
Abstract
Magnesium (Mg2+) homeostasis is impaired following spinal cord injury (SCI) and the loss of extracellular Mg2+ contributes to secondary injury by various mechanisms, including glutamate neurotoxicity. The neuroprotective effects of high dose Mg2+ supplementation have been reported in many animal models. Recent studies found that lower Mg2+ doses also improved neurologic outcomes when Mg2+ was formulated with polyethylene glycol (PEG), suggesting that a PEG/ Mg2+ formulation might increase Mg2+ delivery to the injured spinal cord, compared with that of MgSO4 alone. Here, we assessed spinal extracellular Mg2+ and glutamate levels following SCI in rats using microdialysis. Basal levels of extracellular Mg2+ (∼0.5 mM) were significantly reduced to 0.15 mM in the core and 0.12 mM in the rostral peri-lesion area after SCI. A single intravenous infusion of saline or of MgSO4 at 192 μmoL/kg did not significantly change extracellular Mg2+ concentrations. However, a single infusion of AC105 (a MgCl2 in PEG) at an equimolar Mg2+ dose significantly increased the Mg2+ concentration to 0.3 mM (core area) and 0.25 mM (rostral peri-lesion area). Moreover, multiple AC105 treatments completely restored the depleted extracellular Mg2+ concentrations after SCI to levels in the uninjured spinal cord. Repeated MgSO4 infusions slightly increased the Mg2+ concentrations while saline infusion had no effect. In addition, AC105 treatment significantly reduced extracellular glutamate levels in the lesion center after SCI. These results indicate that intravenous infusion of PEG-formulated Mg2+ normalized the Mg2+ homeostasis following SCI and reduced potentially neurotoxic glutamate levels, consistent with a neuroprotective mechanism of blocking excitotoxicity.
Introduction
M
As in other tissues, Mg2+ homeostasis is critical to normal central nervous system (CNS) function. Whole tissue Mg2+ depletion was observed following injury to spinal cord in rats. 4 Therapeutic effects of Mg2+ supplementation have been tested in preclinical and clinical studies in stroke 5 –7 and traumatic brain injury, 8 –11 and both positive and negative results were reported. Interestingly, the neuroprotective effects of Mg2+ administration were shown in experimental spinal cord injury. 12 –15 However, these effects were achieved only with extremely high doses of MgSO4 (e.g., 600 mg/kg) far exceeding the doses used clinically for patients with preeclampsia or cardiac arrest. Given the unmet need for therapies for acute spinal cord injury and other CNS injury indications and the inherent neuroprotective activity of Mg2+, it is reasonable to consider the development of a Mg2+ formulation that enhances delivery of Mg2+ to the injured CNS.
Polyethylene glycol (PEG) is a common excipient in pharmacologic formulations. It was reported that lower dose of Mg2+, when formulated with PEG, could also provide neuroprotection after spinal cord injury (SCI) in rats. 16,17 AC105 comprises MgCl2 in a PEG3350 formulation. AC105 treatment resulted in reduction of lesion volumes and improvement of neurologic outcomes following SCI in rats, 17 but not in pigs. 18 Although Mg2+ infusion has been shown to be neuroprotective in some animal models, little is known about the distribution of Mg2+ to the CNS following systemic delivery.
In this study, microdialysis was employed to assess whether intravenous AC105 administration enhances delivery of Mg2+ to the extracellular compartment of the compression-injured rat spinal cord, compared with saline or an equimolar MgSO4 infusion. Since glutamate is excitotoxic following CNS injury, we also measured changes in extracellular glutamate concentrations after SCI following vehicle or Mg2+ infusion as MgSO4 or AC105. Although MgSO4 is more commonly used than MgCl2 clinically, the latter might have some advantages in safety. 19 Thus, MgCl2 was selected for formulation with PEG. Our results indicate extracellular Mg2+ concentrations were significantly increased in the rats treated with AC105 over those animals treated with saline or an equimolar Mg2+ infusion alone. In addition, AC105 treatment was found to be superior to saline or MgSO4 administration in blocking glutamate release as measured by extracellular glutamate concentrations following SCI. These results suggest that AC105 provides for enhanced delivery of Mg2+ to injured spinal cord tissue over standard Mg2+ salt infusion alone and inhibits glutamate release after compression injury in spinal cords.
Methods
Microdialysis probe preparation and evaluation
Microdialysis probes were either purchased (Part No. CMA8010631, CMA 31 Linear Microdialysis Probes, Molecular Weight Cut-off 55kD; Harvard Apparatus, Newark, NJ) or custom-made with a regenerated cellulose hollow fiber (Part No. 132295, Molecular Weight Cut-off 18kD; Spectrum Laboratories, Inc., Rancho Dominguez, CA) connected to polyimide tubing (Part No. 141-0001; Vention Medical Co., Salem, NH). With the exception of the central 2 mm of the membrane, exposed microdialysis membranes were coated with a thin layer of epoxy, leaving only the uncoated portion remaining for functional dialytic sampling.
The following chemicals were used in this study: NaCl, NaHCO3, MgCl2 and glucose (J.T. Baker, Philipsburg, NJ), and KCl, KH2PO4, CaCl2, MgCl2, sodium L-glutamate, taurine, GABA, glycine, L-aspartic acid, and L-asparagine (Sigma-Aldrich, St. Louis, MO). AC105 (a sterile formulation of MgCl2 [Part No.: MGCL13/PEG319] and PEG3350 [Part No.: PEG319]) was produced by AAI Pharma Services, Wilmington, NC.
All probes were evaluated for their ability to recover Mg2+ prior to using in in vivo experiments. To do so, artificial cerebrospinal fluid (CSF) solutions (133.0 mM NaCl, 25.6 mM NaHCO3, 4.9 mM KCl, 1.2 mM KH2PO4, 1.9 mM CaCl2, and 10.0 mM glucose) were prepared, containing 0.1, 0.3, 1.0, and 3.0 mM of MgCl2, a concentration range that covers an estimated extracellular level of Mg2+ (0.5–1.2 mM) in spinal cord. 20 The solutions were bubbled with 95% air/5% CO2 for 15 min before use. The probes were incubated in the artificial CSF solutions at 37°C for at least 1 h and then perfused with artificial CSF solution without Mg2+ at a rate of 2 μL/min for 30 min for sampling of standard Mg2+ solutions using an infusion pump (CMA 402 syringe pump, Part No. 8003100; Harvard Apparatus). Mg2+ levels in the solutions collected from the microdialysis probe outlet (dialysates) were assayed using inductively-coupled plasma atomic emission spectrometry (ICP-AES; model Genesis; Spectro, Mahwah, NJ) according to the manufacturer's technical guides and correlated with the known Mg2+ concentrations in the solutions from which they were obtained using a linear regression method.
To evaluate probe glutamate recovery, an additional seven probes were incubated in artificial CSF solutions (133.0 mM NaCl, 25.6 mM NaHCO3, 4.9 mM KCl, 1.2 mM KH2PO4, 1.9 mM CaCl2, 10.0 mM glucose, and 1 mM MgCl2) at 37°C for at least 1 h. These solutions also contained four concentrations of compounds of interest: 1, 3, 10, and 30 μM of sodium L-glutamate, taurine and GABA; 0.67, 2, 6.67, and 20 μM of glycine; 0.33, 1, 3.33, and 10 μM of L-aspartic acid; and 0.1, 0.3, 1 and 3 μM of L-asparagine. The probes were perfused with the above CSF solution without glutamate and the other compounds of interest at a rate of 2 μL/min for 30 min for sampling of standard solutions. The levels of glutamate in the dialysates were measured using ultra performance liquid chromatography (UPLC) with fluorometric detection (model H-Class system; Waters Corporation, Milford, MA) according to the manufacturer's technical applications guide; and their correlation to the corresponding known glutamate concentrations in the solutions from which they were obtained were analyzed with a linear regression method.
Effects of PEG on Mg2+ and glutamate recovery
To test whether PEG affected probe Mg2+ recovery, Mg2+ standard solutions with different PEG concentrations were prepared to mimic the PEG distribution in relevant compartments: body fluid (0.62% PEG) and plasma (6.67% PEG), assumed for the distribution in the current in vivo microdialysis experiments. Microdialysis probes were incubated in the analyte solutions (the artificial CSF containing 1 mM Mg2+ with different PEG3350 concentrations [0%, 0.62%, 3.65% and 6.67%]) at 37°C for at least 1 h and then perfused with an artificial CSF solution lacking Mg2+ at a rate of 2 μL/min for 30 min using an infusion pump. The Mg2+ concentrations in the dialysates and the standard solutions were measured using ICP-AES for comparison of probe Mg2+ recoveries (Mg2+ in dialysates as a percentage of Mg2+ in the standard solutions).
To determine whether PEG interfered with probe glutamate recovery, microdialysis probes were incubated in artificial CSF solutions (standards) containing 3 μM glutamate with or without 0.62% PEG at 37°C for at least 1 h and then perfused with an artificial CSF solution lacking glutamate at a rate of 2 μL/min for 30 min using an infusion pump. The glutamate levels in the dialysates and the standards were assessed with UPLC. The probe glutamate recoveries (glutamate in dialysates as a percentage of glutamate in the standard solutions) were compared in the standard solutions with or without PEG.
Induction of rat spinal cord injury
All animal procedures were approved by the Institutional Animal Care and Use Committee of Acorda Therapeutics Inc. and conducted in accordance with the Guide for the Care and Use of Laboratory Animals (Version 2011), the Animal Welfare Act, and the 2013 American Veterinary Medical Association Guidelines for the Euthanasia of Animals.
Spinal cord injury was induced in female Long-Evans rats according to a method described previously. 21 Briefly, rats (200–250 g) obtained from the Charles River Laboratories (Raleigh, NC) were acclimated to facility conditions for at least 5 days prior to surgery. Cefazolin (50 mg/kg, subcutaneously) and acetaminophen (100 mg/kg, po) were administered preoperatively. Anesthesia was induced and maintained with isoflurane (1–3% to effect) delivered in medical grade oxygen and the skin over the thoracic spine was shaved and sanitized with povidone iodine solution. Each rat was then placed on a heating pad with temperature control setting at 37°C. Rectal temperature, heart rates, and peripheral capillary oxygen saturation (SpO2) were continuously monitored during the surgical procedure. A laminectomy was performed in T9/T10 spinal vertebrae to expose the spinal cord. Two flat blades (width of 4 mm) of a modified cover-slip forceps were inserted into the T9/T10 spinal canal between the lateral aspects of the spinal cord and vertebrae and then compressed laterally for 15 sec to induce a compressing injury in the spinal cord. The forceps blade gap distance was 0.9 mm, confirmed using a micrometer prior to each surgery. Sham-operated rats underwent all the same procedures without spinal cord compression. The surgeon was blinded to the identity of the treatment group.
Microdialysis procedures
Immediately or at 24 h after SCI induction, two microdialysis probes were inserted transversely through the spinal cord, one positioned at the center of the compression site (core area) and the other 3 mm rostral to the compression center (rostral peri-lesion area; Fig. 1). Under a surgical microscope, the probe alignment was carefully adjusted so that only the functional (uncoated) part (2 mm length) was placed within the spinal cord (∼3 mm diameter). After installation, the probes were continuously perfused with artificial CSF lacking Mg2+ and amino acids at a flow rate of 2 μL/min using an infusion pump. Dialysates were collected every 15 or 20 min for 3 h (not including prior 1 h washout) into a 250 μL polyethylene microcentrifuge tube. Once the dialysate sample was collected, a portion (20 μL) was transferred into another microcentrifuge tube for the determination of Mg2+ concentrations by ICP-AES. The remaining dialysate tube was quickly frozen on dry ice and stored at −80°C for subsequent amino acid analysis using UPLC analysis.
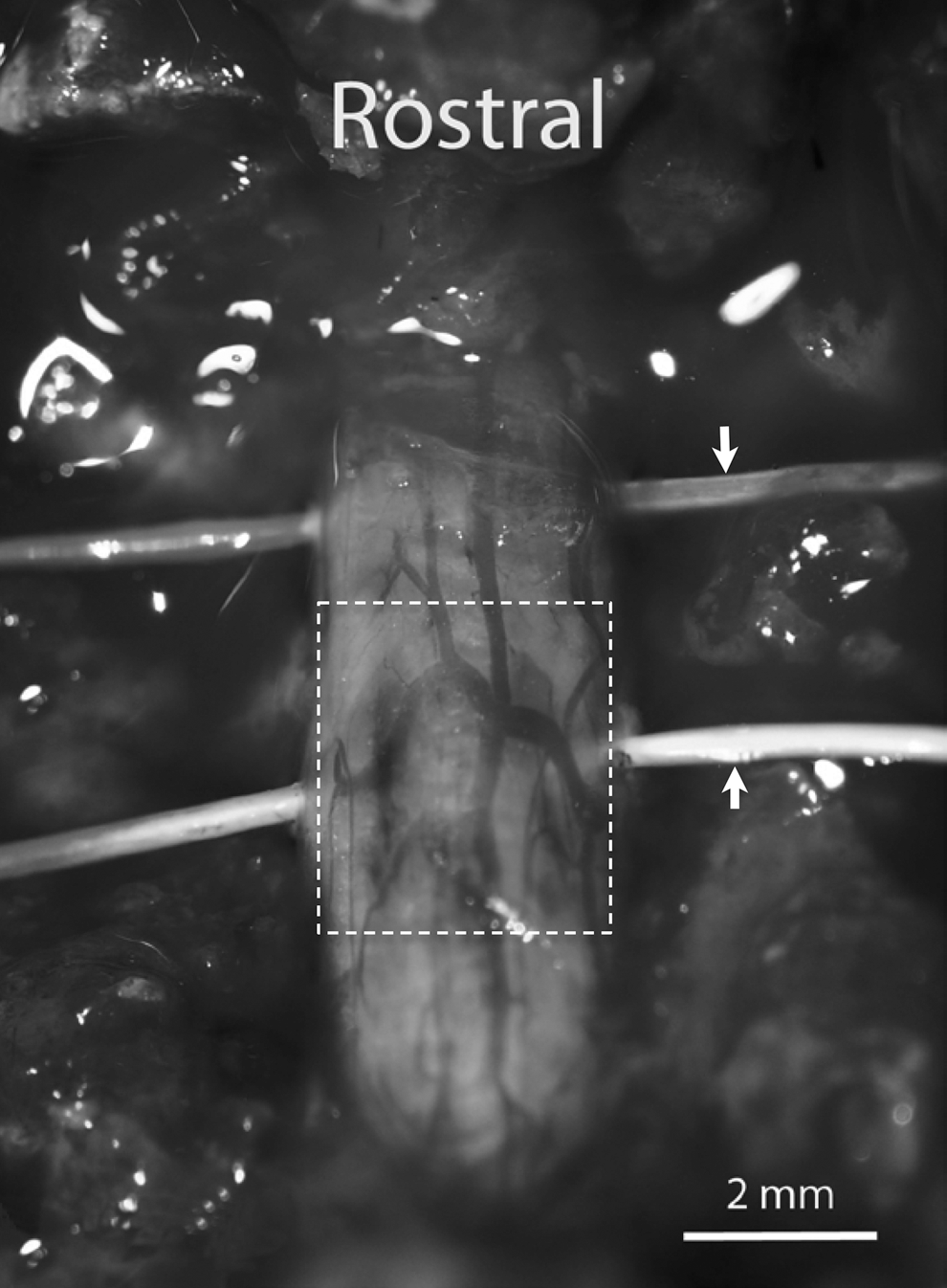
Placement of microdialysis probes. Two microdialysis probes (indicated by arrows, outer diameter 0.26 mm) were transversely implanted in a rat spinal cord after compression injury (outlined with a dotted line); one probe was placed in center of the injury (core area) while the another probe was placed 3 mm rostral to the injury center (peri-lesion zone).
Intravenous drug infusion
The left jugular vein in the rat was catheterized under anesthesia (see above). The catheter (Part No. RJVR-10; SAI Infusion Technologies, Lake Villa, IL) was then connected to a syringe for drug administration with an infusion pump. For the infusions in awake rats, the catheter was subcutaneously tunneled and exteriorized through a scapular incision, and then connected to a small port in a soft harness (Part No.: VAH95AB; Instech Laboratories, Inc., Plymouth Meeting, PA) installed around the animal shoulders. After anesthesia termination, the awake rat was housed in a specially designed cage (Part No.: MTANK; Instech Laboratories, Inc., Plymouth Meeting, PA) where the animal was able to move freely with access to drinking water and food.
Two infusion paradigms were investigated. The first evaluated the effects of a single treatment of AC105 on extracellular Mg2+ changes, while the second examined the effects of multiple successive treatments of AC105 on extracellular Mg2+ and glutamate changes within a 28 h period after SCI. Rats were assigned randomly by an investigator other than the surgeon to receive AC105 (Mg2+ dose of 192 μmoL/kg), MgSO4 (Mg2+ dose of 192 μmoL/kg) or an equivalent amount (5 mL/kg) of sterile normal saline solution. Each treatment group had 12–15 rats. The Mg2+ dose and frequency were selected based on previous studies showing improvement of treatment with PEG/Mg2+ formulation in functional recovery after SCI in rats. 16,17 The solution was intravenously infused over 30 min with a programmable syringe infusion pump (Part No.: 703007; Harvard Apparatus). The first infusion was initiated at 2 h after SCI. For the single treatment paradigm, the solution was administered once only, while for the multiple treatments paradigm, the solution was dosed five times, 6 h apart (Fig. 2).
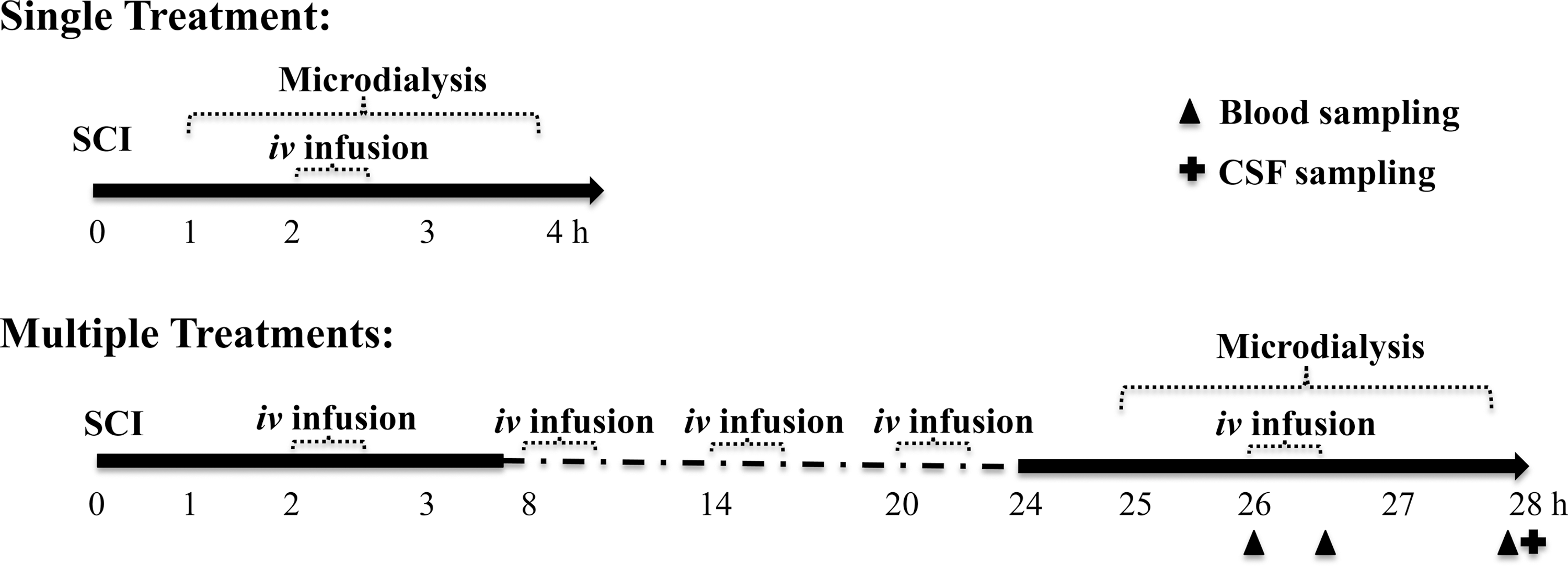
Schematic of the study designs. Experiment 1: single treatment. Microdialysis sampling began 1 h after spinal cord injury (SCI), while intravenous (i.v.) infusion of saline, AC105 or MgSO4 was initiated at 2 h after SCI induction. Experiment 2: multiple treatments. Intravenous infusion was initiated at 2 h after SCI (as in the single treatment phase) and was repeated four times at 6-h intervals. Microdialysis was conducted starting at 25 h after SCI, at a time where four infusions had been completed. The fifth (last) dose was given at 26 h after SCI.
Blood and CSF collection
For the rats administered a series of five saline, MgSO4, or AC105 treatments, blood samples (∼200 μL) were collected into a polyethylene microcentrifuge tube without any anticoagulant immediately before the fifth (the last) drug infusion, at end of the infusion, and at 2 h after the infusion (Fig. 2). A replacement volume of normal saline was administered intravenously after each blood collection to prevent changes in blood volume during sampling. The blood samples were kept at room temperature for 30 min and then centrifuged at 2500 rpm for 20 min at 4°C for serum collection. Terminal CSF samples (∼100 μL) were collected from the cisterna magna at 2 h after the last infusion. Serum and CSF samples were stored at 4°C until Mg2+ concentrations were determined by ICP-AES.
Calculations and statistical analysis
Concentrations of extracellular Mg2+ (mM) in spinal cord tissue were calculated from corresponding concentrations in dialysates, adjusted by previous in vitro calibration. Mg2+ changes in dialysates after drug treatment were expressed as a percentage relative to their pre-treatment baseline levels at each time-point, while the baseline levels were derived from the average of four (for the single treatment setting) or three (for the multiple treatment setting) samples collected during 1 h immediately prior to the drug infusion. The total exposure of extracellular Mg2+ following infusions also was calculated as areas under the curve (AUCs) during the initial 120 min after drug administration.
The extracellular glutamate concentrations were measured and peak exposure, as well as AUCs of the dialysate glutamate over 120 min after start of the last drug or saline i.v. infusion, were obtained. The changes in glutamate also were expressed as percentage relative to its individual pre-treatment baseline, while the baseline was derived from the average of three samples collected during 1 h immediately prior to the drug infusion.
Group mean and standard errors of the mean were calculated for each group. Group means were compared using one-way analysis of variance (ANOVA) followed by Dunnett's or Tukey's multiple comparison tests, or whenever two variables were involved, with two-way ANOVA followed with Bonferroni post-test. Values of p less than 0.05 were considered statistically significant.
Results
Performance of microdialysis probes
Microdialysis was used to sample Mg2+ and glutamate in the extracellular compartment in the rat spinal cord. Prior to their application in live animals, the ability of all microdialysis probes to recover Mg2+ was tested in artificial CSF solutions with different Mg2+ concentrations (0.1–3.0 mM). The results revealed a linear relationship of the probe Mg2+ recovery as a function of Mg2+ concentrations in the dialyzed standard solutions (Fig. 3). In another set of calibration studies, a linear relationship between recovered glutamate levels in dialysates versus glutamate concentrations from dialyzed solutions was observed (Fig. 3). These data show that the probes linearly sampled both Mg2+ and glutamate from solutions external to the membrane across a physiologic/pathophysiologic concentration range.
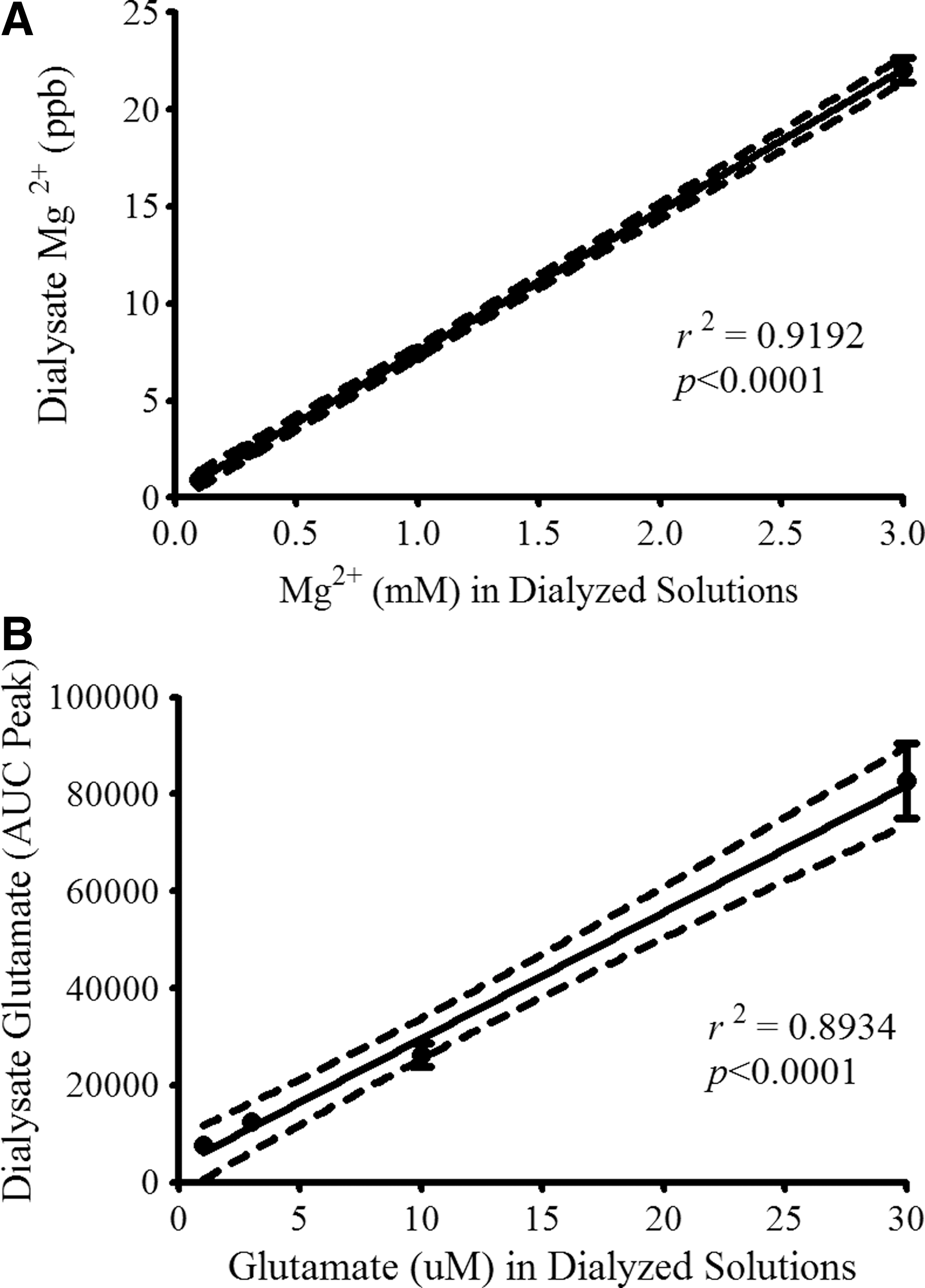
Evaluation of magnesium (Mg2+) and glutamate recovery by microdialysis probes. Prior to their application in in vivo experiments, microdialysis probes (n = 91) were tested in artificial cerebrospinal fluid (CSF) solutions containing Mg2+ at concentrations of 0.1, 0.3, 1.0, and 3.0 mM
Effects of PEG on microdialysis probe recoveries of Mg2+ and glutamate
To address the possibility that the presence of PEG external to the probe membrane could impact Mg2+ and glutamate recoveries, in vitro studies were conducted to compare the recoveries of these analytes by microdialysis probes immersed in solutions containing the same concentrations of Mg2+ and glutamate with or without PEG. The probe Mg2+ recoveries were 8.25 ± 0.65%, 8.10 ± 0.41%, 8.55 ± 0.38%, and 8.64 ± 0.34%, dialyzed in artificial CSF solutions containing 0%, 0.62%, 3.65%, and 6.67% PEG, respectively (n = 16 for each; p > 0.05 by one-way ANOVA). The probe glutamate recoveries were 9.2 ± 1.1% and 10.6 ± 1.1%, dialyzed in artificial CSF solutions containing 0% and 0.62% PEG, respectively (n = 7 for each; p > 0.05 by Student's t-test). No statistically significant difference in probe Mg2+ and glutamate recoveries was observed among the probes tested in the solutions with and without PEG, demonstrating that no PEG-probe interactions contributed to Mg2+ and glutamate recoveries during microdialysis.
Rat physiological parameters during microdialysis procedures
No statistically significant difference in physiological parameters (rectal temperature, heart rate and peripheral capillary oxygen saturation) was detected among the groups of rats with or without SCI before and during the microdialysis procedures (Table 1), suggesting that the changes of Mg2+ and glutamate in extracellular components observed in the current study (see below) were not accounted for by differences in physiological parameters during microdialysis procedures.
Physiological parameters during microdialysis procedures in control (laminectomy only) and SCI rats treated with saline, AC105, or MgSO4. Data are expressed as group mean ± standard error of the mean. n = 12–14 per group.
“0 min” denotes the time to initiate the intravenous infusion during the microdialysis procedures.
Lax., laminectomy; SCI, spinal cord injury; RT, rectal temperature; HR, heart rate; SpO2, peripheral capillary oxygen saturation.
Decreased extracellular Mg2+ levels after SCI
In order to detect alterations of extracellular Mg2+ concentrations, microdialysis probes were implanted transversely in the rat spinal cord at level of T9/10, as shown in Figure 1. A basal level of ∼0.5 mM in extracellular Mg2+ was observed in rats at 25 h after laminectomy without SCI, consistent with the findings (0.5–1.2 mM) reported previously. 20, 22 SCI at 1 h resulted in a significant decline of extracellular Mg2+ concentrations by 69% and 74% in the core and the rostral peri-lesion areas, respectively, compared with the levels of the control animals (laminectomy without SCI at 25 h). Extracellular Mg2+ concentrations also were significantly decreased (∼50%), compared with post-SCI time-matched (25 h) laminectomy controls. These results strongly reflect a disruption of extracellular Mg2+ homeostatic mechanisms following SCI (Fig. 4). Although no laminectomy samples were taken at 1 h, extracellular Mg2+ concentrations within injured spinal cords sampled at 1 h and 25 h, while being less than half those of laminectomy controls, did not significantly differ, suggesting that Mg2+ levels do not spontaneously recover in 24 h.
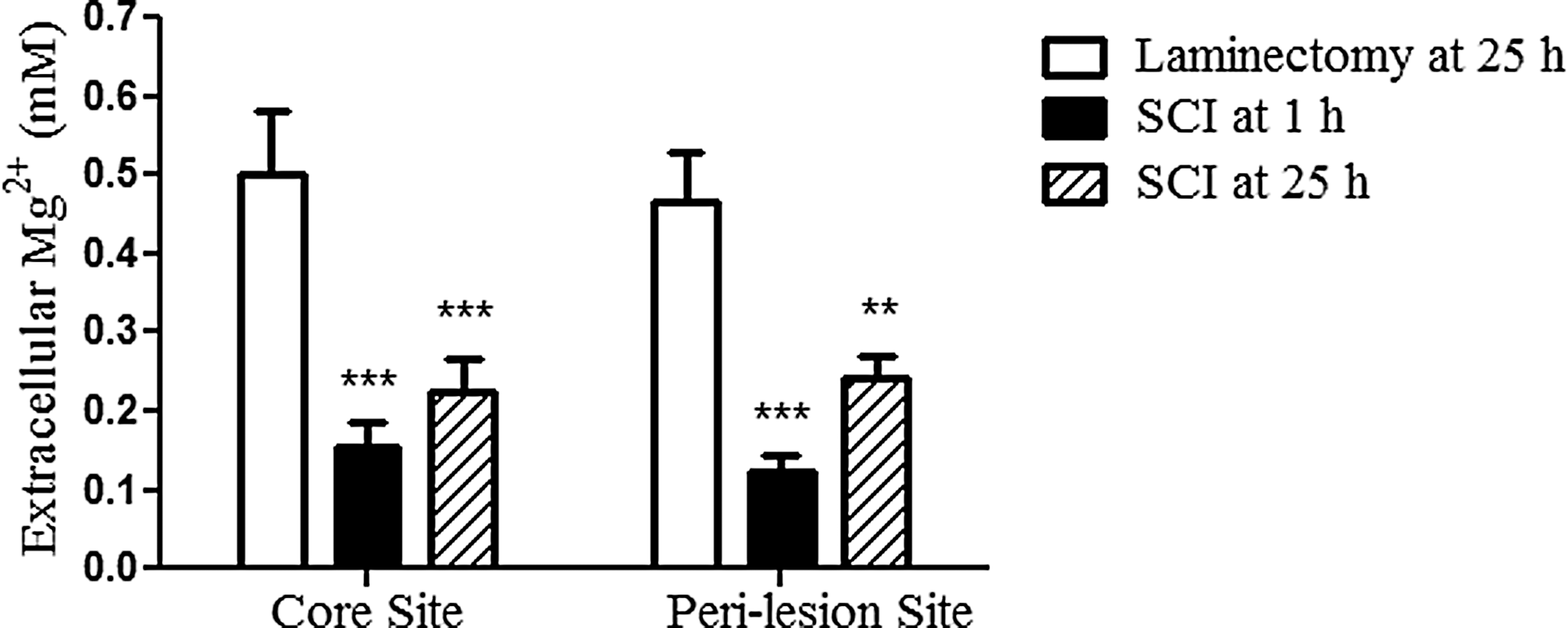
Spinal cord injury (SCI) decreases extracellular magnesium (Mg2+) concentrations. Extracellular Mg2+ levels were measured at the injury core and rostral peri-lesion zone in the spinal cord at 1 h (n = 15) and at 25 h (n = 12) after SCI or at the corresponding areas at 25 h after laminectomy without injury (n = 11). Rats from groups dialyzed at 25 h after laminectomy or SCI were intravenously infused 2 h after SCI or laminectomy with normal saline (5 mL/kg) during four 30 min intervals separated by 6 h. Mg2+ concentration data are expressed as group mean ± standard error of the mean. Statistically significant differences between the SCI group and the Laminectomy group means were detected using two-way analysis of variance followed by Bonferroni post-test. Asterisks ** and *** denote p < 0.01 and 0.001, respectively, vs. the Laminectomy group. No statistically significant difference was detected in mean Mg2+ concentrations between the SCI groups at 1 h vs. 25 h.
Effect of a single treatment with AC105 and MgSO4 on extracellular Mg2+ level
We then examined whether systemic infusion of Mg2+ as MgSO4 or AC105 could restore extracellular Mg2+ levels toward normal. A single i.v. infusion of AC105 (Mg2+ 192 μmoL/kg) significantly increased the extracellular Mg2+ levels from 0.15 and 0.12 mM to 0.30 and 0.25 mM in the core and the rostral peri-lesion areas, respectively. The effect was maximal at 45 min after the infusion and sustained at least through 2 h. However, no significant changes in extracellular Mg2+ levels were observed after infusion of an equimolar MgSO4 or saline solution (Fig. 5). Areas under the concentration-time curve (AUC, an index of extracellular Mg2+ exposure throughout a defined time period) during 120 min after infusion also indicated that a single treatment of AC105 resulted in a significant increase of Mg2+ exposure, compared with saline treatment, while Mg2+ exposure was not significantly changed by MgSO4 treatment (Table 2). When the changes in extracellular Mg2+ levels were expressed as a percentage relative to pre-treatment baseline, the AC105 group produced a 1.7-fold increase in extracellular Mg2+ levels at 45 min after the drug infusion, compared with its pre-treatment level (p < 0.001) in the core area, while MgSO4 and saline treatment did not produce any significant alterations in extracellular Mg2+ levels after the treatments. Similar trends were observed in the rostral peri-lesion area (Fig. 5). These results indicated that intravenous infusion of AC105 significantly enhanced delivery of Mg2+ to the extracellular space of injured spinal cord tissue, compared with equimolar Mg2+ (MgSO4) or saline infusion.
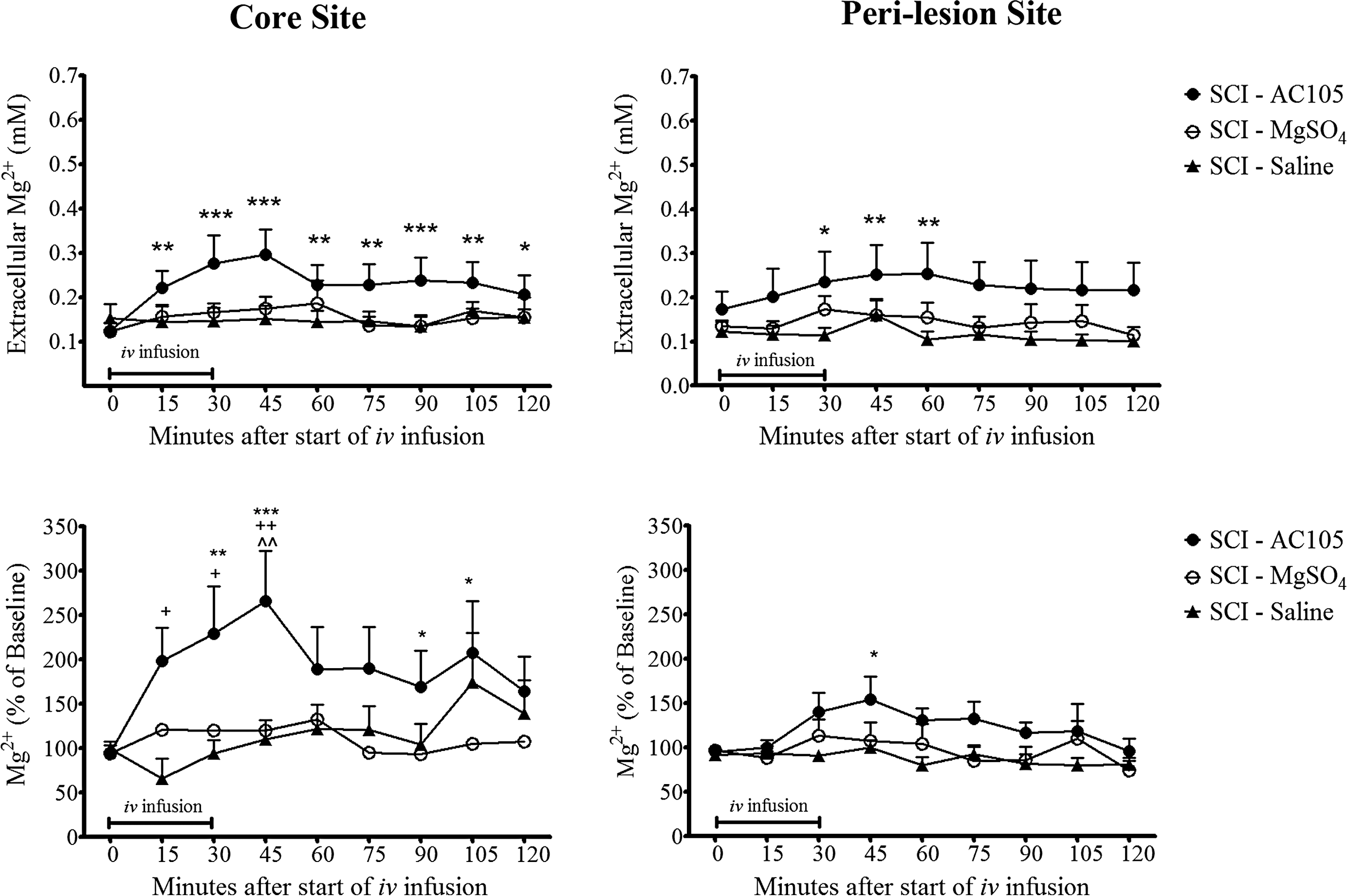
Effects of a single intravenous (i.v.) infusion with AC105 (magnesium [Mg2+] 192 μmoL/kg), MgSO4 (Mg2+ 192 μmoL/kg), or an equivalent volume of saline solution (5 mL/kg) on extracellular Mg2+ levels in rats after spinal cord injury. Changes of extracellular Mg2+ were expressed as concentrations of mM (top panel), and also calculated as percentage relative to the pre-treatment baseline (bottom panel). AC105 treatment resulted in a statistically significant increase in extracellular Mg2+ levels, while no significant increase over baseline was observed with saline or MgSO4 treatment. Asterisks *, **, and *** denote p < 0.05, 0.01, and 0.001 vs. the pre-treatment level at time “0” within the group, respectively, analyzed with repeated one-way analysis of variance (ANOVA) followed by Dunnett's multiple comparison test. Symbols +, ++, and ^^ indicate p < 0.05 and 0.01 vs. the spinal cord injury (SCI)-Saline group, and p < 0.01 vs. the SCI-MgSO4 group, respectively, analyzed using two-way ANOVA followed by Bonferroni post-test. Data are expressed as group mean ± standard error of the mean. n = 14–15 in each group.
Rostral peri-lesion site
Laminectomy-Saline
, **Values of p less than 0.05, 0.01, respectively, compared with SCI-Saline group. Data are expressed as group mean ± standard error of the mean. n = 10–14 per group.
Comparison of extracellular Mg2+ exposure (AUC) and peak levels following a single or multiple intravenous infusions of AC105 (Mg2 + 192 μmoL/kg), MgSO4 (Mg2 + 192 μmoL/kg), or an equivalent amount of saline solution (5 mL/kg) in SCI or laminectomy rats. Mg2+ exposure was expressed as areas under the concentration-time curve (AUC from the time 0 to 120 min after intravenous [i.v.] infusion) or peak level (maximum Mg2+ concentration over 120 min after i.v. infusion). The effect of treatment on differences in AUC was found statistically significant in single treatment (p < 0.05) and in multiple treatments (p < 0.0001) by two-way analysis of variance (ANOVA). The effect of treatment on differences in peak was found statistically significant in multiple treatments (p < 0.0001), but not significant in single treatment, by two-way ANOVA analysis followed with Bonferroni post hoc test.
AUC, area under the curve; SCI, spinal cord injury.
Effect of multiple treatments with AC105 and MgSO4 on extracellular Mg2+ level
We further tested the effects of repeated infusions of Mg2+ on extracellular Mg2+ concentrations in injured spinal cord. Animals were dosed five times, 6 h apart, initiated at 2 h after SCI. Microdialysis procedures were conducted at 25 h after SCI induction, at a time after four treatments had been completed, as described above (Fig. 2).
Control rats (saline-treated laminectomy group) had an extracellular Mg2+ concentration of ∼0.5 mM as assessed by microdialysis, which was about twice that of saline-treated SCI rats. Repeated AC105 treatment significantly increased the extracellular Mg2+ in the center of the lesion to the levels found in laminectomy group (0.5 mM), an effect that was sustained for at least 90 min after the cessation of the last infusion. Repeated MgSO4 treatment (equimolar Mg2+) produced a small but not statistically significant (vs. SCI-saline) increase in extracellular Mg2+ concentration during the 120 min monitoring period, an effect significantly less than that produced by AC105. Similar trends were observed at the rostral peri-lesion area (Fig. 6). The extracellular Mg2+ exposure level expressed in terms of AUC and peak dialysate concentrations during 120 min after initiation of infusion also indicated that multiple treatments of AC105, but not MgSO4, significantly increased the extracellular Mg2+ level, compared with the saline treatment (Table 2). Collectively, these results suggest that multiple infusions of AC105 significantly increased extracellular Mg2+ level in spinal cord after injury. Of note, the extracellular Mg2+ concentration after multiple AC105 infusions approached the levels found in saline-treated laminectomized rats without SCI (Fig. 6; Table 2).
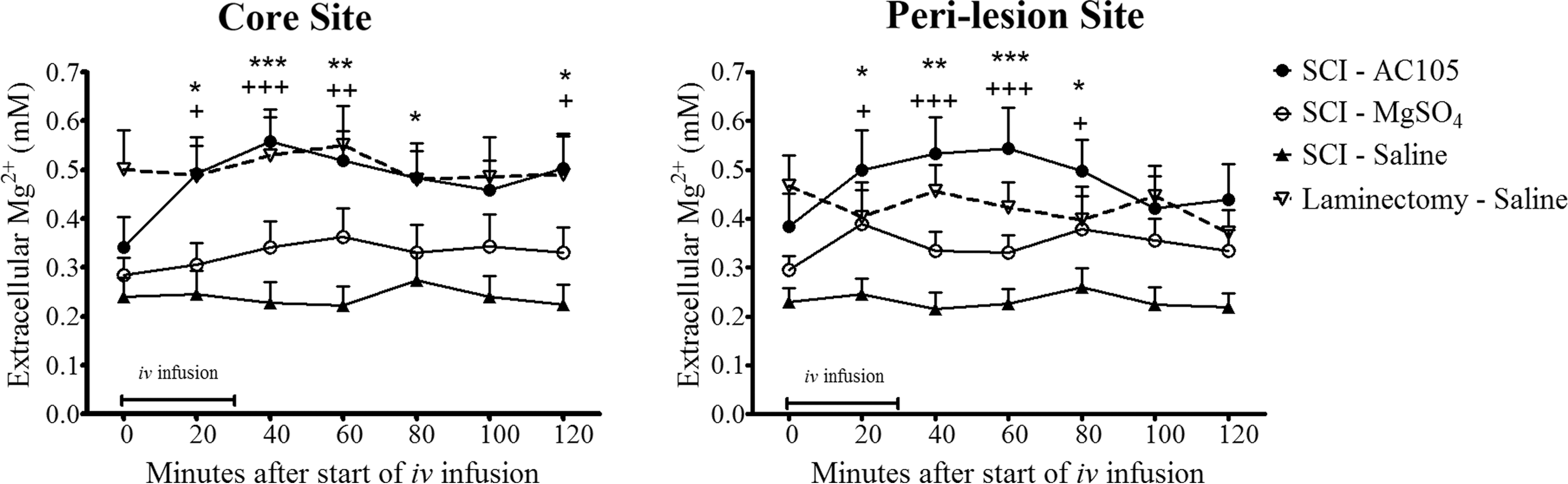
Effects of multiple intravenous (i.v.) infusions with AC105 (magnesium [Mg2+] 192 μmoL/kg), MgSO4 (Mg2+ 192 μmoL/kg), or an equivalent amount of saline solution (5 mL/kg) on extracellular Mg2+ levels in rats after spinal cord injury (SCI). The i.v. infusion was initiated at 2 h after the injury and repeated four times (total of five doses, 6 h apart). The “0” in the X axis indicates 26 h after SCI, at a time four i.v. infusions had been completed. AC105 treatment restored extracellular Mg2+ to levels similar to those measured in the rats with laminectomy alone, but MgSO4 did not. Statistical significances of differences among groups and within a group were detected using two-way analysis of variance (ANOVA) followed by Bonferroni post-test and repeated one-way ANOVA followed by Dunnett's multiple comparison test, respectively. Markers of *, **, and *** denote p < 0.05, 0.01, and 0.001 vs. the pre-treatment level at time “0” within the group, respectively, while +, ++, +++indicate p < 0.05, 0.01, and 0.001 vs. the SCI-Saline group, respectively. Data are expressed as group mean ± standard error of the mean. n = 12–14 in each group.
Serum and CSF Mg2+ concentrations
No significant difference in serum Mg2+ concentrations was detected among the various treatment groups prior to intravenous infusions. However, intravenous infusions of AC105 and MgSO4 resulted in approximately 45% increases (p < 0.001) in serum Mg2+ concentrations for both AC105 and MgSO4 treatment groups at end of the 30 min infusion, verifying that the two groups received equivalent Mg2+ doses. Ninety minutes after the completion of the infusion, serum Mg2+ levels in AC105-treated rats still were significantly higher than those in saline-treated rats, while the Mg2+ levels in MgSO4-treated rats were not significantly elevated, compared with saline-treated rats (Fig. 7). Of interest, no significant difference (p > 0.05 by one-way ANOVA followed by Tukey's multiple comparison test) in CSF Mg2+ concentrations was observed among the groups of laminectomy rats treated with saline (0.81 ± 0.03 mM; n = 10), SCI rats treated with saline (0.85 ± 0.02 mM; n = 11), SCI rats treated with AC105 (1.08 ± 0.13 mM; n = 12), or SCI rats treated with MgSO4 (1.04 ± 0.11 mM; n = 13).
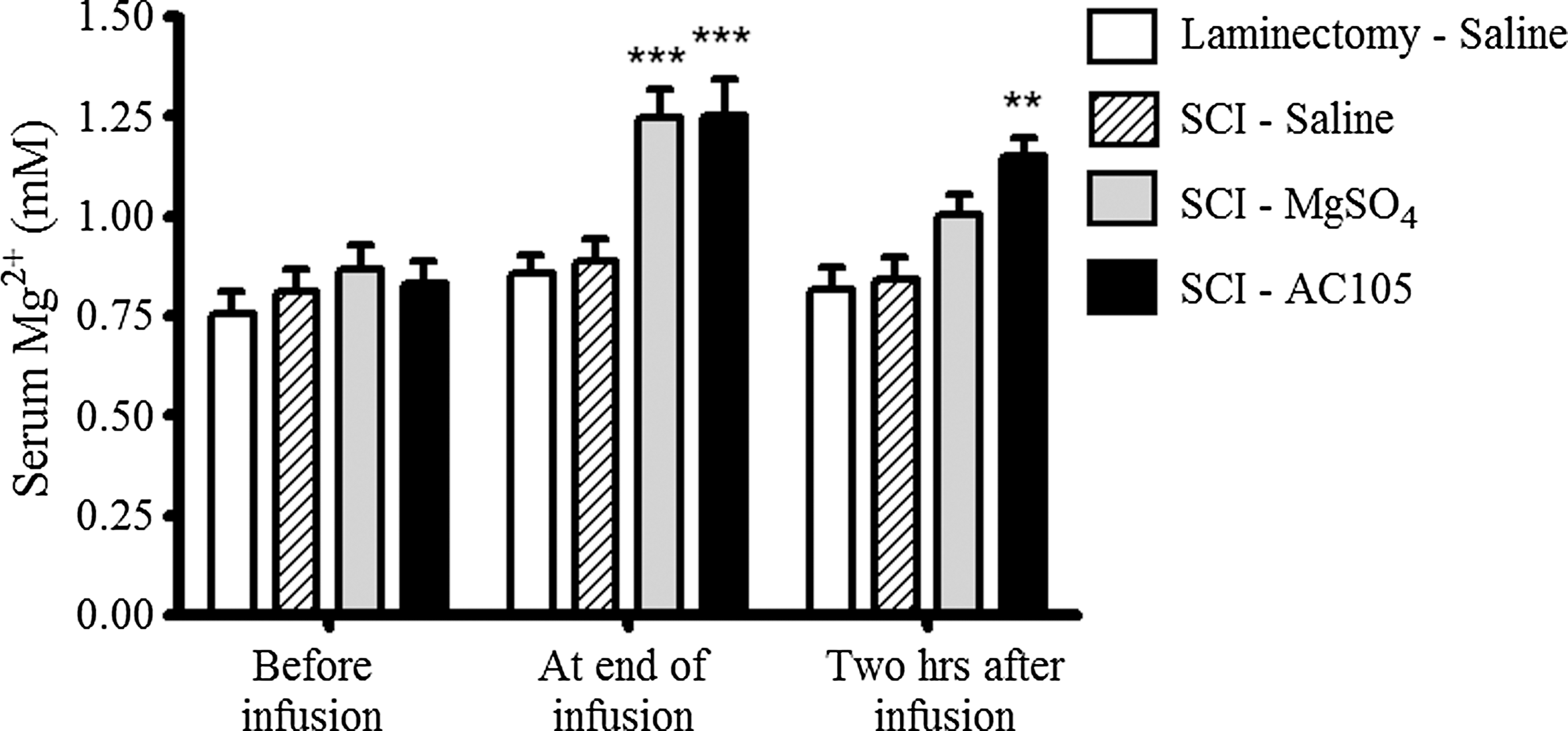
Magnesium (Mg2+) concentrations in serum. AC105 (Mg2+ 192 μmoL/kg), MgSO4 (Mg2+ 192 μmoL/kg), or an equivalent amount of saline solution (5 mL/kg) was intravenously infused over 30 min five times at 6-h intervals. Blood samples were collected immediately before, at end of, and 2 h after the last infusion. Administration of AC105 and MgSO4 resulted in a higher blood level of Mg2+, compared with saline treatment, at end of intravenous (i.v.) infusion. Two-way analysis of variance followed by Bonferroni post-test was performed to detect statistical difference among groups. Markers of ** and *** denote p < 0.01 and p < 0.001, respectively, compared with the Laminectomy-Saline group and the spinal cord injury (SCI)-Saline group. Data are expressed as group mean ± standard error of the mean. n = 14–15 in each group.
Effect of treatments with AC105 and MgSO4 on extracellular glutamate exposure
There was a nearly 20% increase in extracellular glutamate levels in dialysates of spinal cords from saline-treated SCI rats versus saline-treated control rats (sham-operated, laminectomy). This increase was not statistically significant, but was observed in both central and peri-lesion sites. Of note, the elevation in extracellular glutamate level after SCI was abolished by treatment of AC105, but not by administration of an equimolar MgSO4 (Fig. 8). When the glutamate changes were expressed as percentage relative to pre-treatment baseline level, treatment of AC105 elicited a gradual decrease in extracellular glutamate concentrations in the injury core area while infusion of MgSO4 and saline did not. The effect of AC105 treatment on glutamate concentrations vs baseline levels over time was statistically significant, analyzed with two-way ANOVA. At 120 min after infusion, AC105 treatment showed a significant inhibition of glutamate release, compared with the saline treatment after SCI in core area, but not in rostral peri-lesion area (Fig. 8). All together, these data indicate that AC105 reduced extracellular glutamate exposure in parallel with increasing of Mg2+ tissue delivery after SCI.

Extracellular glutamate exposures after multiple treatments of AC105 and MgSO4 in rats. AC105 (Mg2+ 192 μmoL/kg), MgSO4 (Mg2+ 192 μmoL/kg), or an equivalent amount of saline solution (5 mL/kg) was intravenously infused over 30 min for five times with 6 h of interval, initiated at 2 h after spinal cord injury (SCI). The extracellular glutamate levels were measured with ultra performance liquid chromatography and expressed as peak concentration (top panel) and area under the glutamate concentration-time curve (middle panel) over 120 min after start of the last intravenous (i.v.) infusion. The changes in glutamate also were calculated as percentage relative to its individual pre-treatment baseline (bottom panel). Statistical significances of differences among groups were detected using one-way analysis of variance (ANOVA) followed by Tukey's multiple comparison test (top and middle panels) and two-way ANOVA followed by Bonferroni post-test (bottom panel). Overall treatment effect was found statistically significant in comparison of AC105 treatment on glutamate concentrations vs. baseline levels over time in core area after SCI. Marker of * denotes p < 0.05, compared with SCI-Saline group. Data are expressed as group mean ± standard error of the mean. n = 9–14 in each group.
Discussion
Mg2+ plays a critical role in maintenance of a variety of cellular functions. Indeed, Mg2+ is involved in almost every major metabolic and biochemical process within the cells. Mg2+ binds to the nucleotides and stabilizes the natural RNA and DNA structure. 23 Also, Mg2+ is a cofactor or activator for more than 600 enzymes that are essential for life 24 and is involved in regulating intracellular signaling by acting as a physiological calcium antagonist within cells. 25 Moreover, Mg2+ is a key factor in the control of cell proliferation. 26
In the CNS, Mg2+ plays a vital role in neuronal function and abnormalities in Mg2+ homeostasis contribute to CNS dysfunction.
1
–3, 27
Therefore, supplementation of Mg2+ has been considered a therapy for multiple CNS disorders, particularly those involving injury to the central nervous system. Indeed, neuroprotective effects by Mg2+ administration have been reported in experimental models of stroke,
5, 28
–30
traumatic brain injury
8
–11
and spinal cord injury.
12,31,32
It is notable that extremely high doses of Mg2+ (
Treatment with Mg2+ formulated in PEG resulted in smaller lesion size and greater locomotor recovery in a rat thoracic spinal cord injury model, compared with the control of treatment with saline. 16,17 Of note, the Mg2+ dose used in the PEG formulation study is comparable with that used clinically for patients with preeclampsia and cardiac arrest, and is much lower than the doses required to achieve neuroprotection in other studies when Mg2+ was used alone. 12 –14,31 These results suggest that PEG might facilitate delivery of Mg2+ to the injured CNS areas.
The effect of PEG on the delivery of Mg2+ to the extracellular space has not been explored previously. In the current study, we used microdialysis technique to sample the interstitial environment of the normal and injured rat spinal cord for the purpose of measuring extracellular Mg2+ concentrations. SCI produced a significant decline of extracellular Mg2+ level relative to laminectomy control animals, confirming that Mg2+ homeostasis is disrupted following the spinal cord injury. Our data are consistent with a previous report showing declined brain Mg2+ following CNS injury. 33 The reduction in extracellular Mg2+ levels following injury was partially ameliorated by a single treatment of AC105. However, this effect was absent in the groups treated with saline or MgSO4 alone, suggesting PEG, when formulated with Mg2+, enhanced delivery of Mg2+ to the extracellular space of injured spinal cord tissue, compared with equimolar Mg2+ (MgSO4) or saline infusion. With a regimen of multiple administrations, AC105 completely and significantly restored the extracellular Mg2+ concentration after SCI to the levels found in the rats without injury, while saline did not. Multiple infusions of equimolar MgSO4 produced only a small increase in extracellular Mg2+ concentration.
Consistent with a previous report showing sustained glutamate elevation after CNS injury, 34 increased extracellular glutamate levels were observed following SCI in the current study. In parallel to enhancing Mg2+ delivery to the injured tissues, AC105 treatment produced a significant decrease in extracellular glutamate concentration after SCI, observed at 120 min after intravenous infusions. High levels of extracellular glutamate can lead to excitotoxicity in the CNS. When the CNS is injured, glutamate is released from neurons, activates N-methyl D-aspartate (NMDA) receptors, and promotes intracellular calcium overload leading to excitotoxic cell death. 35 Mg2+ is known to inhibit the actions of glutamate at the level of the NMDA receptor, 36 and its role in reducing NMDA-mediated excitotoxicity has been extensively investigated. As a calcium antagonist, Mg2+ blocks glutamate release, 37 inhibits NMDA receptors, 36, 38 –41 and antagonizes the effects of intracellular calcium overload. 42, 43 In addition, Mg2+ modulates vasodilatation and thereby enhances cerebral blood flow. 44 Depletion of extracellular Mg2+ after CNS injury, as seen in the current study, is likely to render neurons vulnerable to NMDA-mediated glutamate excitotoxicity. Thus, blockade of glutamate is one of major mechanisms by which Mg2+ protects CNS from injury.
There are numerous modes for Mg2+ to protect cells of the CNS from injury in addition to inhibition of glutamate-NMDA receptor pathway. For example, Mg2+ inhibits the production of reactive oxygen species 45 and lipid peroxidation. 46 In experimental SCI studies, Mg2+ supplementation was reported to decrease damage to vascular membranes and to preserve the blood–brain barrier, 13,14 normalize energy balance, 47 protect against axonal damage, 12 and reduce apoptosis. 48
In the current study, AC105 increased delivery of extracellular Mg2+ to and reduced extracellular glutamate exposure within the spinal cord after injury. This suggests that the increased delivery of extracellular Mg2+ by PEG formulation ameliorates the effects of glutamate/NMDA receptor-mediated excitotoxicity after CNS injury. This may explain, in part, why supplementation with Mg2+ formulated by PEG showed a tendency to be superior to equimolar doses of Mg2+ as MgSO4 in enhancing behavioral recoveries following rat spinal cord injury in a previous study. 16
The mechanism by which PEG promotes Mg2+ delivery to the injured tissues is not yet clear. Different from other cations like Na+, K+, and even Ca2+, Mg2+ has two hydration shells in a dissolved state, making its hydrated radius about 400 times larger than its dehydrated radius. Therefore, Mg2+ needs to be dehydrated, an energy-consuming process, before passing through channels and transporters. 3 It is not clear yet whether PEG mediates Mg2+ dehydration so as to facilitate transport of Mg2+. On the other hand, PEG was found to more robustly affect Mg2+ concentrations in the lesion core than in the peri-lesion area (Fig. 5). This may indicate that increased vessel permeability after injury may be involved into the process of PEG-mediated Mg2+ delivery. Of interest, we found that the serum Mg2+ concentration at 2 h after infusion was significantly higher in the AC105 group, but not in MgSO4 group, than that in saline group, suggesting that AC105 involves a PEG-Mg2+ complex that reduces systemic Mg2+ clearance (Fig. 7).
In the present study, extracellular Mg2+ levels were measured after infusion of MgCl2/PEG for 120 min only. It appears warranted to continue assessing Mg2+ levels beyond this time-point given an interval of 6 h between infusions in the repeated regimen. These data could help optimize the treatment frequency to maintain Mg2+ homeostasis after SCI. Another limitation of the current study is a lack of simultaneous measurement of CSF concentrations of Mg2+ as opposed to tissue extracellular levels, as comparison of the two assessments may provide relevant information to clinical setting. However, it is not likely that a significant elevation in Mg2+ concentrations would be observed in CSF since CSF samples taken 2 h after infusion of AC105 exhibited no appreciable Mg2+ increase despite a significant and persistent elevation in extracellular Mg2+ concentrations at 2 h, compared with vehicle treatment.
In summary, we report here that extracellular Mg2+ concentrations are depleted after spinal cord injury in rats. Intravenous administration of MgCl2 in a PEG formulation is superior to equimolar MgSO4 infusion alone in raising extracellular Mg2+ concentrations in the injured spinal cord. Multiple infusions of AC105 virtually restore extracellular Mg2+ to normal levels within a 28-h period following SCI. In addition, AC105 treatment significantly decreased extracellular glutamate exposure over time, which may reduce glutamate-NMDA mediated excitotoxicity following injury. Our data demonstrate the importance of formulation refinement and improvement in the development of Mg2+ as a potential therapeutic neuroprotective agent in SCI and other CNS injury.
Footnotes
Acknowledgments
We would like to thank the staff of the Acorda Animal Modeling Facility, including Messrs. Ronald Mertz, Donald Stevenson, and Jaime Muniz, and Ms. Cindy Rodriguez, for their expert animal care and technical support. We would also like to thank Drs. Andrew Blight and Andrew Eisen for their critical review and advice on this manuscript.
Author Disclosure Statement
All authors are employees and stockholders of Acorda Therapeutics, Inc.