
Abstract
Traumatic brain injury (TBI) causes a primary insult and initiates a secondary injury cascade. The mechanisms underlying the secondary injury are multifactorial and may include the aberrant expression of long non-coding RNA (lncRNA) post-TBI. Here, lncRNA microarray analysis was performed to profile the altered lncRNAs in the rat hippocampus after TBI. A total of 271 lncRNA probe sets and 1046 messenger RNA (mRNA) probe sets were differentially expressed after TBI. Gene ontology analysis showed that the main components of the most significantly changed categories were inflammation, DNA transcription, apoptosis, and necroptosis. Additionally, the pathway analysis and the pathway relation network revealed correlated pathways mainly involving inflammation, cell cycle, and apoptosis. A co-expression network of these aberrantly expressed lncRNAs and mRNAs was further constructed to predict the potential function of individual lncRNAs. Sub–co-expression networks were formed for the top three lncRNAs: NR_002704, ENSRNOT00000062543, and Zfas1. Thus, our study demonstrated differential expression of a series of lncRNAs in the rat hippocampus after TBI, which may be correlated with post-TBI physiological and pathological processes. The findings also may provide novel targets for further investigation of both the molecular mechanisms underlying TBI and potential therapeutic interventions.
Introduction
T
Long non-coding RNAs (lncRNAs) of more than 200 nucleotides in length 10,11 lack protein-coding capabilities but can participate in many biological processes that coordinate gene expression. 12,13 Numerous lncRNAs are expressed in the brain. 14 EVF2, for example, is a nervous system-specific lncRNA that regulates the number of GABAergic interneurons in the early postnatal hippocampus and dentate gyrus. 15 The altered genome scale expression of lncRNAs and their potential roles in TBI, however, have not yet been determined.
Aiming to identify the vital lncRNAs involved in TBI and provide novel targets for further investigation, we investigated alterations in lncRNA expression in the rat hippocampus after TBI using microarray analysis. We then predicted the physiological and pathological function of those lncRNAs via the annotation of their co-expressed mRNAs. Sub–co-expression networks for the most significant lncRNAs were constructed. Additionally, four lncRNAs were randomly selected for further validation by quantitative reverse transcription polymerase chain reaction (RT-PCR). With a combination of microarray results, bioinformatics analysis, and subsequent quantitative RT-PCR, we revealed some potential roles of these lncRNAs that are aberrantly expressed after TBI.
Methods
Animals
Studies were performed using male Sprague-Dawley rats (weight, 300–350 g) that were housed for at least 7 days before surgery. Rats were allowed free access to water. Food was withheld overnight before surgery. All animal procedures were approved by the Animal Care and Experiment Committee of Shanghai Jiao Tong University School of Medicine and all experimental procedures were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Surgical preparation and fluid percussion brain injury
Surgical preparation and rat fluid percussion brain injury were performed as previously described. 6,16 In brief, animals were endotracheally intubated and anesthetized with a nitrous oxide/oxygen mixture (70%/30%) containing 2% halothane. During the surgical procedure, the animal body temperature was maintained at 37°C using a thermal heating blanket. A midline scalp incision was performed followed by a 4.8-mm craniectomy drilled on the left hemisphere, 2.5 mm lateral to the midline and midway between the bregma and lambda. A Luer Lock (Becton Dickinson, Mountain View, CA) needle hub was cemented into the craniectomy hole with super glue and dental acrylic. Moderate fluid percussion injury was induced by a swinging pendulum (VCU Biomedical Engineering, Richmond, VA), which was set to an angle of 25°. The peak pressure was 2.5 atmospheres, as measured by an extracranial transducer (Statham PA 85–100; Gould, Oxnard, CA) and recorded on a storage oscilloscope (Tektronix 5111; Tektronix, Beaverton, OR). The sham group received an identical surgical procedure but did not receive a fluid percussion injury.
Considering the future intervention practice, we set a 3 h post-injury as the time-point for the analysis. After a survival period of 3 h, the rats were anesthetized with sodium pentobarbital (100 mg/kg, intraperitoneal). When fully anesthetized, the rats were transcardially perfused with 200 mL of 4°C isotonic saline. The hippocampi from left hemispheres were then rapidly dissected and stored in liquid nitrogen.
Experimental groups and array data production
Twelve rats were randomly divided into two groups of six rats each. The TBI group was subjected to all surgical procedures and fluid percussion injury. The sham group received the identical surgical procedure but did not receive fluid percussion injury. In total, six hippocampi (three from the TBI group and three from the sham group) were used to obtain probes, with one hippocampus per microarray. All 12 samples were used for quantitative RT-PCR assessment.
The ipsilateral hippocampus (left hemisphere) of each rat was dissected for microarray analysis and lysed using TRIzol reagent (Invitrogen, Thermo Fisher Scientific Corporation, Carlsbad, CA), and the total RNA was extracted per the manufacturer's protocol. The total RNA from each sample was then quantified using a NanoDrop 1000, and 1 μg of total RNA from each sample was reverse transcribed into complementary DNA (cDNA). Biotin-labeled complementary RNA (cRNA) was then synthesized from the cDNA with T7 RNA polymerase and biotin-labeled ribonucleotides. The labeled cRNAs were hybridized onto an Affymetrix GeneChip Rat Gene 2.0 ST Array (Affymetrix, Santa Clara, CA). After the slides were washed, the arrays were scanned using a GeneChip Scanner 3000 7G.
Differential expression analysis for lncRNAs and mRNAs
Differential expression analysis was performed using a t-test, and the Q-value (a natural positive false discovery rate analogue of the p value) 17 was used to determine statistical significance. Significantly differentially expressed lncRNAs were defined as having a corrected Q-value <0.25 and absolute fold change >1.2, and significantly differentially expressed mRNAs were included in the analysis with the criteria of a corrected Q-value <0.1 and absolute fold change >1.2. Two-dimensional hierarchical clustering was used to arrange samples into groups based on their expression levels. The findings were illustrated with a volcano plot and the hierarchical clustering of these altered lncRNAs and mRNAs.
Gene ontology analysis and pathway analysis
Gene ontology (GO) analysis is a functional analysis of the differentially expressed mRNAs that is performed by connecting these mRNAs with the GO categories that are derived from Gene Ontology (
LncRNA-mRNA co-expression network analysis
A lncRNA-mRNA co-expression network was constructed according to the normalized signal intensity of these aberrantly expressed lncRNAs and mRNAs. 18 The Pearson correlation coefficients of each dysregulated lncRNA-mRNA, lncRNA-lncRNA, and mRNA-mRNA pair were calculated. Pearson correlation coefficients range from −1.00 to 1.00, and larger absolute values of the Pearson correlation coefficient indicate a stronger interaction. In the present study, the pairs with Pearson correlation coefficients >0.95 or < −0.95 and with a p value <0.05 were chosen to build the co-expression network. In this network, the number of links from each lncRNA to mRNA or to another lncRNA was calculated and defined as the degree of centrality. 19 A higher degree for an lncRNA meant that the lncRNA played a more important role in this network.
Quantitative RT-PCR validation
Quantitative RT-PCR was used to validate the lncRNA expression level from microarray data as described above. In brief, the brain tissue samples from two groups (n = 6 each, including the samples for microarray examination) were lysed using TRIzol reagent (Invitrogen), and then the total RNA was extracted per the manufacturer's protocol. Reverse transcription to cDNA was achieved using a PrimeScriptTM RT Reagent Kit (Perfect Real Time, Takara, China). Quantitative RT-PCR was performed using SYBR Premix Ex Taq II (Tli RNaseH Plus) (Takara, China) in a Thermal Cycler Dice Real Time System II. The primer sequences used for quantitative RT-PCR were as follows: rat Zfas1: 5′-GCT GGT GTA AAT GAT GAC CTC-3′ (forward), 5′-GCC GGT GTT ATC AGA ACT AAC-3′ (reverse); rat Bsr: 5′-CTG ACC AGT GAT GAG AAT TC-3′ (forward), 5′-CTG GAC CTC AGA GTT ACA G-3′ (reverse); rat GAS5: 5′-AGC CCA TGA TGG TAT GAG-3′ (forward), 5′-AGC CTC AGA ACA GTG TT-3′ (reverse); and rat SNHG6: 5′-GCT GGC ACA ATG ATG ACT TAT-3′ (forward), 5′-CTG GCA GCT CAG TCT TAA G-3′ (reverse). All experiments were performed in triplicate, and the target lncRNA expression levels were normalized to their internal control, β-actin. The Ct value for each sample was calculated using the ΔΔCt method, and the results were expressed as 2−ΔΔCT.
Statistical analysis
All statistical analyses were performed using SPSS 16.0 statistics software. Data were presented as the mean ± standard deviation of at least three independent experiments. The statistical significance of differences between the groups was determined using Student's t-test. A p value <0.05 indicated a statistically significant difference.
Results
Differentially expressed lncRNAs in the hippocampus after TBI
The inclusion of differentially expressed lncRNAs that met the criteria of a corrected Q-value <0.25 and absolute fold change >1.2 led us to identify 271 upregulated and no downregulated lncRNA probe sets. We eliminated 55 microRNA or partial cds of mRNA probe sets from the array data, such as NR_031878 and FQ228770 (Fig. 1A; Supplementary Table 1; see online supplementary material at
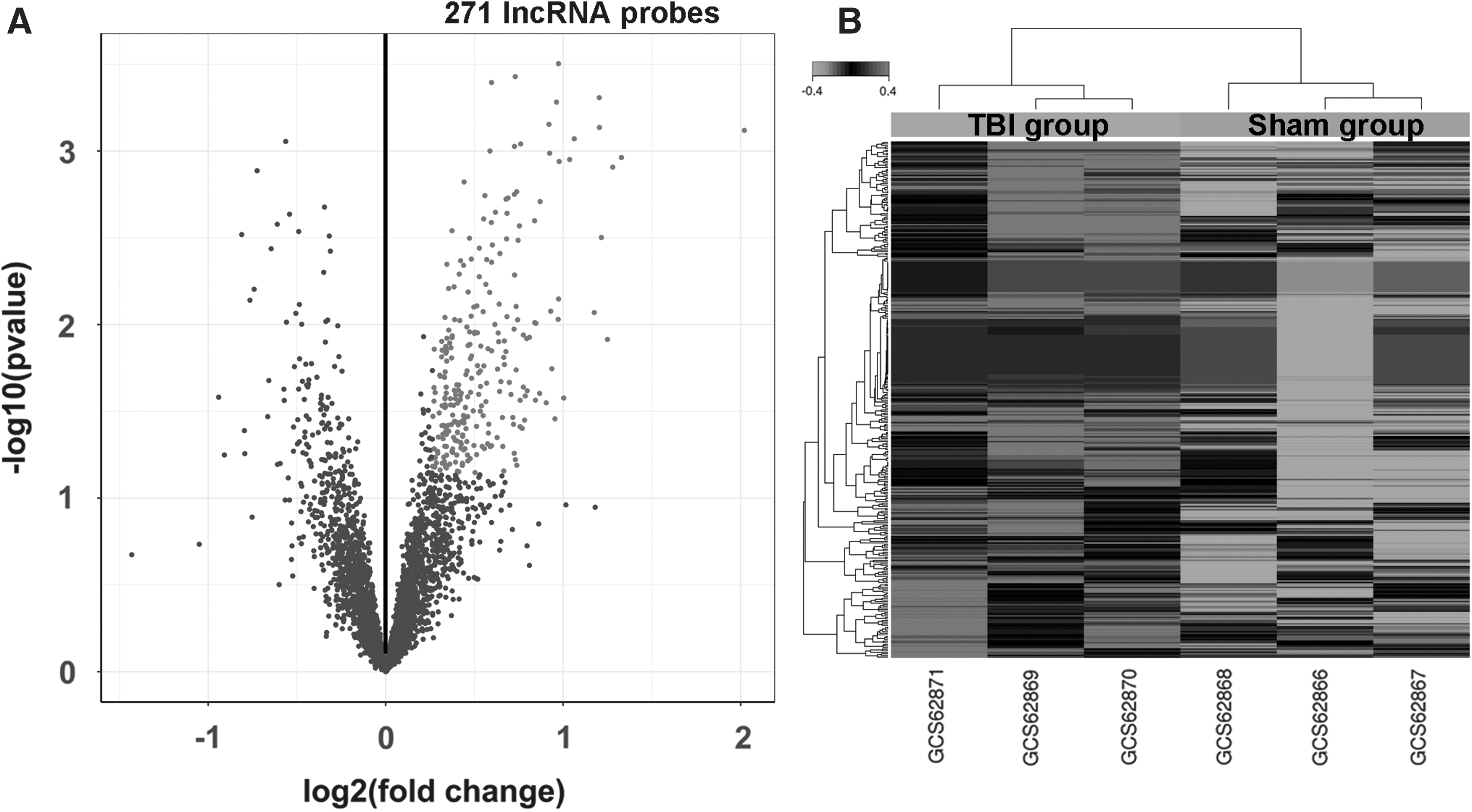
Identification of rat hippocampal long non-coding RNAs (lncRNAs) that are differentially expressed between the TBI and sham groups.
mRNA, messenger RNA; lncRNA, long non-coding RNA.
Differentially expressed mRNAs in the hippocampus after TBI
After moderate fluid percussion brain injury, a total of 756 upregulated and 290 downregulated mRNA probe sets were identified (Fig. 2A; Supplementary Table 2; see online supplementary material at

Identification of rat hippocampal messenger RNAs (mRNAs) that are differentially expressed between the TBI and sham groups.
GO analysis and pathway analysis
When the differentially expressed mRNAs were subjected to GO analysis, three integrated networks were identified. In the molecular function analysis, most of the top 10 categories involved DNA binding and protein binding (Fig. 3A, Supplementary Table 3; see online supplementary material at

Gene ontology (GO) and pathway analysis of long non-coding RNA co-expressed messenger RNAs. The significance of the top 10 GO categories and pathways was calculated and plotted as the -log10(p-value).
Pathway analysis predicted the pathways affected by the aberrantly expressed lncRNAs after TBI. The top three pathways were the mitogen-activated protein kinase (MAPK) signaling pathway, cytokine-cytokine receptor interaction and the nuclear factor κB signaling pathway (Fig. 3D; Supplementary Table 3). Next, the pathway relation network was established based on pathway analysis (Fig. 4; Supplementary Table 3). The main downstream pathways involved the cell cycle, inflammation and apoptosis; this result was consistent with the pathway analysis.
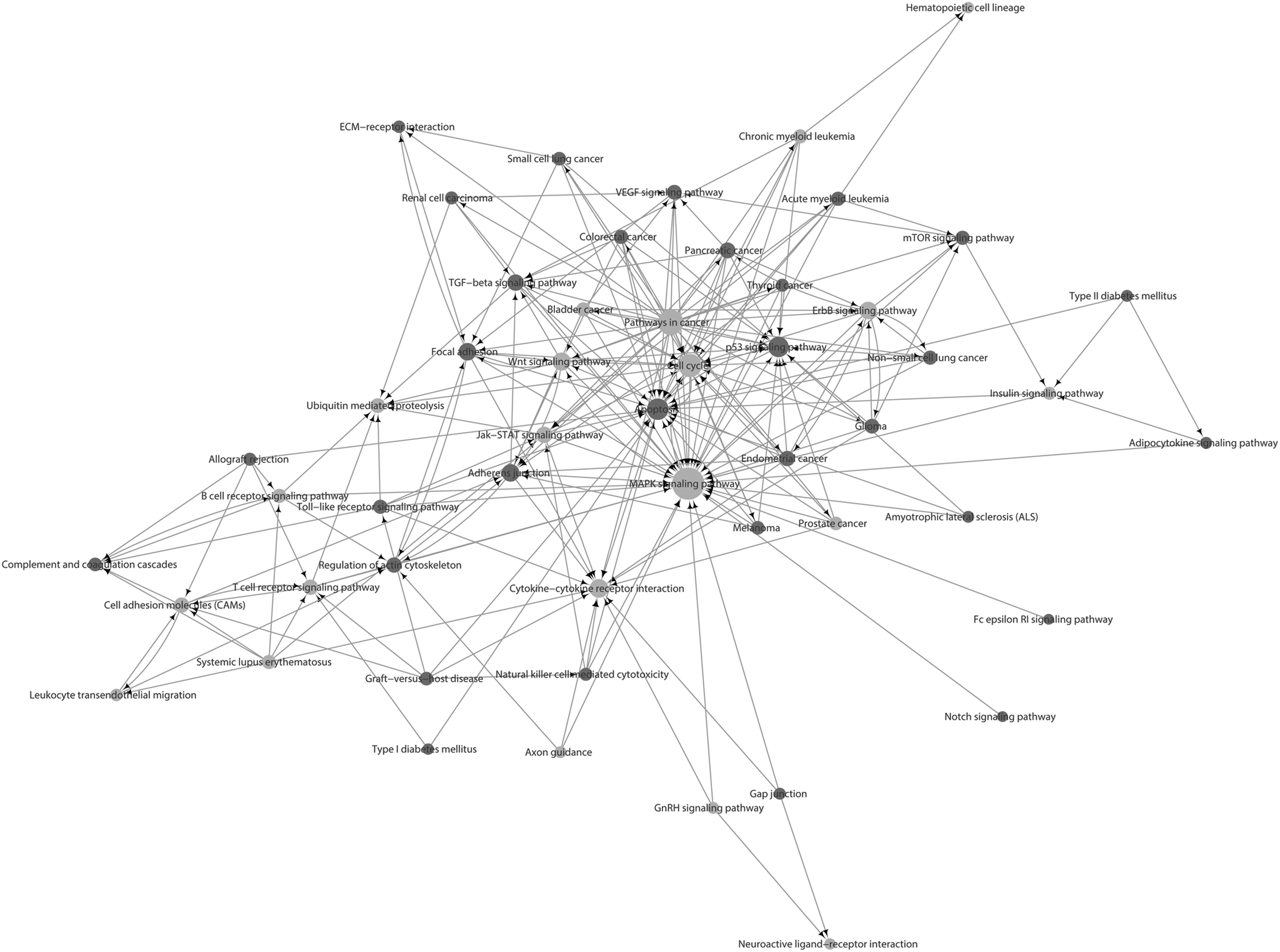
Pathway relation network analysis. The pathway relation network was established based on the result of pathway analysis and a Kyoto Encyclopedia of Genes and Genomes database search. The pathways that contained upregulated genes are shown as red dots. The pathways that contained both upregulated and downregulated genes are shown as yellow dots. The dot size represents the degree of centrality. The arrows point from the upstream pathways to the downstream pathways.
LncRNA and mRNA co-expression network analysis
Co-expression network analysis was performed for these 271 differentially expressed lncRNAs and their co-expressed mRNAs. The entire network consisted of 667 nodes and 2286 connections (Supplementary Fig. 1; see online supplementary material at
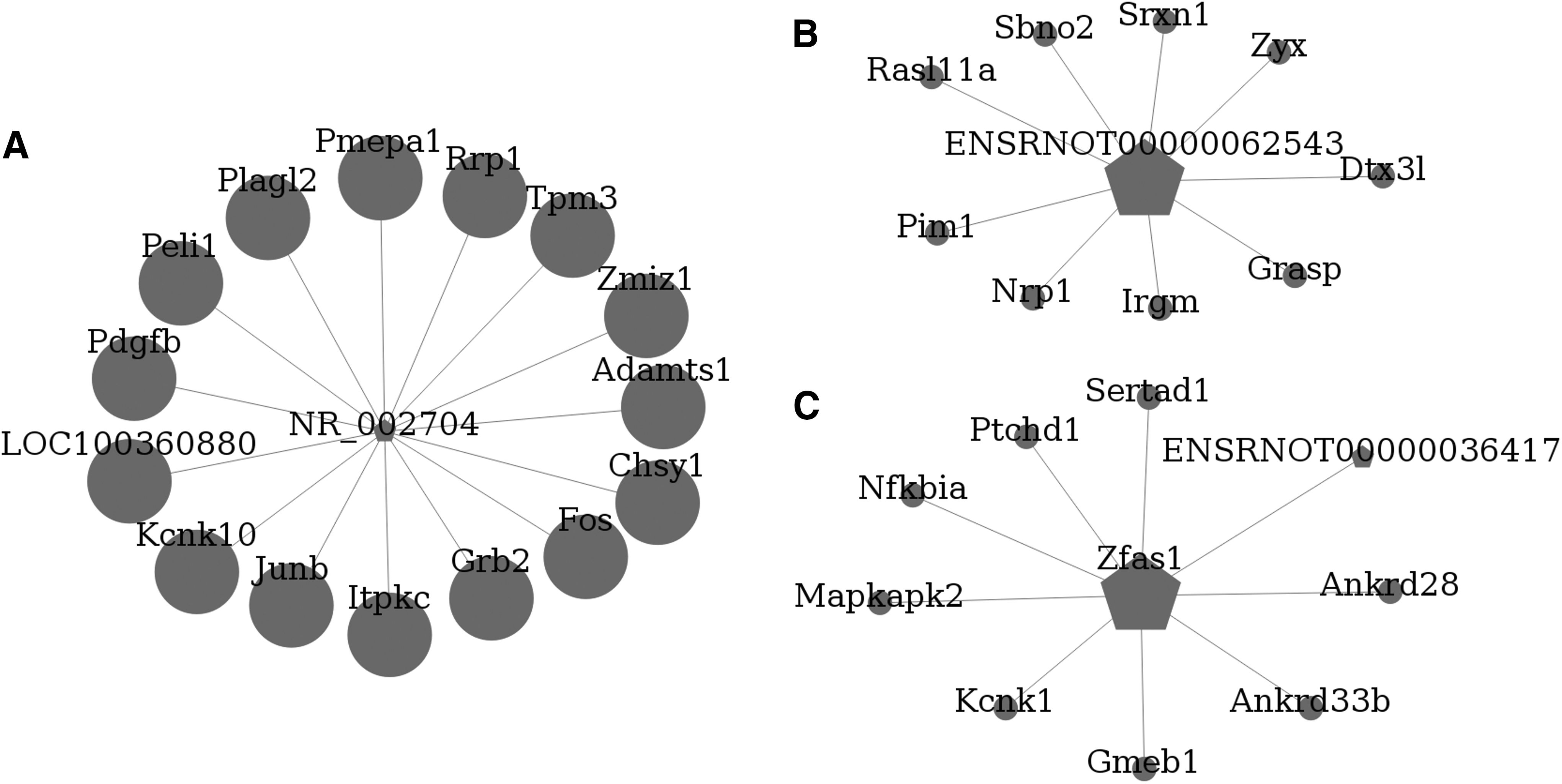
Long non-coding RNA (lncRNA) and messenger RNA (mRNA) co-expression network analysis. The lncRNA-mRNA co-expression sub-network consisted of the three lncRNAs with the highest degree of centrality,
Quantitative RT-PCR validation
Four differentially expressed lncRNAs (Zfas1, Bsr, GAS5, and SNHG6) were randomly selected for validation. The expression levels of these lncRNAs are shown in Figure 6. Consistent with the microarray analysis, the results demonstrated that all four lncRNAs were upregulated in the TBI group, compared with the sham group.
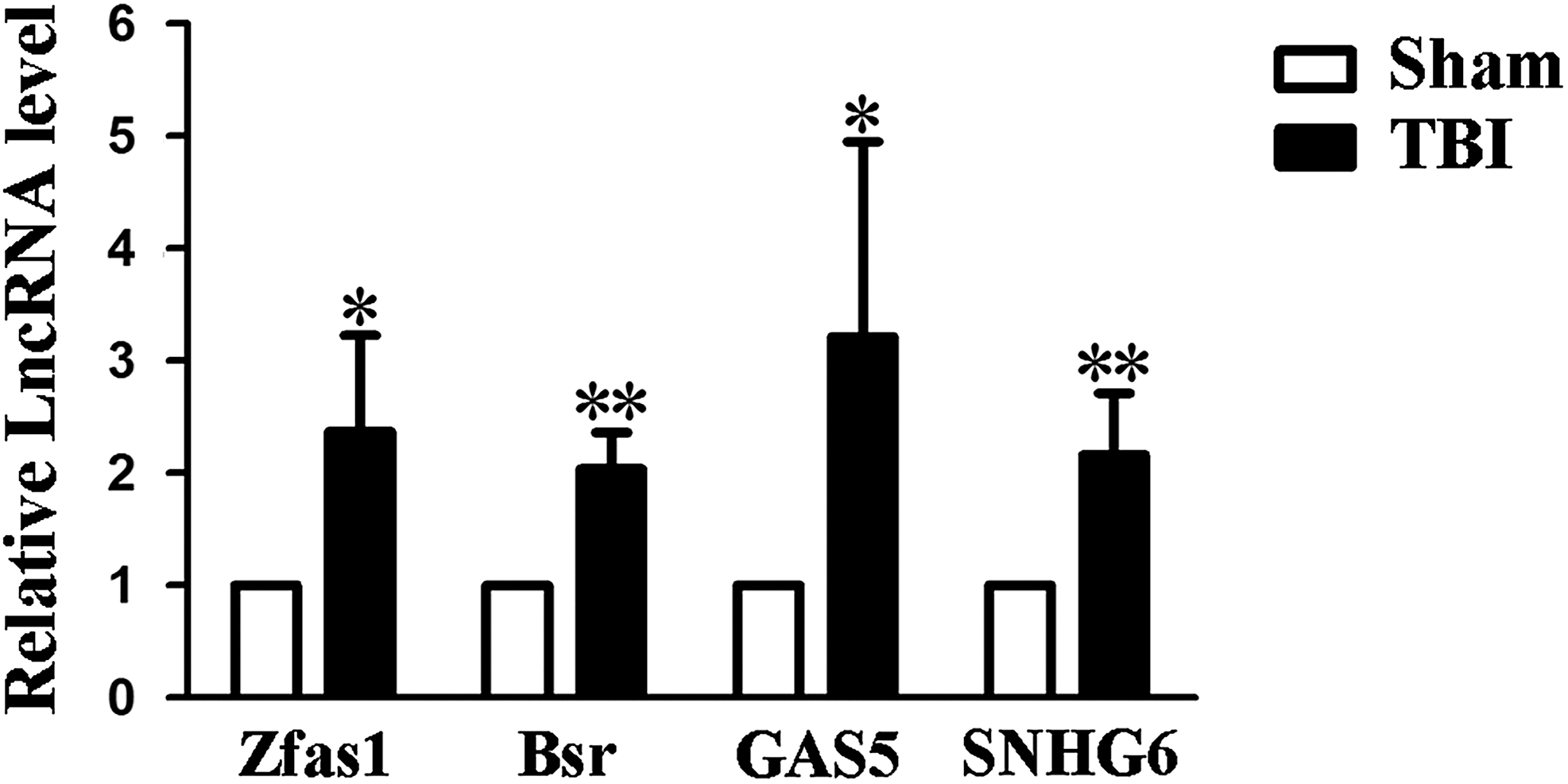
Quantitative reverse transcription polymerase chain reaction (qRT-PCR) validation results of four selected long non-coding RNAs (lncRNAs). The expression level of four selected lncRNAs (Zfas1, Bsr, GAS5, and SNHG6) was validated by qRT-PCR in 12 samples. The target lncRNA expression levels were normalized to their internal control, β-actin. Data are presented as the mean ± standard deviation (n = 6). * p < 0.05, **p < 0.01 significantly different than sham group, Student's t-test (two-tailed).
Discussion
In the present study, we investigated the altered expression profile of lncRNAs after TBI in the rat hippocampus using microarray analysis for the first time, and we predicted their potential roles in the early stage after TBI. In total, 271 lncRNA probe sets and 1046 mRNAs probe sets were differentially expressed following TBI. With further bioinformatics analysis, we achieved novel findings, including the identification of the most significantly changed GO categories, most correlated pathways, core pathways, and specific interaction molecular network for the three lncRNAs with the highest degree of centrality. The expression level of four randomly selected lncRNAs—Zfas1, Bsr, GAS5 and SNHG6—was validated by quantitative RT-PCR, which confirmed the reliability of the microarray results.
A previous study showed that the transcription of lncRNAs might affect the expression of their co-expressed mRNAs at the level of chromatin modification, transcription, and post-transcriptional processing. 20 To gain insight into the potential functions of the aberrantly expressed lncRNAs in the rat hippocampus after TBI, GO and pathway analyses of their co-expressed mRNAs were performed. The most significantly altered categories identified by GO analysis were inflammation, DNA transcription, apoptosis, and necroptosis. These pathological processes are major causes of secondary injury in the early stage after TBI. 21 –23 Thus, we concluded that these aberrantly expressed lncRNAs might play an important role in regulating secondary injury after TBI.
The most highly correlated pathways we identified were mainly involved with inflammation and apoptosis, 24 –26 suggesting that the aberrantly expressed lncRNAs might regulate secondary injury after TBI through effects on these pathways. Based on pathway analysis, we constructed a pathway relation network. The network indicated that the most important core downstream pathways involved the MAPK signaling pathway, apoptosis, cell cycle, p53 signaling pathway, and cytokine-cytokine receptor interaction (Table 1; in degrees were 35, 24, 20, 16, and 15, respectively); thus, the final functions of these aberrantly expressed lncRNAs might be achieved through these downstream pathways.
In a recent study, pathological axonal death after injury was associated with the activation of a MAPK cascade, 27 whereas suppressing the MAPK/nuclear factor κB signaling pathways could attenuate the inflammatory response in TBI and neurodegenerative diseases. 24,28 Moreover, the MAPK pathway mediated traumatic injury–induced apoptosis in astrocytes. 29 In the present study, our data showed that after TBI, the pathways predicted to be downstream of MAPK signaling were cell cycle, apoptosis, Wnt signaling pathway, and p53 signaling pathway (Table 1).
Cell cycle molecules are thought to act as apoptotic mediators for post-mitotic cells that are under stress due to trauma or neurological disease. 30,31 Re-activation of these cell cycle molecules, such as cell cycle–related genes (cyclins A1, A2, D1, E2F1, and PCNA) and proteins (cyclin D1 and CDK4), is observed in dying neurons and contributes to neuronal apoptosis and degeneration in the adult brain. 32,33 Moreover, several studies have found that neuronal damage can be attenuated and functional recovery can be facilitated by treatments with cell cycle inhibitors or selective cyclin-dependent kinase inhibitors. 33,34 In the present study, the pathways that were predicted to be downstream of the post-TBI effects on cell cycle were apoptosis, ubiquitin-mediated proteolysis, and MAPK signaling pathway (Table 1).
The p53 signaling pathway acts as an internal sentinel for cellular stresses, including oncogene activation, hypoxia, or TBI. 35,36 The p53 signaling pathway is associated with cell death, apoptosis, cell cycle arrest, subsequent DNA repair, axonal outgrowth, and neurogenesis after TBI. 37,38 In the present study, the pathways that were predicted to be downstream of the post-TBI effects on the p53 signaling pathway were apoptosis and cell cycle (Table 1).
After TBI, the primary impact initiates cytokine secretion, which leads to activation of brain-resident astrocytes and microglia and recruitment of peripheral blood immune cells to the TBI brain. 39 This inflammatory reaction is one of the main driving forces of secondary injury after TBI. 40 Our data showed that several cytokines and cytokine receptors and their predicted associated lncRNAs might be altered after TBI (Table 1).
From these findings, we knew that most of these aberrantly expressed lncRNAs might participate in secondary injury after TBI by modulating their co-expressed mRNAs, which needs further study. To further investigate the complicated lncRNA and mRNA regulation network and predict potential targets for therapeutic intervention, we performed lncRNA and mRNA co-expression network analysis. The top 10 lncRNAs with the highest degree within the network were NR_002704, ENSRNOT00000062543, Zfas1, ENSRNOT00000053390, ENSRNOT00000036417, ENSRNOT00000052658, ENSRNOT00000054074, ENSRNOT00000053006, ENSRNOT00000062784, and ENSRNOT00000069930, which were identified for the first time in injured brain. The top three lncRNAs and their co-expressed mRNAs were analyzed in detail.
NR_002704 is also known as growth arrest specific 5 (Gas5) and it has mainly been studied as a tumor suppressor regulator in cancer. 41 Several studies have found that Gas5 is involved in the control of cell apoptosis, proliferation, metastasis, angiogenesis, DNA repair, and tumor cell metabolism. 42,43 However, its roles in the early stage after TBI are still unknown. Our data indicated that Gas5 had 15 co-expressed mRNAs that were all upregulated in the early stage after TBI. Among them, a disintegrin and metalloproteinase with thrombospondin motifs 1 (adamts1) is reported to be responsible for the cleavage of chondroitin sulfate proteoglycans (CSPGs), 44 which are the main component of the glial scars. 45 Upregulation of adamts1 may prevent glial scar formation and promote neurite outgrowth and remodeling. The result was consistent with the finding that the maximal spontaneous cortical sprouting occurred in pericontusion regions in the early stage after TBI and accompanied the reduction in extracellular CSPGs. 46 However, that study also mentioned a lack of neurite outgrowth by 28 days after TBI. Whether the extrinsic growth capacity is regulated by Gas5 through adamts1 or Gas5 is downregulated at a later stage after TBI deserves further exploration.
ENSRNOT00000062543 is an lncRNA that has never been studied. Our data identified a total of nine co-expressed mRNAs. Neuropilin 1 (Nrp1) is one of these mRNAs that has been determined to be a regulator that modulates dendrite polarization and axon guidance. 47,48 The possibility that ENSRNOT00000062543 plays a role in axon plasticity after TBI by regulating Nrp1 expression requires further investigation.
Zfas1 functions as an oncogene in several cancers, 49,50 but its potential roles in the early stage after TBI have not been elucidated. Our data indicated that Zfas1 had nine co-expressed mRNAs. Among them, NFKBIA is reported to be a nuclear factor κB pathway inhibitor. 51 Upregulation of NFKBIA may play a role in the attenuation of central nervous system inflammation in the early stage after TBI. Another mRNA, GMEB1, was found to be an endogenous regulator that selectively binds to initiator procaspases and inhibits caspase-induced apoptosis. 52 The possibility that elevated Zfas1 restricts the inflammation and apoptosis reaction after TBI by regulating NFKBIA and GMEB1 expression requires further investigation.
In conclusion, our study identified a series of dysregulated lncRNAs that may be related to the physiological and pathological processes that occur after TBI. We also predicted the potential roles of these aberrantly expressed lncRNAs and the underlying molecular network. The findings may provide novel targets for further investigation of both the molecular mechanisms of TBI and potential therapeutic interventions.
Footnotes
Acknowledgments
We would like to thank Genminix Informatics and Zhi-han Sun for their excellent technical assistance with the data analysis. We also thank Dr. Lei Wang and other members of the Shanghai Institute of Head Trauma for assistance. We thank Dr. Bruce G. Lyeth, Department of Neurological Surgery, University of California Davis, for his critical English language editing. We gratefully acknowledge the following funding sources: the National Natural Science Foundation of China (30901543, 31371406, 81671198, and 81571883) and the National Key Basic Research Program (2012CB518100). JFF also received funding from the Science and Technology Commission of Shanghai Municipality (13ZR1424500), Shanghai Municipal Health Bureau (XYQ2013094) and SMC-Star Award for Young Scholars (B).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.