
Abstract
Although traumatic brain injury (TBI) is now considered a chronic disease, few studies have investigated the long-term behavioral deficits elicited by a well-established rodent model of injury. Here we evaluate behavioral measures, commonly used in TBI research, to determine which tests are useful for studying long-term effects of brain injury in rats. Male Sprague-Dawley rats were handled and pre-trained to neurological, balance, and motor coordination tests prior to receiving parasagittal fluid-percussion injury (FPI), sham injury, or maintenance as naïve cohorts. Rats underwent neuroscore, beam-balance, and beam-walk tests for 3 days after injury. Subsequently, in separate groups at 3, 6, or 12 months, they were re-tested on the same tasks followed by a working memory version of the Morris water maze. On post-injury days (PIDs) 1–3, significant effects of injury on neuroscore, beam-balance, and beam-walk were observed. Differences in the beam-walk task were not detectable at any of the later time-points. However, deficits persisted in beam-balance out to 3 months and neuroscore out to 6 months. Working memory deficits persisted out to 12 months, at which time a reference memory deficit was also evident. These data suggest that balance and motor coordination recovered more quickly than neurological deficits. Furthermore, while deficits in working memory remained stable over the 12-month period, the late onset of the reference memory deficit points to the progressive nature of the injury, or an age/TBI interaction. In conclusion, standard behavioral tests are useful measures of persistent behavioral deficits after parasagittal FPI and provide evidence that TBI is a chronic condition that can change over time and worsen with age.
Introduction
T
Thus, it is imperative to fully understand the long-term behavioral deficits produced by TBI. To do so, it is necessary to thoroughly investigate experimental models of TBI in terms of their usefulness for studying TBI as a progressive, chronic disease. Here, we demonstrate that the parasagital fluid-percussion injury (FPI) model in rats is valuable for the study of persistent behavioral deficits out to one year after injury. Several rodent models of TBI have been developed and are well-established in the scientific literature, including (1) controlled cortical impact injury (CCI),
11
(2) closed head injury (CHI) using such devices as accelerated impactors and weight drop devices,
12
(3) several types of blast TBI,
13,14
and FPI.
15,16
The FPI model remains one of the most popular. Research in FPI using dogs as experimental subjects was first published in the 1950s.
17
Since that time, more than 1,000 articles have been published on research that employed FPI in rats, making it the most studied rodent model of TBI (
All of these models have contributed extensively to our current body of knowledge regarding TBI. However, few reports have investigated outcomes longer than 2 months post-injury and even fewer rodent models of TBI have been evaluated for behavioral deficits that persist or progress over a one-year period (reviewed by Gold and associates in 2013 and Osier and colleagues in 2015). 19,20 In rats, we found three studies that evaluated behavior out to one year after TBI. In 1998 Lindner and co-workers compared a frontal with a lateral cortical contusion injury, using a variety of neurological and behavioral tests, out to one year after injury. 21 They found that cognitive deficits were more robust and enduring than sensorimotor deficits for both injury sites; however, the frontal site produced greater deficits in tests of attention and orientation than the lateral site. In 1999, Dixon and colleagues found deficits in spatial memory performance at one year after controlled cortical impact injury. 22 Pierce and associates in 1998 also reported enduring cognitive and neurobehavioral changes out to one year after a severe lateral FPI. 23
Arguably the most important outcome measure of a TBI model is behavior, as this is the best way to test meaningful brain function. However, the vast majority of FPI experiments with behavioral studies measure outcomes at relatively short time-points after injury, typically from one to 14 days after injury. Thus, we designed this study to behaviorally characterize the chronic effects of the parasagittal FPI model of TBI in rats and to assess whether the behavioral tests we use are sensitive to the chronic effects of injury. We used male Sprague-Dawley rats that were acclimated to handling, pre-trained, and pre-assessed prior to parasagittal FPI, sham injury, or maintenance as an injury-naïve cohort. Rats were evaluated using a short neurological assessment (neuroscore), a balance task (beam-balance), a motor coordination task (beam-walk), and a working memory task (working memory Morris water maze). All rats were assessed on the neuroscore, beam-balance, and beam-walk on Day 0 (prior to surgical preparation or equivalent for naïve animals) and Days 1 through 3 after injury. Subsequently, rats were re-assessed on these tasks in separate groups at 3, 6, and 12 months after injury and prior to the working memory test. Our overall goal was to quantify the degree of deficit and rate of recovery of reflexes, balance, motor coordination, learning, and memory for one year after injury in order to realize a well-defined model of chronic behavioral deficits that will be useful for future testing of treatments for chronic effects of TBI.
Methods
Animals
Adult male Sprague-Dawley rats (300–350 g) were obtained from Harlan (Houston, TX), housed two per cage with food and water ad libitum, and maintained at a constant temperature (21o–23°C) and humidity (45–50%) with lights on 07:00–19:00. Behavioral measures were conducted during the light part of the light/dark cycle. All experiments were conducted in accordance with the National Institutes of Health's Guide for the Care and Use of Laboratory Animals (National Research Council, 2011) and approved by the Institutional Animal Care and Use Committee of the University of Texas Medical Branch, Galveston, Texas.
Surgical preparation
Rats were anesthetized with 4% isoflurane in an anesthetic chamber for 5 min, intubated, and mechanically ventilated with 1.5–2% isoflurane in oxygen:air (50:50) using a volume ventilator (EDCO Scientific, Chapel Hill, NC) set at a tidal volume of 12–15 mL/kg and 50 breaths/min. Rectal and temporalis muscle temperatures were monitored using a telethermometer (Yellow Springs Instruments, Yellow Springs, OH) and maintained between 36.5 and 37.0°C prior to the injury. Rectal temperature was maintained using a thermostatically controlled water blanket (Gaymar, Orchard Park, NY). Body temperatures were maintained between 37.0 and 37.5°C for the duration of anesthesia.
Rats were prepared for parasagittal fluid-percussion TBI as previously described. 15,24 Briefly, rats were placed in a stereotaxic frame and the scalp was sagittally incised. A 4.0 mm diameter hole was trephined into the skull 2.0 mm right of the sagittal suture and midway between lambda and bregma. A modified Luer-Lok syringe hub (Becton-Dickinson, Franklin Lakes, NJ) was placed over the exposed and intact dura, bonded in place with cyanoacrylic adhesive, and covered with dental acrylic. Isoflurane was discontinued. Rats were connected to the trauma device and paw pinch reflexes were tested repeatedly (once per sec) until a withdrawal response was detected, at which point the pendulum was released and the fluid wave was propagated forward, subjecting the brain to a 2.0 atm fluid pulse.
Immediately after delivery of the fluid pulse, rats were removed from the trauma device, placed in a supine position, and monitored until they recovered the righting reflex. The time to recover the righting reflex was recorded. Rats were then replaced on isoflurane anesthesia for removal of the syringe hub and wound closure. Rats were then allowed to recover in a warm, humidified incubator until fully alert and returned to their home cage with the same cage mate. Cage mates were maintained throughout the duration of the experiment.
Experimental design
See Figure 1 for schematic. Rats (n = 131) received moderate parasagittal FPI, sham injury, or were prepared as naïve cohorts. All rats were pre-trained to the neurological scoring, beam-balance, and beam-walking tests one day prior to surgery (Day −1) and assessed for baseline performance on the day of surgery (prior to surgery) or experimental day zero (Day 0). All rats were assessed three times after surgery on post-injury days (PIDs) 1–3 (or the equivalent for naïve rats), then were housed in pairs with the same cage mate and handled twice weekly until the appropriate time-point (3, 6, or 12 months after surgery). The final number of rats in the three groups was: NAÏVE (n = 40), SHAM (n = 39), TBI (n = 37). The number of rats at the three survival times was: 3 months (n = 37), 6 months (n = 38), 12 months (n = 41). Starting 10 days prior to the assigned tissue collection time-point, rats were re-assessed on the neuroscore, beam-balance, and beam-walk (3 months = PIDs 81–83; 6 months = PIDs 171–173; 12 months = PIDs 351–353) rested for 2 days, and then subjected to the working memory version of the Morris water maze for 5 days.

Experimental design work flow. With a goal of 12 rats per group at each time-point, we started with 131 rats that received
Behavioral assessment
All behavioral measures were conducted by a blinded observer.
Neurological function, gross vestibulomotor function, and fine motor coordination were assessed on PIDs 1–3 using a short neurological assessment (neuroscore; modified from Schallert 25 ), the beam-balance task, and the beam-walk task. 26 –28 Reference and working memory were assessed using the working memory version of the Morris water maze. 27,29
Neuroscore
The following reflex tests were administered in order and repeated three times. A normal response received a score of 0, whereas an abnormal response received a score of 1 for each trial of each test for a total possible score of (7 × 3 = 21), the higher the score, the greater the deficit.
1. Forelimb Flexion Test (0–1)
2. Hind Limb Flexion Test (0–1)
3. Visually Triggered Placing Test (0–1)
4. Contact Triggered Placing Test, right (0–1)
5. Contact Triggered Placing Test, left (0–1)
6. Hind Paw Grasping Reflex Test, right (0–1)
7. Hind Paw Grasping Reflex Test, left (0–1)
Beam-balance
The balance beam apparatus consisted of a beam 60 cm in length, 1.75 cm in width, 4.0 cm in height, elevated 90 cm off the floor, with a barrier 30 cm in height, 30 cm in width. The beam was secured to a table and the barrier was attached to the beam so that 50 cm of the beam protruded from the barrier, away from the table, over a cushioned safety box. 15,26,30
Rats were trained 24 h before surgery, and one pre-assessment was made on the day of surgery prior to surgery, or the equivalent, in the case of rats in the naïve group. For training, each rat was placed on the balance beam for a 60 sec trial. The rat was removed from the beam for a brief moment between trials. If the rat failed to balance on its own, it was allowed to fall into the safety box. Timing began when the rat was securely positioned on the beam. Rats were rated using an ordinal scale with (1) being normal balance and (6) being unable to remain on the beam: 1. Balances with steady posture (grooms, climbs barrier) 2. Balances with unsteady posture (grasps sides of beam and/or has shaky movements) 3. Hugs the beam or slips or spins on the beam 4. Attempts to balance, but falls off after 10 sec 5. Drapes over or hangs from the beam, falls off in less than 10 sec 6. Falls off, making no attempt to balance or hang onto the beam
Beam-walk
The beam-walk apparatus consisted of a beam 100 cm in length, 2.5 cm in width, and 4.0 cm in height with four equally spaced pegs (2 cm in height, 25 cm apart) and a black goal box (28 cm in length, 18 cm in height, 18 cm in width) at one end. 15,26,30 The target end of the beam was attached to the goal box that rested on a table. The beam was elevated 1 m off the floor, and a bright light and white noise source were positioned near the starting end. Each rat was trained 24 h before surgery. First the rat was placed in the goal box for 60 sec, after which the rat was removed from the goal box, the light and white noise were turned on, and the rat was placed on the beam at the location of the peg hole closest to the goal box and allowed to walk to the goal box. As soon as the rat's front feet crossed the threshold of the goal box, the light and noise source were turned off. The animal was allowed to rest in the goal box for 30 sec between each trial. This procedure was repeated at each peg location and from the starting position. Smaller pegs (22 cm) were then inserted, and one beam walk was completed, then the longer pegs (38 cm) were inserted. Trials were repeated with the longer pegs until rats completed the task in 10 sec or less (usually approximately three trials). On the day of injury (Day 0), rats underwent a pre-assessment consisting of three timed trials. Three trial assessments were performed on PIDs 1–3.
Rats were retested on the neuroscore, beam-balance, and beam-walk tests the week prior to the Morris water maze working memory test, which was scheduled such that their last day of testing would correspond with the tissue collection date of 3, 6, or 12 months post-injury, sham injury, or the corresponding days for the naïve rats that were housed with the injured and sham-injured rats.
Working memory Morris water maze
In this test, rats were placed in a 1.8 m diameter tank filled with water to a height of 28 cm, 2 cm higher than a hidden platform, which was 10 cm in diameter and 26 cm in height 26,27,29 (Fig. 2). Rats received four pairs of trials for 5 consecutive days. Rats were assigned four starting points (N, S, E, or W) and four platform locations (1, 2, 3, or 4) in a balanced order to avoid starting points too close to platforms. For Trial 1: Rats were placed in the tank facing the wall at the assigned location and allowed 120 sec to find the platform. If the rat failed to find the platform, it was led to the platform by the experimenter's hand; either way, rats were allowed 15 sec to remain on the platform. For Trial 2: Rats were immediately placed back in the same starting position and again allowed 120 sec to find the platform. After Trial 2, the rats rested for 4 min in a heated enclosure followed by the second pair of trials and repeated until four pairs of trials were completed. All rats experienced the same sequence of start points and platform locations, which were randomly selected at the beginning of the experiment. Injured (TBI), sham-injured (SHAM), and age-matched naïve rats that received the same amount of handling (NAÏVE) were tested at 3, 6, and 12 months after surgical preparation, or the equivalent time period for NAÏVE.
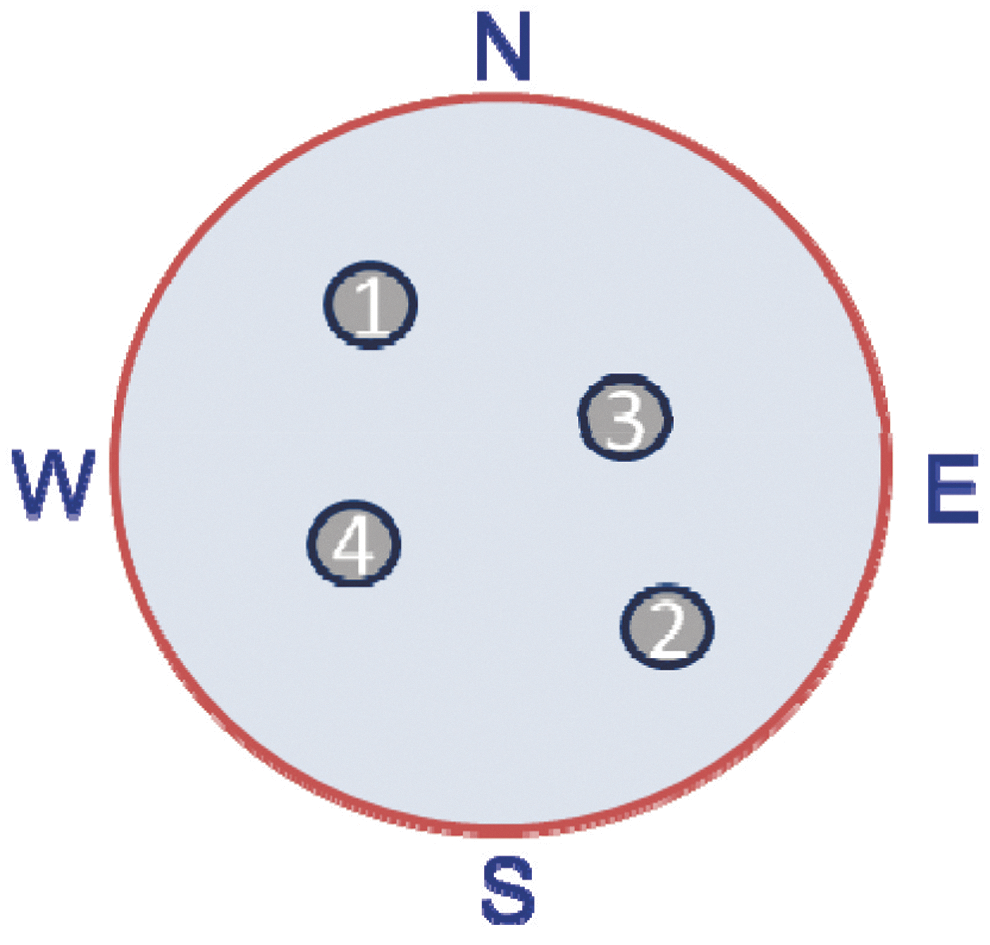
Diagram of platform locations used for the working memory version of the Morris water maze. Rats were given two trials using the same starting point at each platform location with only the 15 sec rest on the platform between trials. After each pair of trials rats were given a 4 min rest in a warming chamber before starting the next pair of trials at a new starting and platform location. Thus each rat experienced four pairs of trials for 5 days in a row.
Statistical analysis
For the neuroscore, beam-balance, and beam-walk assessments, data were expressed as mean ± standard error of the mean (SEM) and were analyzed using a 3 (Injury: NAÏVE, SHAM, TBI) × 4 (Days: 0–3) two-factor analysis of variance (ANOVA) with replication followed by a post hoc homoscedastic two-tailed t test to detect differences between the overall means for each group (NAÏVE, SHAM, TBI).
Working memory water maze results were expressed as the overall mean latency ± SEM for Trial 1 and Trial 2 for each group at each time-point. Analysis consisted of a 3 (Injury: NAÏVE, SHAM, TBI) × 5 (Days: 1–5) two-factor ANOVA with replication followed by a post hoc homoscedastic two-tailed t test to detect differences between the overall means for each group (NAÏVE, SHAM, TBI). Each significant outcome shown is a result of assessing between-groups comparisons separately for Trial 1 and Trial 2 at each time-point (3, 6, 12, months).
Results
A total of 131 rats were employed in this study. Ten rats were excluded because their righting reflex did not meet criterion for this study, 13–20 min; five rats died or were euthanized due to poor health prior to completion of the study. The remaining 116 rats were used in the final analyses. All 116 animals were included in the analyses of behavior on PIDs 0–3, prior to subsequent division into 3, 6, and 12 month groups. Weights are shown in Figure 3.
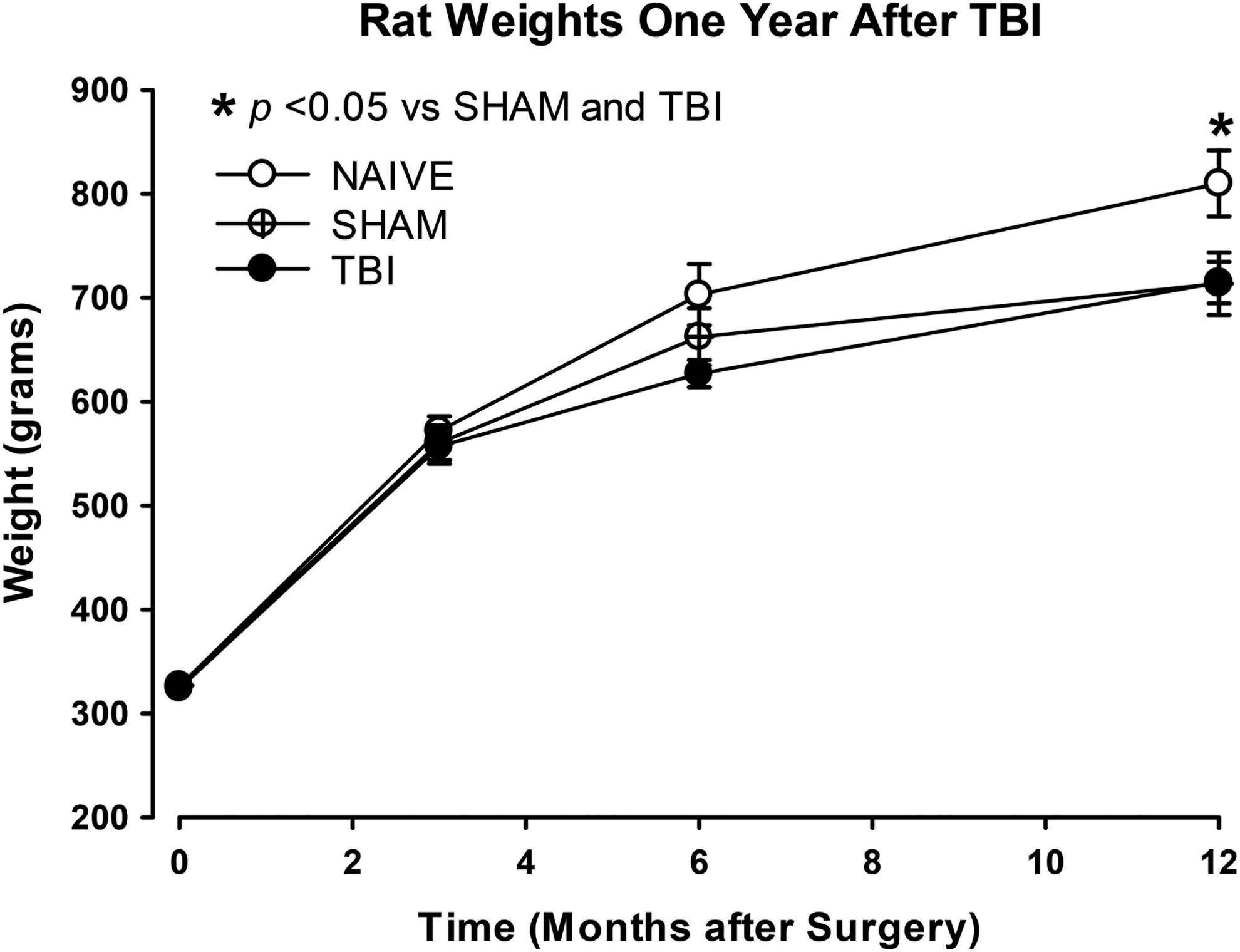
Rat weights from Day 0–12 months. Rats were weighed regularly before and after surgery (Day 0). The Day 0 time-point represents all animals for each group (NAÏVE, n = 40; SHAM, n = 39; TBI, n = 37). Weights are shown at each time-point of assessment: 3 months (NAÏVE, n = 14; SHAM, n = 12; TBI, n = 11), 6 months (NAÏVE, n = 13; SHAM, n = 13; TBI, n = 13), and 12 months (NAÏVE, n = 9; SHAM, n = 13; TBI, n = 14). By 12 months, four rats in the NAÏVE group and one in the TBI group died of “natural causes” or were sacrificed due to poor health. Separate one-way analysis of variances (ANOVAs) were performed at each time-point. There was an overall significant effect of injury status at 12 months (F2,39 = 3.31; p = 0.047). With significant differences between NAÏVE and both SHAM and TBI (*p < 0.05)
Neuroscore
The results of the neuroscore reflex assessment are shown in Figure 4.
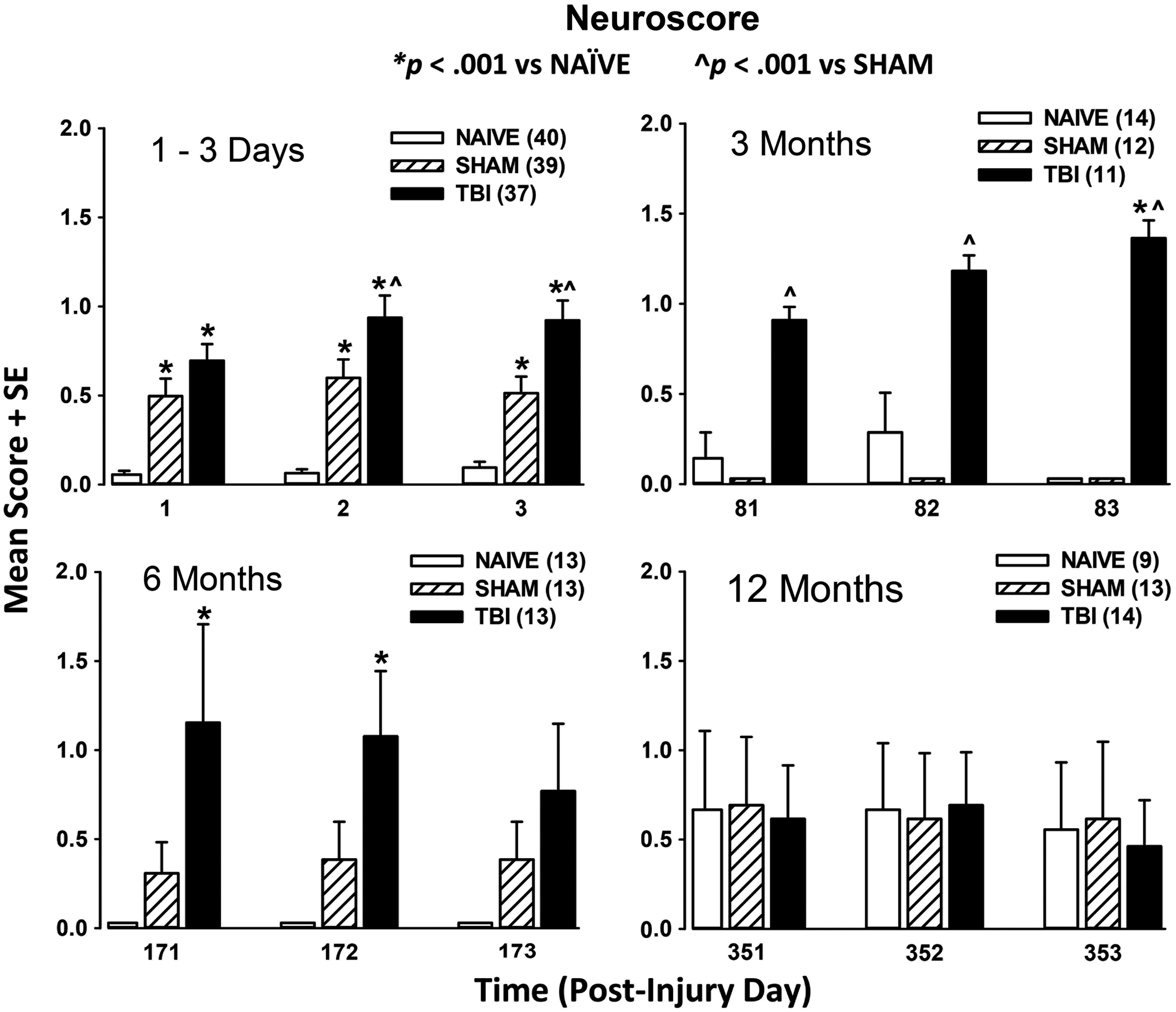
Results of the neuroscore test. All rats were trained to simple reflex testing tasks prior to Day 0 (see text for details on training, testing, and scoring). Results are shown for post-injury days 1–3 (upper left), the 3 month time-point (upper right), the 6 month time-point (lower left), and the 12 month time-point (lower right). The results of the post hoc t test for each time-point are shown on the graphs.
Post-injury days 1–3 (Fig. 4, upper left)
Two-factor ANOVA with repeated measures revealed a significant overall effect of injury (F2,456 = 65.8; p < 0.0001), a significant overall effect of time (F3,456 = 10.4; p < 0.0001), and a significant injury × time interaction (F6,456 = 5.71; p < 0.0001). Post hoc analysis indicated that both injured and sham-injured rats were impaired when compared with naïve rats on the same days. However, the injured rats showed greater deficits when compared with uninjured rats on the second and third day of testing, indicating that the deficits in the injured rats persisted out to 3 days to a greater extent than either of the uninjured groups (SHAM or NAÏVE).
3 month neuroscore (Fig. 4, upper right)
The results for reflex testing at 3 months revealed no significant effect of day (F3,167 = 0.253, n.s.). However, there was a significant effect of injury (F2,167 = 19.51; p < 0.001) and a significant interaction (F6,167 = 3.27; p = 0.005), indicating that deficits in reflexes continue to be detectable 3 months after injury.
6 month neuroscore (Fig. 4, lower left)
The results for reflex testing at 6 months also revealed no significant effect of day (F 3,155 = 0.119, n.s.). However, there was a significant effect of injury (F2,155 = 8.96; p < 0.001) but no interaction (F6,155 = 1.77; p = 0.109), indicating that some deficits in reflexes persist out to 6 months after injury.
12 month neuroscore (Fig. 4, lower right)
The results for reflex testing at 12 months revealed there were no significant effects of day (F 3,156 = 0.461, n.s.), or injury (F2,156 = 0.665, n.s.) and no interaction (F6,167 = 0.428, n.s.). Thus, at 12 months after injury, the neuroscore assessment did not detect an effect of injury on reflexes in this model. These results may be influenced by the effects of aging in that naïve rats also showed minor deficits that were not apparent at the 3 and 6 month time-points.
Beam-balance
The results of the beam balance test are shown in Figure 5.

Results of the beam-balance test. All rats were trained to balance on the beam until they were able to balance for 60 sec for three consecutive trials (see text for details on training, testing, and scoring). On subsequent tests, rats were scored on a scale from 1–6 with 1 signifying normal balance and 6 signifying no effort to stay on the beam. Results are shown for the Day 0 baseline score and post-injury days 1–3 (upper left), the 3 month time-point (upper right), the 6 month time-point (lower left), and the 12 month time-point (lower right). The results of the post hoc t test for each time-point are shown on the graphs.
Post-injury days 0–3 (Fig. 5, upper left)
The results for the beam-balance test were analyzed using a 3 (Injury: NAÏVE, SHAM, TBI) × 4 (Day 0–3) two factor (Injury × Day) ANOVA with repeated measures, followed by a post hoc Student's t test for differences between group means. There was a significant overall effect of injury (F2,456 = 65.8; p < 0.0001), a significant overall effect of time (F3,456 = 10.4; p < 0.0001), and a significant injury × time interaction (F6,456 = 5.71; p < 0.0001), suggesting that both injured and sham-injured rats have balance impairment when compared with naïve rats on the first day after surgery. However, the sham-injured rats recovered to naïve levels by PID 2. Injured rats continued to score significantly worse than sham and naive out to PID 3, as well as compared with themselves prior to injury, indicating that this injury produced deficits in the vestibular system that persisted out to 3 days after injury.
3 month beam-balance (Fig. 5, upper right)
The results for the beam-balance test at 3 months showed a significant effect of day (F 3,167 = 7.89, p < 0.0001), and of injury (F2,167 = 6.30, p = 0.002) but no interaction (F6,167 = 1.65; p = 0.137), suggesting that the balance task becomes more difficult for all rats at 3 months. However, after 2 days of practice, uninjured rats improved performance to a level comparable to their own baseline performance. The performance of injured rats showed no improvement over the 3 day test period.
6 month beam-balance (Fig. 5, lower left)
The results for the beam-balance test at 6 months showed a significant effect of day (F3,144 = 13.2; p < 0.001), but there was no effect of injury (F2,144 = 0.715, n.s.) and no interaction (F6,155 = 0.529, n.s.), indicating that the differences in beam-balance between the baseline scores and the later time-points were not due to injury.
12 month beam-balance (Fig. 5, lower right)
The results for the beam-balance test at 12 months showed a significant effect of day (F 3,156 = 30.23; p < 0.0001), and of injury (F2,156 = 8.49; p < 0.001), but no interaction (F6,167 = 1.11, p = 0.357). These data suggest that whereas the balance task became more difficult for all rats 12 months later (compared with Day 0), after 2 days, naïve rats showed improvement that was not seen in SHAM or TBI rats. This suggests that rats that had a surgical procedure are less resilient compared with naïve rats of the same age. Further, the NAÏVE group weighed significantly more than the other two groups at this time-point, suggesting that weight and age were not a confounding factor for the beam-balance test.
Beam-walk
The results of the beam-walk test are shown in Figure 6.

Results of the beam-walk test. All rats were trained to traverse the beam to reach the escape box and were timed on three consecutive trials for baseline measures (see text for details on training, testing, and timing). Results are shown for the Day 0 baseline latency to traverse the beam and post-injury days 1–3 (upper left), the 3 month time-point (upper right), the 6 month time-point (lower left), and the 12 month time-point (lower right). The results of the post hoc t test for each time-point are shown on the graphs.
Post-injury days 0–3 (Fig. 6, upper left)
The results for the beam-walk task were analyzed using a 3 (Injury: NAÏVE, SHAM, TBI) × 4 (Day 0–3) two factor (Injury × Day) ANOVA with repeated measures followed by a post hoc Student's t test for differences between group means. For PIDs 0–3 there was a significant overall effect of injury (F2,456 = 40.2; p < 0.0001), and time (F3,456 = 9.26; p < 0.001), and a significant injury × time interaction (F6,456 = 4.82; p < 0.0001). Post hoc comparisons between groups revealed the differences shown in Figure 6. Both injured and sham-injured groups were significantly slower at traversing the beam than the corresponding naïve rats on PIDs 1–3. However, injured rats were significantly slower than both naïve and sham-injured rats as well as pre-injury TBI rats, indicating that motor coordination is impaired out to 3 days after injury.
3 month beam-walk (Fig. 6, upper right)
The results for the beam-walk task at 3 months showed a trend toward a significant effect of day (F 3,167 = 2.641; p = 0.051), a significant effect of injury (F2,167 = 4.714; p = 0.010), but no interaction (F6,167 = 1.295; p = 0.262). These results suggest that all groups took longer to traverse the beam when first re-evaluated on PID 81 compared with baseline, possibly due to increased weight, age, or lack of practice. All groups achieved baseline latencies by the third day of testing regardless of injury.
6 month beam-walk (Fig. 6, lower left)
The results for the beam-walk task at 6 months showed a significant effect of day (F3,144 = 13.2; p < 0.001), but no effect of injury (F2,144 = 0.715, n.s.), and no interaction (F6,155 = 0.529, n.s.). Considering that all three groups were significantly slower compared with their baseline measure, and not different from each other, these data suggest that other factors, such as age or increasing weight (an accompaniment of aging in Sprague-Dawley rats) are more significant than injury in altering performance on this task.
12 month beam-walk (Fig. 6, lower right)
The results for the beam-walk task at 12 months showed a significant effect of day (F3,156 = 6.62; p < 0.001), and a significant effect of injury (F2,156 = 5.11; p < 0.001), but no interaction (F6,167 = 0.619, n.s.). Considering that all three groups were significantly slower compared with their baseline measure, these data further support the notion that age and/or weight are of greater significance in reducing performance on this task at 12 months.
Working memory Morris water maze
The results of the water maze test are shown in Figure 7.
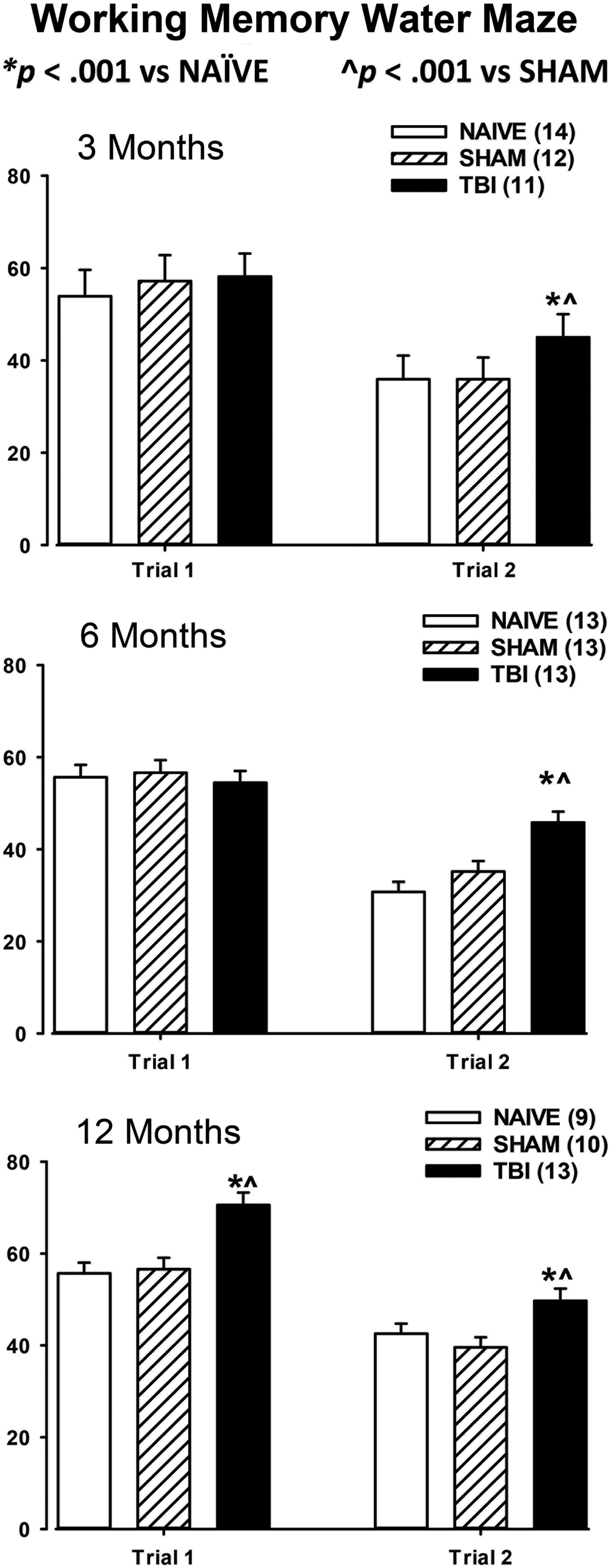
Results of the working memory water maze test. Upon completion of neuroscore, beam-balance, and Beam-walk testing, at the appropriate time after injury rats were tested on a working memory version of the Morris water maze (see text for details). Average cumulative latencies over the 5 day period are presented separately for Trial 1 and Trial 2. Results are shown for the 3 month time-point (upper), the 6 month time-point (middle), and the 12 month time-point (lower). Results of the post hoc t test are shown on the graphs.
3 month water maze (Fig. 7, upper)
Data were analyzed using a 3 (Injury: NAÏVE, SHAM, TBI) × 5 (Days: 1–5) two-factor ANOVA with replication followed by Student's t tests for differences between the means. For Trial 1, there was a significant overall effect of day on latency (F4,839 = 20.47; p < 0.0001), indicating that latencies decreased over time as rats learned the task. However, there was no effect of injury on latency (F2,830 = 0.92, n.s.) and no interaction (F8,839 = 1.15; p > 0.05). For Trial 2 there was a significant effect of day on latency (F4,839 = 39.41; p < 0.0001), a significant effect of injury on latency (F2,839 = 6.74, p = 0.001), and a trend toward an interaction of injury × day, (F8,839 = 1.71; p = 0.094). The effect of day indicates that the rats are learning over the 5 day period. However, the time course for the 3 and 6 month time-points is not presented to focus on the more important differences in latency between groups. The post hoc t test shows significant differences between TBI versus NAÏVE and TBI versus SHAM for Trial 2. These results demonstrate that injured rats took significantly longer than uninjured rats to locate the platform on the second trial, indicating a deficit in working memory.
6 month water maze (Fig. 7, center)
For Trial 1 there was a significant overall effect of day (F 4,765 = 33.13; p < 0.0001), indicating that the rats were learning the task over the 5 day period. There was no effect of injury (F2,765 = 0.037, n.s.), and no interaction (F 8,76 5 = 0.970, n.s.). For Trial 2 there was a significant effect of day on latency (F 4,765 = 43.06; p < 0.0001), a significant effect of injury on latency (F 2, 765 = 15.47; p < 0.0001), but no interaction (F 8, 765 = 0.41, n.s.). These results at 6 months post-injury are similar to the results at 3 months and continue to point to a deficit in working memory in injured rats.
12 month water maze (Fig. 7, lower)
For Trial 1 there was a significant overall effect of day on latency (F4,765 = 20.0; p < 0.0001), again indicating that latencies decreased over time as rats learned the task. There was a significant overall effect of injury on latency (F2,765 = 12.22; p < 0.0001) and a trend toward an interaction (F8,765 = 1.91; p = 0.056), suggesting that injured rats were performing significantly worse in Trial 1. Longer latencies in Trial 1 point to a deficit in reference or long-term memory. For Trial 2 there was a significant effect of day on latency (F4,765 = 47.49; p < 0.0001), a significant effect of injury on latency (F2,765 = 6.15; p < 0.0001), as well as a significant interaction (F8,765 = 2.32; p = 0.019). Taken together, these results suggest that at 12 months after injury, rats are continuing to experience deficits in working memory, as well as demonstrate a first-time deficit in reference (long-term) memory.
The time course for the 12 month working memory water maze test is shown in Figure 8. The left panel shows the time course for Trial 1 for each of the 5 days of testing. Trial 1 is comparable to the classical spatial version of the Morris water maze that tests reference memory. Because normal rats show a decrease in latency to find the platform over the 5 day period on Trial 1 consistently, this indicates a functioning reference memory, an awareness of the possible platform locations retained from previous days of testing. These data are shown to depict the persistent deficit in Trial 1 performance for injured rats at 12 months after injury. The right panel shows the time course for Trial 2 revealing that the deficit in working memory is small, yet persistent and detectable, especially on the second day of testing.
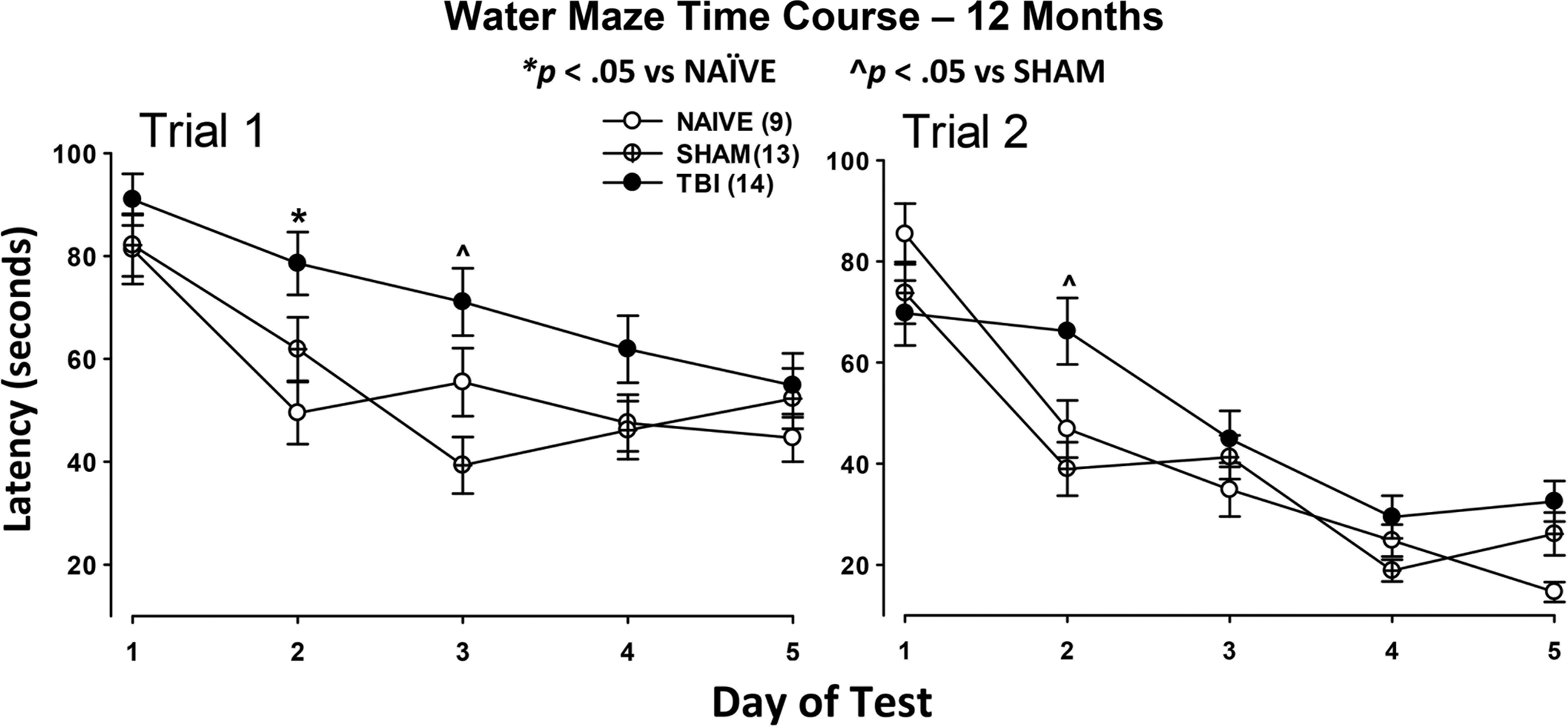
Time course of Trial 1 and Trial 2 for 12 month post-injury groups. The time course for Trial 1 (left) reveals a deficit in reference memory in the TBI group compared with both NAÏVE and SHAM groups as depicted in Figure 7. The post hoc t test detected significant differences on Day 2 between traumatic brain injury (TB) and NAÏVE and on Day 3 between TBI and SHAM. Trial 2 (right) revealed a small deficit in working memory as shown in Figure 7. Post hoc analysis detected a significant difference between TBI and SHAM on Day 2. Differences are depicted on the graphs.
Discussion
These results provide a comprehensive timeline of behavioral deficits in rats after parasagittal FPI using well-established tests to evaluate sensorimotor function, balance, motor coordination, working memory, and reference memory. These tests are valuable tools for evaluating brain function, and have rarely been used at time-points later than 2 months after injury. In humans, a variety of clinical assessments are employed to evaluate patients; these tests are sensitive to deficits even in patients with mild trauma. 31,32 The tests evaluated here, in rodents, also demonstrate sensitivity to injury. Once anesthesia wears off, the injured rats required no intensive care or interventions and looked normal upon casual observation. It is only by probing with specific tests that functional deficits can be observed. Each test employed here detects deficits that result from injury to specific brain areas that reflect related symptoms in humans (Table 1). We utilized the neuroscore to evaluate deficits in reflexes that involve the sensorimotor cortex and thalamus, the beam-balance to measure deficits in the vestibular system, and the beam-walk to detect deficits in the vestibular system, cerebellum, and motor cortex. Additionally, the working memory version of the Morris water maze is sensitive to deficits in the hippocampus and prefrontal cortex. Although other brain areas are likely involved, the areas mentioned here are generally accepted as the primary mediators of these tasks.
To support the translational validity of using experimentally induced brain injury in rats to model chronic effects of TBI in humans, the type of deficit, the brain areas involved, and the test that can be used to measure that deficit in rats are shown, with references.
The neuroscore evaluation was employed early after injury, as well as at later time-points, as repeating this test does not influence the outcome once rats have reached criteria (a score of zero). Deficits in the reflexes tested have been linked to deficits in the sensorimotor cortex and thalamus. These deficits, although significant and consistent up to 6 months after injury, were representative of only one or two small abnormalities, reflective of minor dysfunction. Recovery of injured rats to the same level as age-matched controls (both SHAM and NAÏVE) by 12 months after injury suggests that spontaneous recovery occurred at this level of injury. These deficits indicate a non-debilitating low level of injury. In the forelimb flexion test, first developed to assess deficits in a stroke model, rats with an infarction flexed the contralateral forelimb. 33,34 In rats with a right side FPI (1.9–2.4 atm), Bramlett and colleagues found deficits in contralateral limb placing that recovered during the first week of post-injury testing when tested out to 2 weeks. 35 Deficits in forelimb flexion and placing observed here are likely due to injury of the cortex adjacent to the fluid-percussion location. This area includes the sensorimotor forelimb area. 34
Persistent dysfunction of balance, including dizziness, is often a result of moderate TBI in humans. 36 One way to assess balance, which indicates functioning of the vestibular system, in rats, is using the beam-balance task. 26,37 This task, as with neuroscore testing, is not confounded by repeated testing, so this test is valuable for repeated testing at later time-points. Our data are consistent with that of Beaumont and associates. 38 They found, using different levels of impact-acceleration injury in rats, that beam-balance scores had not returned to normal by 31 days after injury (except in their mildest injury group), suggesting that persistence of beam-balance deficits depends on the level of severity of the injury.
Deficits in gross and fine motor skills often persist after TBI in humans. 36 Motor coordination can be assessed in rats using the beam-walking task. 28 This task requires both gross and fine motor coordination, as well as balance, and thus requires the coordination of multiple brain areas such as the sensorimotor cortex, the cerebellum, the vestibular system, and the thalamus. Others have found deficits in the beam-walk that recovered partially over a 2 week post-injury period. 35 Or, that by 31 days after injury, more severely injured groups had not returned to their baseline latencies; however, the moderately injured group did return to baseline. 38 This suggests that the beam-walk may be sensitive to the long-term effects of severe injuries but not mild injuries. The idea that beam-walking is mediated in part by the motor cortex and can be disrupted by injury to the cortex is supported by a lesion study that demonstrated damage to the cortical hindlimb area that is situated in the parietal cortex produced deficits in beam-walking performance. 39
Memory loss and malfunction are two of the most prevalent and persistent challenges facing patients after TBI. 36 Although different types of memory can be impaired by TBI in humans, working memory is particularly vulnerable. Working memory dysfunction is often overlooked upon casual observation in humans and can be difficult to measure in both humans and animals. Working memory is a system that holds and processes new and previously stored information transiently. It involves storing and manipulating information and executing a response, such as completing a task in the face of interferences such as delays or distractions. This differs from short-term memory, which serves as temporary information storage only.
In regard to rat behavior, Aggleton and co-workers 40 described working memory as retaining information that is useful for only one trial of an experiment but becomes irrelevant or misleading upon subsequent trials, so appropriate forgetting is mandatory. An example is discrete-trial alternation, which requires remembering the previous trial. These concepts were crucial in the design of the working memory version of the Morris water maze that we used here. 27,41 The paradigm we used has built-in measures of learning as well as working and reference memory, making it especially useful for studying TBI in rats. 27 Our results indicate that working memory deficits are evident at 3 months after injury and persist to the same degree out to one year after injury. Injured rats were able to learn the working memory task, but it took them longer than the uninjured rats—this is similar to what is seen in many patients suffering from chronic effects of brain injury. They are often able to learn new tasks, but require more time compared with uninjured cohorts. 42 In fact, there is continued debate as to whether the activity of working memory tasks in and of themselves has therapeutic potential to improve working memory. 43 Thus, the performance of the task over several days may contribute to improving the performance in brain-injured rats.
As opposed to working memory, reference memory refers to storage of information that remains stable over time, such as the information relevant to all trials of the spatial/reference memory version of the water maze. Reference memory is trial independent and remains relevant for many trials or an entire experiment, and is required to learn the rules of a task. 44,45 Some argue that the learning of consistent response associations (reference memory) does not require the hippocampus. 40 However, the classical, spatial-reference memory version of the water maze is hippocampal dependent because of the spatial nature of the information requiring the use of the directional cells of the hippocampus. Nevertheless, lesion studies have demonstrated that spatial navigational deficits can result from dysfunction in several brain areas, one of which is the hippocampus, 46 which is now well known to be particularly vulnerable to TBI. 47,48 In fact, TBI-induced spatial navigational deficits can occur in the absence of gross structural damage. 49 Subregions of the cerebral cortex are also likely to be involved such as the thalamus with its important corticothalamic circuitry. 46 What is notable in this study is that there were no apparent deficits in reference memory in injured compared with uninjured rats (either sham or naïve) until 12 months after injury, strongly indicating an interaction between brain injury and aging. 50
Conclusion
These results verify that parasagittal FPI produces deficits in multiple brain areas that are detectable using behavioral assessments out to 12 months after injury, making parasagittal FPI and the well-described behavior paradigms of neurological scoring, beam-balance, beam-walk, and the Morris water maze, working memory version, appropriate for investigating the chronic effects of TBI in rats.
Footnotes
Acknowledgments
We thank Maggie Parsley, for surgical preparation of subjects and assistance with scheduling, Maria-Adelaide Micci, PhD, for assistance in proofreading and editing the manuscript, and June Guptarak, PhD, for assistance with experimental design and behavior testing.
These studies were completed as part of a team funded by The Moody Project for Translational Traumatic Brain Injury Research.
Author Disclosure Statement
No competing financial interests exist.