
Abstract
Cervical spinal cord injury (cSCI) occurs in over half of all cases of traumatic spinal cord injury (SCI), yet we lack therapies that can generate significant functional recovery in these patients. The development of animal models of cSCI will aid in the pre-clinical assessment of therapies and in understanding basic pathophysiological mechanisms. Here, we describe a clinically relevant model of cervical contusion-compression injury in the mouse. Using a modified aneurysm clip, we generated a bilateral, incomplete injury that mimics contusion-compression injuries most commonly observed in humans. We followed the recovery of injured and sham-operated (laminectomy-only) animals for 8 weeks post-surgery. Behavioral tests, including the Basso Mouse Scale (BMS), wire hanging, grip strength, and CatWalk automated gait analysis, showed that while natural recovery is limited, it occurs in a clinically relevant window during the subacute phase of injury (7–14 days post-SCI). BMS scoring demonstrated that, while injured animals are ambulatory, they do not recover normal locomotor ability. CatWalk analysis quantitatively showed a loss of coordination and motor ability, with minimal recovery. The wire hanging and grip strength tests confirmed a significant decrease in forelimb motor strength in injured animals. Histological analysis carried out during the subacute phase (7-day time point) and chronic phase (8-week time point) demonstrated that the lesion epicenter is formed by 7 days post-SCI. Volumetric analysis of protein kinase C gamma (PKCgamma)–stained axons revealed that this injury results in significant damage to the corticospinal tract caudal to the injury site. Finally, we used quantitative real-time polymerase chain reaction to show that genes associated with inflammation and glial scarring are upregulated as a result of injury. This study confirms that we can effectively model bilateral cervical injury in the mouse and provides a framework for future studies using this model to assess therapies.
Introduction
I
Methods
Animal care
Adult female C57Bl/6 mice (18–20 g) were obtained from Charles River Laboratories (Wilmington, MA). All animal care procedures were created according to the Guide to the Care and Use of Experimental Animals designed by the Canadian Council of Animal Care. All procedures were vetted and approved by the University Health Network Animal Care Committee.
Clip-compression spinal cord injury
Our laboratory has extensively characterized the use of a modified aneurysm clip to generate a bilateral contusion-compression injury in rats and mice. Anesthesia was induced using inhalant isoflurane (1–2%) delivered in a 1:1 mixture of 02/N20. The incision area was shaved and disinfected using 10% povidone followed by 70% ethanol. A midline skin incision was made in the lower cervical and upper thoracic areas between C5 and T2. The three muscle layers covering the dorsal aspect of the vertebrae were retracted in preparation for the removal of the C6 and C7 laminae. Similar to the surgical procedure for thoracic injury in mice, a two-level laminectomy was performed with microscissors. Following removal of the C7 and C6 laminae, a microrongeur was used to widen the laminectomy for better visualization of the nerve roots. While this process is key for the correct placement of the clip, the surgeon must ensure that the articular processes and facet joints are left intact to prevent instability of the spinal column and development of neuropathic pain. The C6 spinal level was compressed for 40 sec with a 5.25-g clip. The injury site was covered by a small piece of Surgifoam® (Ethicon, Inc., Somerville, NJ), and the overlaying muscle and skin were sutured with 5-0 Softsilk® sutures (Covidien Ltd., Dublin, Republic of Ireland). Pain was treated post-operatively with analgesics (subcutaneous buprenorphine, 0.05 mg/kg, two times daily for 3 days). Subcutaneous saline was administered twice-daily to prevent dehydration. For recovery, animals were housed in standard mouse cages with absorbent bedding (5 animals per cage) and placed in a chamber that maintained the temperature at 27°C and humidity at 50%. Animals had their bladders manually expressed three times daily until the return of normal bladder function. Clavaomox was delivered in drinking water to prevent infection post-operatively.
Neurobehavioral testing
Basso Mouse Scale scoring
Animals were evaluated once per week by two observers. Mice were placed individually in an open field where locomotor ability was scored. Motor function was scored on an 11-point scale (0–10). Animals with normal locomotion achieve a score of 10. The scale evaluates parameters including joint movement, stepping ability, coordination, and trunk stability. Eleven injured animals and 5 laminectomy-only animals were scored at each time point.
Grip strength testing
This test is used to assess forelimb motor function based on the ability of the animal to exert a pulling force on a metal grid attached to an electronic grip strength meter (SDI Grip Strength System model DFM-10; San Diego Instruments, San Diego, CA). Animals were tested once per week; each trial consisted of five separate pulls. The highest and lowest forces were omitted and the remaining three were averaged. Grip strength was recorded for 11 injured animals and 5 laminectomy-only animals at each time point.
Wire hang test
This test is designed to assess sensorimotor function. Mice have a natural grasping instinct, and therefore no training is required for this test; however, to minimize stress, animals were habituated to this task in two to three sessions before beginning data collection. Animals were placed on top of a wire cage lid and shaken gently three times to encourage them to grasp onto the wire. The cage lid was then inverted so that the animals were suspended over a pillow (blue bench diaper) at a hight of 15 cm. After inversion of the cage lid, the hanging time was recorded in seconds. Nine injured animals and 5 laminectomy-only animals were evaluated at each time point.
Automatic gait analysis (CatWalk)
Gait testing was performed using the CatWalk system (Noldus Information Technology, Wageningen, The Netherlands). Animals traverse a horizontal glass plate as a fluorescent light is projected across it. Footfalls displace the fluorescent light and are recorded by a camera positioned beneath the walkway. Raw data were collected and analyzed using CatWalk software. For analysis, crossing speed was standardized, and at least three consecutive step cycles were required for the analysis of one crossing. The software labeled paw prints automatically. We selected the following parameters for analysis: stride length; print area; print width; print length; swing speed; maximum area of contact; and intensity. Paw intensity is measured in arbitrary units (a.u.). Six injured animals and 5 laminectomy-only animals were evaluated at each time point.
Tissue processing
Animals received an overdose of inhalant isofluorane in preparation for transcardial perfusion with 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS; pH 7.4). Spinal cords were removed and post-fixed in 4% PFA for a maximum of 4 h. Spinal cords were transferred to 30% sucrose and incubated overnight. In total, a 2-cm section of the cervical cord spanning the injury epicenter was embedded on dry ice using Shandon M-1 tissue embedding medium (Thermo Fisher Scientific, Waltham, MA). Cryosections were cut at a thickness of 30 μM and mounted on Fisherbrand® Superfrost Plus charged glass slides (Thermo Fisher Scientific).
Quantitative lesion analysis
To visualize axons and myelin, frozen sections were stained with anti-neurofilament (NF) antibody and eriochrome cyanine (EC). Sections were incubated overnight at 4°C with mouse monoclonal anti-NF antibody (Sigma-Aldrich, St. Louis, MO) at a concentration of 1:500. Sections were incubated for 1 h at room temperature in goat anti-mouse horseradish peroxidase (1:300). NF staining was visualized using diaminobenzidine (DAB; Vector Laboratories, Burlingame, CA). Sections were stained in 0.2% EC solution for 30 min and differentiated in 5% iron alum for 10 min. Tissue was further differentiated in borax ferricyanide for 15 min. Tissue was dehydrated in a graded series of ethanol and xylene. Using the Cavalieri Estimator function in the Stereo Investigator software (MBF Bioscience, Williston, VT), area and volume measurements were obtained for individual tissue sections. Spared white matter was defined as any region with normal EC staining and the presence of NF-positive axons. Spared gray matter was defined as any area displaying normal cytoarchitecture and where neuronal cell bodies could be easily identified. Lesional tissue was classified by a complete loss of normal cytoarchitecture, a fibrous appearance, and a decrease in, or complete absence of, neuronal cell bodies. Damaged white matter was identified based on the loss of EC staining, decreased density of NF-positive axons, the presence of vacuoles, and irregular or fibrous tissue. Lesion analysis was performed at 7 days post-SCI using 4 injured animals and at 8 weeks post-SCI using 5 injured animals.
Immunohistochemistry and quantification
Frozen sections (30 μM thick) mounted on glass slides were air dried for 10 minutes. Sections were rehydrated in PBS for 10 min and then blocked with a solution of 5% normal goat serum and 0.3% Triton-X in PBS at room temperature for 1 h. Sections were incubated with primary antibody in a humidified chamber overnight at 4°C, and rabbit polyclonal PKCγ (1:500) was used to mark fibers of the dorsal CST. Following overnight incubation, sections were washed three times in PBS and secondary antibody was applied. Secondary antibodies were diluted at a concentration of 1:300 in a solution of 5% normal goat serum in PBS. Sections were incubated in secondary antibody for 1 h at room temperature and washed three times in PBS. Nuclei were counterstained with 4’,6-diamidino-2-phenylindole (1:1000). Sections were mounted in Mowiol and cover-slipped. The volume and area of the dorsal CST in naïve animals (n = 4) and C6-injured animals at 7 days (n = 4) and 8 weeks post-SCI (n = 4) were determined using the Cavalieri Estimator function in the Stereo Investigator software (MBF Bioscience).
RNA isolation, complementary DNA synthesis, and real-time polymerase chain reaction quantification of gene expression
Mice were sacrificed at 3, 7, 14, 28, and 56 days post-injury, and a 7-mm sample of the spinal cord containing the epicenter of the injured tissue was extracted for RNA analysis. Total RNA from each individual sample was extracted using mirVana PARIS RNA and Native Protein purification kit (Ambion AM1556; Ambion, Inc., Austin, TX). RNA purity and quantity was assessed with a 2000C NanoDrop. Complementary DNA (cDNA) synthesis was performed using the SuperScript® VILO™ cDNA Synthesis Kit from Thermo Fisher Scientific. Relative quantification of gene expression was performed using the 2−ΔΔCT method. Table 1 lists the Applied Biosystems Inc. (Hercules, CA) TaqMan probes used in this study. Cyclophilin A (peptidylprolyl Isomerase A [PPIA]) was used as the endogenous control because its threshold cycle (CT) values were consistent and did not change across samples. For all time points, both C6-injured and laminectomy-only animals were analyzed. The animal numbers in each group are summarized in Table 2.
Assay ID, gene symbol, and amplicon length (base pairs) for five probes used in qRT-PCR analysis.
qRT-PCR, quantitative real-time polymerase chain reaction; PPIA, peptidylprolyl isomerase A; IL1B, interleukin-1 beta; TNF, tumor necrosis factor; IL16, interleukin-16; GFAP, glial fibrillary acidic protein.
qRT-PCR, quantitative real-time polymerase chain reaction; SCI, spinal cord injury; C6, cervical level 6.
Statistical analyses
Statistical analyses were performed using SigmaStat software (Systat Software, Inc., San Jose, CA). All data are represented as the mean ± standard error of the mean. p < 0.05 was considered significant.
Results
Modified surgical procedure allows for extradural clip placement at cervical level in the mouse
Our group has modified the clip-compression model, originally designed for rats, in order to perform injuries in mice. The primary modification has been adjustment of the dimensions of the clip. The mouse clip is made up of two curved blades that are tapered at their ends. The length of the blades has been shortened to 1.3 mm, and the thickness has been reduced to 0.3 mm at the blade tips (Fig. 1A). 16 Our group has successfully used the modified clip to perform injuries at the thoracic level in mice. 17,18 To use the clip at the C6 to C7 spinal level in the mouse, we did not change the clip dimensions. Instead, we modified our surgical procedure to allow for insertion of the clip in an area where large nerve roots pose challenges for the extradural clip placement. Similar to the surgical procedure for thoracic injury in mice, a two-level laminectomy is performed (Fig. 1C,D). We use a modified laminectomy procedure that creates enough space for clip placement while leaving the articular processes and facet joints intact to prevent instability of the spinal column and the development of neuropathic pain (Fig. 1B). To assess the general health of animals, we recorded body weight at 1 week post-SCI and 8 weeks post-SCI in C6-injured animals (n = 9) and laminectomy-only animals (n = 5; Fig. 1E). Injured animals are able to gain weight over the recovery period. Laminectomy-only animals display a similar pattern of weight gain over the recovery period. Basso Mouse Scale (BMS) scoring is typically used to assess locomotor function in models of thoracic SCI. We observed that BMS provides a useful measure of overall locomotor function of cervically injured mice (Fig. 1F). While injury reduces BMS score by approximately half, animals remain ambulatory. There is a slight, but significant, improvement in BMS score that occurs between the first and second week post-injury.

C6 clip-compression model in the mouse. (
Clip-compression injury triggers lesion formation beginning at 7 days post–spinal cord injury
Following contusive SCI, mice do not develop the cystic cavity that is characteristic of SCI in rats and humans. Instead, the injury epicenter in mice is characterized by the formation of a dense lesion core. Using myelin-specific EC in combination with NF staining, we have quantified the volume of lesional tissue, preserved gray matter, preserved white matter, and lesioned white matter in injured animals at 7 days (n = 4) and 8 weeks post-SCI (n = 5). Figure 2B shows an inset of normal white matter in the naïve spinal cord. Lypophilic EC stains myelin dark purple. Myelin is tightly packed around dark brown axons stained with NF and a DAB secondary (Fig. 2B). NF staining allows for the visualization of large motor neurons in the ventral horns of the white matter (Fig. 2C). At 7 days post-injury, a lesion core of dense fibrotic tissue has formed primarily in the ventral part of the cord (Fig. 2D). The dorsal horns of the gray matter and some dorsal white matter are preserved. Areas of lesioned white matter surround the lesion core. In these regions, white matter has a vacuolated appearance and contains hypertrophic axons (Fig. 2E). We have defined lesional tissues as areas where there is a complete loss of normal cytoachitecture, a decrease in, or the absence of, neuronal cell bodies, and that contain fibrous scar tissue. Once the lesion epicenter has formed at 7 days post-SCI, its size and morphology remain constant. There is no significant difference in the volume of lesional tissue at 7 days and 8 weeks post-SCI (Fig. 2J). Similarly, there is no significant change in the volume of preserved gray and white matter over the 8-week recovery period (Fig. 2G,H). There is a trend toward a decrease in the amount of lesional tissue at 8 weeks post-SCI (Fig. 2I); however, this trend is not significant. The lesion epicenter extends for approximately 1.8 mm along the rostrocaudal axis. The extent of the lesion epicenter remains constant from 7 days to 8 weeks post-SCI.
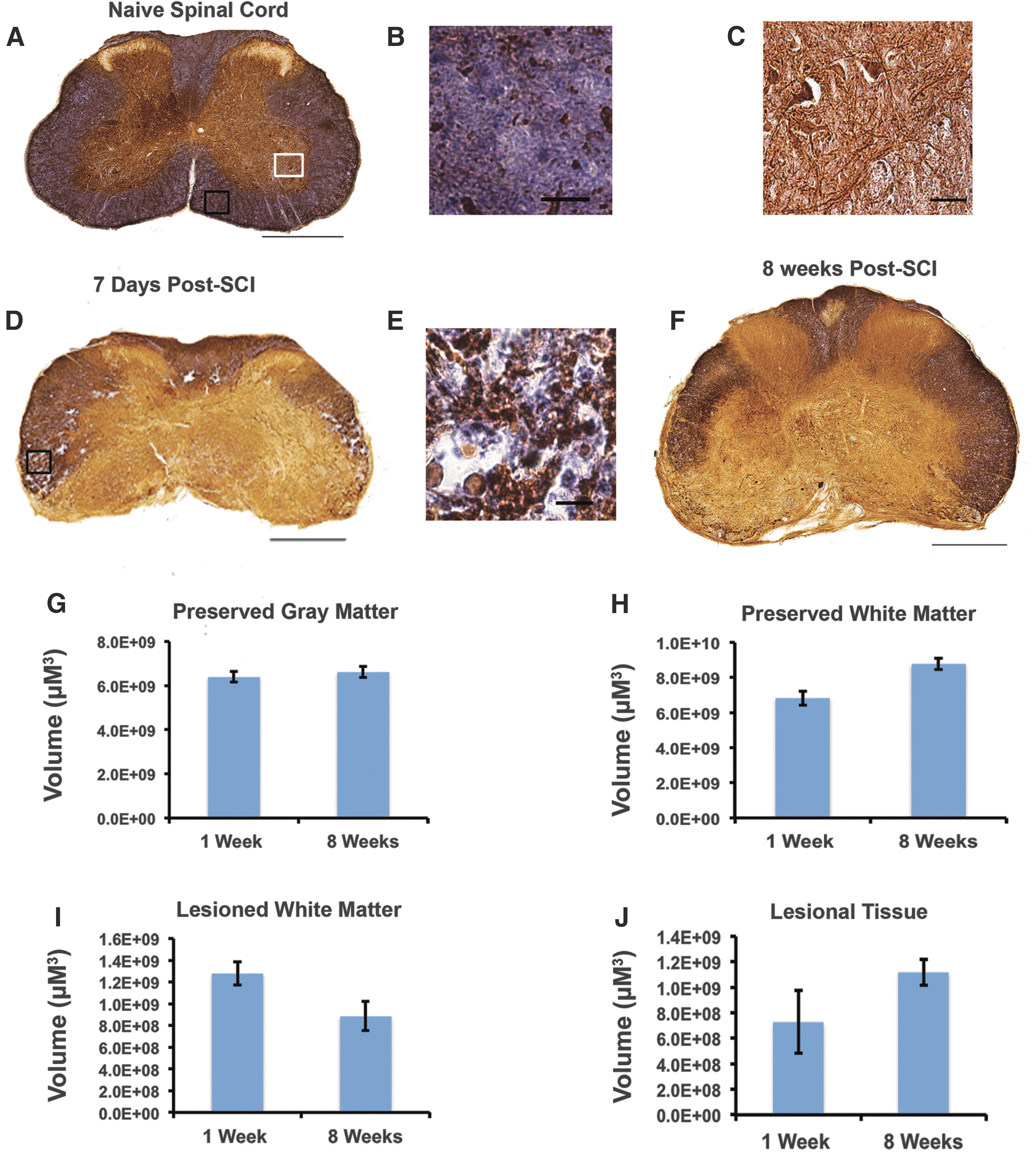
C6 clip-compression results in scar tissue formation at the lesion epicenter beginning at 7 days post-injury. (
Cervical level injury causes significant damage to the dorsal corticospinal tract
The CST is a key descending pathway that plays a significant role in motor control of the forelimbs and digits in humans and rodents. In the mouse spinal cord, the CST is found in the medial dorsal white matter. Here, we use the γ isoform of protein kinase C enzyme (PKCγ) to mark lamina II spinal cord interneurons and CST axons. 19,20 Previous work has shown that clip-compression injury results in Wallerian degeneration of CST fibers caudal to the injury and axonal “die-back” rostral to the injury. 21 First, we were interested in confirming these effects in the mouse model. Next, we aimed to assess any changes in the morphology of the dorsal CST over the course of recovery that might be an indication of injury-induced plasticity. In our model, PKCγ stained the dorsal CST and lamina II interneurons (Fig. 3A). Quantification of the total area of PKCγ-positive axons in the dorsal white matter shows a significant decrease in injured animals as compared to naïve animals (n = 4; Fig. 3B,C). Analysis of the CST area along the rostrocaudal axis shows that at 7 days (n = 4) and 8 weeks-post SCI (n = 4) the CST is completely absent caudal to the injury (Figure 3D). The significant decrease in PKCγ-positive CST axons remains constant during the recovery period. This observation suggests that if there is injury-induced plasticity occurring, that it must be detected using more-sensitive assays. In addition, the permanent loss of CST axons is consistent with very limited recovery of forelimb and locomotor function.
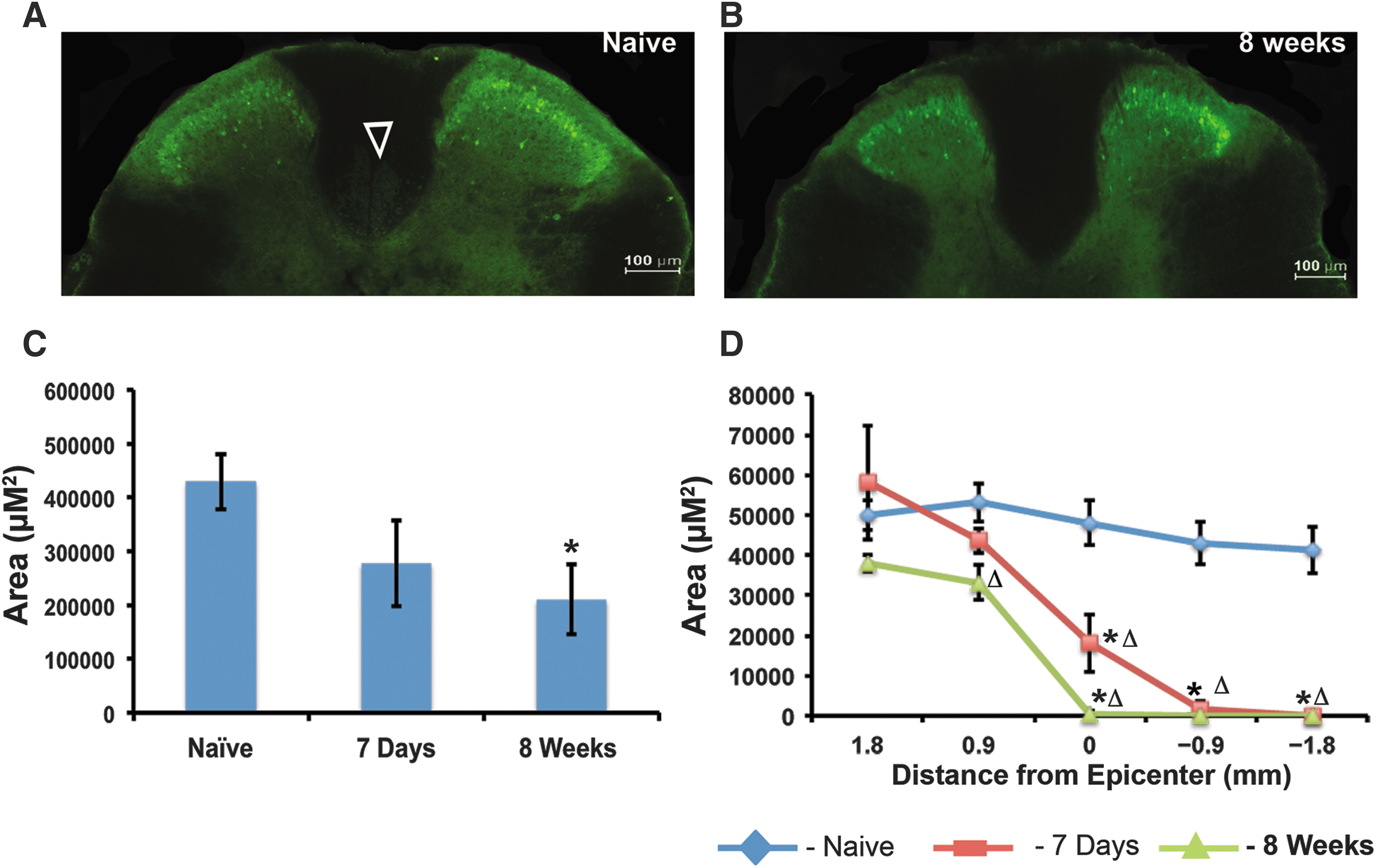
C6 clip-compression injury causes significant loss of PKCγ-positive corticospinal tract fibers (CSTs) below the level of injury. (
Cervical level injury results in sensorimotor deficits and gait dysfunction
The wire hanging test was used to characterize the effects of C6 injury on sensorimotor function (Fig. 4A). We tested animals pre-injury (baseline) and then on a weekly basis for 8 weeks post-SCI. At every time point, injured animals (n = 9) were compared to laminectomy-only controls (n = 5). Animals were placed on a standard cage top and inverted. The ability to maintain a grasp on the metal bars of the cage top was used as a measure of strength and motor function in the forelimbs and hindlimbs. To obtain readings in animals with normal grasping ability (baseline and laminectomy only), animals were allowed to hang for a maximum of 120 sec and were then removed from the metal grid. At 1 week post-injury, animals were able to grasp, on average, for 28 sec (± 8.1 sec). In comparison to laminectomy-only controls, this represents a 75% drop in grasping ability. At 8 weeks post-injury, animals were able to grasp for 24.8 sec (± 7.9 sec). To summarize, the wire hanging test demonstrates that injured animals show no significant improvement in grasping ability over the 8-week recovery period (two-way ANOVA with Tukey's post hoc, p < 0.05). To confirm results from the wire hanging test, grip strength testing was performed on injured animals (n = 11) and laminectomy-only animals (n = 5) each week, for 8 weeks. Grip strength testing showed that injury results in an almost 50% decrease in grip strength that does not return to baseline levels (Fig. 4B).

Sensorimotor deficits and loss of interlimb coordination show limited recovery. (
Automated gait analysis was performed using the CatWalk system. We performed CatWalk analysis pre-injury (baseline), 2 weeks, and 8 weeks post-SCI. At every time point, injured animals (n = 6) were compared to laminectomy-only controls (n = 5). First, we analyzed the effect of injury on coordinated movement using the regularity index and relative print position. Intact animals have a regularity index of 90–100%. Injured animals experience loss of coordination that results in disruption of regular step patterns and a lower regularity index. C6-injured mice have a regularity index of 70% at 2 weeks post-injury (Fig. 4D). We observed a slight, but not significant, increase in regularity index by the end of the recovery period. To ensure safe paw placement, rodents place the hindpaw in the same position as the previous forepaw. Injured animals lose this ability and display an increased distance between the placement of forepaws and hindpaws on the same side of the body. C6-injured animals placed forepaws and hindpaws twice as far apart, as compared to laminectomy-only controls (Fig. 4E,F). Paw placement values do not return to baseline levels over the 8-week recovery period. Representative walking patterns qualitatively show the loss of coordinated movement in C6-injured animals (Fig. 4B).
Analysis of paw statistics obtained using CatWalk provides a more detailed picture of the gait deficits that result from C6 clip-compression injury. We observed the expected decrease in hindlimb and forelimb stride length that is associated with contusion injury in rodents (Fig. 5A,B). Forelimb stride length displayed a larger decrease as compared to hindlimb stride length. In injured animals, forelimb stride length was decreased by more than 50% in comparison to laminectomy-only controls at 2 and 8 weeks post-SCI. Hindlimb stride length was decreased by approximately 27% at 2 and 8 weeks post-SCI. Contusion injury significantly decreased swing speed in both the hindlimbs and forelimbs. However, the forelimbs showed a greater decrease in swing speed as compared to the hindlimbs. At 2 and 8 weeks post-SCI, forelimbs display an approximately 35% decrease in swing speed as compared to laminectomy-only controls (Fig. 5C). In the hindlimbs, swing speed is decreased by 24% at 2 and 8 weeks post-SCI, as compared to laminectomy-only controls (Fig. 5D). The duration of the stance phase increased significantly in both forelimbs and hindlimbs as a result of injury. In contrast to the previous two parameters, injury has a greater effect on stance phase in the hindlimbs than in the forelimbs (Fig. 5E,F).
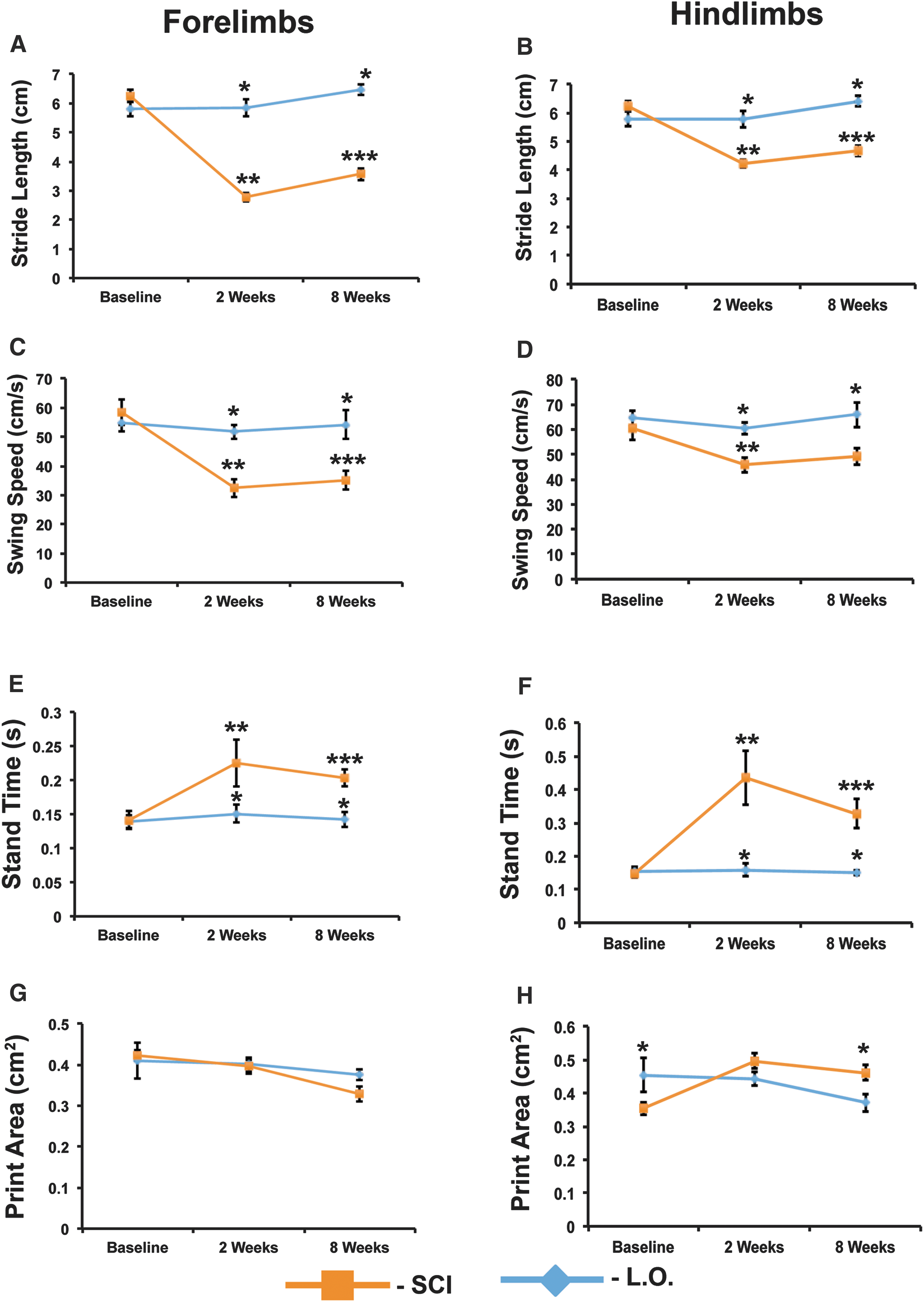
C6 clip-compression results in gait deficits that show limited recovery. (
Previous work in thoracic SCI has shown that print size will increase in response to contusion injury. 22 However, our CatWalk analysis did not reveal any significant changes in print size. Forelimb print area is almost completely unchanged in injured animals as compared to laminectomy-only animals (Fig. 5G). Hindlimb print area shows a small, but significant, increase at 8 weeks post-SCI (Fig. 5H). Print length and width are also unaffected in injured animals. Forelimb print width is 0.6 cm (± 0.03 cm) in injured animals and 0.7 cm (± 0.01 cm) in laminectomy-only animals at 8 weeks post-SCI. Hindlimb print width is 0.79 cm (± 0.02 cm) in injured animals and 0.75 cm (± 0.04 cm) in laminectomy-only animals at 8 weeks post-SCI. Similarly, forelimb print length is 0.9 cm (± 0.02 cm) in injured animals and 0.9 cm (± 0.01 cm) in laminectomy-only animals at 8 weeks post-SCI. Hindlimb print length is 1.2 cm (± 0.02 cm) in injured animals and 0.96 cm (± 0.02 cm) in laminectomy-only animals at 8 weeks post-SCI. In our model, print intensity showed a slight, but not significant, decrease in the forepaws of injured animals at 8 weeks post-SCI (207.9 ± 0.8 a.u.), as compared to baseline (212.9 ± 1.7 a.u.). Hindlimb print intensity was decreased slightly in injured animals at 8 weeks post-SCI (218.6 ± 2.5 a.u.), as compared to baseline (226.7 ± 2.2 a.u.). However, in contrast to what we observed in the forelimbs, two-way analysis of variance (ANOVA) analysis with Tukey's post hoc showed that the decrease in print intensity in the hindlimbs was significant (p = 0.026).
Genes associated with glial scarring and inflammation show similar expression patterns during the chronic phase of injury
Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was used to analyze the expression of glial fibrillary acidic protein (GFAP), tumor necrosis factor alpha (TNFα), interleukin (IL)-6, and IL-1β in spinal cord lysates at 3 days, 7 days, 14 days, 4 weeks, and 8 weeks post-SCI. The 3-day time point represents the acute phase, the 7- and 14-day time points encompass the subacute phase, and the 4- and 8-week time points span the chronic phase. Post-SCI, astrocytes enter a reactive state in which they upregulate the expression of GFAP. At 3 days post-SCI, GFAP expression is increased 3-fold as compared to laminectomy control animals. The increase in GFAP expression in not maintained throughout the subacute phase; at 14 days post-SCI, the increase in GFAP expression is not significant in comparison to laminectomy-only controls. During the chronic phase of injury, the levels of GFAP expression begin to increase. At 8 weeks post-SCI, the levels of GFAP expression return to the same levels observed at 3 days post-SCI (Fig. 6A). Members of the TNF family of cytokines display early upregulation as a result of SCI in mice. 23 In accord with previous work, our analysis showed a 4-fold increase in TNFα expression at 3 days post-SCI. During the subacute phase of injury, the levels of TNFα expression decrease. However, in the chronic phase of injury, TNFα expression increases to levels higher than those observed during the acute phase (Fig. 6B). Surprisingly, the levels of IL-6 and IL1β are not significantly higher when compared to laminectomy-only animals. IL-6 expression peaks during the subacute phase at 2 weeks post-SCI (Fig. 6C), whereas IL1β expression is highest during the chronic phase at 8 weeks post-SCI (Fig. 6D).
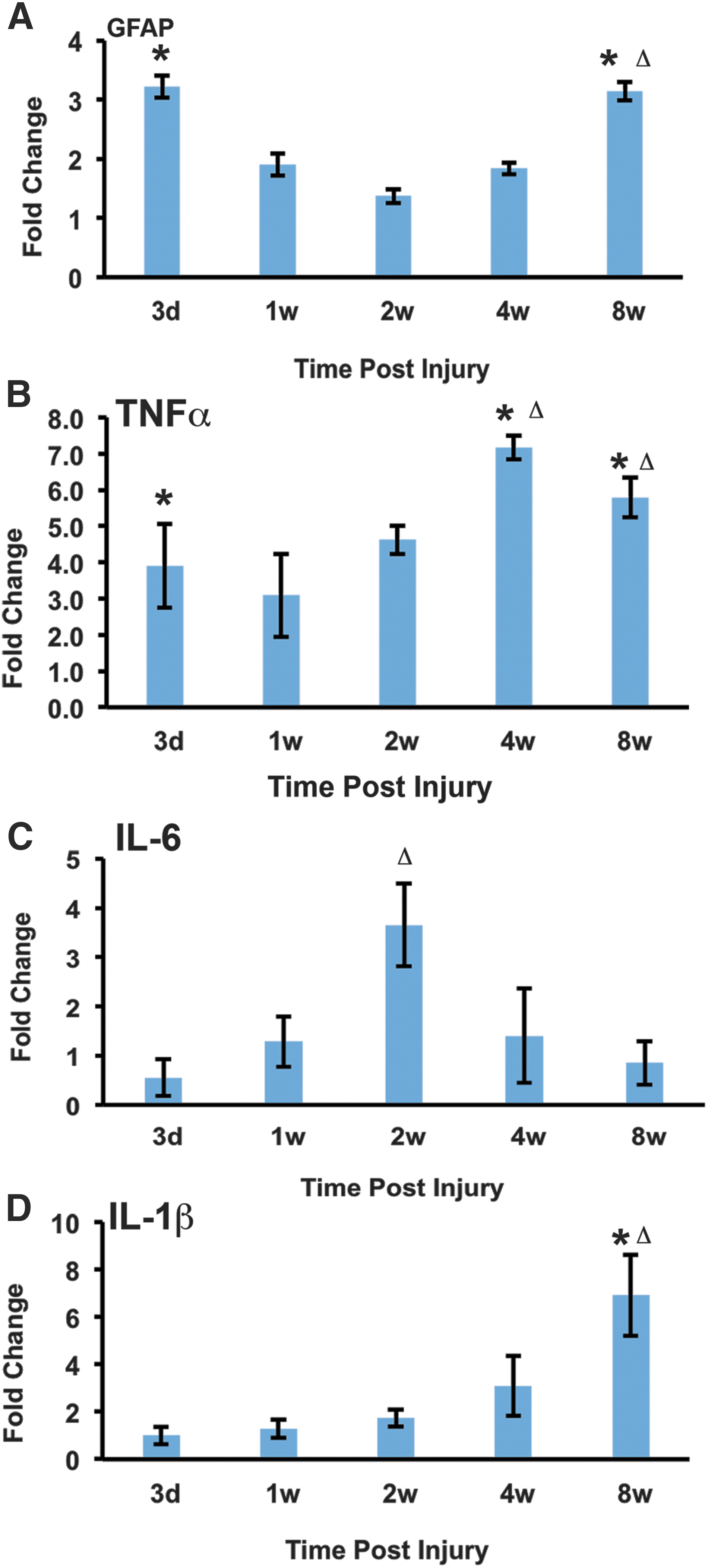
Genes associated with glial scarring and inflammation show similar expression patterns during the chronic phases of injury. (
Discussion
Here, we have described a clinically relevant mouse cervical contusion injury model. Using a modified aneurysm clip with a 5.25-g closing force, we can generate a bilateral, incomplete injury at the C6 level. In developing this model, one of our primary goals was to ensure high survival rates by optimizing the severity of the injury. It has been suggested that models of cervical injury in the mouse must be unilateral in order to preserve function and prevent mortality. 24 This work demonstrates that we can generate bilateral cervical injury while preserving some locomotor ability and preventing high levels of mortality. Based on pilot studies, we determined that using a 5.25-g clip with a 40-sec compression time resulted in measureable neurological deficits, while maintaining acceptable survival rates of 80% or higher. In comparison to previous work done by our group using a model of mouse thoracic injury, this is the lowest clip strength we have ever used. Another key aspect of this model was the development of a laminectomy technique that would allow for successful clip placement, while at the same time being minimally invasive. Due to the size and positioning of nerve roots in the cervical region, there is very little space to insert the clip. Optimal clip placement, with minimal damage to the nerve roots, requires good visualization. To achieve this, we created a wider laminectomy using a microrongeur. During the laminectomy, care is taken to keep the facet joints and the majority of the articular processes intact; this helps to maintain spinal column stability and prevents the development post-operative pain.
Mice display a unique response to contusion SCI that is not observed in other species. In other mammals (e.g., rats, hamsters, and monkeys) and in humans, SCI typically results in the formation of a cystic cavity surrounded by a spared white matter. In contrast, mice develop a fibrotic scar at the lesion core. 25 The response to SCI in mice is thought to be a result of an accelerated wound-healing process that involves changes in vasculature and extracellular matrix deposition. 26 Here, we show that the lesion epicenter is filled with what appears to be scar tissue by 7 days post-SCI. Our lesion analysis is based on morphological criteria defined by EC staining for myelin and NF staining for axons, a method that has been used in other studies of mouse models of SCI to quantify the extent of injury. 27,28 Further immunohistochemical analysis, looking at the expression of markers such as GFAP, chondroitin sulphate proteoglycan, fibronectin, and collagen at the injury epicenter, are required to confirm the presence of scar tissue. We observed that the size of the lesion epicenter remains constant for up to 8 weeks post-SCI. Further, there are no significant changes in preserved gray or white matter over the 8-week recovery period. Based on our histological analysis, we conclude that the lesion epicenter forms during the first week post-injury and undergoes limited remodelling from the acute to the chronic stages of injury. In contrast, following cervical clip-compression injury in the rat, the volume of lesional tissue decreases whereas the volume of the cavity increases over an 8-week recovery period. 11 This supports evidence that mice have an accelerated wound-healing process compared to rats.
We captured gene expression changes in our model using qRT-PCR. For this analysis, we chose a set of four candidate genes whose expression has been studied in rat and mouse models of SCI. For this analysis, we chose a small subset of genes that could provide preliminary mechanistic data in our model. This analysis does not confirm, using immunostaining, that astrogliosis contributes to the formation of the lesion epicenter. However, we know that this process is a major player in SCI pathophysiology. Therefore, our gene expression analysis looked at a primary marker of astrogliosis—GFAP. We show that, in our model, GFAP expression is significantly upregulated in the acute and chronic phases of injury. Inflammatory cytokines, including TNFα, are known to be one of the signals that trigger reactive astrogliosis. 29 We confirm that TNFα expression is upregulated as a result of injury, with the most significant upregulation occurring in the chronic phase of injury. This coincides with significant increases in GFAP and IL1β in the chronic phase. This observation supports the concept that the microenvironment of the chronically injured spinal cord is refractory to regeneration, attributable to inflammation and establishment of the glial scar. Therefore, qRT-PCR analysis confirms that C6 clip compression in the mouse results in expected changes in expression of genes associated with inflammation and glial scarring. However, further analysis by immunohistochemistry and Western blot is required to confirm these results.
To assess neurobehavioral deficits, we choose the grip strength test to assess forelimb function, the wire hanging test to provide a general picture of the effects of this injury on sensorimotor function, and the CatWalk test to provide quantitative measures of gait. BMS scoring was also used to provide a general picture of health condition and locomotor ability. In developing a model of cervical injury, our focus was on creating deficits in forelimb function. The grip strength test is an accepted means by which to assess neuromuscular function of the forelimbs in mice and rats post-SCI. 13,24 Here, we show that forelimb strength is reduced by approximately half and displays a small, but not significant, improvement that occurs primarily in the subacute phase of injury. The wire hanging test has been used to test motor strength in mice. 30 This test has drawbacks as a measure of forelimb function because animals are able to grasp with both their forepaws and hindpaws. However, this test has been used to assess forelimb function in some studies. For instance, this test was used to show that loss of cutaneous input attributed to ablation of spinal interneurons that regulate hand grasping causes impaired hand function in mice. 31 According to data from the wire hanging test, there is very limited natural recovery of motor strength and gripping ability post-SCI.
Cervical injury disrupts ascending and descending sensorimotor pathways required for normal locomotion. Here, we present the first CatWalk analysis of mice with bilateral cervical contusion injury. Regularity index and paw placement distance were used as measures of interlimb coordination. The regularity index measures how animals use regular step patterns. Intact rats and mice predominantly use the “Ab” step pattern. 22 The significant decrease in regularity index post-injury reflects an increase in missteps and a shift from the predominant “Ab” walking pattern to other walking patterns. Our analysis of regularity index showed that, in some cases, injured mice predominantly used the rotary patterns, which are only rarely observed in intact animals. The decrease in regularity index and increase in paw placement distance are also observed in rat models of contusion injury. Dorsal transection injury results in a decreased regularity index. However, following a recovery period, this parameter is greatly improved. 22 This indicates that sparing of the ventral tracts is enough to allow for functional recovery. Therefore, the minimal natural recovery in our model is most likely a result of more widespread damage to dorsal and ventral sensorimotor pathways. One of the most surprising observations from the CatWalk analysis was that this injury did not affect print size. Our analysis showed no change in print length, width, and area. In a rat model of thoracic contusion injury, print area is increased in both the forelimbs and hindlimbs. 22 Moreover, our previous analysis of bilateral contusion-compression cervical injury in the rat showed a significant decrease in forelimb and hindlimb print area. 11 We expected to see a similar decrease in print area in our mouse model. Based on these observations, we can conclude that print size is not the most instructive parameter for the assessment of locomotor changes in this model. We were also surprised to see only modest decreases in print intensity. In our rat model of cervical contusion-compression injury, there is a significant drop in print intensity. 11 Decreases in print intensity have been correlated with increased mechanical allodynia in models of SCI. 32,33 One possibility is that CatWalk analysis alone is not sensitive enough to detect subtle increases in mechanical allodynia, and, as a consequence, print intensity did not show dramatic changes in our study. However, to confirm this, future studies using von Frey testing of the forelimbs and hindlimbs in cervically injured mice will be required. In summary, the limited natural recovery we see across all behavioral measures is consistent with other models of contusive SCI and is, most likely, a result of widespread damage to ascending and descending sensorimotor pathways. Most important, our neurobehavioral analysis supports the clinical relevance of this model in two main ways. First, this injury creates significant neurological deficits that would be able to show a treatment effect in studies on putative therapies. Next, although natural recovery is not dramatic, there are slight and occasionally significant improvements that occur during the subacute phase of injury, which is a clinically relevant time point for therapeutic interventions.
We followed the weight of our animals over the recovery period in order to rule out the possibility that lack of natural recovery in behavioral tests was attributed to weight loss or an inability to gain body weight. 34,35 Injured animals displayed a significant weight gain at 8 weeks post-SCI, indicating that failure to gain weight or weight loss is not responsible for limited functional recovery.
This injury model was designed to create forelimb deficits by affecting motor neuron pools in the C6 region of the spinal cord that control hand and arm function. Retrograde tracing studies have demonstrated that the ventral horns of the gray matter in the C7 to C5 segments contain motor neurons controlling lower forelimb muscles, including the extensor carpi radialis brevis (dorsal upper forelimb) and the flexor carpi radialis (ventral lower forelimb). 36,37 In fact, clip-compression injury resulted in the most significant tissue damage to the ventral gray matter. Therefore, we can link deficits in forelimb function to loss of innervation of muscles of the lower forelimb.
In addition to direct loss of motor neurons, neurobehavioral deficits are a result of the disruption of a number of targets, including sensorimotor pathways, second-order sensory neurons, and interneurons. Our analysis shows that there is significant loss of dorsal CST fibers caudal to the injury as early as 7 days post-SCI. Limited regeneration of the CST is likely a key factor in the minimal natural recovery we see in our model. The lack of natural recovery also suggests that spared pathways, including the rubro- and reticulospinal tracts, are not able to compensate for damage to the CST. Further retro- and anterograde tracing studies are required to determine the degree of sparing in these pathways.
In conclusion, this model addresses the lack of clinically relevant bilateral cervical injury models in the mouse. Although similar weight-drop models have been characterized, ours is the first to use clip compression to achieve a bilateral injury at C6 in mice. Further, we present the first detailed gait analysis in a bilateral cervical contusion injury in the mouse. We clearly demonstrate that this model will be a useful tool for pre-clinical testing of therapies for cSCI by showing that modest natural recovery occurs during the clinically relevant subacute period, and that behavioral deficits are significant enough to reveal treatment effects. Finally, increased emphasis on the development of mouse models of cSCI will allow for future studies using transgenic animals, which will provide mechanistic insights that are critical for the identification of therapeutic targets.
Footnotes
Acknowledgments
The authors acknowledge Sadiya Yousef and Behzad Azad for their work on the animal care required for these experiments. Priscilla Chan is acknowledged for her contribution to the qRT-PCR analysis.
Author Disclosure Statement
No competing financial interests exist.