
Abstract
Blocking of Connexin43 hemichannels, the main gap junction protein located on astrocytes in the central nervous system, has been shown to reduce neural injury in a number of models. We demonstrated previously that local administration of a Connexin43 mimetic peptide, Peptide5, reduces secondary tissue damage after spinal cord injury (SCI). Here, we investigated whether acute systemic delivery of Peptide5 is also protective in a model of SCI. Rats were subjected to a mild spinal cord contusion using the Multicentre Animal Spinal Cord Injury Study impactor and were injected intraperitoneally with Peptide5 or a scrambled peptide immediately and at 2 h and 4 h post-injury. Rats were tested for locomotor recovery and pain hypersensitivity and euthanized at 8 h, 24 h, two weeks, or six weeks post-injury. Compared with control rats, Peptide5 treated rats showed significant improvement in hindlimb locomotor function between three and six weeks post-injury and reductions in at-level mechanical allodynia at weeks one and six post-injury. Immunohistochemistry showed that Peptide5 treatment led to a reduction in total Connexin43 and increased phosphorylated Connexin43 at 8 h compared with scrambled peptide. At two and six weeks, lesion size, the astrocytic and the activated macrophage, and/or microglial response were all decreased in the Peptide5 animals. In addition, neuronal cell numbers were higher in the Peptide5 animals compared with the scrambled peptide treated rats at two and six weeks. These results show for the first time that systemic administration of Peptide5 to block the pathological opening of Connexin43 hemichannels is a feasible treatment strategy in this setting, ameliorating the secondary SCI.
Introduction
T
Gap junctions are specialized intercellular connections found in a multitude of animal species that enable direct and facilitated passage of various ions and small cytoplasmic molecules between two adjacent cells. 11 –14 A gap junction consists of two hexameric hemichannels inserted in the cell membrane, and each hemichannel is composed of six oligomerized connexin proteins. 15 Connexin hemichannels from adjacent cells are able to dock with each other to form functional gap junction channels. 16,17 In the spinal cord, Connexin43 is the major protein of both gap junctions and unopposed hemichannels on astrocytes and endothelial cells.
In pathological conditions, gap junction-mediated communication is involved in secondary cell death. Gap junctions have been considered to transmit neurotoxins including glutamate, nitric oxide, lactate, arachidonate, ammonia, and reactive oxygen species, as well as calcium waves from the initial injured cells to the adjacent healthy cells, known as the bystander effect. 18 –20 More recently, the opening of uncoupled hemichannels has been considered the principle perpetuator of secondary injury. Hemichannel opening facilitates the release of neurotoxins and adenosine triphosphate (ATP) and establishes calcium waves, and this can then affect surrounding intact cells. 21 –25
Connexin43 mRNA and protein levels are elevated after SCI 19,26 –29 and can be reduced using antisense oligodeoxynucleotides or gene knock-out to improve functional recovery. 26,30 –32 Mimetic peptides have also been used in experiments to modulate both gap junction coupling and hemichannel opening. 33 –37 These are short peptide sequences designed to match portions of the extracellular loops of the connexin proteins. Both gap junctions and hemichannels can be regulated by connexin mimetic peptides independently of each other depending on the concentration and incubation time. 28,36,38
A series of Connexin43 mimetic peptides has been designed against the two extracellular loops of Connexin43 protein, and one of these, referred to as Peptide5 (sequence VDCFLSRPETK), shows particular efficacy in regulating both Connexin43 gap junctions and hemichannels in a dose dependent manner 28 and improves cellular and functional recovery when delivered directly to a spinal cord contusion 29 without appearing to affect other connexin isoform gap junctions or hemichannels. Peptide5 has also been shown to be neuroprotective in retinal ischemia, 39 –41 cerebral ischemia, 42 and an epileptiform lesion model. 36 Additional mimetic peptides against Connexin43 protein 43 –48 have also been used for other indications but not for SCI repair. It is also possible that other pharmacological agents may be developed to regulate downstream effects of Connexin43 hemichannel opening 24 ; for example, a P2X2 receptor agonist has been used to block ATP released through these hemichannels. 49
Importantly, it is feasible for Peptide5 to reach injury sites systemically where the capillary bed has been compromised in the injured spinal cord, thus offering a practical therapeutic approach for people with SCI. In the current study, we have focused our investigation on determining whether systemic delivery of Peptide5, rather than local delivery, influences Connexin43 expression and recovery after SCI. We demonstrate that Peptide5 also improves cellular and functional outcomes when delivered systemically, as well as reducing chronic pain in an in vivo contusion model of rat SCI.
Methods
Experimental groups
There were 104 adult female Sprague-Dawley rats weighing 250–300 g used (Animal Resource Centre, Perth, Australia). All experimental protocols and procedures used in the study were approved by the Animal Care and Ethics Committee of the University of Technology Sydney, in accordance with the guidelines of the National Health and Medical Research Council of Australia. Rats were anesthetized with 4% isofluorane in oxygen (1 L/min) and then maintained at 2% isofluorane in oxygen (1 L/min). They were then placed on a heated pad and their body temperature was monitored throughout the surgery.
Anesthetic (bupivacaine, 0.5%, 0.02 mL) was injected locally at all surgical sites before incisions were made. Skin and muscle layers were incised over the dorsal midline and retracted, and a T10 laminectomy was performed to expose the spinal cord. The Multicentre Animal Spinal Cord Injury Study impactor device was used to produce a mild contusion injury (6.25 mm drop, 10 g weight) to the cord at T10. The mild SCI was chosen for this study because it results in less tissue disruption compared with more severe injuries but still has measurable locomotor and sensory deficits. The surgical wound was then sutured closed in layers. Control rats (Shams) had laminectomies but were left uninjured.
Peptide treatment
The dose of peptide used was to give a blood concentration of 100 μM, as shown to be effective in our previous study. 41 The following dosing regimen was used to maintain this concentration based on an expected half-life of the about 2 h in rat serum. The rats were administered intraperitoneally three doses of Peptide5 or scrambled peptide (SP) at 0 h (10 mg/kg, 0.5 mL), 2 h (5 mg/kg, 0.5 mL), and 4 h (2.5 mg/kg, 0.5 mL) after surgery. Peptide5 was designed to match a portion of the extracellular loop 2 of Connexin43 (sequence VDCFLSRPTEKT) as described previously. 28 A scrambled control peptide with the same amino acids but in a different order was designed as a control treatment to Peptide5 (sequence RFKPSLCTTDEV). Both peptides were solid phase synthesized by Mimotopes Pty Ltd., Australia, using Fmoc chemistries on a Protein Technology, Symphony instrument, purified by high-performance liquid chromatography and the structure confirmed by mass spectrometry analysis.
Post-operative care
Animals were housed individually for seven days post-surgery and then rehoused in groups of four. They were kept in a holding room with controlled temperature (22–25°C), humidity, and light cycle (12 h on/12 h off) and had access to food and water ad libitum. Post-operatively, animals were administered subcutaneously antibiotics (cephalothin sodium, 0.33 mg/kg,), analgesic (buprenorphine hydrochloride, 0.03 mg/kg), and supplementary fluid (Hartman replacement solution, 3.8 mL) twice daily for three days. Antibiotics were continued until urine was clear. Bladders were expressed manually twice daily for 1–2 weeks until the normal voiding response returned.
Behavioral assessments
After the surgical procedures, each animal was assessed for functional recovery using an open field and a horizontal ladder testing paradigm. Animals were tested at days 1, 3, 7, 14, 21, 28, 35, and 42 of the post-operative recovery period. Hindlimb locomotor function was assessed using the Basso-Beattie-Bresnahan (BBB) Locomotor Rating Scale for the open-field test 50 and by calculating the percentage of error hindlimb steps for the horizontal ladder test. 51 All tests were digitally recorded, and assessments were performed by two independent reviewers who were blinded to the groups. The scores from the two reviewers had a high correlation (R2 = 0.879) without statistically significant differences (p = 0.480), so the average scores were used for all further analysis.
Mechanical hypersensitivity tests were conducted a week before the surgical injury to determine the baseline level and then once weekly after surgery, for up to six weeks. The rats were placed on an elevated wire mesh platform in individual Perspex cubicles and were allowed to acclimatize to the pain testing apparatus for 40 min in a quiet and temperature controlled room. Mechanical hypersensitivity was measured at level (injury site) and below level (hindpaws) using a series of calibrated von Frey filaments with incremental stiffness ranging from 0.02 g to 60 g. The von Frey filaments were applied serially according to the up-down method, and the 50% withdrawal threshold was calculated, as described previously. 52
At-level pain was tested via application of the von Frey filaments, three times on both the medial and lateral side, approximately 3 cm away from the injury site. Below-level pain was tested via application of the von Frey filaments to the plantar surface of both hindpaws. Positive pain responses included licking, flinching, vocalization, and lifting of the leg in the case of below-level pain. Responses that occurred independent of the filament were not recorded.
Immunohistochemistry
At 8 h (n = 8), 24 h (n = 32), two weeks (n = 32), and six weeks (n = 32) after surgery, animals were euthanized with sodium pentobarbital intraperitoneally (100 mg/kg) and perfused intracardially with 200 mL of heparinized saline followed by 400 mL 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Approximately 20 mm of spinal cord centered at T10 was dissected, removed, and post-fixed in 4% paraformaldehyde overnight and then transferred to 30% sucrose. Spinal cords were sectioned horizontally (from the dorsal to ventral side) on a cryostat at 15 μm onto Matsunami Platinum PLC-14 and 2% gelatine-coated glass slides alternatively. Sections were mounted in a series of 10 so that each section on a slide was at 150 μm intervals. Mayer's hematoxylin and eosin (H&E) staining was performed to assess tissue morphology and determine the lesion area.
Immunohistochemistry was performed by first incubating slides in 5% normal goat serum (NGS) in phosphate buffered saline with Triton X-100 at pH 7.4 (PBST) for 30 min. Before the primary antibody incubation, heat-induced antigen retrieval was performed in citrate buffer (10 mM citric acid, pH 6.0) for 10 min in a pressure cooker for NeuN staining instead of NGS blocking. Primary antibodies were diluted in phosphate buffer with 5% normal goat serum (PBG) and incubated overnight at 4°C.
For the early time points (8 h and 24 h), primary antibodies used were rabbit anti-Connexin43 (1:2000, Sigma-Aldrich, Germany) to show levels of Connexin43 and rabbit anti-phosphorylated Connexin43 (1:100, Santa Cruz, Texas). This second antibody recognizes Connexin43 phosphorylation on Ser368, which has been shown to change Connexin43 hemichannels from an open to a closed state. 29,53 For two and six weeks, primary antibodies used were rabbit anti-glial fibrillary acidic protein (GFAP) (1:1000, Dako, Denmark) to label astrocytes, mouse anti-neuronal nuclei (NeuN) (1:1000, Millipore, Germany) to label neurons, mouse anti-CD68 (ED1) (1:2000, AbD Serotec, UK), and rabbit anti-ionized calcium-binding adapter molecule 1 (Iba1) (1:1000, Abcam, UK) to demonstrate activated microglia and macrophages.
Slides were then washed in PBST and incubated with goat anti-rabbit Alexa Fluor 488 (Connexin43, GFAP and Iba1; 1:200, Life Technologies), goat anti-rabbit Alexa Fluor 568 (phosphorylated Connexin43; 1:200, Life Technologies), goat anti-mouse Alexa Fluor 488 (NeuN; 1:200, Life Technologies), or goat anti-mouse Alexa Fluor 568 (ED1; 1:200, Life Technologies) in PBG for 2 h at room temperature. Slides were washed with PBST and counterstained by Hoechst (1:5000, Invitrogen) for 10 min. Primary antibody was omitted from negative controls. All slides were coverslipped in fluorescent mounting medium (Dako, Denmark).
Image analysis
Imaging was performed using an Olympus BH-2 light microscope with a PixeLINK Pl-A662 camera (H&E staining) or Olympus BX-51 light microscope with an Olympus U-RFL-T fluorescence burner and appropriate filters (immunohistochemistry staining). Injury on H&E stained sections was observed as hemorrhage, axonal swellings, cells with no nuclei or pyknotic nuclei, areas of cellular debris, or cavity formation. Sections were imaged at 150 μm intervals through the dorsal-ventral plane, and the injury area was measured on each section using Image J software. The lesion size was then calculated for each spinal cord as an approximate volume using these measurements at the lesion center and 3.5 and 7 mm distal to the lesion center.
Spinal cord sections were aligned using the central canal as an anatomical landmark for immunohistochemistry analysis. Images were taken adjacent to the lesion and at both 3.5 and 7 mm distal to the lesion center in both rostral and caudal directions.
Image analysis was performed using ImageJ software (National Institutes of Health, version 2X). Analysis of Connexin43, p-Connexin43, and astrogliosis was undertaken by measuring the staining intensity (mean grey scale value) in 10x images of midline sections. All slides were stained at the same time, and the images used for quantification were taken at the same microscope settings. In addition, the background black balance was normalized for each image to minimize any variation between slides.
For activated microglia quantification, all cells were identified with nuclear Hoechst stain and co-labeled with both ED1 (phagocytic macrophages) and Iba1 (microglia), counted manually in 40x images of midline sections, and reported as number of cells/field of view. Neurons were identified using NeuN and counted manually in 40x images of midline sections and in sections at 300 μm dorsal and ventral to the midline and are reported as number of cells/field of view.
Statistics
Statistical analysis was performed using Graphpad Prism (GraphPad Software, Inc, version 6). Data are expressed as mean ± standard error of the mean and analyzed using unpaired t tests, two-way analysis of variance (ANOVA) for the spinalized rats only with “treatment” and “time” as the two factors, two-way ANOVA at the different time points with “treatment” and “injury status (sham or SCI)” as the two factors. Finally, all the behavior data were analyzed using two-way ANOVA with treatment and injury status as the two factors, and using t tests to compare the two groups at each time point. Differences were considered to be statistically significant at p < 0.05.
Results
Surgery
All surgical procedures and intraperitoneal peptide injections were conducted without incident. None of the rats undergoing sham surgeries showed any indications of SCI or any post-operative complications. The impactor parameters for the SCI rats in the Peptide5 (n = 28) and SP (n = 28) groups were not significantly different (p > 0.05) for impact velocity (0.319 ± 0.004 m/sec vs. 0.319 ± 0.04 m/sec), compression (1.502 ± 0.025 mm vs. 1.505 ± 0.025 mm), or compression rate (0.281 ± 0.004 m/sec vs. 0.281 ± 0.004 m/sec). After SCI, rats showed immediate loss of locomotor function that gradually improved over the study period consistent with a mild SCI. Bladder function and weight gain returned to normal within one week to 10 days for all rats.
Connexin43 and phosphorylated Connexin43 at 8 h post-injury
Analysis of Connexin43 protein and phosphorylated Connexin43 protein (closed hemichannels) was undertaken at 8 h post-injury using immunohistochemistry on the midline sections. Connexin43 expression, as measured by gray scale values of immunohistochemistry labeling intensity, were lower in the Peptide5 (n = 4) treated animals compared with the SP (n = 4) control group in both gray (63.35 ± 6.68 vs. 49.51 ± 2.69) (p < 0.01) and white matter (46.91 ± 5.43 vs. 28.41 ± 2.57) (p < 0.001) (Fig. 1A, 1B). In contrast, there was an increase in the level of Connexin43 phosphorylation at 8 h after injury in the Peptide5 treated animals (n = 4) compared with the animals receiving SP control treatment (n = 4) with an increase in overall labeling intensity from 30.93 ± 2.53 to 48.61 ± 1.71 in gray matter (p < 0.001) and from 15.75 ± 1.96 to 24.90 ± 1.28 in white matter (p < 0.001) (Fig. 1C, 1D).
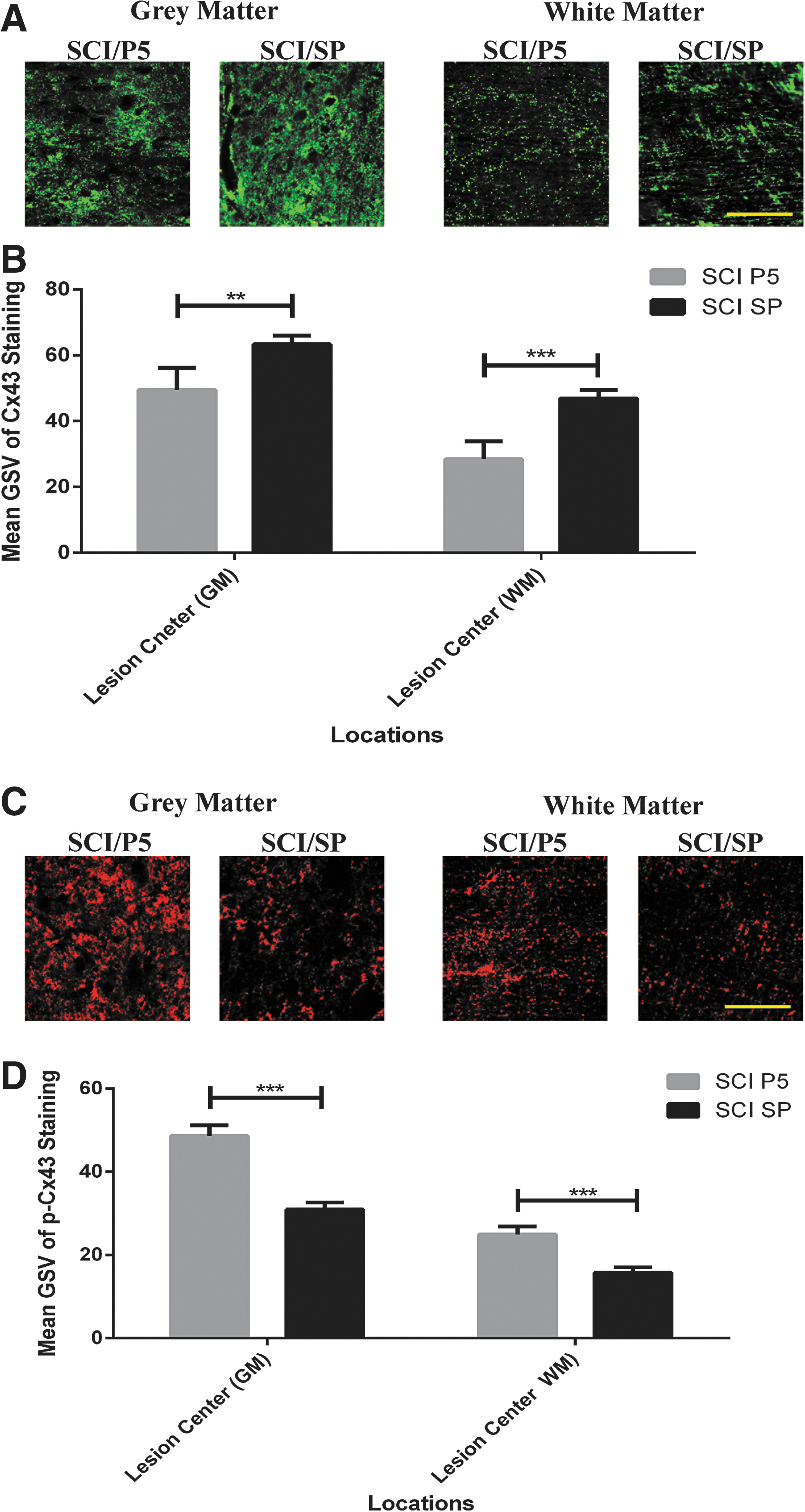
Changes in Connexin43 and phosphorylated Connexin43-Ser368 levels at 8 h after spinal cord injury (SCI). (
Lesion size
There was no histological evidence of any tissue damage in the sham animals at any time point. For the SCI groups, there was an obvious lesion apparent from 24 h post-injury that increased in size and changed in morphology over the six week period. At the earlier time points, there was more hemorrhage visible, and over time this changed to include more cell loss and increased cellular debris before development of a clearly defined central cavity. Lesion size was significantly reduced by the systemic administration of Peptide5 in comparison with the SP control treatment at all time points post-injury.
Longitudinal H&E stained sections including the lesion are shown for two weeks (Fig. 2A) and six weeks (Fig. 2B) after SCI. There were significant differences between treatment groups (p < 0.001) but not over time using two-way ANOVA tests. At two weeks post-SCI, the lesion was 6.72 ± 1.24 mm3 in the SP control group (n = 8), while the Peptide5 treatment (n = 8) was able to limit the lesion size to 2.58 ± 0.89 mm3 (p < 0.01) (Fig. 2C). By six weeks post-injury, the lesion was found to be 7.55 ± 1.55 mm3 in the SP treated animals (n = 8) while the lesion in the animals receiving Peptide5 treatment (n = 8) was limited to 3.60 ± 0.30 mm3 (p < 0.01) (Fig. 2C).
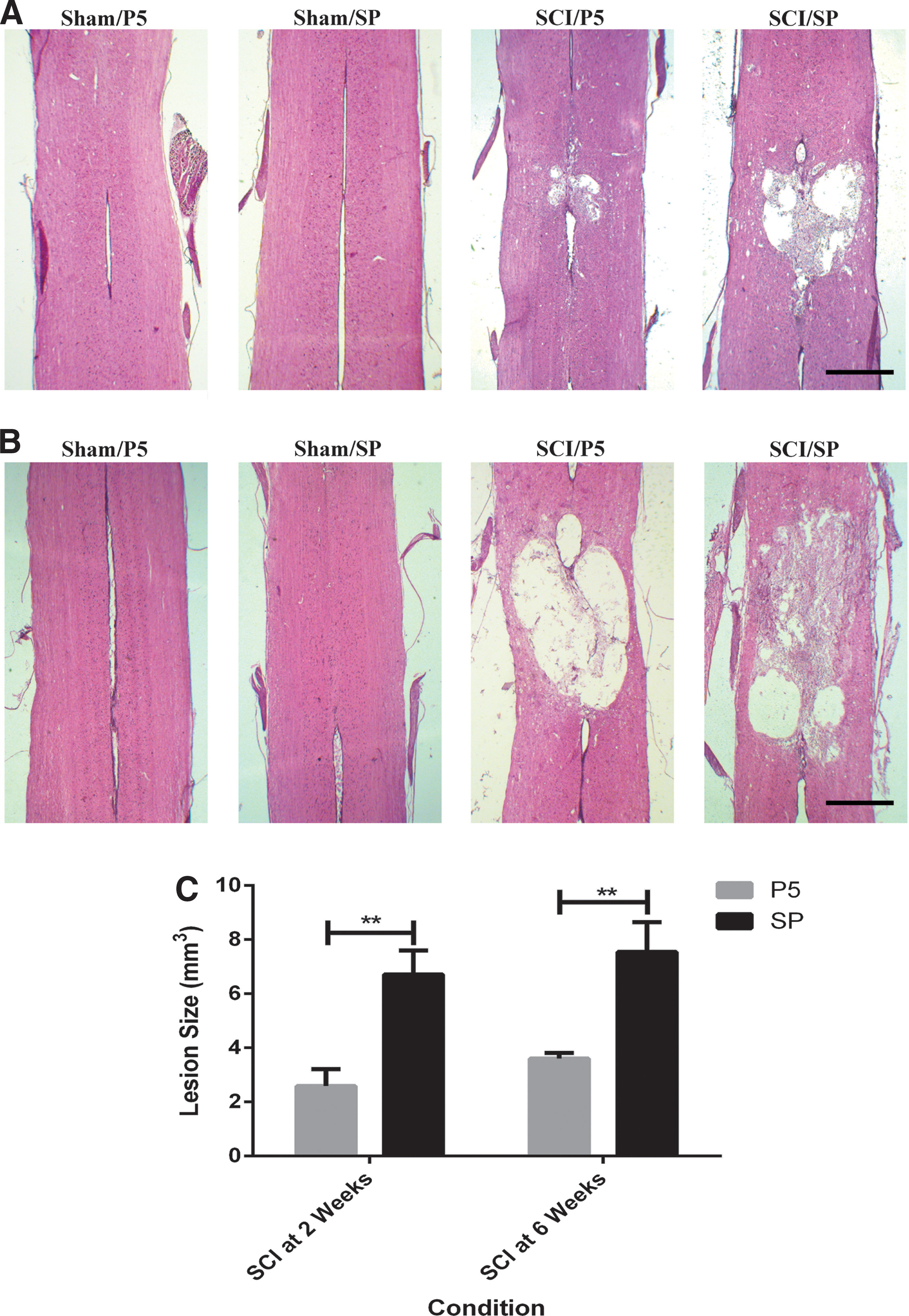
Longitudinal sections at the midline level of the spinal cord stained with Mayer hematoxylin and eosin showing the lesion at (
Astrogliosis (GFAP immunohistochemistry)
Astrogliosis (astrocyte activation) is known to be a key response after SCI and ultimately results in the formation of a glial scar starting from approximately one week. GFAP immunohistochemistry sections at the midline level were used to evaluate the effect of Peptide5 on astrogliosis after SCI (Fig. 3A for representative sections at two weeks post-surgery). The GFAP staining intensity at the lesion edge was quantified in mean gray scale value for statistical analysis (Fig. 3B, 3C).
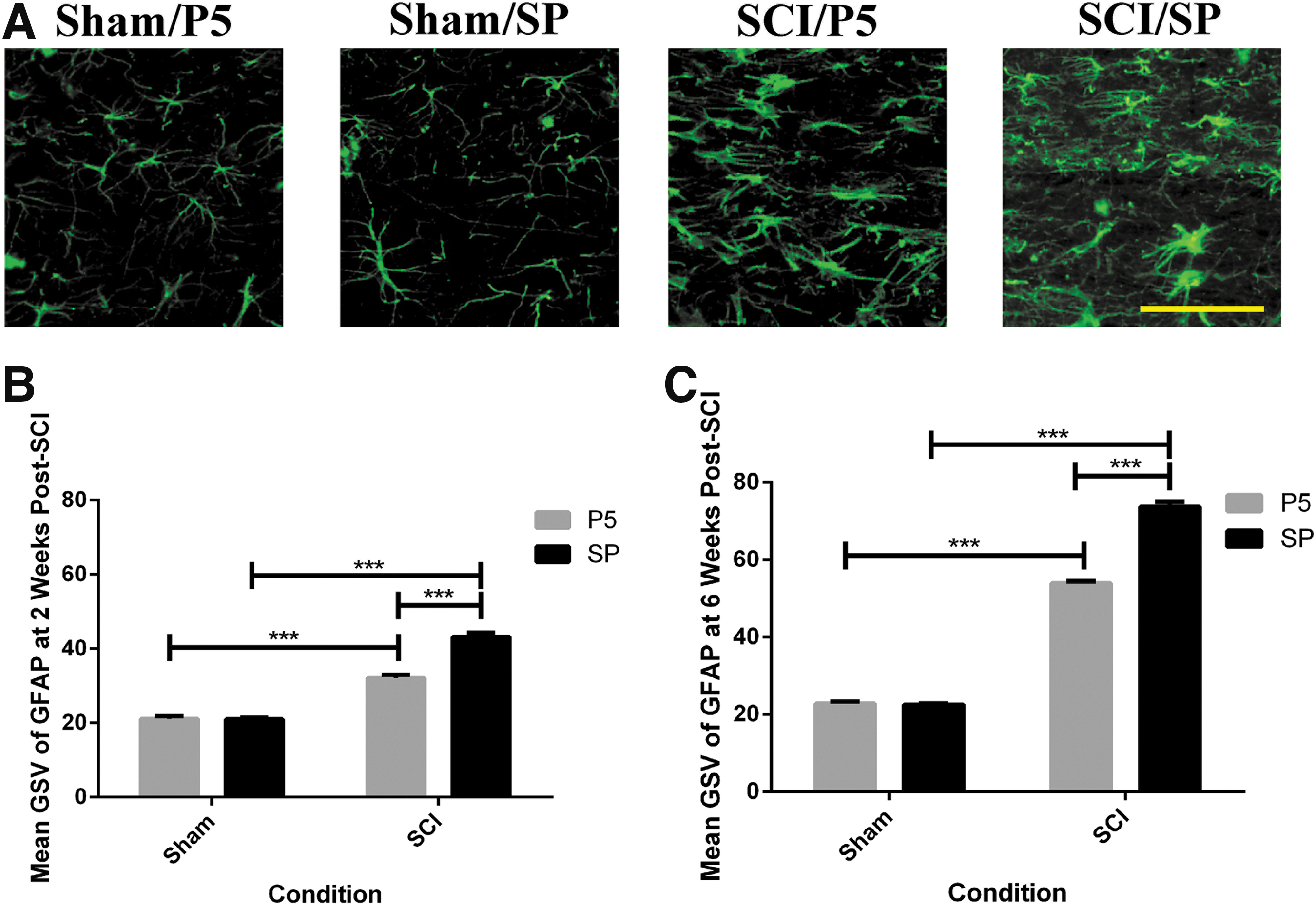
Glial fibrillary acidic protein (GFAP) expression at the lesion edge after contusion spinal cord injury (SCI). (
The normal patterns and intensity of astrocyte staining were seen in the sham spinal cords, and these measurements were used as the baseline for comparison for injury and for treatment. The level of GFAP immunoreactivity in the SP control animals at two and six weeks post-injury (n = 8 at each time point) was increased 2.1 and 3.3 times, respectively, compared with the sham groups (n = 8 at each time point) (two weeks, p < 0.001; six weeks, p < 0.001 for injury status). The Peptide5 treatment resulted in a significantly reduced level of GFAP immunoreactivity compared with SP treated animals with SCI at both post-injury time points (n = 8 at each time point) (two weeks, p < 0.001; six weeks, p < 0.001 for treatment) with levels of 1.5 and 2.4 fold decrease compared with that seen in sham animals, respectively (n = 8 at each time point).
Microglial/macrophage activation (Iba1 & ED1 immunohistochemistry)
One of the events that occurs during secondary SCI is the activation of microglial cells and the recruitment and activation of macrophages. These cells are involved in the inflammatory cascades that follow injury and are generally thought to be detrimental to recovery. Activated microglia and/or macrophages were identified using Hoechst (nuclear staining) and double labeling with Iba1 and ED1 (Fig. 4A for representative sections at two weeks post-surgery). Because activated microglia and macrophages share similar cellular markers and switch their genotypes and phenotypes frequently, it is difficult to distinguish between them. We therefore considered the Iba1+/ED1+ cells in the injured spinal cords as activated microglia and/or macrophages.
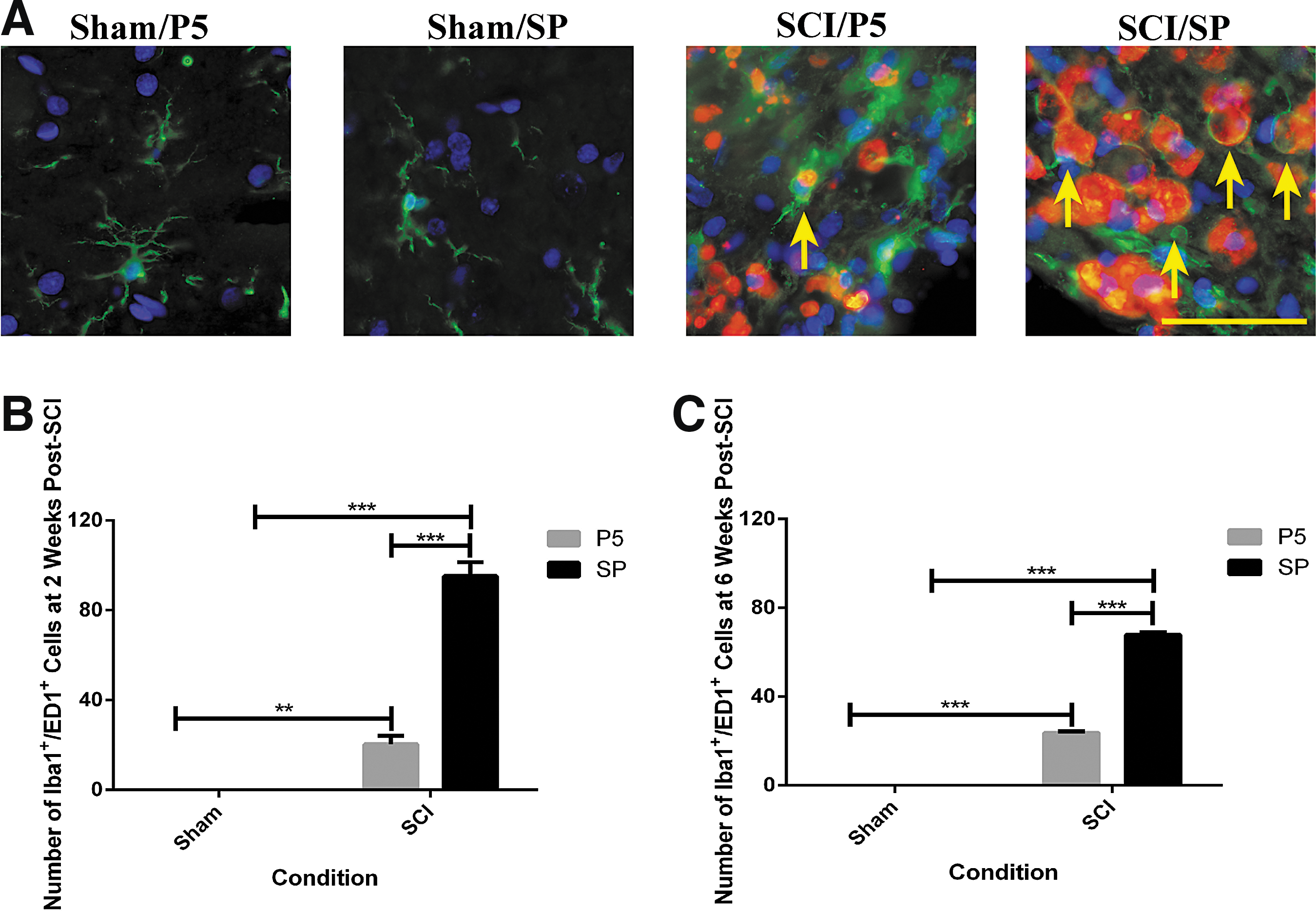
Activation of microglia and/or macrophages at the lesion edge after contusion spinal cord injury (SCI). (
The activated microglia and/or macrophages were observed in all areas of both gray and white matters, and the quantification of midline sections was conducted at the lesion edge where the cells double labeled with Iba1 and ED1 appeared in the greatest number (Fig. 4B and C). There were no Iba1+/ED1+ cells in any of the sham surgery spinal cords. In the SCI rats treated with SP (n = 8 at each time point), the number of Iba1+/ED1+ cells increased to 95 ± 6 cells/field of view at two weeks (p < 0.001) and 68 ± 1 cells/field of view at six weeks (p < 0.001 for injury status). Peptide5 treatment (n = 8 at each time point) reduced the number of Iba1+/ED1+ cells at both two weeks (20 ± 4 cells/field of view, p < 0.001) and at six weeks (24 ± 1 cells/field of view, p < 0.001 for treatment).
Neuronal survival (NeuN immunohistochemistry)
A significant loss of neuronal cells during the secondary phase of SCI is known to be a consequence of inflammatory responses including microglial/macrophage activation. Because Peptide5 treatment via systemic delivery resulted in a reduced level of activated microglia and/or macrophages in the secondary phase, it was important to determine whether neuronal survival was enhanced; for this, we used NeuN immunohistochemistry (Fig. 5A for representative sections at two weeks post-surgery). The surviving neuronal nuclei were identified by NeuN positive staining and then quantified at 7 mm to the lesion center at the midline level (Fig. 5B, 5C). There were no significant differences between the cell counts at 7 mm rostral and caudal to the lesion center using the paired t test; therefore, the mean of rostral and caudal measurements for each sample was used for further analysis.
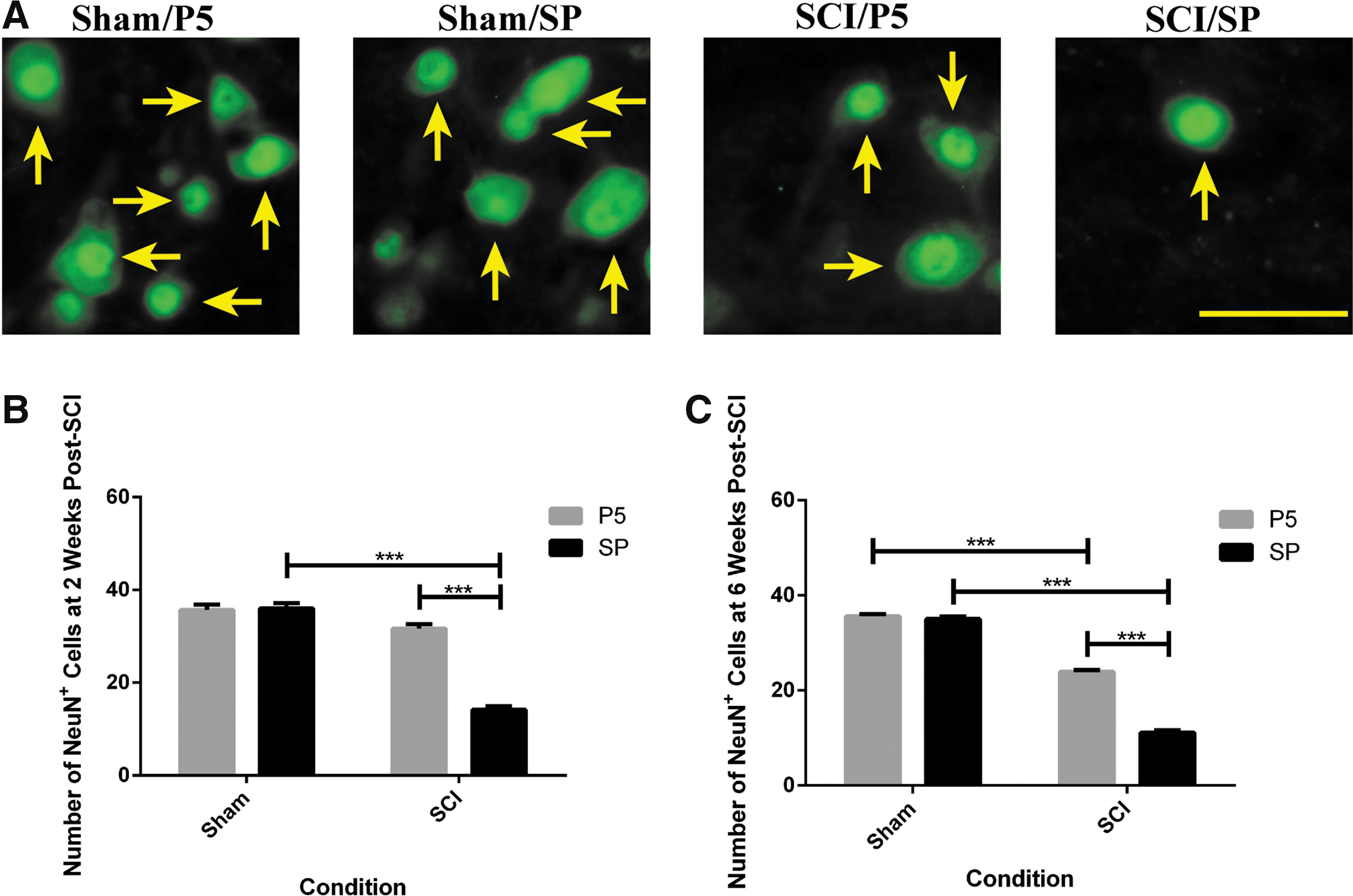
Surviving neuronal cells after contusion spinal cord injury (SCI). (
Neuronal cell number and distribution was not altered over time in the sham animals (36 ± 0 cells/field of view). As anticipated, the number of neurons per field of view in the SP treated animals (n = 8 at each time point) significantly decreased to 15 ± 1 cells/field of view at two weeks (p < 0.001) and 11 ± 1 cells/field of view by six weeks (p < 0.001 for injury status) post-injury. In Peptide5 treated animals (n = 8 at each time point), this number was retained at 32 ± 1 cells/field of view at two weeks (p < 0.001) and 24 ± 0 cells/field of view by six weeks (p < 0.001 for treatment) post-injury compared with the injured animals that received SP control treatment (n = 8 at each time point).
The number of neurons in the dorsal horn showed a similar pattern to that at the midline level, but the neurons in the ventral horns did not decrease significantly after this mild contusion injury at the dorsal level of the spinal cord (data not shown). Therefore, the Peptide5 treatment via systemic delivery was able to promote neuronal survival during the secondary phase of SCI.
Motor function (open field and error ladder)
We used two behavioral assessments—open field and horizontal ladder tests—to determine whether Peptide5 treatment led to an improvement in functional recovery. The animals with SCI treated with Peptide5 (n = 8) showed a significant improvement in hindlimb locomotor function in both tests from week three compared with the injury animals treated with SP (n = 8) (Fig. 6A, 6B). There was no deficit in hindlimb locomotion in any of the sham animals regardless of treatment (n = 8 for each treatment) for both behavioral assessments.
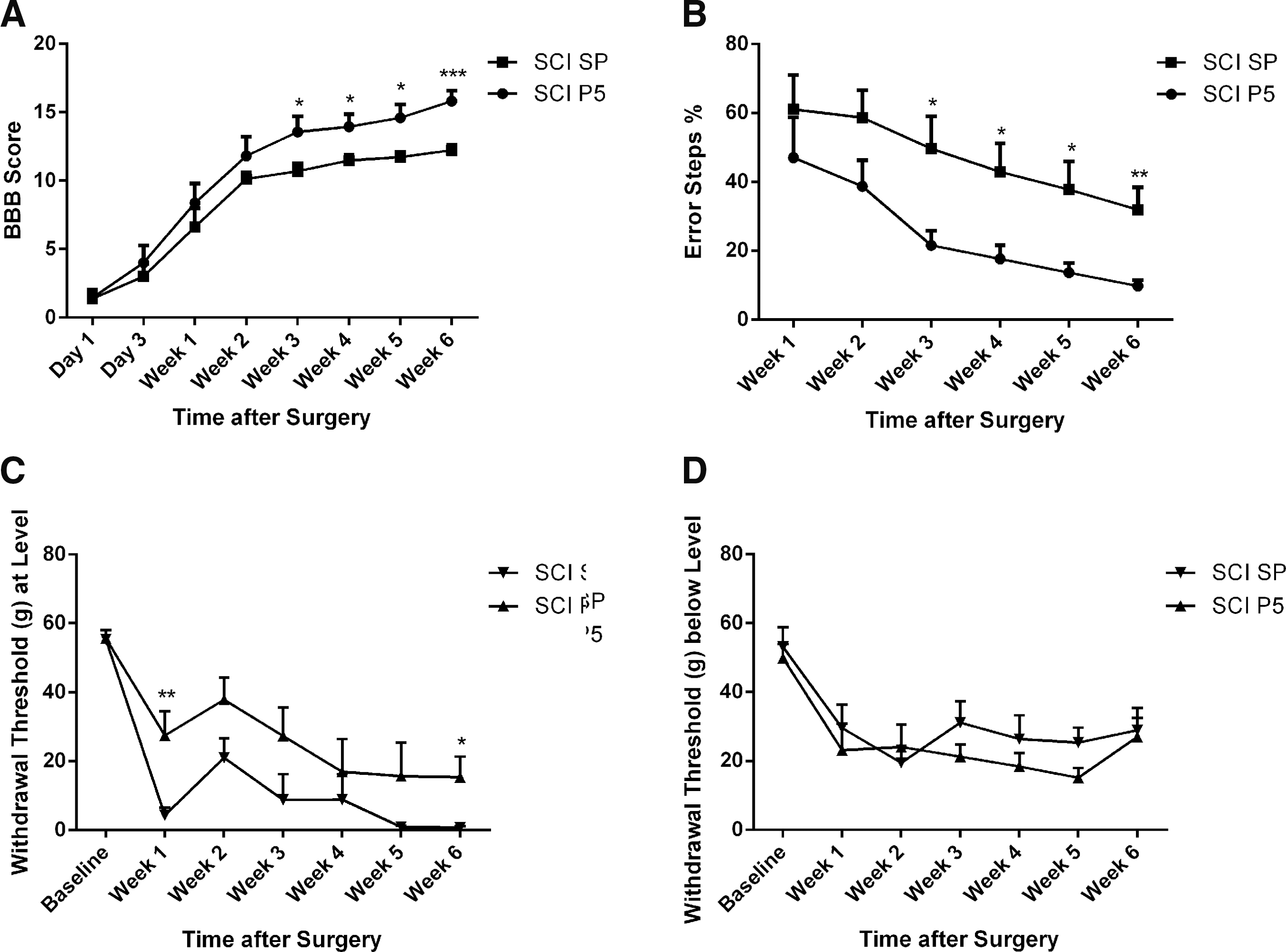
Behavioral assessments of hindlimb locomotor function after contusion spinal cord injury (SCI). (
In the open field test, injury groups displayed improvements in the BBB scores from day one to week six (SCI/Peptide5, 1.44 ± 0.50 to 15.81 ± 0.76, p < 0.01; SCI/SP, 1.00 ± 0.67 to 12.00 ± 0.41, p < 0.01) as expected after a mild contusion SCI (Fig. 6A), but Peptide5 treated injury animals showed improved hindlimb locomotor function when compared with the SCI animals treated with SP with significant differences observed from week three (13.56 ± 1.14 vs. 11.00 ± 0.57, p < 0.05) to week six (15.81 ± 0.76 vs. 12.00 ± 0.41, p < 0.001) at which time point behavioral assessment ceased.
Similar results were obtained with a horizontal ladder test (Fig. 6B). All injury animals with Peptide5 or SP treatments exhibited a reduction in the percentages of error steps from week one to six (SCI/Peptide5, 47.20 ± 11.68% to 9.64 ± 1.66%, p < 0.001; SCI/SP, 61.09 ± 10.02% to 32.04 ± 6.58%, p < 0.001). The injury animals treated with Peptide5, however, displayed fewer hindlimb error step percentages compared with those that received the SP control treatment with significant differences seen from week three (21.65 ± 4.25% vs. 49.72 ± 9.42%, p < 0.05) to week six (9.64 ± 1.66% vs. 32.04 ± 6.58%, p < 0.01). Differences in the percentages of error steps at weeks one and two were seen between Peptide5 and SP treated injury animals; however, these differences were not statistically significant (p > 0.05).
Sensory function (mechanical allodynia)
Spinal astrogliosis is well regarded as a contributor to the development of neuropathic pain states via the release of small excitatory molecules through opened hemichannels after SCI. 54 Therefore, we investigated whether treatment with Peptide5 alleviates mechanical pain hypersensitivity after SCI. At-level hypersensitivity demonstrated by reduced withdrawal thresholds after the injury developed in rats with SCI. Compared with SP treated controls (n = 16), treatment with Peptide5 (n = 16) significantly improved at-level pain with a 41% increase in the withdrawal threshold at week one (p < 0.01) and a 27% increase in the withdrawal threshold at week 6 (n = 8) (p < 0.05) after SCI (Fig. 6C). Below-level mechanical pain hypersensitivity was also measured on the hindpaws (Fig. 6D). There was no significant difference in paw withdrawal thresholds between the treatment groups during the course of the six weeks, however (p > 0.05) (Fig. 6D).
Discussion
We have demonstrated previously that the local application of Peptide5 at a low concentration within the first 24 h after traumatic SCI reduces tissue swelling, lesion spread, inflammation, astrogliosis, neuronal cell death, and functional deficits in ex vivo and in vivo models. 28,29 Acute administration of Peptide5 at low doses has also been shown to be neuroprotective in models of retinal and cerebral ischemia. 39 –41,55 –57 The local administration of Peptide5, however, is not practical for use in spinal patients because this would necessitate a highly invasive surgical procedure to expose the injury area or involve another procedure such as inserting an intrathecal catheter.
Upregulation of Connexin43 has been demonstrated early after SCI. 19,27,29 Therefore, application of Peptide5 should occur within 8 h, before maximal upregulation of Connexin43 protein, to be most effective. Unfortunately, many patients with SCI arrive in emergency departments with traumatic co-morbidities or life-threatening injuries and a timely spinal cord procedure is not always possible. If the acute clinical management of a patient with SCI includes an open surgical procedure such as decompression or stabilization, there may be an avenue for application of either an Connexin43 antisense oligodeoxynucleotide in a topical gel 26 or a direct application of Peptide5. 29 Unfortunately, in most cases, these procedures do not occur early enough for effective regulation of Connexin43 protein.
Because mimetic peptides have been shown to permeate the lesion site of spinal cords through the disrupted blood spinal cord barrier, 58 vascular administration of Peptide5 is a suitable approach for early delivery to the lesion. Our findings demonstrate the neuroprotective effects of Peptide5 via this systemic delivery is in agreement with our previous studies using direct peptide administration. 28,29
Effect of Peptide5 on Connexin43 protein in the spinal cord
The increase in total Connexin43 protein level after injury was suppressed with administration of Peptide5 and phosphorylated Connexin43 was higher in the spinal cords at 8 h after Peptide5 administration. There is substantial evidence to indicate an increase of Connexin43 protein after brain ischemia 59 –63 and SCI. 19,26 –29 Down-regulation of connexin or blocking gap junction channels using gap junction blockers has been shown to be beneficial. 26,63 –65 Other studies using gene knockout models have demonstrated conversely an increase in lesion size resulting from complete loss of Connexin43 stopping normal gap junctional communication necessary for spatial buffering and neuronal survival at the later stages of recovery. 66 –68
The exact mechanisms of Peptide5 are still being elucidated, but at the concentration used in this study, it is acting to block Connexin43 hemichannels rather than affecting intact gap junctions. 28 It is thus logical to assume that Peptide5 is acting by blocking Connexin43 hemichannels leading to a reduced propagation of neurotoxins to the surrounding healthy cells and extracellular matrix. 29,40,41,55,69
Neither the sham animals treated with Peptide5 nor the animals treated with SP showed any evidence of adverse effects in this study or in other studies using Peptide5. 41,57 We do not think that Peptide5 will affect the uninjured tissue nor should it act globally on gap junctions in other organs at the concentration we are administering. Certainly, higher doses may prove deleterious, and this has been shown to be the case in a fetal model of ischemia in sheep. 62 While long-term effects are unknown at this stage and future studies will have to determine whether Peptide5 treatment has any effects on the levels of other connexins in the injured spinal cord, only transient block is required to obtain the improved outcomes described.
Targeted tissue effects of Peptide5 in injured spinal cord tissue
Injured spinal cords treated with Peptide5 had smaller lesions, decreased astrogliosis, less inflammatory cell infiltration and activation, and increased neuron survival, all indicating reduced effects of secondary injury and lesion spread. There is increasing evidence to suggest that the uncontrolled opening of Connexin43 hemichannels causes release of neurotoxins and ATP from activated microglia and/or macrophages, thus propagating secondary injury. 22 –25,37 In addition, many of these neurotoxins including glutamate and reactive oxygen species have been shown to increase the opening of Connexin43 hemichannels, 21,22,70 –72 thus forming a positive feedback loop. We show here that blocking Connexin43 hemichannels with Peptide5 halts the amplification of injury and reduces a number of downstream effects that would normally be part of the secondary injury cascade, including astrogliosis.
Normally, astrogliosis will commence in the rat spinal cord from about three days after injury and lead to glial scar formation, a major physical and chemical barrier to neural regeneration. 73 –77 The physical barrier acts to prevent remyelination and regrowth of axons, 78 –80 while the chemical barrier adds to the nerve regrowth inhibitory microenvironment by producing a number of molecules such as chondroitin sulfate proteoglycans, tenascins, and bone morphogenetic proteins. 4,81 –86 The reactive astrocytes in astrogliosis are believed to be adverse for neuroregeneration by inhibiting axonal regeneration and oligodendrocyte differentiation from precursor cells, leading to remyelination failure, as well as generating and secreting nitric oxide, a neurotoxin that damages local neural cells contributing to the secondary injury. 74,84,87 –89
Phagocytes found at the lesion area of CNS injury arise from two sources: resident microglia proliferating in situ and infiltrating macrophages recruited from the bloodstream. Both become activated and remain for several weeks after injury in response to the pro-inflammatory environment at the site of injury. 90 –93 With the systemic administration of Peptide5, there was a decrease in the number of activated microglia and/or macrophages present in or adjacent to lesions. Studies have shown that activated microglia and/or macrophages are involved in an acute inflammatory response to neurotrauma, such as synthesis and release of cytokines and chemokines. 94 –99 These pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1ß, and IL-6 are believed to cause neuronal and glial toxicity. 1,99 –102
The activated microglia and/or macrophages have been reported to increase Connexin43 expression and gap junction coupling in response to a trauma in the CNS. 103 The decrease in activated microglia and/or macrophages in Peptide5 treated spinal cords may not only be a downstream consequence of a decrease in astrocytic Connexin43, but may also result from Connexin43 hemichannel closure blocking pro-inflammatory signaling molecule release from the activated microglia and/or macrophages themselves.
Death of neuronal cells after CNS injury is thought to be a consequence of a complex cascade of events involving uncontrolled opening of Connexin43 hemichannels, spread of inflammation, and release of neurotoxic molecules. 12,22,104 There is growing evidence to suggest that a number of neurotoxins are synthesized and released from the activated microglia and/or macrophages in response to injury, including glutamate, nitric oxide, pro-inflammatory cytokines, ATP, and prostaglandins. 31,101,102,104 –106 These neurotoxic molecules enhance the opening of astrocytic Connexin43 hemichannels, resulting in a further release of neurotoxins from the injured astrocytes. 107,108 Thus, there is a positive feedback loop developed in the lesion area to decrease the neuroprotective functions of astrogliosis, leading to an increase of neuronal cell death via the activation of amyloid-β peptide 25 –35 and pannexin 1 hemichannel pathways. 107,109
Behavioral improvements
Most importantly, behavioral assessments over a six-week period after injury showed significant improvement in hindlimb locomotor ability in the Peptide5 treated animals. Given the time frame of the current study, we believe this represents a reduction in tissue damage that translates directly to a reduction in the degree of functional deficit seen after injury. By three weeks after injury, the Peptide5 treated animals reached an average BBB score of 14, which corresponds with consistent weight-supported plantar steps. This is a significant improvement in rat locomotor function and affords the rat increased mobility and activity compared with a control injury group BBB score of less than 9, where rats have no weight bearing stepping at all. 50 In the error ladder test, we also saw improvements in function with the Peptide5 treatment that were indistinguishable from the sham operated rats.
At-level pain after SCI has been reported previously to be the most common of neuropathic pain types (41% of subjects), with an early onset and pain described as severe or excruciating. 110,111 In the current study, we demonstrated that Peptide5 treatment was able to alleviate at-level pain during the early (one week) and later (six week) stages of recovery after SCI, confirming the role of connexin hemichannels in the development of neuropathic pain. 112
Treatment with Peptide5 had no effect on paw withdrawal threshold in response to mechanical stimuli. It is noteworthy that interpretation of spinal-mediated reflex response may be problematic when assessing below-level SCI pain because of the development of hyperactive reflex circuitries. 113 Hindpaw stimulation caused increased innate reflex responses in the absence of increased brainstem responses, thus reflecting a spasticity syndrome rather than pain-like behavior. 113 In addition, in humans, below-level pain is reported to occur months to years after SCI, in contrast to the days/weeks for the development of at-level pain. 111 Although promising, further studies are required to determine the effects of Peptide5 treatment on the full spectrum of SCI-induced pain behaviors.
The findings of the current study provide strong in vivo data supporting the benefits of systemic delivery of the Connexin43 mimetic peptide, Peptide5, in modulating Connexin43 hemichannels as a feasible approach for acute SCI. These findings support the need to further investigate the timing of administration, the optimal dose, pharmacokinetics, and administration route for humans and to further evaluate the mode of action of Peptide5 to begin translation to clinical trials.
Footnotes
Acknowledgments
This work was supported financially by the Spinal Cord Injury Network, Australia and New Zealand and the CatWalk Spinal Cord Injury Trust. We acknowledge the University of Technology Sydney for the financial support in the form of postgraduate research scholarships for Yilin Mao. The authors would like to thank staff at the Ernst Facility, University of Technology Sydney, for their care of the experimental animals.
Author Disclosure Statement
Prof. Colin Green is a founding scientist of CoDa Therapeutics, Inc. (USA) that holds the intellectual property rights to Peptide5. For the remaining authors, no competing financial interests exist.