
Abstract
Reactive astrogliosis has occurred after intracerebral hemorrhage (ICH). Leukemia inhibitory factor (LIF) can act as a modulator for glial gene expression. Signal transducer and activator of transcription 3 (STAT3) is a critical regulator of reactive astrogliosis. The present study tested whether endogenous LIF acted on ICH-induced reactive astrogliosis via the STAT3 signaling pathway. Rats were divided into three experimental groups: 1) Rats received either an ICH or a needle insertion (sham), 2) Rats received 100 ng LIF or an equal volume of phosphate-buffered saline (PBS) by direct infusion into the lateral ventricle (LV) after ICH, and 3) AG490 (0.25 mg/kg) was injected into the LV to block STAT3 signaling. Brains were perfused to identify proliferating cell nuclear antigen (PCNA)+/glial fibrillary acidic protein (GFAP)+nuclei. The expression of GFAP, LIF, LIF receptor (LIFR), glycoprotein 130 (gp130), and phospho-STAT3 (p-STAT3) was evaluated by immunohistochemistry and Western blot, respectively. After ICH, the number of the PCNA+/GFAP+ nuclei and the expression of GFAP, LIF, LIFR, gp130, and p-STAT3 were increased. Moreover, LIF increased the number of PCNA+/GFAP+ nuclei and the expression of GFAP, LIFR, gp130, and p-STAT3. The number of PCNA+/ GFAP+ nuclei and GFAP protein levels were attenuated markedly after inhibition of p-STAT3. Together, these data suggest that LIF contributes to ICH-related reactive astrogliosis via activation of STAT3 signaling.
Introduction
I
After central nervous system (CNS) injury, reactive astrogliosis occurs with increased expression of specific markers, such as glial fibrillary acidic protein (GFAP) and various extracellular matrix molecules such as chondroitin sulphate proteoglycans (CSPGs) and in severe cases, glial scar is formed surrounding the injury site. 4 Glial scar not only creates a physical barrier for axonal sprouting, but also produces CSPGs, which can convert the axons into a dystrophic state, and leads to regeneration failure. 5 Therefore, a favorable environment could be achieved by modulating reactive astrogliosis and growth inhibitory molecule production, which should be beneficial for functional recovery after ICH; however, cellular and molecular mechanisms leading to the process after ICH remain poorly defined.
Leukemia inhibitory factor (LIF) has been proved to regulate astrocyte activation via modulating GFAP expression both in vitro and in vivo during development and in adulthood as well as following cortical injury. 6 –8 The cell-type specific effects of LIF are mediated by a heterodimeric complex of LIF receptor (LIFR) and glycoprotein 130 (gp130). 9 Recent studies have demonstrated that expression of LIF and its receptor components is upregulated after cerebral ischemia; 10,11 however, the alteration of LIF and its receptor components after ICH have been poorly understood.
Among signaling pathways for modulating reactive astrogliosis, signal transducer and activator of transcription 3 (STAT3) has gained the most attention. 12 In the CNS, STAT3 is expressed by astrocytes, neurons, and other cell types, 13 and activation of STAT3 by phosphorylation increases markedly after CNS insults. 14 –17 After cerebral ischemia, the induction of phosphorylated STAT3 was significantly detected on the ischemic side in the high-LIF group after LIF injection. 11 However, it is unclear what effect LIF has on reactive astrogliosis after ICH and whether STAT3 signaling pathway involves the astrogliosis. Accordingly, we aimed to elucidate the effects of endogenous LIF on ICH-induced reactive astrogliosis and to further define the signaling pathways.
Methods
Animal preparation
Adult male Sprague–Dawley (SD) rats weighing 250–300 g were housed under diurnal lighting conditions. This study was conducted in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH Publication No. 85–23, revised 1996). All experiments were approved by the Institutional Animal Care and Use Committee of Central South University (201403164).
Surgical procedure of ICH
ICH was conducted as described in detail previously. 18 Animals were anesthetized with an intraperitoneal injection of chloral hydrate (400 mg/kg), and then were fixed onto a stereotactic frame (Stoelting) in the prone position. After an incision was made on the scalp, a small cranial burr was drilled near the right coronal suture 3.2 mm lateral to the midline. A 26-gauge needle was inserted stereotaxically into the right globus pallidus (1.4 mm posterior and 3.2 mm lateral to bregma, 5.6 mm ventral to the cortical surface), and 100 μL of non-heparinized autologous whole blood was taken from the femoral artery and was injected into the globus pallidus over 2 min, then the needle was slowly removed over another 10 min to prevent backflow.
Experimental designs
Rats were randomly divided into three groups. Group 1: the rats were administered 100 μL autologous blood (ICH group) into the right globus pallidus (n = 60). Sham controls received only a needle insertion (n = 20). Group 2: the rats received 100 ng LIF (Chemicon, Temecula, USA, 100 ng dissolved in phosphate-buffered saline [PBS] for a total volume of 10 μL, n = 20) or an equal volume of PBS (control group, n = 20) by direct infusion into the right lateral cerebral ventricle (LV) (bregma; 0.8 mm posterior, −4.8 mm dorsoventral, −1.5 mm lateral) immediately after ICH. 11 Group 3: to further define the role of STAT3 in reactive astrogliosis in vivo, AG490 (Sigma-Aldrich, St. Louis, MO) was used to block STAT3 signaling and 2 mg AG490 was dissolved in 1 mL dimethyl sulfoxide (DMSO). AG490 (0.25 mg/kg) was stereotactically injected into the LV immediately after ICH (n = 20), and the control group received an equal volume of vehicle (n = 20). 19
Specimen preparation
Animals were deeply anesthetized with chloral hydrate (800 mg/kg). For immunohistochemistry, animals (n = 10 per time point) were transcardially perfused with 0.9% saline followed by 250 mL ice-cold 4% paraformaldehyde in 0.1 M phosphate buffer (PB) (pH = 7.4). The excised brains were post-fixed in the same fixative for 2 h, and then embedded in paraffin. All tissues were cut into 5 μm coronal sections. For Western blot, after rats (n = 10 per time point) were perfused, the brains were immediately removed, and the tissues in the striatum adjacent to the hematoma were dissected and stored at −196°C in liquid nitrogen.
Functional assessment
According to the literature reported by Hua and coworkers, 20 the use of a forelimb asymmetry test was adopted. Every rat was laid in a transparent cylinder 20 cm in diameter and 30 cm in height. A mirror was placed near the cylinder at an angle convenient for observing the forelimb movement of the rat and at the same time, simultaneous recording was performed by video camera. The test lasted 10 min. During the testing time, behavior of the rat was quantified by counting the occasions that the rat's forelimb touched the cylinder wall while it was in an orthostatic position with a balanced gravity center. The occasions of contact with the unimpaired (ipsilateral) forelimb were recorded as I, those with the impaired forelimb (contralateral to the blood injection site) were recorded as C, and those with both forelimbs were recorded as B. Then the forelimb asymmetric use rate (AUR) was calculated by the following formula: AUR = [I/(I+C+B)]-[C/(I+C+B)].
Immunohistochemistry
Expression of GFAP, chondroitin sulphate proteoglycans (CSPGs), and microtubule-associated protein 2 (MAP2)
Immunohistochemistry was performed as previously described. 21 The primary antibodies were mouse anti-GFAP (1:500, Santa Cruz Biotech, CA), mouse anti-CSPG (monoclonal, 1:100, Sigma-Aldrich), or rabbit anti-MAP2 (a marker for axonal growth and synaptic plasticity, monoclonal, 1:100, Santa Cruz Biotech). AF488-conjugated goat anti-mouse antibody (1:100, Jackson Immuno Research Laboratories, West Grove, PA) and Cy3-conjugated goat anti-rabbit antibody (1:100, Jackson Immuno Research Laboratories) were used as secondary antibodies. 4, 6-diamino-2-phenyl indole (DAPI, 10 mg/mL, Sigma-Aldrich) was included in the secondary antibody incubation for labeling nuclei. Finally, the slides were washed and cover-slipped with antifade mounting media. The sections were observed under an Olympus BX51 fluorescent microscope (Olympus, Japan) connected with a computer screen.
Proliferation of GFAP
Cell proliferation was determined based on the immunostaining of the endogenous cell proliferative marker-proliferating cell nuclear antigen (PCNA), and activated astrocytes were determined based on the immunoreactivity of GFAP. Double immunolabeling was performed. The sections were first incubated for 1 h at room temperature with a mixture of two primary antibodies against mouse anti-GFAP (1:250) and rabbit anti-PCNA (1:250, Santa Cruz Biotech). AF488-conjugated goat anti-mouse antibody (1:100) and Cy3-conjugated goat anti-rabbit antibody (1:100) were used as secondary antibodies. DAPI was used to label nuclei. These sections were scanned using a laser scanning confocal microscope (LSM-510, Zeiss). For a negative control, 1% bovine serum albumin (BSA) was used instead of the primary antibody in each experiment.
PCNA+/GFAP+ nuclei close to the hematoma were counted in four 250 × 250 μm areas in 10 sections through the stroke for each animal at × 40 objective magnification with Motic Images Advance 3.2 image analysis software by a researcher blind to the experimental cohort, and the data were presented as the number of nuclei per mm2 (N/mm2).
Expression of LIF, LIFR, gp130, and phospho-STAT3 (p-STAT3)
To observe spatial profiles of LIF, LIFR, gp130, and p-STAT3, immunohistochemistry was used for detection their expression. 22 The primary antibodies (all obtained from Santa Cruz Biotech) were used as follows: rabbit anti-LIF (polyclonal, 1:100), rabbit anti-LIFR (polyclonal, 1:100), mouse anti-gp130 (monoclonal, 1:100), and goat anti-p-STAT3 (Tyr 705) (polyclonal, 1:100).
To determine whether LIF, LIFR, gp130, and p-STAT3 were co-expressed with GFAP, immunofluorescence double labeling was used. The sections were first incubated for 1 h at room temperature with a mixture of two primary antibodies against mouse (or rabbit) anti-GFAP (1:250) and one of the following antibodies: LIF (1:50), LIFR (1:100), gp130 (1:100), and p-STAT3 (Tyr705) (1:50). AF488-conjugated goat anti-mouse antibody (1:100) and Cy3-conjugated goat anti-mouse, goat anti-rabbit, and donkey anti-goat antibodies (1:100) were used as secondary antibodies. DAPI (10 mg/mL) was used to label nuclei. These sections were scanned using a laser scanning confocal microscope (LSM-510, Zeiss). For a negative control, 1% BSA was used instead of the primary antibody in each experiment.
Western blot
To detect the levels of GFAP, LIF, LIFR, gp130, and p-STAT3 (Tyr705), protein was extracted from the tissues in the striatum adjacent to the hematoma, and Western blot analysis was performed. 10 Briefly, 50 μg proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to a Hybond-C pure nitrocellulose membrane (Amersham). Monoclonal mouse antibodies for GFAP (1:1000) and gp130 (1:500), polyclonal rabbit antibodies for LIF (1:250) and LIFR (1:500), polyclonal goat antibodies for p-STAT3 (Tyr705) (1:250), and β-actin (1:200, Santa Cruz Biotech, CA) were used. Quantitative densitometric analysis of Western blots was performed with a computerized digital image analysis system.
Statistical analysis
All data are expressed as mean ± standard deviation and analyzed with Student's t test and one way analysis of variance (ANOVA), followed by Scheffe's post-hoc test. Differences were considered significant at p < 0.05.
Results
ICH induced reactive astrogliosis
In the sham control group, astrocytes were characterized by small bodies and slender processes. After ICH, several astrocytes in the perihematoma region became activated with hypertrophied morphology, and the reactive phenotype with enlargement of the cell bodies was extensively overlapping other at day 14. Meanwhile, the results of Western blot showed that GFAP level was weak in the sham group. In contrast, GFAP level was increased notably at day 3 compared with the control group and reached a significant high level at day 14 post-ICH (Fig. 1A, p < 0.05).

Intracerebral hemorrhage (ICH) induced reactive astrogliosis.
Next, we determined whether an increase in GFAP immunoreactivity was associated with increased cell proliferation after ICH. In the sham control group, PCNA-positive nuclei were sporadically observed, and few PCNA+/GFAP+ nuclei could be detected. However, the nuclei stained with PCNA appeared in GFAP-positive activated astrocytes at day 3, and reached a maximal level at day 14 post-ICH (Fig.1B, p < 0.01).
Finally, the spatial distributions of CSPG and MAP2 were observed by Immunofluorescent double labeling. The results showed that a diffuse extracellular deposit of CSPG was seen close to the lesion site. Surprisingly, most MAP2 immunoreactivity was encompassed by CSPG (Fig. 2).
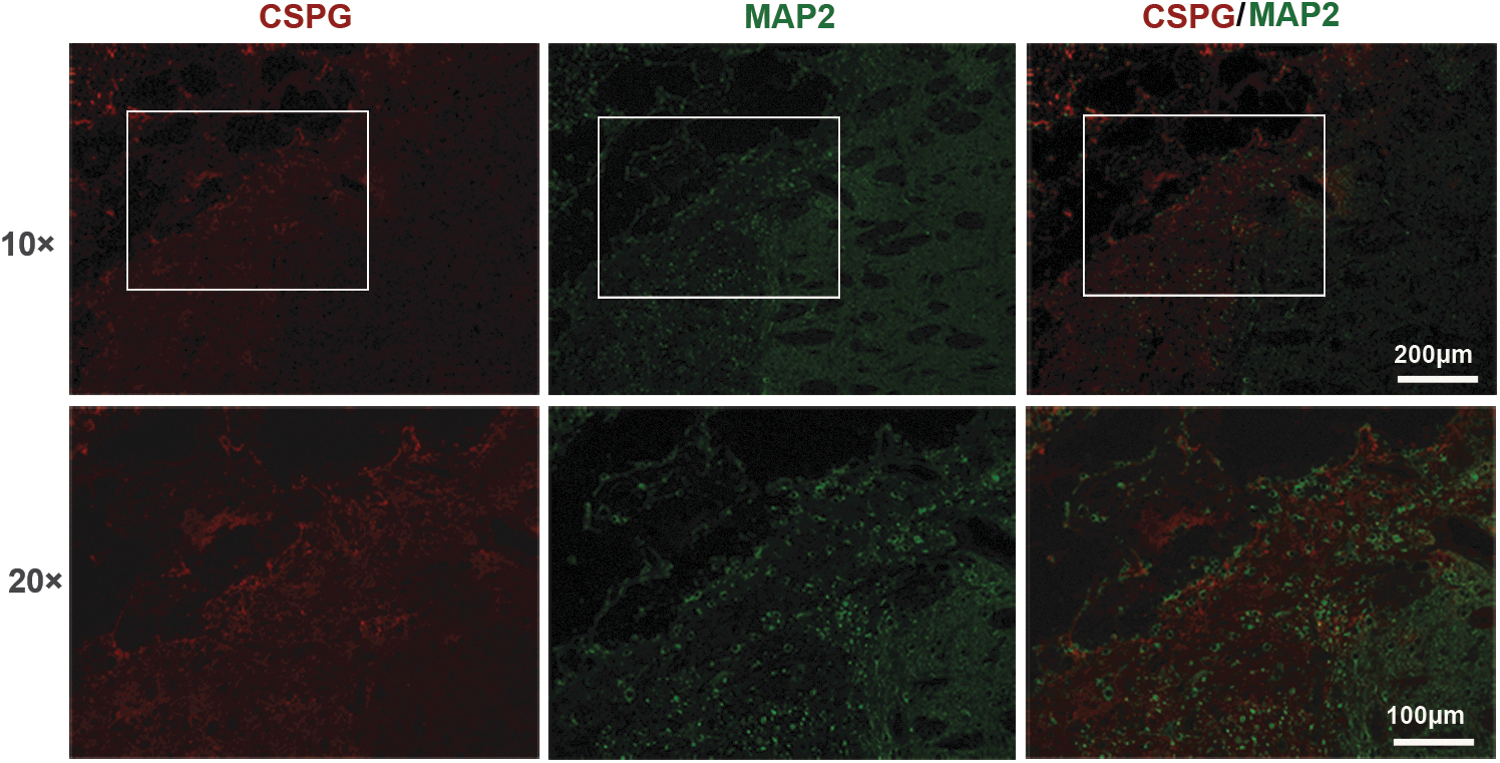
The spatial distributions of chondroitin sulphate proteoglycans (CSPGs) and microtubule-associated protein 2 (MAP2) after intracerebral hemorrhage (ICH). Immunofluorescent double labeling showed distribution of CSPG (red) and MAP2 (green) 14 days after ICH. Scale bar = 100 μm (20 × ), 200 μm (10 × ).
ICH induced increased expression of LIF, LIFR, gp130, and p-STAT3
In the sham group, immunoreactivity of LIF, LIFR, and gp130 was unclearly detected in any regions, and some sporadic p-STAT3-positive cells were observed. After ICH, LIF-, LIFR-, gp130-, and p-STAT3 (Tyr 705)-positive cells were detected at day 3. Immunoreactivity of LIF, LIFR, and gp130 was confined to the cytoplasm of cells in the perihematoma region, whereas p-STAT3 (Tyr 705) immunoreactivity was mainly located in nuclei, and double immunolabeling showed that immunoreactivity of LIF, LIFR, gp130, and p-STAT3 (Tyr 705) was co-localized with GFAP-immunoreactive astrocytes (Fig. 3A).
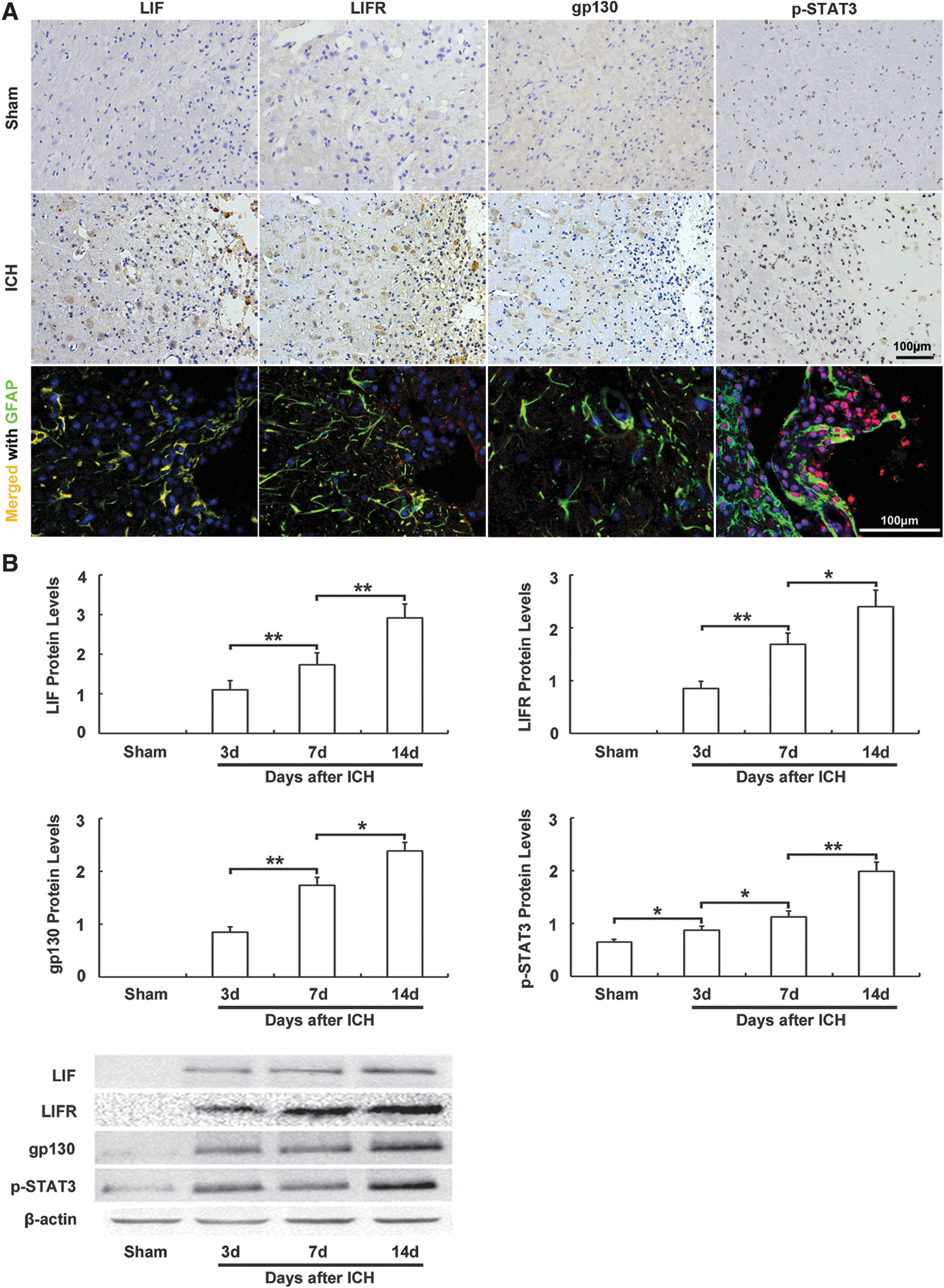
Expression of leukemia inhibitory factor (LIF), LIF receptor (LIFR), glycoprotein 130 (gp130), and phospho-signal transducer and activator of transcription 3 (p-STAT3) after intracerebral hemorrhage (ICH).
As there were significant changes in immunoreactivity of LIF, LIFR, gp130, and p-STAT3 (Tyr 705) around the hematoma, we further evaluated their protein levels at the basal ganglion ipsilateral to the ICH by Western blot. In sham-operated animals, no LIF, LIFR, or gp130 signal could be detected. After ICH, the levels of LIF, LIFR, gp130, and p-STAT3 (Tyr 705) were increased persistently from day 3 to day 14 (Fig. 3B, p < 0.05)
LIF contributed to reactive astrogliosis; upregulation of LIFR, gp130, and p-STAT3; and neurotoxicity after ICH
To further examine the effect of LIF on the reactive astrogliosis, exogenous LIF was infused into the LV. The present study showed that LIF increased the number of PCNA+/GFAP+ nuclei, and upregulated GFAP protein levels (Fig. 4A, p < 0.01).

Leukemia inhibitory factor (LIF) contributed to reactive astrogliosis; upregulation of LIF receptor (LIFR), glycoprotein 130 (gp130), and phospho-signal transducer and activator of transcription 3 (p-STAT3); and neurotoxicity after intracerebral hemorrhage (ICH).
We next evaluated the effect of LIF on the expression of LIFR, gp130, and p-STAT3. Exogenous LIF notably unregulated the expression of LIFR, gp130, and p-STAT3 (Tyr 705) after ICH (Fig. 4B, p < 0.05).
Finally, the effect of LIF on neurological deficits was estimated. As showed in Figure 4C, the ICH group was distinctly more impaired than the sham group at the corresponding time points (p < 0.01). When rats were treated with exogenous LIF, the AUR of the ICH+LIF group was significantly higher than that of ICH+vehicle group (p < 0.05).
STAT3 was required for reactive astrogliosis
To further define the role of STAT3 in reactive astrogliosis in vivo, AG490 was used to block STAT3 signaling. After AG490 treatment, the number of PCNA+/GFAP+ nuclei was decreased significantly, and GFAP expression was downregulated as well (Fig. 5, p < 0.01).

Signal transducer and activator of transcription 3 (STAT3) was required for reactive astrogliosis. The number of proliferating cell nuclear antigen (PCNA)+/glial fibrillary acidic protein (GFAP)+ nuclei and GFAP protein levels in the ipsilateral basal ganglia at 14 days after injection of AG490 or vehicle (control) into the lateral ventricle. (**p < 0.01, n = 10).
Discussion
In the present study, we found that not only upregulation of GFAP and the morphological appearance of reactive astrogliosis, but also increased PCNA+/GFAP+ activated astrocytes were prominently detected within the perihematoma tissue after ICH, which suggested the presence of reactive astrogliosis, which is consistent with previous studies. 23 –25 Upregulation of GFAP in reactive astrocytes is perhaps the best-known hallmark of reactive astrogliosis and glial scar, which consists predominately of reactive astrocytes, microglia, and extracellular matrix molecules, especially CSPGs. Knockout mice lacking GFAP presented lower than normal levels of astroglial activity, with a reduced astrocytic hypertrophy and a defective scar formation in vivo. 23 However, so far, the effect of reactive astrogliosis and glial scar in the injured brain or spine remains controversial. 26 The reactive process is reported to be neuroprotective, 27 and functions in isolating and protecting the uninjured tissue from injury, inhibiting the spread of inflammation and regulating the extracellular milieu in certain conditions, 28 but excessive reactive astrogliosis is detrimental and contributes to neuronal damages. 29 Further, Munakata and coworkers 25 reported that inhibition of transient receptor potential canonical 3 could attenuate astrogliosis and improve outcomes after ICH. Although the functional significance of reactive astrogliosis was not addressed in the present study, we observed that most MAP2 immunoreactivity was encompassed by CSPG. Accordingly, it was supposed that CSPG associated with reactive astrogliosis clearly inhibited neurite outgrowth. A body of evidence has shown that reactive astrogliosis leads to formation of glial scar at the lesion site and overproduction of CSPGs, which hamper axonal regeneration and reconstruction of circuitry as a local biochemical and physical barrier. 30 In contrast, attenuation of reactive astrogliosis can enhance locomotion recovery following spinal cord injury (SCI). 31
LIF, as a member of the interleukin-6-type cytokine family, exerts its effect on binding to LIFR and gp130. 9 Herein we observed a strong upregulation of LIF, LIFR, and gp130, primarily within GFAP-positive astrocytes, beginning at 3 days post-ICH, which was consistent with previous studies. 10,11 Therefore, it was speculated that astrocyte-specific expression of LIF bound to the receptors on astrocytes through an autocrine means. We next demonstrated that LIF upregulated the expression of LIFR/gp130 as well as GFAP. Moreover, previous studies showed that LIF-, LIFR-, or gp130-deficient mice had a diminished astrocytic response compared with wild type (WT). 8,32,33 Based on these findings, we speculated that the LIFR/gp130 induced by LIF injection had a significant effect on reactive astrogliosis after ICH. LIF has been known to be neuroprotective against cerebral ischemia. 10 However, LIF overexpression caused the development of severe hindlimb motor dysfunction after SCI, 9 and our data indicated that LIF was detrimental against ICH. One possible explanation for this discrepancy may be in the different stroke models used, and LIF may exert pleiotropic effects on cell function in different biological contexts.
We further defined the signaling pathways for reactive astrogliosis induced by LIF, and found that the upregulation of p-STAT3 was closely correlated with the expression of gp130, and that LIF increased gp130 and p-STAT3 protein levels. Activation of the gp130 homodimer could lead to the initiation of STAT signaling, 34 whereas inhibition of gp130 abolished STAT3 activation. 35 Therefore, it was supposed that LIF might regulate the expression of p-STAT3 through gp130. STAT3 is a critical regulator of astrogliosis and scar formation after SCI, and reactive astrogliosis required STAT3 phosphorylation on Tyr-705 but not on Ser-727. 36,37 p-STAT3 dimerizes, translocates to the nucleus, binds to the GFAP promoter, and, finally, promotes the transcription of GFAP. 38 Deletion of STAT3 significantly attenuated astrocyte activation after SCI. 34 After cerebral ischemia, STAT3 is activated in GFAP+ astrocytes. 39 Consistent with these studies, the current study observed that p-STAT3 was mainly located to nuclei and expressed in GFAP+ astrocytes. Inhibiting STAT3 phosphorylation led to a marked decrease of PCNA+/GFAP+ activated astrocytes and downregulation of GFAP. Taken together, our data suggested that STAT3 signaling was required for the reactive astrogliosis induced by ICH.
Conclusion
We demonstrated for the first time that ICH led to a prolonged, sustained upregulation of LIF and its receptor component expression that may contribute to the development of reactive astrogliosis via activation of STAT3 signaling (Fig. 6). This information could lead to new approaches to modulating the time span and severity of reactive astrogliosis, consequently fine-tuning neuronal tissue repair and regeneration.
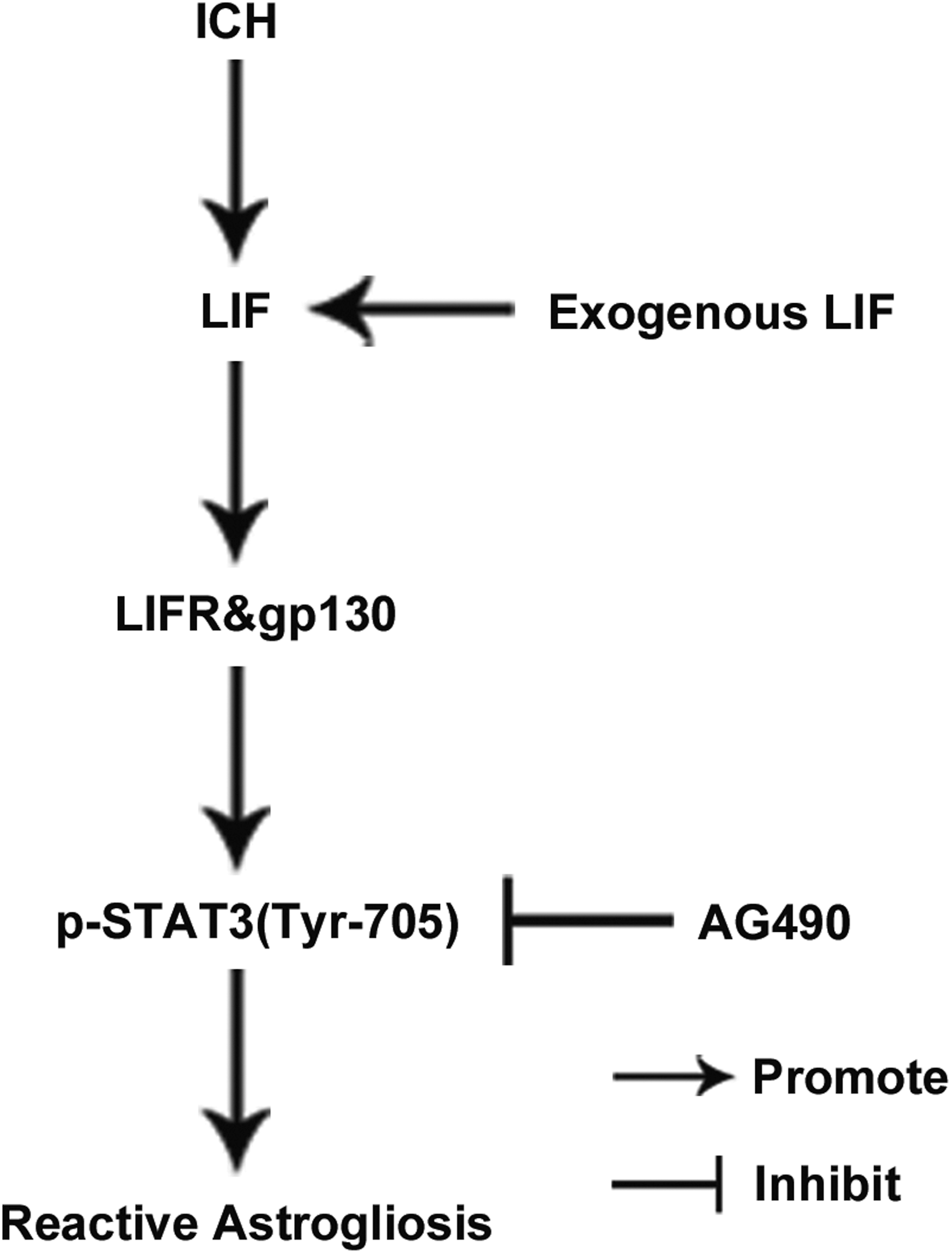
The upregulation of leukemia inhibitory factor (LIF) plays a crucial role in reactive astrogliosis via activation of signal transducer and activator of transcription 3 (STAT3) signaling following intracerebral hemorrhage (ICH).
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 30400581, 30873221, 81173175, and 81202625), the Project for New Century Excellent Talents (NCET-11-0522), and the Hunan Provincial Natural Science Foundation (Grant Nos. 07JJ5007 and 10JJ2023). Data supporting the conclusions of this article are presented in the manuscript. Conception and design: H.J. Zhou and T. Tang; acquisition of data: X. Yang, H.J. Cui, A.L. Yang, and J.K. Luo; analysis and interpretation of data: H.J. Zhou, T. Tang; drafting the article: H.J. Zhou; critically revising the article: T. Tang, J.K. Luo, Q.M. Zhang, J.H. Zhou, and Q. Zhang; reviewed final version of the manuscript and approved it for submission: H.J. Zhou, T. Tang, X. Yang, H.J. Cui, J.K. Luo, Q.M. Zhang, J.H. Zhou, and Q. Zhang; statistical analysis: H.J. Zhou; study supervision: T. Tang.
Author Disclosure Statement
No competing financial interests exist.