
Abstract
The present study was designed to establish a midcervical contusion model that can simulate long-term respiratory deficits, and investigate the breathing pattern during vagal-mediated respiratory reflexes following midcervical contusion. Moderate and severe (impactor height: 6.25 or 12.5 mm) contusion was induced at midline C3-4 spinal cord in adult Sprague–Dawley rats. The ventilatory behaviors of unanesthetized were evaluated by whole body plethysmography at 1 day and 1, 2, 4, and 8 weeks post-injury. The tidal volume was decreased and frequency was increased in contused animals compared with uninjured animals at the acute injury state. At 8 weeks post-injury, respiratory frequency was similar between groups; however, contused animals had lower tidal volume. The pulmonary chemoreflex induced by intrajugular capsaicin (1.5 μg/kg) injection and the Hering–Breuer inflation reflex induced by increasing positive end-expired pressure (9 cm H2O) were evoked in anesthetized animals at 3 days, or 2 or 8 weeks post-surgery. The result showed that capsaicin induced a significant prolongation of the expiratory duration in uninjured animals; however, this response was greatly attenuated in contused animals at 3 days post-injury. Increased positive end-expired pressure also caused an increase in the expiratory duration in uninjured and moderately contused animals; however, severely contused animals exhibited an attenuated response. At 2 and 8 weeks post-injury, both the pulmonary chemoreflex and the Hering–Breuer inflation reflex were similar between uninjured and contused animals. These data suggested that midcervical contusion can cause a long-term respiratory impairment and a transiently attenuation of vagal-mediated respiratory reflexes.
Introduction
I
Lung vagal afferents primarily divide into slowly adapting pulmonary stretch receptors, rapidly adapting receptors, and bronchopulmonary C fibers. These afferents can detect mechanical and/or chemical stimuli in the lung under physiological and pathological conditions and then trigger regulatory (e.g., Hering–Breuer reflex) and/or defensive (e.g., pulmonary chemoreflex) reflexes. 30 –32 The functional role of lung vagal afferents on the respiratory function following spinal cord injury has been studied by our and other groups. Golder and coworkers showed that unilateral hemisection at the second cervical spinal cord (i.e., C2 hemisection) reduced the tidal volume and increased the respiratory frequency; however, this rapid shallow breathing pattern was not observed after bilateral vagotomy. 22 Our recent studies indicated that pulmonary chemoreflex activated by bronchopulmonary C fibers was attenuated during both acute and chronic injury state following C2 hemisection. 13,21,33 In addition, lung inflation-induced reflex was also blunted at 2 and 8 weeks post-C2 hemisection. 34 These data suggested that basal vagal inputs have a substantial impact on spinal cord injury-induced respiratory deficiency, and that lung vagal-medicated respiratory reflexes were altered after cervical spinal cord injury. However, the abovementioned experiments were conducted in the C2 hemisection injury model, which interrupts the bulbospinal respiratory pathway but preserves the intact spinal respiratory motoneurons. The functional role of vagal afferents on the breathing pattern in the midcervical contusion model, which not only influence the bulbospinal respiratory pathway but also directly damage the phrenic motoneuron, remains unclear. Accordingly, the second aim of the present study was to investigate the ventilator responses of lung vagal-mediated respiratory reflexes of contused animals after injury.
Methods
Animals
A total of 145 adult male Sprague–Dawley rats purchased from BioLASCO Taiwan Co., Ltd received C3-C4 laminectomy surgery (i.e., uninjured group, n = 36) or cervical contusion at midline C3-C4 spinal cord (n = 109). Two injury severities were made by using different heights of the impact rod (severe contusion [12.5 mm], n = 71; moderate contusion [6.25 mm] n = 38). All experimental procedures were approved by the Institutional Animal Care and Use Committee at National Sun Yat-sen University.
Spinal cord contusion surgery
All rats were anesthetized using xylazine (10 mg/kg, s.c., Rompun®, Bayer) and ketamine (140 mg/kg, i.p., Ketalar®, Pfizer) at 9–10 weeks of age. An incision of dorsal skin and muscle was made after absence of a toe-pitch withdrawal reflex. The C3-C4 vertebral bones were immobilized by the Cunningham spinal adaptor (Stoelting Co.) which was mounted on a stereotaxic instrument (David Kopf Instruments). The animal was then placed on a stage of the Multicenter Animal Spinal Cord Injury Study (MASCIS) Impactor System (Model II, W.M. Keck Center for Collaborative Neuroscience) followed by C3-C4 laminectomy. The contusion injury was performed by releasing the impact rod (weight: 10 g; diameter: 2 mm) at the height of 6.25 mm or 12.5 mm above the surface of midline C3-C4 spinal cord. If animals stopped breathing after contusion, the animal would be mechanically ventilated by the rodent ventilator (Harvard Apparatus) until spontaneously breathing was resumed. The overlying muscle and skin were sutured with 4-0 chromic (UNIK) and 4-0 nylon (UNIK) sutures, respectively. Animals were then received subcutaneous injections of yohimbine (1.2 mg/kg, Tocris) to reverse the action of xylazine, and lactated Ringer's solution (5 mL, Nang Kuang Pharmaceutical Co., Ltd) to prevent dehydration. A single bolus injection of buprenorphine (0.03 mg/kg, Shinlin Sinseng Pharmaceutical Co., Ltd) was used for analgesia after animals woke up. Postoperative care protocols, including oral administration of Nutri-cal (1–3 mL, EVSCO Pharmaceuticals) and lactated Ringer's solution injection (5 mL, s.c.), were performed daily until recovery of volitional eating and drinking. One moderately contused and 34 severely contused animals did not survive at 1 day following injury. These animals were excluded from the further experiments.
Breathing patterns in unanesthetized animals
The breathing pattern (e.g., respiratory frequency, tidal volume, and minute ventilation) of unanesthetized and unrestrained animals (uninjured, n = 10; severe contusion [12.5 mm], n = 11; moderate contusion [6.25 mm] n = 11) were measured at 1 day and 1, 2, 4, and 8 weeks after spinal cord surgery using the whole body plethysmography system (Buxco® FinePointe Whole Body Plethysmography, Data Sciences International). The temperature, humidity, and airflow of the chamber (#PLY4213, volume: 3.9 L) were calibrated by the standard procedure indicated by the manual (Buxco FinePointe Whole Body Plethysmography, Data Sciences International). The body temperature of the animal was recorded and used to calibrate the breathing pattern parameters. The animal was then placed in the chamber and exposed to 60 min normoxic gas (21% O2, 79% N2) by flushing compressed gas mixtures (2.5 L/min) into the chamber. The animals used for the whole body plethysmography measurement were also used to monitor the cardiorespiratory patterns under the anesthetized condition at 8–9 weeks post-injury (discussed subsequently).
Cardiorespiratory patterns in anesthetized animals
The cardiorespiratory pattern were measured at 3 days (uninjured, n = 8; moderate contusion [6.25 mm] n = 8; severe contusion [12.5 mm], n = 8), 2 weeks (15.2 ± 0.3 days) (uninjured, n = 10; moderate contusion [6.25 mm], n = 10; severe contusion [12.5 mm], n = 10) and 8 weeks (59.2 ± 0.6 days) (uninjured, n = 10, moderate contusion [6.25 mm], n = 11; severe contusion [12.5 mm] n = 11) after spinal cord surgery. On the day of the experiment, the animal was anesthetized with urethane (1.6 g/kg, i.p., Sigma) and placed in a supine position after an absence of the toe-pitch withdrawal reflex. The rectal temperature was monitored by a rat temperature probe and maintained at 37 ± 1°C by a temperature controller (model TC-1000, CWE Inc.). An endotracheal tube (PE-240, Clay Adams) was inserted into the trachea below the larynx and connected to a respiratory flow head (MLT1L, ADInstruments) coupled with a spirometer (FE141, ADInstruments) for respiratory flow measurement. The femoral artery (PE-50) and right jugular vein were catheterized (PE-100, Clay Adams) for blood pressure monitoring (Transducer: DTX-1; Amplifier: BPM-832, CWE Inc.) and drug administration, respectively. A 26 gauge needle with a PE-50 tube was inserted in the endotracheal tube to detect the tracheal pressure by connecting the pressure monitor system (Transducer: DTX-1; Amplifier: BPM-832, CWE Inc.). A two way non-rebreathing valve (Model 2300, Hans Rudolph, Inc.) was connected to the flow head to regulate the end-expired pressure (see Lung vagal-mediated reflexes). A hyperoxic gas (50% O2, balance N2) was introduced to the animal through the inspiratory port of the two way valve. All physiological signals (e.g., respiratory airflow, tracheal pressure, and blood pressure) were digitized using CED Power 1401 (Cambridge Electronic Design Limited) and the PowerLab data acquisition system (ADInstruments) at a sampling rate of 1000 Hz, and recorded in a computer by Spike 2 (Cambridge Electronic Design Limited) and LabChart software.
Lung vagal-mediated reflexes
After a stable recording of the cardiorespiratory pattern under anesthetized conditions, two different approaches (i.e., lung inflation and intrajugular capsaicin administration) were used to trigger lung vagal-mediated reflexes. 1) Lung inflation was induced by inserting the outlet tube of the two way valve into a graduated cylinder of water at a depth of 9 cm for 10 sec. This approach can modulate the activity of mechanical-sensitive vagal afferent and evoke the Hering–Breuer inflation reflex by increasing the end-expired pressure. 34,35 2) A single bolus intrajugular injection of capsaicin (1.5 μg/kg) was used to evoke the pulmonary chemoreflex through activation of lung chemical-sensitive afferents. 13,21,33,36 These two approaches were randomly applied to the animal, and an interval of 10–20 min elapsed to prevent interaction between two stimuli. Bilateral vagus nerves were separated from the carotid artery at the cervical region and then sectioned at 10 min after the last stimuli. Both lung inflation and capsaicin administration were applied again under vagotomized conditions to confirm that the effects of capsaicin and lung inflation were mediated by vagal afferents. The arterial blood parameters (partial pressure of arterial oxygen [PaO2], partial pressure of carbon dioxide [PaCO2], and pH) were measured by the blood analyzer (i-STAT, Abbot) during the baseline under vagal-intact and vagotomized conditions.
Spinal cord histology
After termination of physiological measurements under anesthesia, the animals were systemically perfused with heparin-saline followed by 4% paraformaldehyde (Alfa Aesar) and then 10% sucrose (Sigma) in 4% paraformaldehyde. The C2-C5 spinal cord was dissected and placed in 30% sucrose in phosphate buffered saline (UniRegion Bio-Tech) for dehydration. After the spinal cord sank, the spinal cord was cryoprotected and transversely sectioned into 40 μm slices (CM 1850, Leica) and mounted on glass slides (S7441, Matsunami). The spinal cord slices were stained with cresyl violet (Acros Organics) and imaged using a microscope (BX43, Olympus) connected to a charge-coupled device (CCD, DS-5Mc, Nikon). The lesion epicenter was defined as the section with the largest lesion cavity relative to total spinal cord area in the same section. The area of the spinal cord section and lesion empty cavity was manually outlined and quantified using ImageJ software every five sections rostral and caudal to the lesion epicenter. The lesion area was presented as a percentage of spinal cord section area.
Phrenic motoneuron labeling
A subset of animals (uninjured, n = 8; severe contusion [12.5 mm], n = 8; moderate contusion [6.25 mm] n = 8) were used to examine the impact of cervical contusion on the number of phrenic motoneurons. At 8 weeks post-injury, animals were anesthetized with xylazine (10 mg/kg, s.c.) and ketamine (140 mg/kg, i.p.). A laparotomy was performed and the abdominal muscle was retracted to expose the diaphragm, and 40 μL of monosynapic retrograde tracer (cholera toxin subunit B conjugated with Alexa Fluor 594 [#C34777, ThermoFisher Scientific]) was painted on the bilateral diaphragm. 37,38 Animals were anesthetized with urethane (1.6 g/kg, i.p.) at 72 h (72.2 ± 0.5 h) after tracer application. The procedure of perfusion and spinal cord tissue preparation was similar as described in the Spinal cord histology section. The C2-C5 spinal cord was horizontally sectioned into 40 μm slices (CM 1850, Leica), mounted on glass slides (S7441, Matsunami glass, Ind., Ltd.) and cover-slipped with mounting medium containing 4',6-diamidino-2-phenylindole (DAPI, Sigma). The tracer-labeled phrenic motoneurons were observed by a microscope (BX43, Olympus) with a fluorescence filter cube (U-FMCHE, Olympus). The image was taken by CCD (DS-5Mc, Nikon) and the number of tracer-labeled phrenic motoneurons was counted manually.
Data analysis and statistics
The breathing pattern (e.g., respiratory frequency, tidal volume, and minute ventilation) of unanesthetized rats were calculated by FinePointe software (Data Sciences International). These data were exported to an Excel file and averaged over 10 min during the normoxic breathing. The cardiorespiratory patterns of anesthetized rats were calculated by Spike 2 scripts. The respiratory cycle duration (Tcycle) was defined as the period between the onset of two successive inspirations. The inspiratory duration (TI) was defined as the period when the respiratory airflow is below zero, and the expiratory duration (TE) was calculated as “Tcycle - TI.” The tidal volume is derived from integration of the inspiratory flow. The mean arterial pressure (MAP) was calculated as “1/3 systolic blood pressure + 2/3 diastolic blood pressure.” The heart rate (HR) was determined as “60/cardiac cycle duration.” The cardiorespiratory patterns were averaged over 10 sec before lung inflation or capsaicin administration as the baseline value (BL). The response to lung inflation was analyzed by averaging data during the period when the lung was inflated by increasing end-expired pressure. The response to capsaicin administration was evaluated by the immediate cardiorespiratory response (e.g., prolongation of TE, bradycardia, and hypotension) after capsaicin treatment. A few animals (uninjured animals at 3 days [n = 1] and 2 weeks [n = 3] post-injury; moderate contused animals at 8 weeks post-injury [n = 1]) exhibited a rapid shallow breathing pattern after capsaicin administration. These animals were excluded from further analysis. 21 The data during and/or following vagal-mediated reflexes were expressed as absolute value and a percentage of the BL.
A two way mixed design analysis of variance (ANOVA) followed by a Student–Newman–Keuls (SNK) post-hoc test was used to analyzed the body weight, the breathing pattern of unanesthetized rats, the intensity of vagal-mediated reflexes of anesthetized rats, and the lesion area. A one way ANOVA followed by a SNK post-hoc test was used to evaluate the baseline cardiorespiratory pattern of anesthetized rats and the number of phrenic motoneurons between animal groups.
All data are expressed as the mean ± standard error of the mean. A p value <0.05 was considered statistically significant.
Results
Body weight
The body weight of animals used for the whole body plethysmography at different time points after spinal surgery is provided in Table 1. The weight was similar between groups at 1 day following spinal surgery, but animals that received severe contusion had a lower body weight than animals that received sham surgery and moderate contusion from 1 to 8 weeks post-surgery (p < 0.05, Table 1). Although contusion injury generally reduced the body weight, the body weight could still gradually increase in both contused groups (p < 0.05, Table 1).
Values are means ± SE. The sample size for each group is 11.
p < 0.05 vs. uninjured; #p < 0.05 significant difference between moderate contusion (6.25 mm) and severe contusion (12.5 mm).
p < 0.05 vs. 1 day; b p < 0.05 vs. 1 day and 1 week; c p < 0.05 vs. 1 day, and 1 and 2 weeks; d p < 0.05 vs. 1 day, and 1, 2, and 4 weeks; e p < 0.05 vs. 1 and 2 weeks.
Breathing patterns in unanesthetized animals
Representative examples of the respiratory airflow of unanesthetized animals recorded by whole body plethymography is shown in Figure 1A. At 1 day post-injury, the respiratory frequency of contused animals (moderate contusion: 138 ± 4 breaths/min; severe contusion: 143 ± 10 breaths/min) was significantly higher than in uninjured animals (113 ± 5 breaths/min) during the normoxic breathing (i.e., baseline) (p < 0.05, Fig. 1B), and the tidal volume of contused animals (moderate contusion: 1.06 ± 0.02 mL; severe contusion: 0.89 ± 0.04 mL) was significantly lower than that of uninjured animals (1.34 ± 0.06 mL) (p < 0.05, Fig. 1B). The breathing frequency of contused animals returned to the normal value at 1 week post-injury and was similar to that of uninjured animals at 2–8 weeks post-injury. Although the tidal volume of contused animals can gradually increase during a subchronic to chronic injury state, the tidal volume of contused animals (moderate contusion: 1.5 ± 0.1 mL; severe contusion: 1.7 ± 0.1 mL) remained lower than that of uninjured animals (2.2 ± 0.1 mL) at 8 weeks post-injury (p < 0.05, Fig. 1B). The minute ventilation of contused animals during normoxic breathing was not significantly different from that of uninjured animals at 1 day and 1 week following injury (Fig. 1B); however, the insufficiency of the minute ventilation was revealed in contused animals at 4 weeks (uninjured: 207 ± 17 mL/min; moderate contusion: 157 ± 7 mL/min; severe contusion: 142 ± 7 mL/min) and 8 weeks post-injury (uninjured: 217 ± 17 mL/min; moderate contusion: 172 ± 10 mL/min; severe contusion: 147 ± 9 mL/min) (p < 0.05, Fig. 1B).
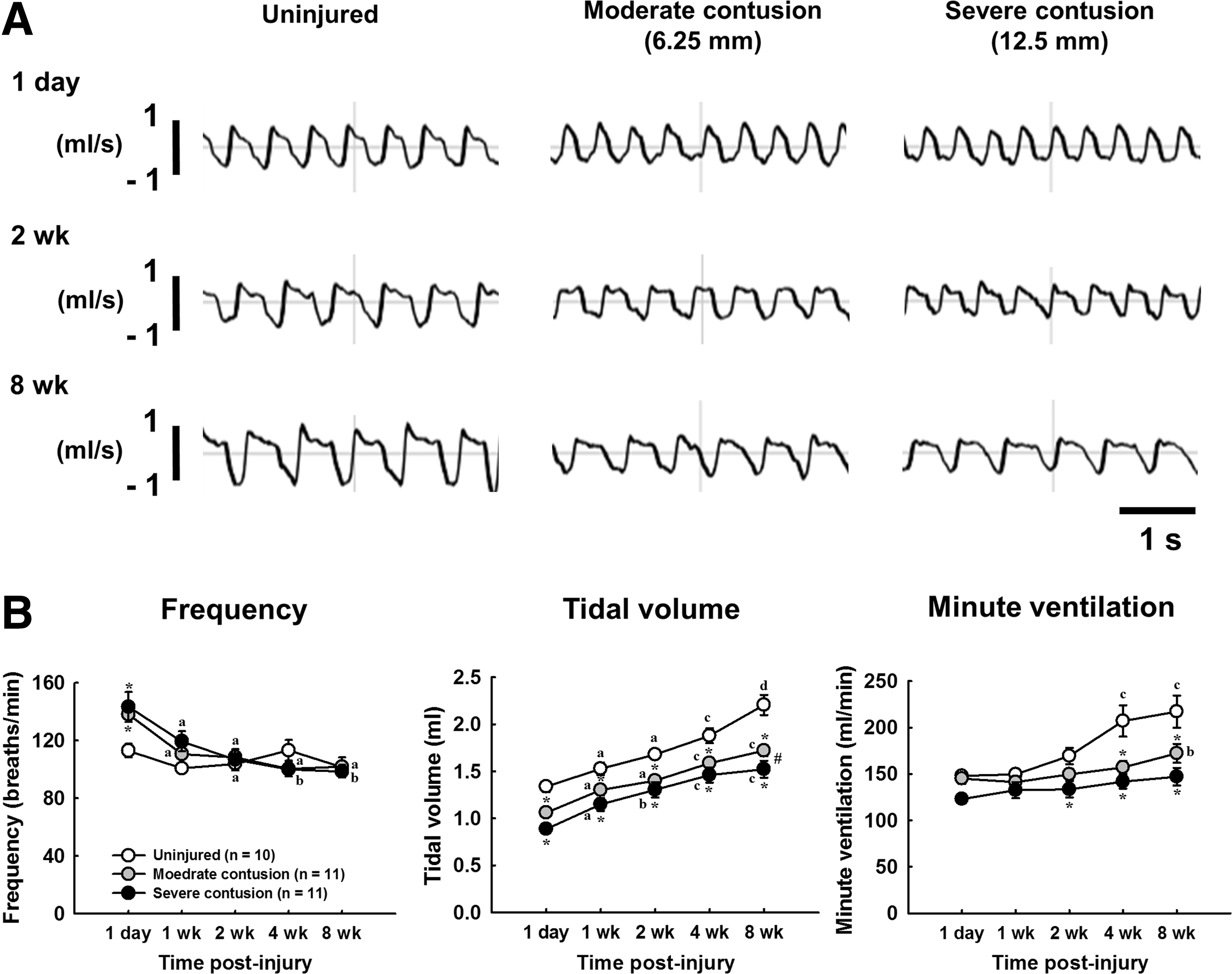
The representative example of respiratory airflow recorded by whole body plethysmography at baseline (21% O2, 79% N2) in uninjured and contused animals at 1 day, 2 weeks, and 8 week post-injury
Cardiorespiratory patterns in anesthetized animals
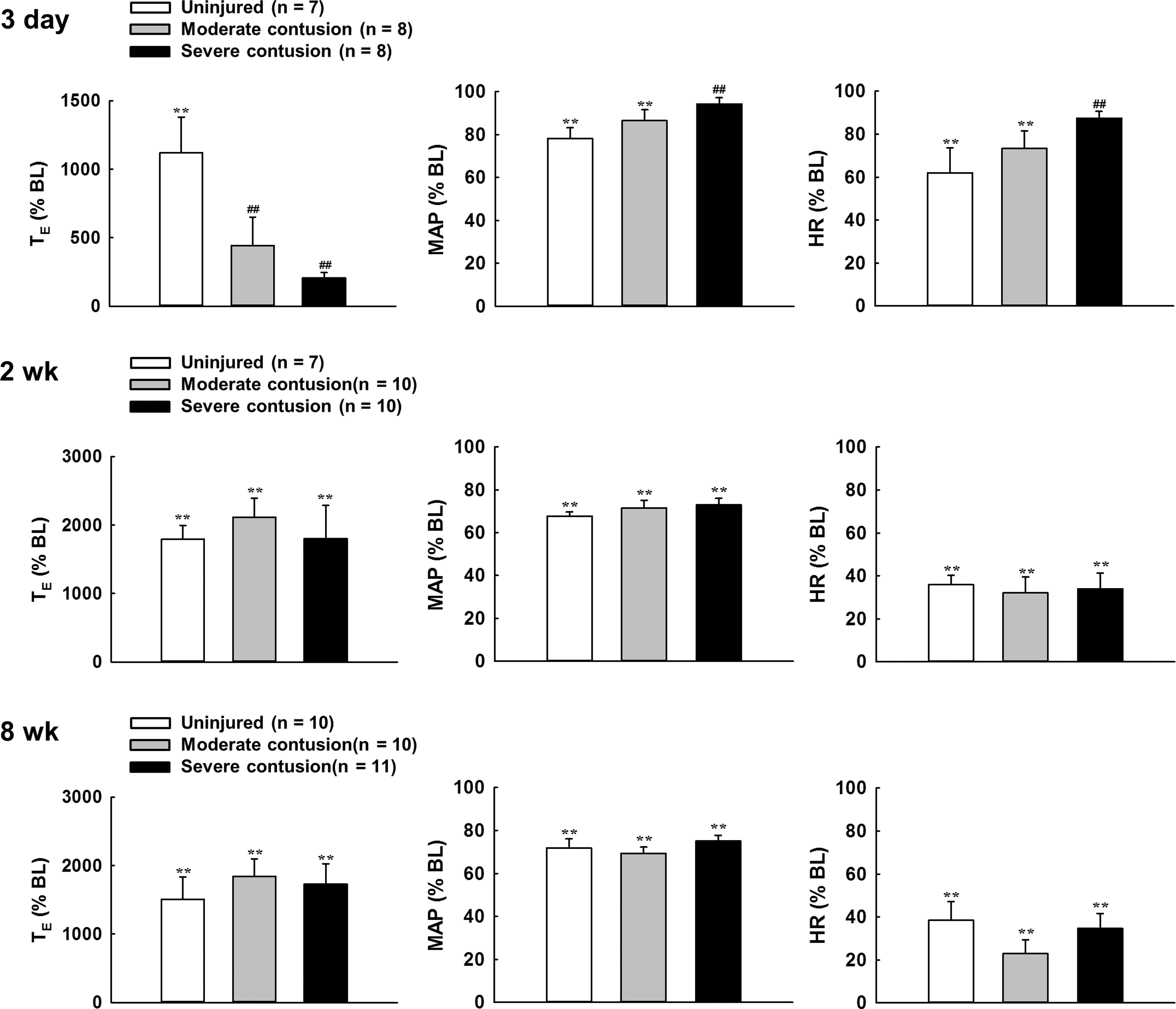
Representative examples of the respiratory airflow recorded in anesthetized animals at 3 days, 2 weeks, and 8 weeks post-injury are presented in Figures 2 and 3. At 3 days post-injury, the minute ventilation was significant lower in both contused animals (moderate contusion: 185 ± 7 mL/min; severe contusion: 132 ± 8 mL/min) than in uninjured animals (218 ± 10 mL/min) (p < 0.05, Table 2). The reduction of the minute ventilation was primarily the result of lower tidal volume in contused animals. Specifically, the tidal volume of both moderately (1.4 ± 0.1 mL) and severely (1.2 ± 0.1 mL) contused animals was significantly lower than that of uninjured animals (1.7 ± 0.1 mL) (p < 0.05, Table 2). The alteration of ventilation following contusion caused a significant change in the blood gas parameters. The PaCO2 of severely contused animals (66 ± 4 mm Hg) was significantly higher than that of uninjured (48 ± 3 mm Hg) and moderately contused (52 ± 3 mm Hg) animals (p < 0.05, Table 3), whereas PaO2 and pH were significantly lower in severely contused animals (p < 0.05, Table 3).
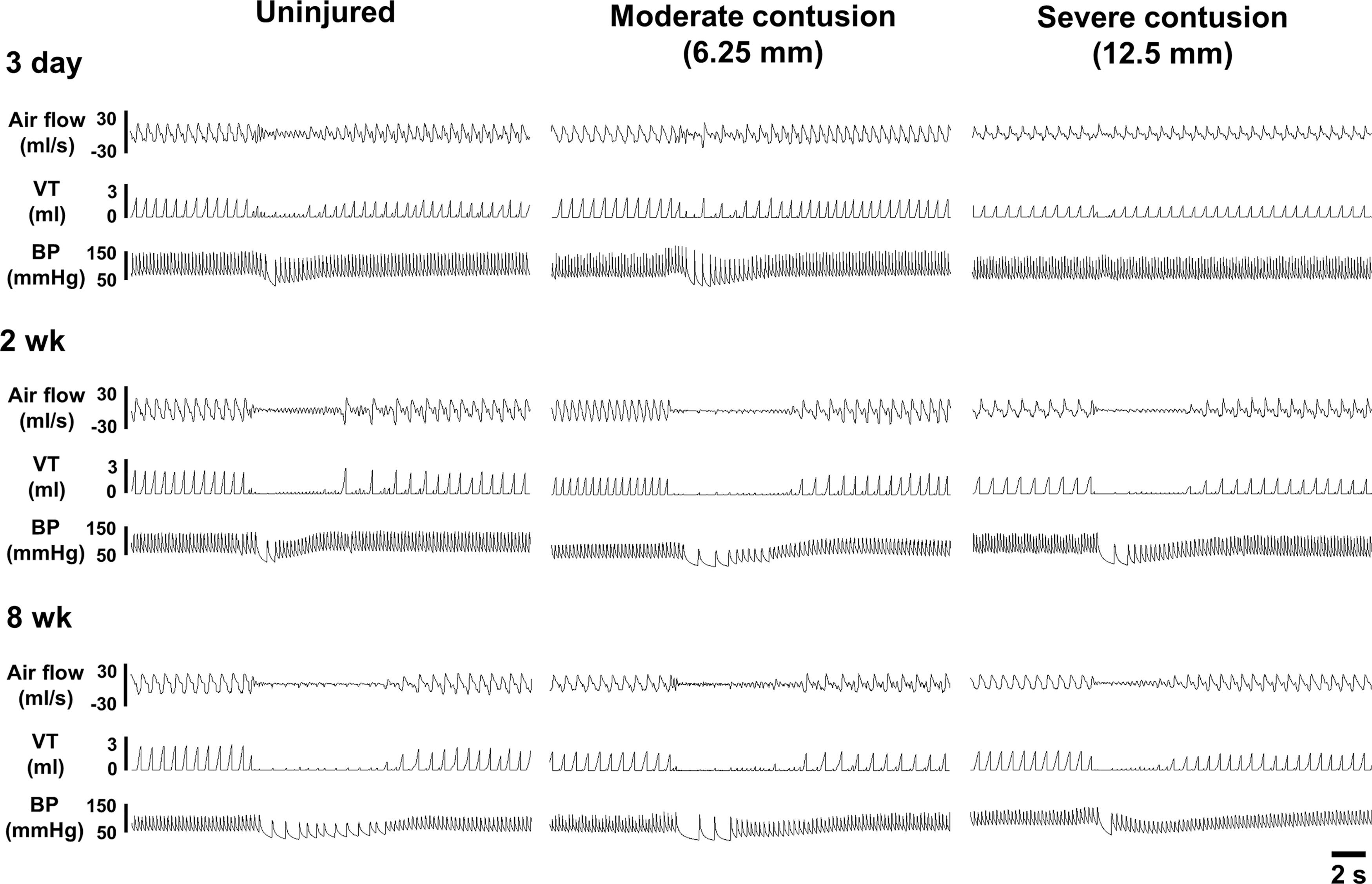
Representative examples of the cardiorespiratory pattern before, during, and after capsaicin-induced pulmonary chemoreflex in anesthetized uninjured and contused animals at 3 days, 2 weeks, and 8 weeks post-surgery. VT, tidal volume; BP, blood pressure.
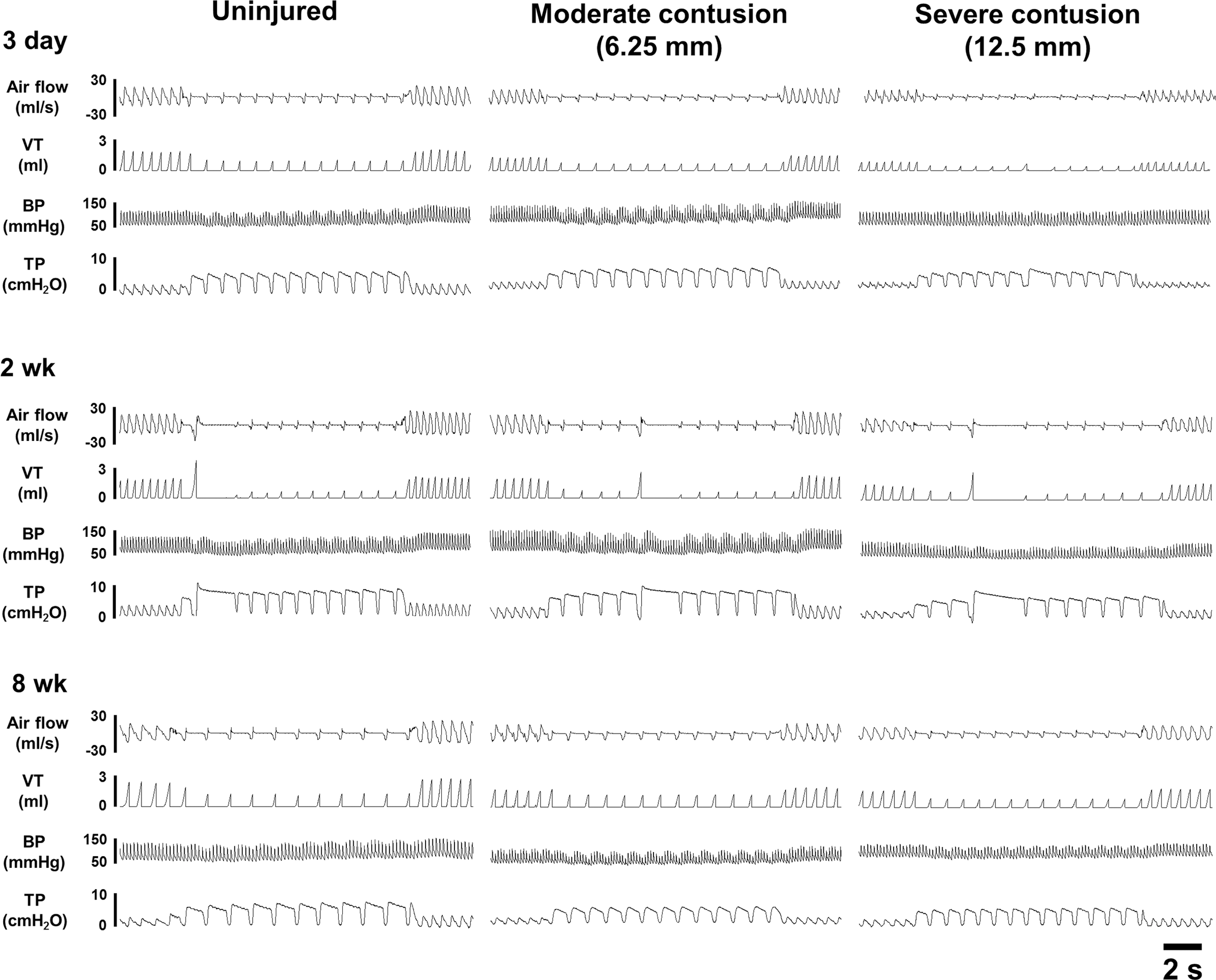
Representative examples of the cardiorespiratory pattern before, during, and after Hering–Breuer inflation reflex in anesthetized uninjured and contused animals at 3 days, 2 weeks, and 8 weeks post-surgery. VT, tidal volume; BP, blood pressure; TP, tracheal pressure.
Values are means ± SE.
p < 0.05; ** p < 0.01 vs. uninjured.
#p < 0.05; ##p < 0.01 significant difference between moderate contusion (6.25 mm) and severe contusion (12.5 mm).
n, number of animals; f, respiratory frequency; VT, tidal volume; MV, minute ventilation; MAP, mean arterial blood pressure; HR: heart rate.
p < 0.05; ** p < 0.01 vs. uninjured; #p < 0.05; ##p < 0.01 vs. vagal-intact;
θp < 0.05 vs. moderate contusion (6.25 mm).
The tidal volume remained lower in both contused animals (moderate contusion: 1.8 ± 0.1 mL; severe contusion: 1.4 ± 0.1 mL; uninjured: 2.2 ± 0.1 mL) at 2 weeks post-injury, but only severely contused animals had lower minute ventilation than uninjured animals (p < 0.01, Table 2). At 8 weeks post-injury, the breathing pattern of moderately contused animals was comparable to that of uninjured animals. However, severely contused animals still had a rapid shallow breathing pattern (frequency: 124 ± 6 breaths/min; tidal volume: 1.9 ± 0.1) compared with uninjured animals (frequency: 108 ± 4 breaths/min; tidal volume: 2.3 ± 0.1) (p < 0.05, Table 2). Despite that there were still some deficiencies in ventilatory behaviors at 2 and 8 weeks post-contusion, the blood gas parameters (PaO2, PaCO2, pH) were similar between uninjured and contused animals (Table 3).
The mean arterial blood pressure and HR were similar between uninjured and contused animals at 3 days and 2 weeks post-injury. At 8 weeks post-injury, we noticed that the HR was significantly higher in contused animals (moderate contusion: 377 ± 9 beats/min; severe contusion: 354 ± 7 beats/min) than in uninjured animals (326 ± 11 beats/min) (p < 0.05; Table 2) despite that the mean arterial blood pressure of contused animals was similar to that of uninjured animals.
Pulmonary chemoreflex
Representative examples of capsaicin-induced pulmonary chemoreflex are represented in Figure 2. Intrajugular capsaicin administration can induce the pulmonary chemoreflex characterized by apnea, bradycardia, and hypotension. 13,21,33 The expiratory duration was significantly prolonged from 0.25 ± 0.01 to 2.94 ± 0.77 sec (1119 ± 261% BL) in uninjured animal at 3 days post-injury (p < 0.01, Fig. 4); however, capsaicin-induced apneic response was attenuated in both moderately (0.24 ± 0.01 sec; 443 ± 208% BL) and severely (0.31 ± 0.03 sec; 206 ± 40% BL) contused animals (p < 0.01, Fig. 4). Similarly, the capsaicin-induced hypotension and bradycardia were also blunted in contused animals. Specifically, the mean arterial blood pressure decreased from 95 ± 5 mm Hg to 74 ± 6 mm Hg (78 ± 5% BL) in uninjured animals, but was able to still be maintained at 87 ± 5% BL and 94 ± 3% BL in moderately and severely contused animals, respectively (Fig. 4). The heart rate was significantly decreased in both uninjured (62 ± 11% BL) and moderately (73 ± 8% BL) contused animals (p < 0.01, Fig. 4), but the bradycardia response was significantly attenuated in severely contused animals (87 ± 3% BL) (p < 0.01, Fig. 4).
Changes in expiratory duration (TE), mean arterial blood pressure (MAP) and heart rate (HR) induced by capsaicin injection in uninjured and contused animals at 3 days, 2 weeks, and 8 weeks post-injury. Data were presented as a percentage of the baseline value (%BL). **p < 0.01 vs. BL. ## p < 0.01 significant differences from the uninjured animal.
The pulmonary chemoreflex could be evoked in both contused groups during the subchronic to chronic injury state. Specifically, the expiratory duration of moderately contused animals was prolonged to 2115 ± 276% BL and 1841 ± 254%BL at 2 and 8 weeks post-injury, respectively. The severely contused animals also exhibited a similar apneic response (2 weeks: 1798 ± 489% BL; 8 weeks: 1726 ± 298% BL). Unlike blunted pulmonary chemoreflex at 3 days post-contusion, the intensity of the pulmonary chemoreflex (e.g., apnea, hypotension, and bradycardia) was similar between contused and uninjured animals at 2 and 8 weeks post-injury (Fig. 4).
Hering–Breuer inflation reflex
Representative examples of Hering–Beruer inflation reflexes are presented in Figure 3. Increase of end-expired pressure induced a significant prolongation in the expiratory duration and reduction in both inspiratory duration and respiratory frequency (Fig. 5). At 3 days post-injury, the expiratory duration was significantly increased from 0.25 ± 0.01 sec to 0.80 ± 0.12 sec (316 ± 40% BL) in the uninjured group and from 0.26 ± 0.01 sec to 0.92 ± 0.11 sec (363 ± 46% BL) in the moderate contusion group during increase of end-expired pressure; however, this response was significantly attenuated in the severely contused group (230 ± 17% BL) (p < 0.05, Fig. 5). The inspiratory duration was significantly reduced to 0.22 ± 0.01 sec to 0.20 ± 0.004 sec (89 ± 3% BL) in uninjured animals as expected (p < 0.01, Fig. 5). However, the inspiratory duration was not significantly reduced during lung inflation in either the moderately (97 ± 2% BL) or the severely (106 ± 5% BL) contused group (Fig. 5).
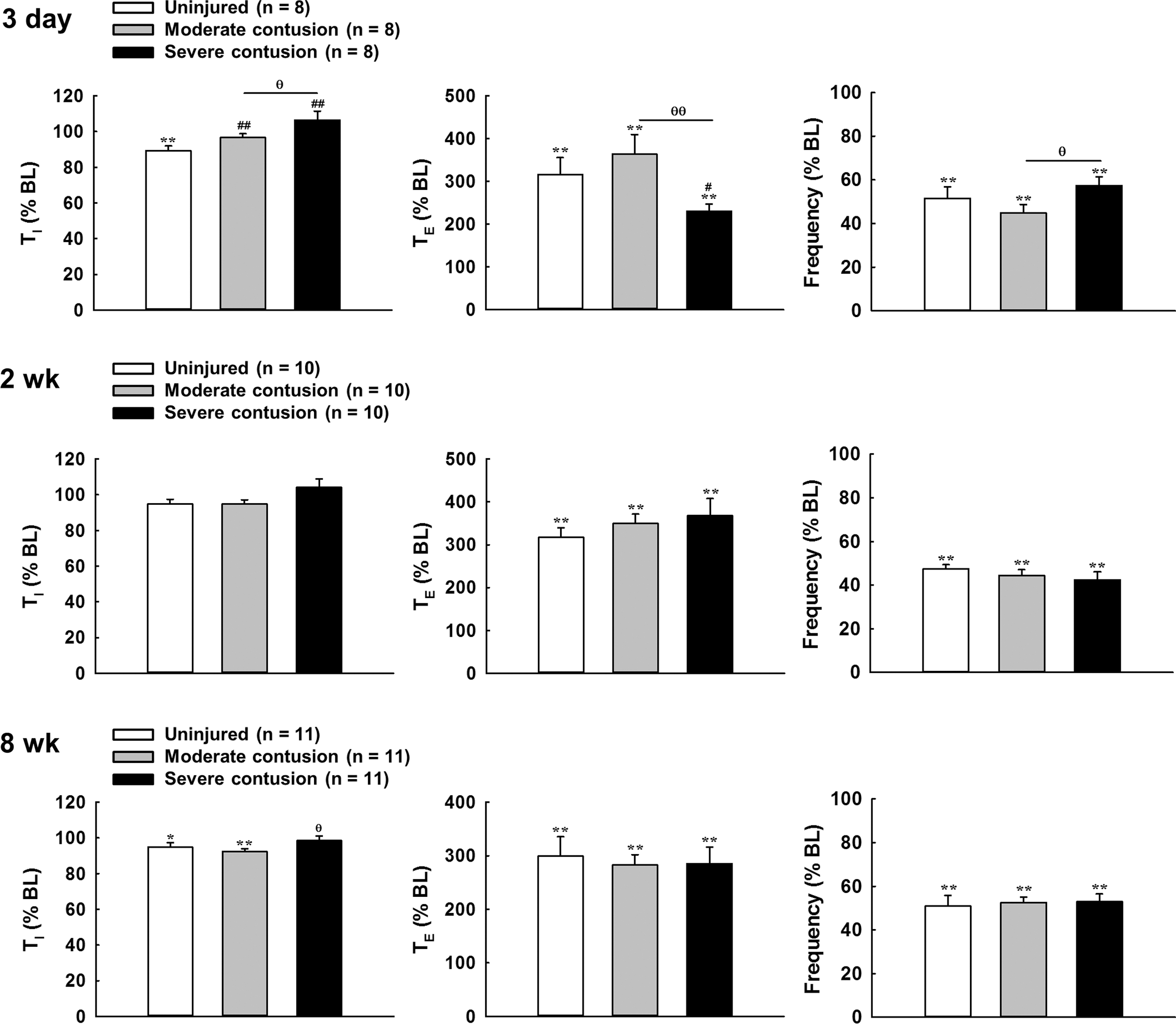
Changes in the inspiratory duration (TI), expiratory duration (TE), and respiratory frequency induced by lung inflation in uninjured and contused animals at 3 days, 2 weeks, and 8 weeks post-injury. Data were presented as a percentage of the baseline value (% BL). *p < 0.05; **p < 0.01 vs. BL. # p < 0.05; ## p < 0.01 significant differences from the uninjured animal. θ p < 0.05; θθ p < 0.01 significant differences between moderately (6.25 mm) and severely (12.5 mm) contused animals.
At 2 weeks post-surgery, increased end-expired pressure can induce a similar and significant prolongation of the expiratory duration in uninjured (317 ± 23% BL), moderately contused (349 ± 22% BL) and severely contused (368 ± 40% BL) animals (Fig. 5), which caused a significant reduction in the respiratory frequency (uninjured: 47 ± 2% BL; moderate contusion: 44 ± 3% BL; severe contusion: 42 ± 4% BL) during lung inflation (p < 0.01, Fig. 5). The alteration of the expiratory duration and respiratory frequency was also similar between uninjured and contused animals at 8 weeks post-injury (Fig. 5).
Vagotomy
Bilateral cervical vagotomy removed vagal afferent inputs and induced a substantial change in the breathing patter and blood gas parameters (Tables 2 and 3). The respiratory frequency was significantly reduced because of the prolongation of both inspiratory and expiratory duration in all groups at three time points. The respiratory frequency was similar among groups at three time points; however, the tidal volume of severely contused animals was significantly lower than that of uninjured and moderately contused animals at 3 days and 2 weeks post-injury (p < 0.05, Table 2). The capacity of increasing the tidal volume following vagotomy was lower in severely contused animals (156 ± 5% BL) than in uninjured (169 ± 4% BL) and moderately contused (182 ± 8% BL) animals at 3 days post-injury (p < 0.05). Interesting, vagotomy-induced increase in tidal volume was somewhat greater in severely contused animals (2 weeks: 208 ± 9% BL; 8 weeks: 196 ± 5% BL) than in uninjured (2 weeks: 179 ± 6% BL; 8 weeks: 175 ± 7% BL) animals during the subchronic to chronic injury state (p < 0.05).
Both pulmonary chemoreflex and Hering–Breuer inflation reflex were greatly attenuated following bilateral cervical vagotomy as expected, suggesting that these two respiratory reflexes were primarily mediated by activation of vagal afferents (data not shown).
Contusion severity
Representative examples of transverse sections of severely contused spinal cord at 3 days, 2 weeks, and 8 weeks post-injury are shown in Figure 6A. At 3 days post-injury, the lesion epicenter was filled with infiltration of blood and the lesion cavity was 14 ± 2% and 17 ± 3% of the whole spinal cord section area in moderately and severely contused animals, respectively (Fig. 6B). At 2 weeks post-injury, the lesion severity was increased (moderate contusion: 39 ± 3%; severe contusion: 49 ± 4%) and the lesion extended to ∼1–2 mm from the lesion epicenter. This lesion pattern maintained at 8 weeks post-injury (moderate contusion: 40 ± 3%; severe contusion: 50 ± 3%), suggesting the histopathological progress of the contused spinal cord primarily occurred during the acute to subchronic injury state (Fig. 6B). In addition, the lesion severity of severe contused animals was generally greater than for moderately contused animals around the lesion epicenter (p < 0.05, Fig. 6B).
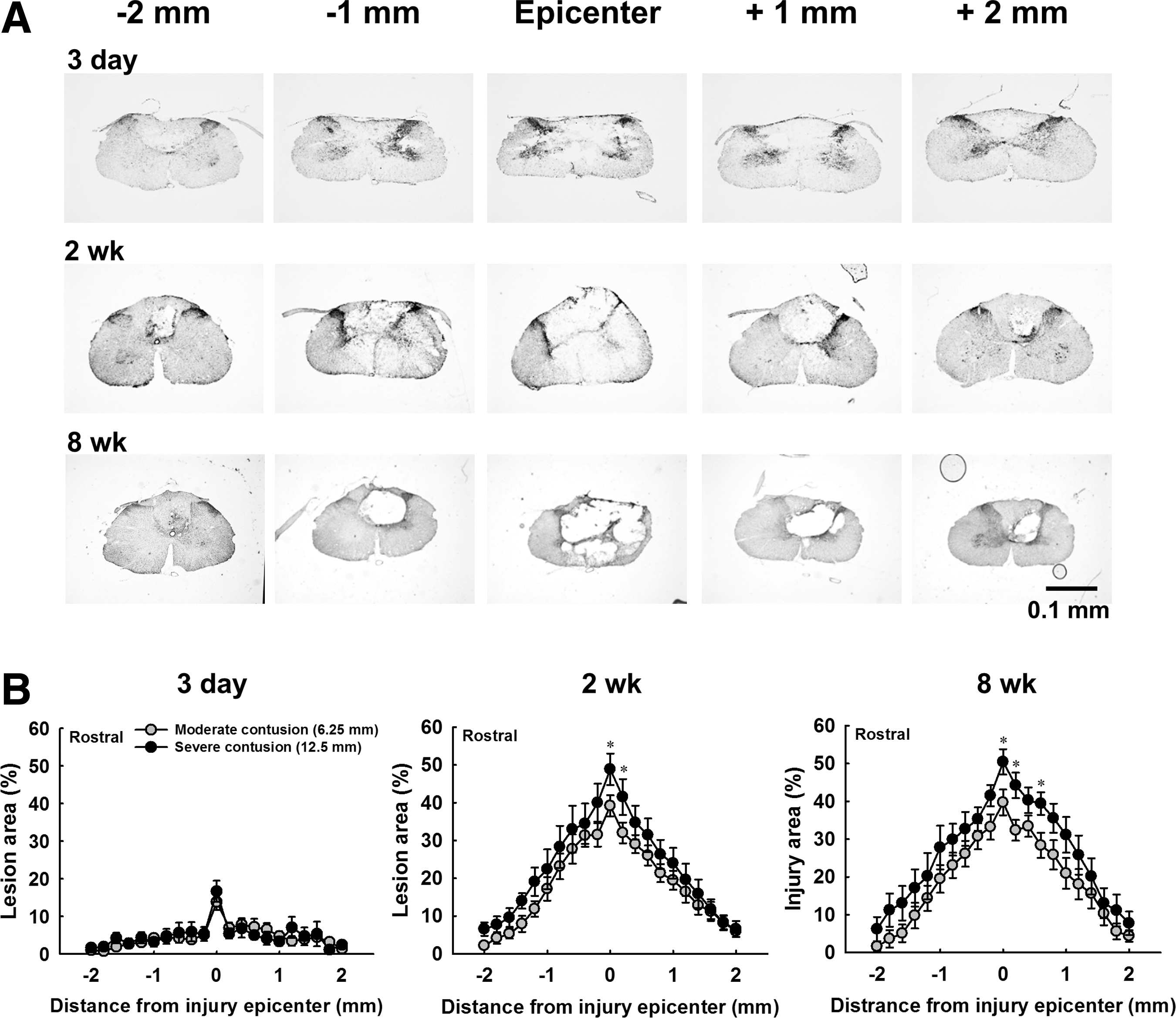
Cervical spinal cord histology in severely contused animal at 3 days, 2 weeks, and 8 weeks post-contusion.
Phrenic motoneuron pool
Representative examples of cholera toxin subunit B labeled phrenic motoneurons in uninjured and contused animals are shown in Figure 7A. The number of labeled phrenic motoneurons was 489 ± 31 in uninjured animals, and the phrenic motoneuron number was significantly reduced in both moderately (382 ± 29) and severely (331 ± 15) contused animals (p < 0.01, Fig. 7B). In addition, the phrenic motoneuron number was similar between the left and right sides in both contused groups, suggesting that the impact rod evenly damaged the bilateral spinal cord (Fig. 7C).
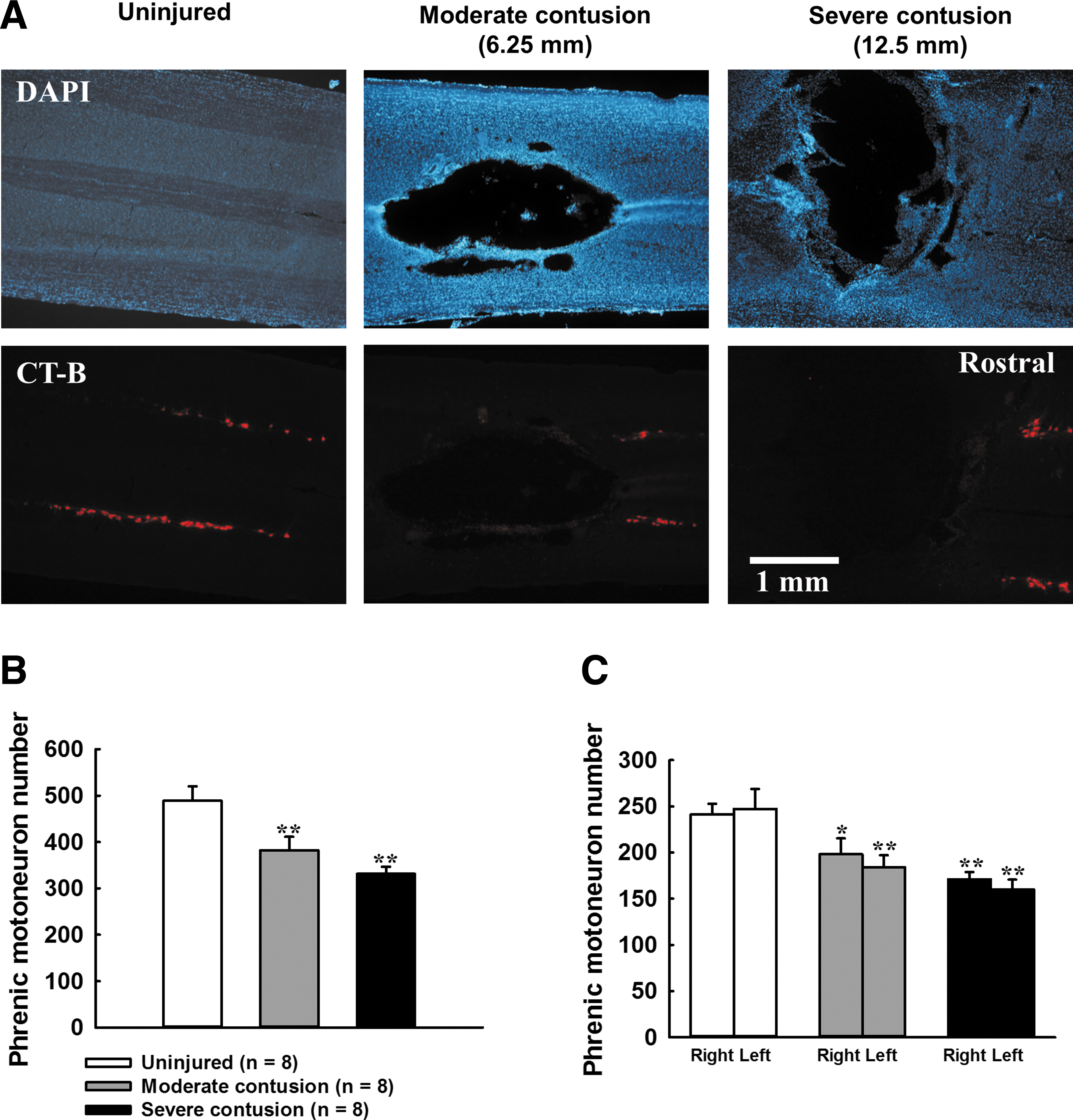
Phrenic motoneuron pool in uninjured and contused animals at 8 weeks post-surgery.
Discussion
Our results demonstrated that the cervical contusion model used in the present study caused a significant lesion in midcervical spinal cord and loss of phrenic motoneurons, which resulted in a persistent reduction (i.e., 8 weeks post-injury) in the tidal volume during normoxic and hypercapnic breathing in unanesthetized rats. Further, lung vagal-mediated pulmonary chemoreflex and Hering–Breuer inflation reflex were attenuated during the acute but not the subchronic and chronic injury states with rats under anesthesia. These results suggest that cervical spinal contusion not only impacts the baseline breathing pattern but also influences the respiratory reflexes that were not mediated by the spinal pathway.
Critique of methods
Several studies have reported that cervical contusion can specifically compromise the phrenic motor system (i.e., phrenic bursting and diaphragm EMG activity) but the ventilatory behavior was only transiently changed after injury. 7,8,27,39 However, the contused animals in the present study had a significantly lower tidal volume than uninjured animals from 1 day to 8 weeks post-injury, suggesting that our cervical contusion injury model can cause a persistent impairment in the ventilation. We proposed that two factors may contribute to long-term respiratory impairment in the current study. First, the current experiment used male rats as an injury model. A previous study demonstrated that female rats exhibited a better recovery in tidal volume and phrenic activity during hypercapnia following high cervical hemisection, 40 suggesting that female hormones may be benefit respiratory recovery. In other words, male rats may sustain a more severe and longer-term respiratory impairment after cervical spinal cord injury. Our data showed that the tidal volume remained lower than in uninjured animals at 8 weeks post-injury; however, reduction of tidal volume in female contused rats was not significant or only occurred at 1 day post-contusion. 7,8 Second, respiratory motor outputs may be differentially influenced by unilateral versus midline spinal injury. Previous studies reported that there would be a compensatory response of uninjured side phrenic motor outputs following unilateral hemisection. 41 However, the midline spinal contusion caused a significant lesion in bilateral gray matter, and resulted in a loss of bilateral phrenic motoneurons. Accordingly, bilateral phrenic motor outputs were compromised, and may not be able to sustain the normal ventilation in the present study.
Alteration of breathing pattern after midcervical contusion in unanesthetized rats
The present study demonstrated that the breathing pattern was differentially expressed during the acute versus the subchronic and chronic injury states following midcervical contusion. At 1 day post-injury, the contused animal showed a higher respiratory frequency than uninjured animals during the baseline. Golder and coworkers had demonstrated that the rapid breathing pattern of C2 hemisected rats became normal after bilateral vagotomy. 22 Therefore, we speculated that the reduced tidal volume in contused animals may modulate the inputs from the pulmonary stretch receptors and then increase the respiratory frequency. In addition, cervical contusion also damaged the dorsal column and dorsal horn, which may compromise the somatic afferent ascending pathway and, in turn, influence respiratory frequency. 42 Moreover, homeostasis of blood gas and pH was not stable during the acute contused injury phase. Activation of central and/or peripheral chemoreceptors could augment respiratory frequency following cervical contusion.
Respiratory frequency returned to the normal value at 2 weeks post-injury; however, the tidal volume remained lower than in uninjured animals from 2 to 8 weeks post-injury. The reduction of tidal volume could partially have resulted from loss of phrenic motoneurons following contusion, because our histological data indicated that the number of phrenic motoneurons of contused animals was only ∼68–78% of that of uninjured animals. However, Nichols and coworkers demonstrated that the baseline tidal volume can remain at the normal value after selective respiratory motoneuron death induced by intrapleural injections of cholera toxin B fragment conjugated to saporin. 43 Therefore, reduction of phrenic motoneurons themselves may not be severe enough to reduce the tidal volume. We did observe that a portion of lateral and ventral funiculus was also impaired at the lesion epicenter, which can interrupt the bulbospinal respiratory pathway to phrenic and intercostal motoneurons below the injury. Therefore, a persistent impairment of tidal volume in the present study could be caused by both phrenic motoneuron loss and interruption of the respiratory pathway following cervical contusion.
Alteration of breathing pattern after midcervical contusion in anesthetized rats
Respiratory frequency was similar between uninjured and contused animals; however, the tidal volume was lower in contused animals at 3 days and 2 weeks post-injury. Unlike the rapid shallow breathing pattern that was observed during the acute injury phase in unanesthetized rats, anesthetized rats showed rapid shallow breathing in the chronic injury state. In addition, we noticed that the tidal volume of moderately contused animals was comparable to that of uninjured animals at 8 weeks post-injury under anesthesia; however, unanesthetized contused rats still exhibited a significantly lower tidal volume. These differences in the breathing pattern between unanesthetized and anesthetized animals suggested that the impact of contused injury on the respiratory function could be confounded by the anesthetic.
Alteration of lung vagal-mediated reflexes after cervical contusion
The present study demonstrated that both capsaicin-induced pulmonary chemoreflex and lung inflation-induced Hering–Breuer reflex were attenuated in contused animals during the acute injury phase. In a high cervical hemisection animal model, the pulmonary chemoreflex disappeared at 1 day post-injury and remained blunted at 2 and 8 weeks post-injury. 13,21,33 Moreover, Hering–Breuer inflation reflex was blunted at 2 weeks post-injury in C2 hemisected rats. 34 These data suggested that lung vagal-mediated reflexes would be impaired during the acute cervical spinal cord injury state regardless of injury type (e.g., hemisection vs. contusion) or location (e.g., high vs. midcervical region).
Several factors may cause alteration of lung vagal-mediated reflex following cervical spinal cord injury. First, Schelegle and coworkers demonstrated that both Hering–Breuer inflation reflex and pulmonary chemoreflex were blunted following bleomycin-induced lung fibrosis. 44 Therefore, alteration of the breathing pattern and respiratory muscle and lung mechanics following spinal cord injury may be involved in modulation of lung vagal-mediated respiratory reflexes. 24 Second, blood gas parameters were significantly changed during the acute injury phase. The relative lower PaO2 and higher PaCO2 of severely contused animals suggested that activity of both peripheral and central chemoreceptors were maintained at a higher level, which can increase the central respiratory drive and may override the respiratory inhibitory inputs during lung vagal afferent inputs. 45 Third, lung vagal-mediated reflexes are initiated and/or modulated by the central respiratory-related region in the medulla and pons (e.g., nucleus tractus solitarius and dorsolateral pons). 46 Several reports had shown that respiratory neuroplasticity could be evoked in these central respiratory regions under experimental conditions. 47,48 Moreover, Felix and coworkers had demonstrated that spinal cord injury can induce alteration in cellular signaling of supraspinal regions. 49 Therefore, attenuation of lung vagal-mediated reflexes could be partially the result of changes in activity of the central respiratory circuit.
Recovery of lung vagal-mediated reflexes following cervical contusion
Our present data demonstrated that both the pulmonary chemoreflex and the Hering–Breuer inflation reflex were attenuated during the acute injury state but recovered to the normal level during the subchronic to chronic injury state. Similarly, high cervical spinal hemisection also caused a transient attenuation of lung vagal-mediated reflexes. 13,21 These data suggested that there is a time-dependent change in the elements of the respiratory reflex arc (e.g., lung vagal afferent activity, central integration region, and spinal motor outputs) following cervical spinal cord injury. In addition, we noticed that the tidal volume remained lower in severely contused animals at 2 and 8 weeks post-injury even when the lung vagal-mediated reflexes resumed. In addition, whole body plethysmography results demonstrated that the respiratory frequency of contused animals was similar to that of uninjured animals at 1–8 weeks post-injury; however, the tidal volume of contused animals was lower than that of uninjured animals from 1 day to 8 weeks post-injury. These observations indicated that the recovery of respiratory frequency was faster than the tidal volume after cervical contusion.
Physiological significance
The present study established a midcervical contusion model that can induce a long-term respiratory impairment. This model is more clinically relevant than a traditional unilateral hemisection model, and can enable evaluation of potential therapy (e.g., neurorehabilitative training, cellular replacement, pharmacological approach) 50 –52 on respiratory function following cervical spinal injury. Our previous studies already demonstrated that lung vagal-mediated reflexes were blunted in a high cervical spinal hemisection model. 13,21,33,34 The current data extended this observation, showing that midcervical contusion also resulted in attenuation of lung vagal-mediated reflexes. These data suggested that attenuation of lung vagal-mediated respiratory reflexes may be a general phenomenon after cervical spinal cord injury. Clinical reports revealed that airway protective reflexes were weak in spinal cord injured patients, 53 which may further cause lung inflammation and result in severe pulmonary complications. Accordingly, future therapeutic strategies for respiratory recovery after spinal cord injury should also evaluate the recovery status of respiratory regulatory and defensive reflexes.
Footnotes
Acknowledgments
Support for this work was provided by grants from the National Health Research Institutes (NHRI-EX105-10223NC), Ministry of Science and Technology (Most 102-2320-B-110-004-MY3 & Most 105-2628-B-110-002-MY3), and NSYSU-KMU Joint Research Project (105-I004).
Author Disclosure Statement
No competing financial interests exist.