
Abstract
We previously demonstrated that the inflammatory cytokine interleukin-6 (IL-6) activates the Janus kinase (JAK)-signal transducer and activator of transcription (STAT) signaling pathway in fibroblasts within the outer membranes of chronic subdural hematomas (CSDHs), and the activation of this pathway may induce CSDH outer membrane growth. The inhibitory system for this signal transduction pathway is unknown. CSDH fluids were obtained from 10 patients during trepanation surgery as the case group, and cerebrospinal fluid (CSF) samples were obtained from seven patients suffering from subarachnoid hemorrhage (SAH) on Day 1 as the control group. The concentrations of IL-6, soluble IL-6 receptor (sIL-6R), and soluble gp130 (sgp130) in CSDH fluid and CSF were measured using enzyme immunoassay kits. The co-localization of IL-6 and sgp130 in CSDH fluid was examined by immunoprecipitation. The expression levels of STAT3, JAK2, suppressor of cytokine signaling 3 (SOCS3), and protein inhibitor of activated Stat3 (PIAS3) in the outer membranes of CSDHs were examined by immunostaining. Soluble IL-6R and sgp130 concentrations in CSDH fluid were significantly higher than those in CSF after SAH. Sgp130 and IL-6 were co-immunoprecipitated from CSDH fluid. Immunostaining revealed STAT3, JAK2, SOCS3, and PIAS3 expression in fibroblasts located in the outer membranes of CSDHs. Soluble gp130 binds to IL-6/sIL-6R and acts as an antagonist of the JAK/STAT signaling pathway. SOCS3 also binds to JAK and inhibits its signaling pathway. In addition, PIAS3 regulates STAT3 activation. These factors might down-regulate the IL-6/JAK/STAT signaling pathway in fibroblasts within CSDH outer membranes. Therefore, these molecules may be novel therapeutic targets for the inhibition of CSDH growth.
Introduction
F
IL-6 induces the intracellular signaling pathway via a heterodimeric signaling complex consisting of the IL-6 receptor (IL-6R) and glycoprotein 130 (gp130). Then, the Janus kinase (JAK)-signal transducer and activator of transcription (STAT) signaling pathway, which regulates gene expression and cellular proliferation and differentiation, is activated. 5,6 We previously detected the activation of JAK/STAT signaling by IL-6 in fibroblasts and endothelial cells within the outer membrane of CSDHs, which suggests that JAK/STAT signal transduction may play an important role in the enlargement of the outer membrane. 7,8 Both IL-6R and gp130 exist in soluble and membrane-bound forms, which makes them highly dynamic, and they are important regulators of IL-6 signaling. 9 Interventions against IL-6/IL-6R/gp130 signaling, namely, the inhibition of IL-6 activity, were recently shown to be therapeutically effective in autoimmunity, inflammation, and cancer. 10 The protein inhibitors of activated STAT (PIAS) and the suppressor of cytokine signaling (SOCS) are negative regulators of JAK/STAT signal transduction. 6,11 The inhibition of activated STAT proteins and the modulation of their regulators have been shown to inhibit tumor cell growth; consequently, such molecules are new targets for anticancer therapy. 12 –14 However, the regulatory mechanisms of the IL-6/gp130/JAK/STAT3 signaling pathway in the outer membrane of CSDHs have not been clarified.
Therefore, the present study examined the expression of regulatory molecules of the JAK-STAT3 signaling pathway in the fluids and outer membranes of CSDHs. We performed enzyme immunoassay (EIA), immunoblot, and immunohistochemical analyses on the fluids and outer membranes of CSDHs.
Methods
Patients
Ten patients (six men and four women; 59–79 years of age; mean age, 68 years) with a CSDH confirmed by computed tomography (CT) or magnetic resonance imaging (MRI) were enrolled in this study. All patients underwent burr hole drainage surgery under local anesthesia at Aichi Medical University Hospital. All patients had a history of mild head injury without any hemostatic disorder, and no patients had received antiplatelet or anticoagulation therapy. The Ethics Committee of Aichi Medical University approved this clinical study.
Materials
All chemicals, unless otherwise specified, were obtained from Sigma Chemicals (St. Louis, MO).
Analysis of IL-6, soluble IL-6R, and soluble gp130
CSDH fluids were collected during trepanation surgery for all cases. For the control group, cerebrospinal fluid (CSF) samples were obtained through a cisternal drainage tube on Day 1 from seven patients undergoing neck clipping for subarachnoid hemorrhage (SAH). All samples were immediately centrifuged and the supernatants were stored at −80°C until analysis. The concentrations of IL-6, sgp130, and sIL-6R in CSDH fluid were measured by EIA (R&D Systems, Minneapolis, MN). The limits of detection for that assay were 0.70 pg/mL for IL-6, 0.08 ng/mL for sgp130 and 6.5 pg/mL for sIL-6R.
Immunoprecipitation of CSDH fluid
CSDH fluid samples from seven patients were used for further studies. CSDH fluid (500 μL) was incubated with IL-6 polyclonal antibodies (2 μL; Cell Signaling) at 4°C with gentle shaking for 1 h. Then, protein G sepharose (20 μL; GE Healthcare, Buckinghamshire, UK) was added to the mixture, and it was incubated for an additional hour at 4°C. The immunocomplexes were then washed with phosphate-buffered saline and boiled in 50 μL of Laemmli sample buffer for 7 min. All sample buffers were centrifuged and the supernatants were subjected to 7.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The proteins were then transferred to polyvinylidene difluoride membranes under wet conditions and incubated with primary monoclonal antibodies against gp130 (Millipore, Darmstadt, Germany) at a dilution of 1:750 overnight at 4°C. After washing, the membranes were incubated with secondary antibodies conjugated to horseradish peroxidase (Sigma) at a dilution of 1:3,000 for 30 min at room temperature. The reactions were developed with ECL (GE Healthcare). The bands were scanned with the image analyzer LAS-4000 (GE Healthcare). HeLa whole cell lysate served as the positive control.
Histological examinations
To study the cellular localization of STAT3, JAK2, SOCS3, and PIAS3, immunohistochemical staining was performed at room temperature following the avidin-biotinylated peroxidase complex (ABC) technique (n = 3). To preserve the outer membranes of the CSDH samples, they were incubated in 10 mL of ice-cold 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 3 h. Serial axial cryostat sections (10 μm) were placed on slides for staining. Non-specific immunoreactivity was blocked by incubation with goat serum for 30 min. The samples were treated with primary antibodies against STAT3 (Cell Signaling Technology) at a dilution of 1:300; JAK2 (Cell Signaling Technology) at a dilution of 1:50; and SOCS3 (Santa Cruz Technology, Dallas, TX) and PIAS3 (Cell Signaling Technology) both at a dilution of 1:100 overnight at 4°C. After washing, the samples were incubated with biotinylated anti-rabbit IgG for 1 h and then ABC for 1 h. Sera for the blocking step, biotinylated antibodies, and ABC were purchased from Vector Laboratories (Burlingame, CA). The reaction products were developed by incubating the sections in 0.05% 3.3′-diaminobenzidine tetrachloride and 0.01% H2O2 in 50 mM Tris-HCl (pH 7.5) for 10 min.
Statistical analysis
Data are expressed as the means ± the standard error. Significant differences between groups were assessed using the Mann-Whitney U test. The results were considered significant at p < 0.05.
Results
Concentrations of IL-6, sIL-6R, and sgp130
The IL-6 concentration was not significantly different between the CSDH fluid samples and the CSF samples from SAH patients (SAH CSF; Fig. 1A). However, the concentrations of sIL-6R and sgp130 were significantly higher in the CSDH fluid samples than in the SAH CSF samples (Fig. 1 B and 1C, respectively).
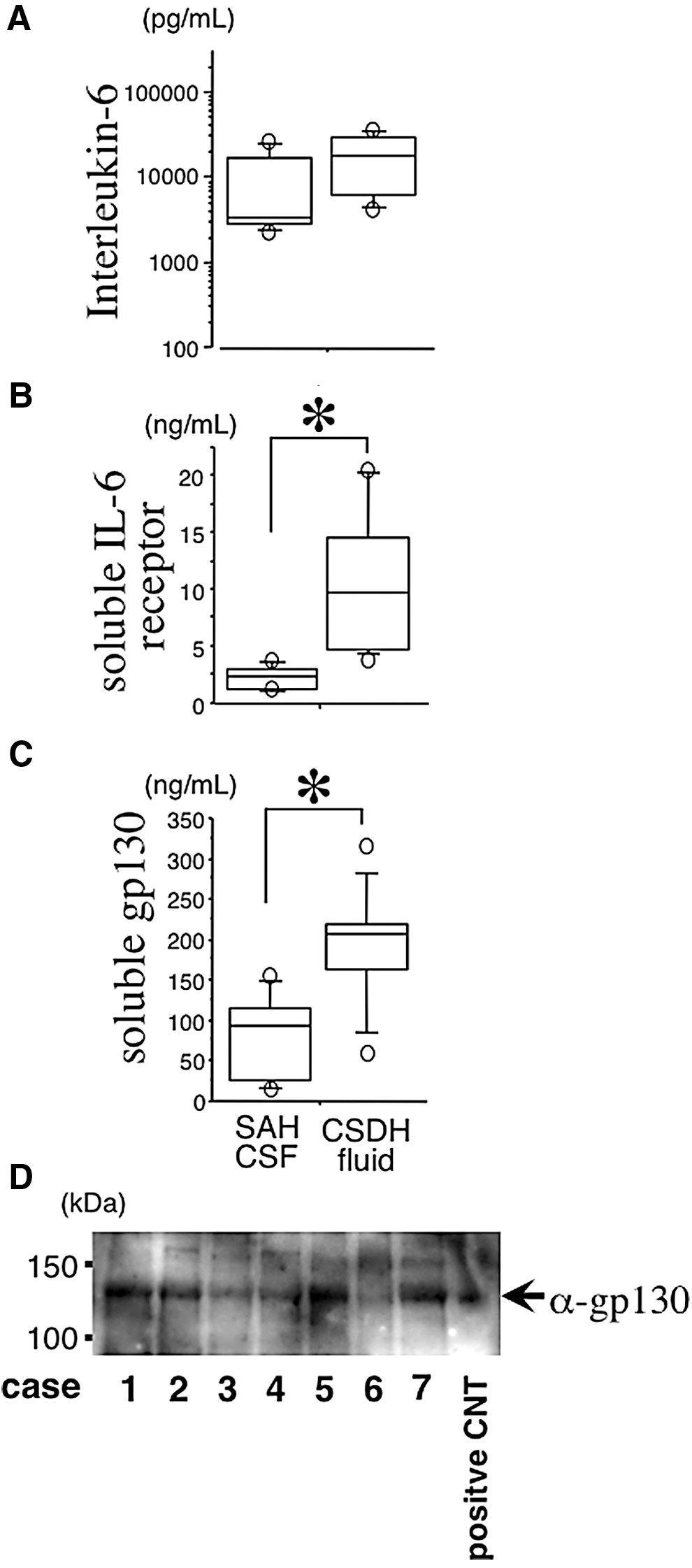
Concentrations of interleukin-6 (IL-6), soluble IL-6 receptor, and soluble gp130 (
Co-localization of IL-6 and sgp130 in CSDH fluid
Sgp130 bound to IL-6/sIL-6R was previously identified as an antagonist. We examined the interaction of IL-6 with sgp130 in CSDH fluid by immunoprecipitation. As shown in Figure 1D, sgp130 was detected in some cases, suggesting that sgp130 may regulate IL-6 signal transduction in CSDHs.
Histological observations
Hematoxylin and eosin staining of CSDH membranes showed that fibroblasts are located between collagenous fibers (Fig. 2A). STAT3, JAK2, SOCS3, and PIAS3 were primarily observed in fibroblasts (Fig. 2C, 2D, 2E, and 2F, respectively), and for the negative controls without primary antibodies, the fibroblasts were consistently negative for the markers listed above (Fig. 2B). STAT3, JAK2, SOCS3, and PIAS3 molecules also were observed in inflammatory cells, such as neutrophils, eosinophils, lymphocytes, macrophages, and plasma cells (Supplementary Fig. 1; see online supplementary material at

Hematoxylin and eosin staining demonstrating the presence of fibroblasts, collagenous fibers, inflammatory cells, and vessels
Discussion
In this study, we confirmed that sIL-6R and sgp130 expression levels were significantly higher in CSDH fluid than in SAH CSF; however, IL-6 expression was not significantly different between CSDH fluid and SAH CSF. The co-localization of IL-6 and sgp130 was confirmed by immunoprecipitation. Immunohistochemistry showed that PIAS3 and SOCS3 are mainly expressed in fibroblasts but also are expressed in inflammatory cells.
Several studies have shown that inflammatory cytokines and cyclooxygenase-2 (COX-2), known indicators of local inflammation, may play an important role in CSDH growth. 4,8,15,16 Further, anti-inflammatory cytokines in CSDH fluid help reduce the recurrence of CSDH. 17 STAT proteins are latent cytoplasmic transcription factors that become phosphorylated when signaling pathways are activated by various inflammatory cytokines, especially IL-6. IL-6 first binds to the IL-6R on the surface of the target cells. The IL-6/IL-6R complex then associates with the signal transducing membrane protein gp130, which is the second subunit of the IL-6 receptor complex. Dimerization of gp130 results in the transduction of an IL-6 signal and the subsequent phosphorylation of JAK. Activated JAK phosphorylates STAT at its tyrosine residue. Then, phosphorylated STAT3 dimerizes and translocates to the nucleus where it regulates target genes by binding to their DNA promoter regions. 6 Consistent with previous reports, our immunohistochemical assay showed that both JAK2 and STAT3 are expressed in fibroblasts. However, the details of the regulatory system that generates exceptionally high IL-6 levels in CSDH fluids are still unclear.
IL-6R and gp130 have both membrane-bound and soluble forms. 18 Membrane-bound gp130 is present on most, if not all, cells in the body; however, membrane-bound IL-6R expression is more limited and is predominantly confined to hepatocytes, neutrophils, and some lymphocytes. 18 IL-6 binds to sIL-6R and exerts its action through membrane-bound gp130, a process referred to as trans-signaling. 18 Consequently, soluble IL-6R is considered to have agonistic properties; however, the activity of the IL-6/sIL-6R complex is regulated by the soluble form of gp130 (sgp130). 18 Soluble gp130 binds to the IL-6/sIL-6R complex and prevents it from binding to membrane-bound gp130. This inhibits IL-6 signal transduction, suggesting that sgp130 acts as an antagonist of IL-6 signaling. 18 In inflammatory neurological diseases, such as Guillain-Barré syndrome, meningitis, and multiple sclerosis, sIL-6R expression is increased in the CSF, which suggests that the IL-6/sIL-6R complex plays a pivotal role in the central nervous system (CNS). 19 In multiple sclerosis, sgp130 expression is decreased in the CSF; this suggests that the effects of multiple sclerosis in the CNS may be due to a failure to antagonize IL-6. 20 The intracisternal injection of IL-6 induces long-lasting cerebral vasospasm. 21 Immediately after the onset of subarachnoid hemorrhage, the concentration of IL-6 in the CSF increases, and it remains high until the chronic stage of the disease is reached. During the course of SAH, the concentration of sIL-6R in the CSF does not change significantly; however, the concentration of sgp130 in the CSF decreases significantly after Day 5. 22 Therefore, IL-6 signals may be more easily transmitted after Day 5, which may result in cerebral vasospasm. Considering these previous data and those of our study, IL-6 signal transduction is regulated by sIL-6R and sgp130, and the distortion of the ratio of sIL-6R to sgp130 may be responsible for the increase in IL-6 signal transduction in CSDH fluids.
The stringent regulation of signal transduction pathways are necessary for appropriate cellular and physiological responses to cytokine stimulation to occur. PIAS3 inhibits the transactivation of a STAT3-responsive reporter gene and the DNA binding activity of STAT3. 23 Phosphorylation of STAT3 at Tyr705 induces the rapid formation of the PIAS3-STAT3 complex within 5 min and reduces STAT3-mediated gene transcription. 24 Glioblastoma is an aggressive and infiltrative malignant astrocytic glioma due to its high degree of cellularity, vascular proliferation, and necrosis. IL-6 plays an important role in the malignant progression of astrocytomas through the activation of STAT3. 25 Inhibition of STAT3 activation inhibits proliferation and induces apoptosis in glioblastoma multiforme cells. 26 PIAS3 expression was shown to be significantly lower in glioblastoma tissues than in control tissues. 27 These data suggest that the loss of PIAS3 contributes to enhanced STAT3 transcriptional activity and subsequent cell proliferation in glioblastoma and that therapies targeting the STAT3 signaling pathway may be potent interventions. Our data showed that PIAS3 is expressed in fibroblasts, which suggests that the growth of fibroblasts may also be regulated by PIAS3.
SOCS3 proteins are inducible inhibitors of cytokine signaling. SOCS3 binds to specific JAK-cytokine receptor complexes to control cytokine signaling by inhibiting JAK enzymatic activity. 28 STAT3 also plays an important role in skin remodeling and wound healing. Deletion of SOCS3 in keratinocytes resulted in aberrant STAT3 activation, epithelial hyperproliferation, and impaired wound healing, which suggests that SOCS3 helps prevent excessive IL-6 signaling. 29 Rheumatoid arthritis (RA) is one of the most immune-mediated diseases. Antigen-induced arthritis (AIA) is an animal model for arthritis, and its histopathological features closely resemble those of RA. IL-6 plays a crucial role in cartilage destruction and AIA development. 30 IL-6 also induces the proliferation of synovial fibroblastic cells in co-operation with sIL-6R. 31 The JAK/STAT3 pathway has critical roles in inflammation and joint destruction in RA. 32 The periarticular injection of a SOCS3 adenovirus was shown to drastically reduce the severity of arthritis and joint swelling. 32 Recently, tocilizumab, a monoclonal antibody against the IL-6 receptor, and tofacitinib, a JAK inhibitor, were recommended for the management of RA by the European League against Rheumatism (EULAR). 33 The IL-6-JAK-STAT3 signaling pathway plays an important role in RA. In our study, SOCS3 also was found to be expressed in fibroblasts within the outer membrane of CSDHs, which suggests that both PIAS3 and SOCS3 may be involved in the inhibition of fibroblast growth.
This study has several limitations. First, our findings are purely observational. We examined the presence of regulators of the JAK/STAT3 signaling pathway in the outer membranes and fluid of CSDHs; however, we did not perform any assays of the function of IL-6 while bound to sgp130 or any animal experiments to corroborate our findings. Further, the sample size was not sufficiently large to allow us to determine the roles of the assayed molecules in inflammatory cells in CSDHs. Additional definitive experiments are necessary to determine whether these molecules are associated with the regulatory system of the JAK/STAT signaling pathway in the outer membranes of CSDH.
In summary, this study was the first to examine the expression levels of sIL-6R and sgp130 in CSDH fluid and those of SOCS3 and PIAS3 in fibroblasts in the outer membranes of CSDHs. In particular, sIL-6R and sgp130 were shown to be co-localized with IL-6 in CSDH fluids and may inhibit IL-6 signal transduction. sIL-6R and sgp130 may precisely regulate the transduction of the IL-6/JAK/STAT signaling pathway in the outer membranes of CSDHs and control the growth of CSDHs. However, other unknown molecules may also be involved in the regulation of the JAK/STAT pathway. Additional in vivo studies or clinical trials of CSDH are necessary to determine if the IL-6/JAK/STAT signaling cascade is regulated by sIL-6R and sgp130. Interventions targeting sIL-6R and sgp130 may be beneficial for the treatment of recurrent and intractable CSDHs in the near future.
Footnotes
Acknowledgments
This work was supported in part by a Japanese Grant-in-Aid for Scientific Research (C), Grant Number 26462174 (K.O.), and by a grant (K.O.) from the General Insurance Association of Japan. The authors thank Hisae Inui for her technical assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.