PCL, paracentral lobule; MFG, middle frontal gyrus; IFGtr, inferior frontal gyrus pars triangularis; MCC, middle cingulate cortex; PCC, posterior cingulate cortex; PCUN, precuneus.
Ath-mTBI, football players with concussion history; Ath, football players without concussion history; B, bilateral; CA2-3/DG, cornu ammonis 2-3 and dentate gyrus of hippocampus; HC, healthy controls; IFGtr, inferior frontal gyrus pars triangularis; MCC, middle cingulate cortex; MFG, middle frontal gyrus; OFC, orbitofrontal cortex; PCC, posterior cingulate cortex; PCL, paracentral lobule; PCUN, precuneus; R, right; SMA, supplementary motor area.
Interaction of neuroprotective indices and group on fcMRI
A secondary aim of the current study was to determine whether plasma levels of neuroprotective KYN measures (i.e., KYNA/QUIN and KYNA/3HK) moderated the relationship between fcMRI in football players with and without a history of concussion relative to non-football players (i.e., interaction of group by KYN indices). Information on clusters with significant interactions including the statistics for the slope comparisons and simple slopes for each group are presented in Table 3. A significant interaction between KYNA/QUIN and group was observed for pgACC connectivity to the right middle MFG, the left lateral and medial superior frontal gyrus (SFG), and the right anterior insula (aINS) extending into the pars opercularis of the inferior frontal gyrus (IFGop) (Fig. 2). Follow-up analyses showed that the relationship between KYNA/QUIN and pgACC connectivity to the MFG was significantly different in Ath and HC relative to the relationship observed in Ath-mTBI participants, whereas the relationship did not differ between HC and Ath groups. Simple slope analyses showed that there was a significant inverse relationship between pgACC-MFG connectivity and KYNA/QUIN in Ath-mTBI, a significant positive relationship in HC, and a nonsignificant positive relationship in Ath.

Interaction between group and neuroprotective kynurenine indices. Regions where there was a significant interaction between group and the ratio of kynurenic acid to quinolinic acid (KYNA/QUIN) on connectivity of the left pregenual anterior cingulate cortex (pgACC) and right cornu ammonis (CA) 2-3 and dentate gyrus of the hippocampus are shown. Fisher's z-scored connectivity is plotted against KYNA/QUIN for football players with a concussion history (Ath-mTBI), football players without a concussion history (Ath), and for non-football players (healthy controls [HC]). The p value for each slope is shown in Table 3. MTG, middle temporal gyrus; SFG, superior frontal gyrus; aINS, anterior insula; IFGop, inferior frontal gyrus pars opercularis; MOG, middle occipital gyrus; SOG, superior occipital gyrus; FuG, fusiform gyrus; LiG, lingual gyrus; IOG, inferior occipital gyrus.
Ath-mTBI, football players with concussion history; Ath, football players without concussion history; aINS, anterior insula; CA2-3/DG, cornu ammonis 2-3 and dentate gyrus of hippocampus; FuG, fusiform gyrus; HC, healthy controls; IFGop, inferior frontal gyrus pars opercularis; IOG, inferior occipital gyrus; KYNA/QUIN, ratio of plasma concentrations of kynurenic acid to quinolinic acid; L, left; LiG, lingual gyrus; MFG, middle frontal gyrus; MOG, middle occipital gyrus; pgACC, pregenual anterior cingulate cortex; R, right; SFG, superior frontal gyrus; SOG, superior occipital gyrus.
The relationship between KYNA/QUIN and pgACC connectivity to the SFG was also different in Ath and HC than in Ath-mTBI, whereas HC and Ath did not differ. There were negative associations between pgACC-SFG connectivity and KYNA/QUIN in Ath, a nonsignificant negative association in HC, and a significant positive association in Ath-mTBI. Finally, the relationship between KYNA/QUIN and pgACC connectivity to the aINS/IFGop was significantly different in Ath and HC than in Ath-mTBI, and different in HC than in Ath. Ath-mTBI and Ath had significant and nonsignificant negative relationships between pgACC-aINS/IFGop connectivity and KYNA/QUIN, respectively, whereas HC showed a positive relationship.
There was also a significant interaction between KYNA/QUIN ratio and group for right CA2-3/DG connectivity to the left middle and superior occipital gyri (MOG/SOG) and the right fusiform gyrus (FuG), lingual gyrus (LiG), and inferior occipital gyrus (IOG) (Fig. 2). Follow-up analyses showed that the relationship between KYNA/QUIN and connectivity to the MOG/SOG was significantly different in Ath and Ath-mTBI than in HC, and different in Ath than in Ath-mTBI. Simple slope analyses revealed that both Ath and Ath-mTBI had significant inverse relationships between connectivity to the MOG/SOG and KYNA/QUIN, whereas HC had a nonsignificant positive relationship. For connectivity between the right CA2-3/DG and FuG/LiG/IOG, the relationship with KYNA/QUIN was significantly different in Ath and Ath-mTBI relative to HC, and different in Ath relative to Ath-mTBI. There was a significant inverse relationship between FuG/LiG/IOG connectivity and KYNA/QUIN in both Ath and Ath-mTBI, but a nonsignificant positive relationship in HC. There were no significant interactions between group and KYNA/3HK on fcMRI for any seed-region.
Main effects of neuroprotective kynurenine indices on fcMRI
Although it was not a primary or secondary goal of the current work, we observed several clusters in which KYN pathway measures were inversely associated with connectivity when combined across all groups (see Table 4 for list of regions; Fig. 3). For the ratio of KYNA/3HK, there was an inverse relationship with connectivity of the left pgACC seed-region, the left OFC seed-region, and the right motor cortex seed-region. There were no significant associations between either left or right CA2-3/DG connectivity and KYNA/3HK. Supplementary analyses demonstrated that KYNA and 3HK had opposite relationships with connectivity of the pgACC, OFC, and motor cortex seed-regions, with stronger connectivity in the identified clusters positively correlating with 3HK and negatively correlating with KYNA (Table S1) (see online supplementary material at
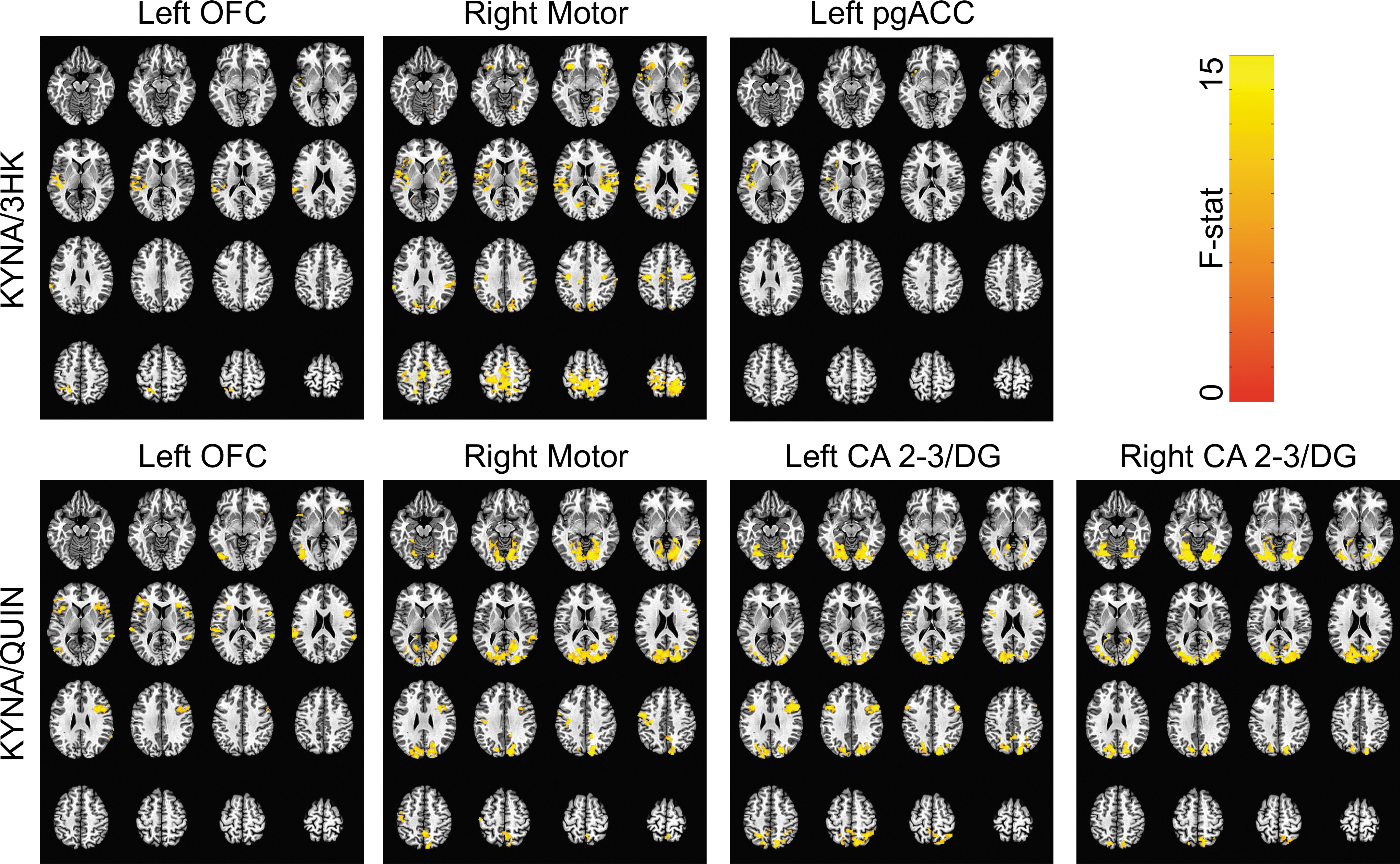
Functional connectivity associated with neuroprotective kynurenine indices across all groups. Clusters in which connectivity to seed-regions in the left orbital frontal cortex (OFC), right motor cortex, left pregenual anterior cingulate cortex (pgACC), and left and right cornu ammonis areas 2-3 and dentate gyrus of the hippocampus (CA 2-3/DG) were significantly associated with either the ratio of kynurenic acid to 3-hydroxykynurenine (KYNA/3HK) or the ratio of kynurenic acid to quinolinic acid (KYNA/QUIN).
aINS, anterior insula; B, bilateral; CA2-3/DG, cornu ammonis 2-3 and dentate gyrus of hippocampus; CAL, calcarine sulcus; CUN, cuneus; FuG, fusiform gyrus; IFGop, inferior frontal gyrus pars opercularis; IFGor, inferior frontal gyrus pars orbitalis; IFGtr, inferior temporal gyrus pars triangularis; IOG, inferior occipital gyrus; IPL, inferior parietal lobule; KYNA/QUIN, ratio of plasma concentrations of kynurenic acid to quinolinic acid; KYNA/3HK, ratio of plasma concentrations of kynurenic acid to 3-hydroxykynurenine; L, left; LiG, lingual gyrus; MOG, middle occipital gyrus; MTG, middle temporal gyrus; OFC, orbitofrontal cortex; PCUN, precuneus; PCL, paracentral lobule; pgACC, pregenual anterior cingulate cortex; pINS, posterior insula; PoG, postcentral gyrus; PrG, precentral gyrus; R, right; ROL, rolandic operculum; SMA, supplementary motor area; SMG, supramarginal gyrus; SOG, superior occipital gyri; SPL, superior parietal lobule; STG, superior temporal gyrus.
Discussion
The primary finding of this study is that collegiate football players both with and without a self-reported history of SRC exhibited varied levels of disrupted functional connectivity to regions with previously demonstrated structural abnormalities. In addition, this study presents the first evidence of relationships between blood levels of neuroactive KYN metabolites and functional connectivity that differ among groups with varying levels of football experience and concussion history. Collectively, these results suggest that football exposure even in the absence of self-reported concussion history is associated with long-term alterations in intrinsic functional connectivity. Further, our results highlight the potential moderating effects of neuroactive kynurenine metabolites on resting state functional connectivity secondary to concussion and/or football exposure.
Group differences in fcMRI
Football players with a history of concussion (median 5 months prior to scanning) had increased connectivity between the right motor cortex to bilateral supplementary motor areas and the paracentral lobule relative to football players without a concussion history and non-football players. These findings are consistent with our previous finding of a thinner motor cortex in football players with a concussion history relative to those without in the same sample,
10
as well as several other previously published studies. For example, altered neurometabolites have been reported within the 1st week of concussion in the primary motor cortex relative to controls,
36
and altered resting state connectivity metrics in the motor cortex have been reported at various time points following mTBI and more severe TBI relative to controls.
37
–40
Consistent with the current work, persistent elevations in intracortical inhibition and reduced glutamate levels have been reported in the motor cortex in athletes with a history of concussion.
41
–44
In the current study, supplementary analyses found a nonsignificant trend for an inverse relationship between time since the most recent concussion and motor cortex connectivity to SMA/PCL (see Supplementary Text) (see online supplementary material at
Importantly, we also found that football players, both with and without a history of concussion, had, relative to non-football players, increased connectivity of the left OFC to the right middle and inferior frontal gyri, as well as left CA2-3/DG of the hippocampus to the MCC and PCC extending into the right precuneus (PCUN). Qualitative inspection of the left OFC and CA2-3/DG reveals that both seed-regions have strong connectivity to the DMN (see Fig. S3), including the mPFC, PCC, and bilateral angular gyri. 45 The increased connectivity between the lateral frontal cortex and the DMN is consistent with previous findings in mTBI patients relative to controls. For example, increased connectivity of the DMN to the frontoparietal and salience networks has been observed in mTBI patients with memory complaints at ∼35 days post-injury, 46 in mTBI patients at ∼11 days and 3–5 months post-injury, 47 in asymptomatic mTBI patients at ∼6 months post-injury, 48 and in asymptomatic collegiate athletes between 3 weeks and 6 months post-concussion. 49 In addition, a recent study found increased connectivity between the mPFC of the DMN and the left inferior frontal gyrus in asymptomatic hockey players with a history of concussion, on average 40 months prior to study relative to teammates with no history of concussion. 6 We extend this previous work by showing that football players without a history of concussion also have altered functional connectivity of the OFC and CA2-3/DG of the hippocampus relative to non-football players, suggesting that football exposure alone without a previous diagnosis of mTBI may alter functional connectivity within frontal and medial temporal structures.
The fact that altered functional connectivity in the OFC was observed in football players with and without a history of concussion is significant, because only football players with a history of concussion had significantly thinner OFC relative to non-football athletes. 10 The observation of functional but not structural differences could reflect differences in the sensitivity of the respective measures. For example, structural MRI does not currently have the resolution to detect subtle dendritic or axonal changes that may affect measures of functional connectivity. Alternatively, it is possible that functional changes associated with multiple years of tackle football exposure precede structural changes, similar to findings that functional activation changes are present in the absence of mesial temporal lobe gray matter changes in asymptomatic adults at genetic risk for Alzheimer's disease. 50,51 Future longitudinal studies of football players with no history of concussion are needed to answer this question.
In contrast to the OFC, we have previously reported smaller hippocampal volumes and, specifically, smaller CA2-3 and DG volumes in football players with and without a concussion history relative to non-football players. 8,9 Here, we also report differences in football players with and without a history of concussion relative to non-football players in left CA2-3/DG functional connectivity to the MCC, PCC, and right PCUN in the same sample. Previous studies have shown abnormalities in hippocampal functional connectivity following mTBI relative to controls, including reduced interhemispheric connectivity in asymptomatic athletes at 10 days post-concussion, 52 decreased connectivity to the inferior frontal gyrus and increased connectivity to FuG and PCUN within 24 h following mTBI, 53 and reduced connectivity to DMN at 7 days following sport-related concussion. 54 Importantly, these studies were conducted at the acute and subacute stages following injury, which could potentially explain the apparent discrepancy between the previous studies documenting reduced connectivity and the results of increased connectivity reported here. In the current study, the areas of increased CA2-3/DG connectivity were in and near the borders of the DMN extending into neighboring networks such as the frontoparietal network, as previously defined (see Fig. S3 for group connectivity maps). 55 One hypothesis is that this reflects the de-differentiation of typically well-defined resting state networks because of the persistent neuronal and metabolic effects of mTBI and subconcussive brain injuries, analogous to similar hypotheses put forth in aging. 56 However, these findings could also reflect the persistent recruitment of additional brain regions as a compensatory mechanism for the observed structural changes or other effects associated with football exposure. 57 Regardless, our data from the current sample suggest that repeated exposure to blows to the head below the threshold of a diagnosable concussion (i.e., football exposure without concussion history) may alter both hippocampal structure and functional connectivity. 9,10 Additional studies are needed to determine how persistent these changes are and whether abnormalities observed in young athletes are related to abnormalities in brain structure and function that have been reported in retired athletes. 58 –61
It is important to note that the regions in which we have identified group differences in functional connectivity also differed structurally among groups, with the exception of the previously discussed findings in the OFC. 9,10 Postmortem evidence of cortical thinning in severe TBI has been linked to reductions in the loss of cortical neurons, 62 and additional factors such as a loss of neuropil have been linked with volumetric differences in other diseases. 63 Therefore, it is unsurprising that neuronal differences and/or differences in neuropil would be associated with differences in functional connectivity of the affected region. However, gray matter volume reductions have been observed without concomitant changes in functional connectivity, for example, in temporal lobe epilepsy. 64 The impact of coincidental volumetric differences on functional connectivity remains unclear, and is worthy of further study.
Potential effects of neuroprotective indices on fcMRI
The secondary goal of the current work was to test the hypothesis that the long-term effects of football exposure and concussion history on fcMRI are moderated by neuroprotective KYN metabolite ratios (i.e., KYNA/QUIN and KYNA/3HK). Although neither KYNA/QUIN nor KYNA/3HK moderated the group differences in connectivity in the regions observed in the primary analyses, the relationship between KYNA/QUIN and functional connectivity to the left pgACC and right CA2-3/DG seed-regions differed as a function of football experience and concussion history. Therefore, our data suggest that individual differences in the relative blood levels of neuroprotective to neurotoxic KYN metabolites are inversely associated with abnormalities of resting state functional connectivity commonly reported following mTBI in football players with (for all observed clusters) and football players without (for right CA2-3/DG clusters) a history of concussion.
For example, as discussed, previous research has demonstrated that patients with mTBI commonly have increased connectivity between the DMN and salience or frontoparietal networks as well as decreased connectivity between DMN regions. 47 –49 Here, football players with a previous concussion who had lower blood levels of KYNA relative to QUIN had greater connectivity between the pgACC and regions of the ventral attentional network, also commonly referred to as the salience network or cingulo-opercular network (i.e., MFG and aINS/IFGop). 55,65,66 Conversely, football players with a concussion history who had lower levels of KYNA/QUIN had less connectivity between the pgACC seed-region and the SFG, regions that are both part of the DMN. Our results also suggest that a history of concussion and subconcussive hits alters the relationship between neuroactive KYN metabolites and functional connectivity between the right CA2-3/DG and the visual cortex, which has previously been shown to be sensitive to the effects of concussion. 52
Although the current study is ultimately unable to determine why football players with and without a history of concussion have different relationships between connectivity and neuroactive KYN metabolites than controls, several intriguing hypotheses emerge. The generation of the blood-oxygen-level dependent (BOLD) signal is predominantly reflective of glutamatergic neurotransmission and more specifically the cycling of glutamate and glutamine. 67,68 Whereas KYNA is an NMDA receptor antagonist, QUIN is an NMDA receptor agonist that also potentiates glutamate release and impairs astrocytic reuptake of glutamate. 20 Therefore, chronic activation of the neurotoxic branch of the KYN pathway via repeated inflammatory events, such as from concussion and subconcussive injuries, 69 –71 may facilitate abnormal functional connectivity patterns relative to those observed in healthy controls without concussion or football exposure. However, it is also conceivable that individual differences in KYN metabolites arising from factors such as genetics, history of infection, and physical or psychological stress may influence sensitivity to repetitive brain injury.
An alternative explanation for the group-specific relationships between connectivity and KYNA/QUIN is that repeated brain injuries can result in disruption of the blood–brain barrier, allowing for the passage of KYN metabolites (i.e., KYNA and QUIN) into the brain. 72 Both imaging and blood markers of altered blood–brain barrier have been reported in football players without documented concussion. 73,74 Moreover, QUIN itself has also been shown to alter blood–brain barrier permeability. 75 Bidirectional translational research is needed to determine the causal factors behind the group-specific relationships between kynurenine metabolites and functional connectivity observed in the current study.
Interestingly, we observed no significant interactions between football exposure and KYNA/3HK on functional connectivity. This could possibly reflect the fact that the actions of QUIN, as well as KYNA, are more directly associated with NDMA receptor activity, and, therefore, variations in the BOLD signal, whereas 3HK primarily acts as a free radical generator and does not directly influence glutamatergic neurotransmission. 21,22,68 Moreover, the majority of previously reported abnormalities in KYN pathway metabolites following brain injury have been in QUIN, which may be more sensitive to injury than 3HK. For example, multiple human studies have reported increased QUIN in CSF following TBI, 76 –79 and we have previously reported decreased plasma levels of KYNA/QUIN and increased QUIN in concussed football athletes across the 1st month post-injury relative to healthy football athletes. 80 As previously reported, we found no differences in KYNA/QUIN in the current sample, although we did observe higher QUIN in football players with a concussion history relative to those without. 9 Current results suggest that repetitive brain injury with and without frank concussion alters the effects of neuroactive kynurenine metabolites on brain function even in the absence of group differences in these metabolites.
Finally, we also report several significant relationships between KYNA/3HK and/or KYNA/QUIN and widespread functional connectivity across all groups combined. Interestingly, all significant associations were negative for both KYNA/3HK and KYNA/QUIN relative to every seed-region. It may be noteworthy that with the exception of the observed relationship between connectivity of the right motor cortex seed-region to other sensorimotor areas, all other regions with significant relationships to KYNA/3HK or KYNA/QUIN were in functional networks distinct from the primary network of the relevant seed-region, commonly the ACC, aINS, superior temporal gyrus (STG)/rolandic operculum (ROL), and visual cortex. Additional studies are needed to clarify the relationship between neuroactive KYN metabolites and functional connectivity in healthy cohorts. Nevertheless, these results are the first evidence, to our knowledge, to suggest that the relative balance between KYNA and neurotoxic KYN metabolites could affect intrinsic brain connectivity.
Limitations
Exposure history (i.e., concussion history and tackle football experience) was reliant on self-report, and it is possible that athletes overestimated or underestimated the number of previous concussions. 81 Cytokine data were not available for the current sample, and we are unable to determine whether similar relationships are present between functional connectivity and these inflammatory mediators. Further, we were only able to assess KYN metabolites in the blood. The extent to which systemic KYN levels reflect levels in the brain is unknown, although tryptophan, KYN, and 3HK can cross the blood–brain barrier, and other work has demonstrated both increased QUIN and decreased KYNA in both the CSF and serum in depressed patients relative to controls. 25,82
Another limitation of the current work is the mean difference in age across groups, although age was included as a covariate in all analyses. There was also a group difference in BMI. However, numerous studies have demonstrated that BMI is a misleading metric in football athletes, which is not indicative of adiposity. 83,84 Further, it is possible that other factors associated with athletic competition could explain some of the observed differences relative to the healthy control group, including physical fitness, activity level, and proprioception. Future studies with groups better matched in age, more validated measures of adiposity, and confounds associated with athletic competition are merited. Finally, attention-deficit/hyperactivity disorder (ADHD) was not an exclusionary criterion for the current study. However, supplementary analyses confirmed that excluding the three participants with ADHD resulted in similar findings to those of the full sample (see Supplementary Text).
Conclusion
Current results suggest that football exposure with and even without a self-reported history of concussion can have a significant long-term effect on intrinsic brain connectivity. Although the clinical significance of these findings is uncertain, it is possible that functional brain changes are present in young athletes with repeated exposure to brain injury before the manifestation of symptoms that have been reported in some retired contact sport athletes. 85,86 Further, the observation of altered intrinsic functional connectivity in football players with and without a concussion history may have important public health implications, highlighting the need for safety initiatives that limit overall exposure, rather than focusing on concussion management alone. The current results also highlight the potential role of neuroactive KYN pathway metabolites in altered functional connectivity associated with contact sport exposure. Additional studies are needed to replicate these results and further delineate the role of neuroactive KYN metabolites in the neurofunctional sequelae of repeated brain injury.
Footnotes
Acknowledgments
This research was conducted using internal funds from the Laureate Institute for Brain Research, which is supported by The William K. Warren Foundation. Dr. Meier received support for this work through a project funded through the Research and Education Program, a component of the Advancing a Healthier Wisconsin endowment at the Medical College of Wisconsin. The authors thank the psychiatric assessment team and MRI technicians at the Laureate Institute for Brain Research; Dr. Rashmi Singh and Dr. Patrick Bellgowan for their assistance with data collection; and Christopher Nerio, David Polanski, Dr. Thomas Allen, and Dr. Lamont Cavanagh for athlete referral and injury oversight.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.