
Abstract
Ventral root avulsion induces dramatic loss of the affected spinal cord motoneurons. The neuroprotective effect of riluzole has been previously proven on the injured motoneurons: the vast majority of them can be rescued even when they have no possibility to regenerate their axons. In this study the number of injured motoneurons rescued by riluzole treatment and their capacity to reinnervate the denervated forelimb muscles was investigated. Surgical reconnection with a peripheral nerve graft between the affected spinal cord segment and the C7 spinal nerve was established immediately or with 1- and 3-week delay after avulsion. Avulsion and immediate reconnection of the motoneuron pool to the spinal nerve resulted in moderate reinnervation of the spinal nerve (281 ± 23 standard error of mean [SEM] retrogradely labeled motoneurons), whereas treatment of the injured motoneurons with riluzole yielded considerably higher numbers of reinnervating motoneurons (548 ± 18 SEM). Reconnection of the motor pool with the C7 spinal nerve with 1-week delay allowed fewer motor axons to reinnervate their targets in control and riluzole-treated animals (159 ± 21 vs. 395 ± 16 SEM). A clinically relevant 3-week delay in reconnection further reduced the number of reinnervating motoneurons (76 ± 22 SEM), but riluzole pre-treatment still enabled a significant number of rescued motoneurons (396 ± 17 SEM) to regenerate their axons into the C7 spinal nerve.
These results show that those injured adult motoneurons that are rescued by riluzole treatment started immediately after the avulsion injury are able to reinnervate their targets even if they are provided with a conduit several weeks after the primary injury. This finding suggests that partial rescue of injured motoneurons with riluzole in patients who suffered a brachial plexus avulsion injury may provide an available pool of surviving motoneurons for late reconnection/reimplantation surgeries.
Introduction
A
The time window between avulsion and the reconnection of the remaining motor pool with the peripheral targets appears to be a crucial factor for a successful outcome. 26 By 2 weeks after avulsion injury the number of surviving motoneurons dramatically decreases in the affected ventral horn of experimental animals. 4,9,10 In the cases of human brachial plexus injuries the main attempt is to stabilize the polytraumatized patient, and it is not always even possible to diagnose early the neurological deficit of the plexus and initiate a restorative plexus surgery. Accordingly, it may take several weeks before surgical interventions may be carried out to connect the surviving motoneurons to their target muscles or nerves.
We have shown in our previous studies that systemic administration of riluzole in animals that had their lumbar 11 or cervical 9 ventral root avulsed and reimplanted prevented the death of the majority of motoneurons even if the onset of the treatment was delayed by 10 days. 10 Moreover, evidence has also been provided that motoneurons with avulsed axons survive even in cases when their axons are prevented from regeneration. These recent findings suggest that a vast majority of motoneurons can be rescued from cell death initiated by the interruption of their whole axon. Moreover, their perikarya may retain the capacity to regenerate their axons at later time-points provided the cell body remains intact. 9 The aim of our study was to reveal whether a significant portion of the spinal C7 motoneuron pool rescued by riluzole after ventral root avulsion can be used for the reinnervation of the brachial plexus. Moreover, the study aimed at showing that the denervated forelimb muscles achieve satisfactory functional reinnervation after 1- or 3-week delay in reconnection of the motor pool with the C7 spinal nerve.
Methods
Surgery
Altogether 35 Sprague-Dawley female rats (weight at time of surgery: 170–180 g; Biological Services, University of Szeged) were used in this study. Five intact animals were used for counting the C7 motoneuron pool. In 30 animals the right C7 ventral root was avulsed from the cord and these animals were used to set up six experimental groups, each group consisting of five animals (Fig. 1).

Schematic diagrams show the experimental procedures and the time course of various interventions in the different experimental paradigms.
All the operations were carried out under deep ketamine-xylazine combination anesthesia (ketamine hydrochloride: 90 mg/kg body weight, Ketavet, Pharmacia & Upjohn Co.; xylazine: 5 mg/kg body weight, Rompun, Bayer Co.). Animals received post-operative pain therapy in form of a single daily dose of meloxicam (0.75 mg/kg body weight, Metacam, Boehringer Ingelheim) administered for 3 days post-operatively.
In all experimental animals a laminectomy was performed under a surgical microscope at the level of the C5–6 vertebrae. The identification of these vertebrae was based on the location of the T2 vertebra, equipped with a long spinous process. The dura mater was opened and the right C7 ventral root was pulled out after cutting the C7 dorsal root (Fig. 1A). To prevent regeneration into the avulsed root, it was bent and placed further away from the site of injury. In groups 1 and 2 (immediate reconnection groups; group 2 animals received riluzole treatment) the ventral ramus of the C7 spinal nerve was dissected from a ventral approach between the anterior and middle scalenus muscles.
The ventral ramus was axotomized and an autologous common peroneal nerve graft (about 25 mm in length, harvested from the right hindlimb of the same animal) was placed between the spinal cord and the C7 ventral ramus to establish a conduit for the regenerating motor axons (Fig 1A). The medial stump of the nerve graft was implanted ventrolaterally into the spinal cord just dorsally to the original exit zone of the C7 ventral root. To avoid damage to the cord, a small myelotomy groove was created on the ventrolateral surface of the cord and an 0.5-mm long segment of the nerve graft was gently inserted into the hole by using a watchmaker's forceps. Special care was taken to avoid damage to the motoneuron pool or to the implanted nerve segment during this procedure. The distal stump was sutured to the C7 ventral ramus with 10-0 Ethilon sutures (Ethicon). The spinal cord was covered with the remaining dura, the wound was closed, and the animals were allowed to recover. Removal of the common peroneal nerve for autografting from the right hindlimb did not cause serious long-term deficit in the locomotor pattern of the hindlimb and did not affect the motor performance of the forelimbs.
To reproduce clinically relevant situations in the experimental paradigms, in the other experimental groups the peroneal nerve grafting (reconnection of the motor pool and the freshly cut C7 spinal nerve with a freshly harvested peroneal nerve graft) was carried out with a delay of 1 week (groups 3–4) and 3 weeks (groups 5–6) after avulsion injury under the same anesthesia protocol as described above (Fig. 1B–D). Animals in groups 2, 4, and 6 received riluzole treatment for 3 weeks starting at the day of avulsion (in group 1 this was identical with the date of reconnection). Animals in groups 1, 3, and 5 served as controls for their treatment groups, that is, they received no riluzole treatment (Fig. 1D). The experiments were carried out with the approval of the Committee for Animal Experiments, University of Szeged regarding the care and use of animals for experimental procedures. All the procedures were carried out according to the Helsinki Declaration on Animal Rights.
Riluzole treatment
Animals were treated with riluzole (2-amino-6-trifluoromethoxy-benzothiazole, kind gift of Tocris Cookson Ltd., Langford, UK; 4mg/kg) for 3 weeks. Riluzole treatment started immediately on the day of avulsion surgery and the drug was injected intraperitoneally daily for 1 week and every second day for the next 2 weeks (Fig. 1D). This treatment protocol was based on the successful riluzole treatment described in our earlier publications. 9,11 The dose of riluzole was established from data obtained from our earlier and other laboratories' experiments. 9,11,14,27 –29 It has also been taken into consideration that 5 mg/kg riluzole administered intraperitoneally in rats produces a significant riluzole level in the brain 30 suggesting that this dose is able to induce therapeutic effects. Control animals in groups 1, 3, and 5 received no injections in place of riluzole.
Retrograde labeling and immunohistochemistry
Three months after the reconnection surgery (i.e., peripheral nerve grafting, Fig. 1D) the animals were deeply anesthetized with ketamine-xylazine. On the operated side the ventral ramus of the C7 spinal nerve was sectioned just distal to the coaptation site (Fig. 1E) and the proximal stump of the nerve was covered with a few crystals of Fast Blue (FB; Illing Plastics GmbH, Breuberg, Germany). In two animals both from groups 1 and 2 the forelimb muscles known to receive innervation from the C7 spinal segment 31 (triceps brachii, palmaris longus, flexor digitorum profundus, flexor carpi radialis, extensor digitorum superficialis and profundus, extensor pollicis longus, and extensor carpi ulnaris) were injected with 2% aqueous suspension of DiY 2 days prior to FB labeling. As FB labels the cytoplasm and DiY accumulates in the nucleus of the labeled cells, double retrograde labeling of the same motoneurons could be performed. Five days after the application of FB the animals were re-anesthetized and perfused transcardially with 4% paraformaldehyde in 0.1 mol/L phosphate buffer. A 2-mm long segment of the nerve graft was removed and immersion fixed in 2.5% phosphate-buffered glutaraldehyde (Fig. 1E). The C7 motoneuron pool of intact animals was labeled as described above and these animals were also allowed to survive for 5 days.
The cervical part of the spinal cords, with the reimplanted nerve segment was removed and kept in fixative (4% phosphate-buffered paraformaldehyde) overnight at 4°C. The cords were then immersed in 30% sucrose in phosphate-buffered saline (PBS). Serial 25-μm thick cryostat sections were cut, mounted on gelatinized slides, and examined in an Olympus BX51 fluorescence microscope (Olympus Ltd., Tokyo, Japan). The number of retrogradely labeled cells was determined in 25-μm thick cryostat sections. To avoid double counting of the same neuron present in consecutive sections, the retrogradely labeled neurons were mapped and drawn with the aid of an Olympus camera Lucida (Olympus Ltd., Tokyo, Japan), and their locations within the ventral horn were compared with those of labeled neurons in the previous section. 9,10 Further help for the identification of labeled motoneurons was provided by the presence of cell nuclei, left unlabeled by FB. All sections from the C7 motoneuron pool were used. This method provided similar numbers of motoneurons in the C7 spinal segment as reported by earlier studies (see Results).
Three spinal cords from all groups were then further processed for choline acetyltransferase (ChAT) immunohistochemistry. Sections processed for ChAT immunohistochemistry were pre-incubated in 3% normal goat serum for 1 h, then incubated with a polyclonal goat anti-ChAT antibody (Merck/Chemicon, Hofheim, Germany, 1:100) overnight at room temperature. The immune reaction was completed by using the avidin-biotin technique (reagents were purchased from Vector Labs, Burlingame, CA) and finally tyramide-amplified with the Cyanine3 TSA kit (Tyramide Signal Amplification, Perkin Elmer, Waltham, MA). The number of ChAT-stained motoneurons in the pools where retrogradely labeled cells were found was also determined both on the operated and control sides. Some sections were stained with cresyl violet to assess the morphology of the spinal cord. Sections were photographed using an Olympus DP70 digital camera mounted on the microscope. Digital images were resized and their contrast and brightness adjusted.
Semithin sections
The peripheral nerve graft segments were thoroughly rinsed in PBS, osmicated, dehydrated, and embedded in Durcupan™ ACM (Fluka GmbH). Semithin sections (400-nm thick) were cut on a Leica Ultracut-R ultramicrotome (Leica GmbH) and stained according to Rüdeberg 32 (mixture of 0.1% methylene blue and 0.1% thionine in 0.1 mol/L phosphate buffer). This mixture is preferably used to visualize intact and lesioned nervous tissue. The myelinated axons were counted by using the ImageJ (NIH) open source image analysis program. To gain more information about the extent of myelination in the grafts, the g-ratio (the ratio of inner axonal to the outer diameter) of the myelin sheaths (for details see Chomiak and Hu 33 ) was determined from selected areas of the nerve cross semithin sections.
Functional analysis
The forelimb movements of the operated animals were monitored every week after a 3-week recovery period. The degree of dorsiflexion in the wrist joint and the extent of flexure contracture developed in the same joint were observed, and the pellet reaching test was performed. The grades used for the extent of dorsiflexion and flexure contracture are shown in Table 1. To perform pre-training for the pellet reaching test, rats were briefly trained 1 week before the first operation to reach through a slot in a plexiglass box for food pellets that they grasp and then place in their mouth for eating. Rats were mildly deprived of food for few days before the test. After the operation rats were allowed to regenerate their motor axons into the implanted nerve segment and proximal muscles for 3 weeks and then they were trained again. Pellet reaching was considered successful if the rat was able to grasp the food in on to two reaches and release the food into the mouth. 34 The percentage of the success was determined by 10 trials.
The table details the grading system used to assess the extent of dorsiflexion and contraction in the wrist joint on the operated side throughout the study.
Statistical analysis
The analysis of variance (ANOVA) test along with the Tukey's all pairwise multiple comparison procedure were used to compare the groups of data of parametric nature. The data of the functional tests were computed according to the repeated measures analysis of variance (repeated measures ANOVA) followed by Fisher's least significant difference (LSD) post hoc analysis. All data in this study are shown as mean ± SEM.
Results
Observations on the movement pattern of operated animals
After the first surgery all animals developed a partial paralysis in the operated forelimb. Animals without riluzole treatment, whose C7 ventral root was avulsed and the spinal cord–C7 spinal nerve reconnection was performed after a 1- and 3-week delay (groups 3 and 5) developed a marked atrophy in the extensor musculature of the upper limb. Accordingly, the wrist joint and the toes were fixed in a low-grade flexion contracture by 6–7 weeks after surgery (grade 1 for group 5 and 0.3 for group 3 animals). This contracture further impaired with time up to grade 1.4 and 0.4 (group 5 vs. group 3, Fig. 2). These animals were able to grasp the food in the pellet reaching test with very low efficacy (up to 15% by week 12) and were not able to dorsiflex their wrist joint more than 30% at any time (grade 1 of dorsiflexion). In contrast, animals that had their C7 spinal nerve reconnected to the spinal cord immediately after avulsion (group 1) performed considerably better in the functional tests, for example, they developed minimal contracture and produced a 20% success rate in the pellet reaching test by week 12.
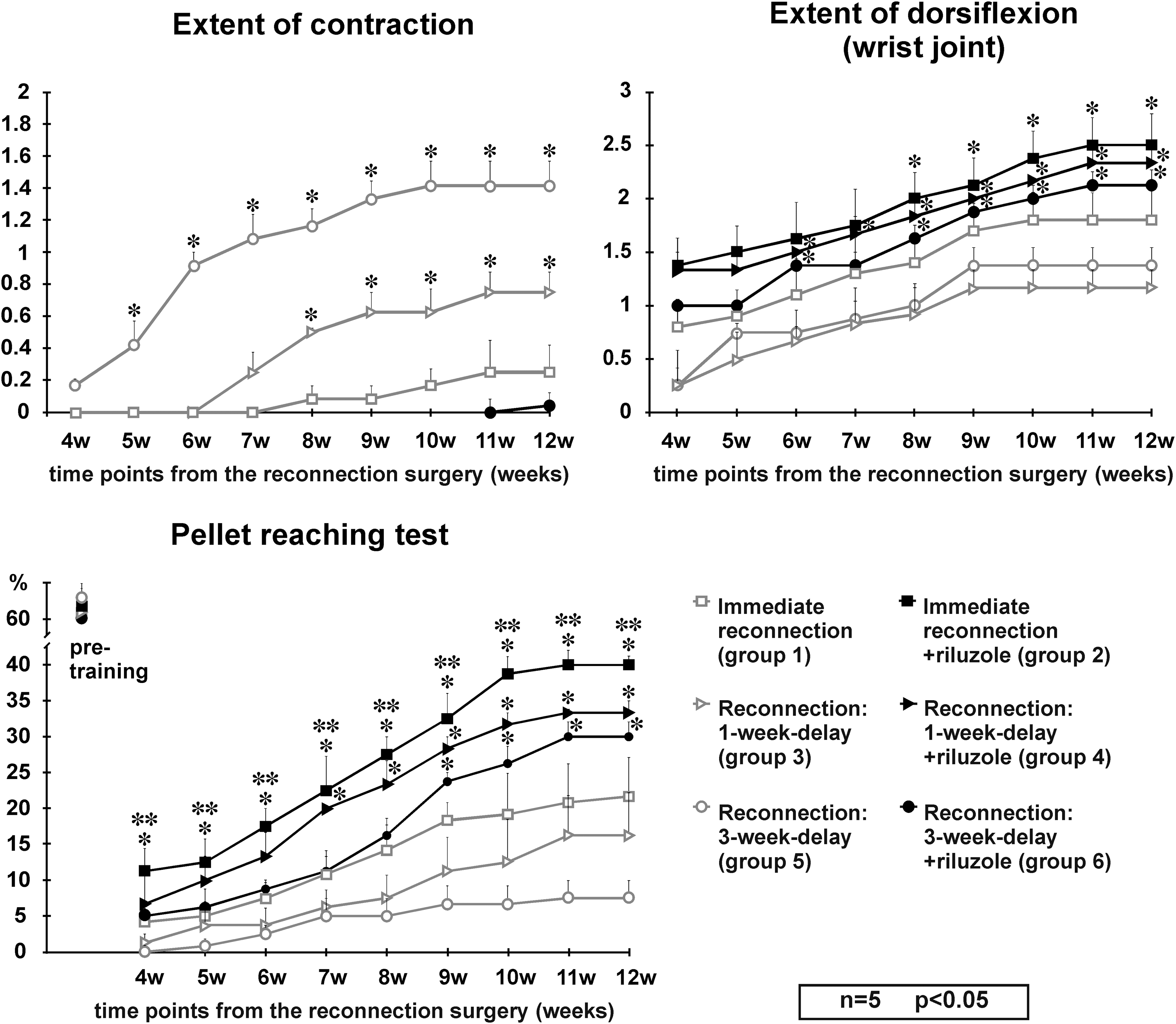
Functional evaluation of experimental animals during their 12-week survival. The upper left panel shows the extent of contracture developed by animals in groups 1, 3, 5, and 6 (group 2 and 4 animals were not included as they did not have any contracture throughout the survival period). Animals that received riluzole treatment were void of contracture except one animal that showed minimal contracture in the wrist joint. Significant difference was found from early time-points onward between the riluzole-treated groups and their controls. The right upper panel displays the extent of dorsiflexion in the wrist joint as a function of time in experimental animals belonging to all groups. Lower panel presents the results of the pellet reaching test in all experimental groups. Note that despite the significant improvement observed in groups 2 and 4, the recovery of this function does not reach the pre-training values (∼62%). *: significant difference between the riluzole-treated groups and their controls; **: significant difference between groups 2 and 6 (immediate reconnection + riluzole treatment vs. reconnection after 3-week delay + riluzole treatment; repeated measures ANOVA followed by Fisher's least significant difference [LSD] post hoc analysis, values are shown as mean ± SEM). ANOVA, analysis of variance; SEM, standard error of the mean.
On the other hand, all the animals that received riluzole treatment and had their C7 motoneuron pool connected to the target muscles (groups 2, 4, and 6) started to recover from paralysis from the 4th week following surgery, but near complete recovery took a few more weeks. By the end of the survival period they were able to walk without major deficit and during locomotion extensively dorsiflexed their wrist joint (grade 2–3). Most of these animals developed no contracture, except for one animal in group 6 which showed minimal contracture of the wrist joint (grade 1, Fig. 2). They produced an extensive dorsiflexion and performed very well in the pellet reaching test (30–40% success rate as compared with the 60% rate of animals in the pre-training phase).
Retrograde labeling: regeneration of the axons of the cervical motoneurons following C7 ventral root avulsion and reconnection surgery
First we have determined the number of resident motoneurons in the C7 motoneuron pool through the use of retrograde labeling of the ventral ramus of the C7 spinal nerve as we performed in our previous study. 9 We found the average number of retrogradely labeled motoneurons in the C7 spinal segment to be 881 ± 35. This number of C7 motoneurons correlated with the motoneuron numbers taken from other publications 35,36 and with that of our earlier counts. 9
The effect of riluzole treatment on injured motoneurons after immediate or delayed reconnection of the damaged C7 motoneuron pool with the C7 ventral ramus was studied in the next series of experiments. In animals whose avulsed C7 ventral root was reconnected straight after avulsion with the C7 spinal nerve and received riluzole treatment (group 2), 548 ± 18 retrogradely labeled motoneurons were found, indicating that nearly two-thirds of the total population of C7 motoneurons were able to grow axons into the re-established C7 spinal nerve (Fig. 3). Without riluzole treatment (group 1) the number of retrogradely labeled neurons decreased to 281 ± 23, and this number correlated well with our earlier findings. 9 Double labeling experiments have shown that in these two groups 78.5 ± 0.7% and 79 ± 0.4% (group 2 and group 1 animals, respectively) of the FB-labeled motoneurons were co-labeled with DiY. This finding indicates that the vast majority of the reinnervating axons present in the nerve graft were able to reach the forelimb muscles and reinnervated them.
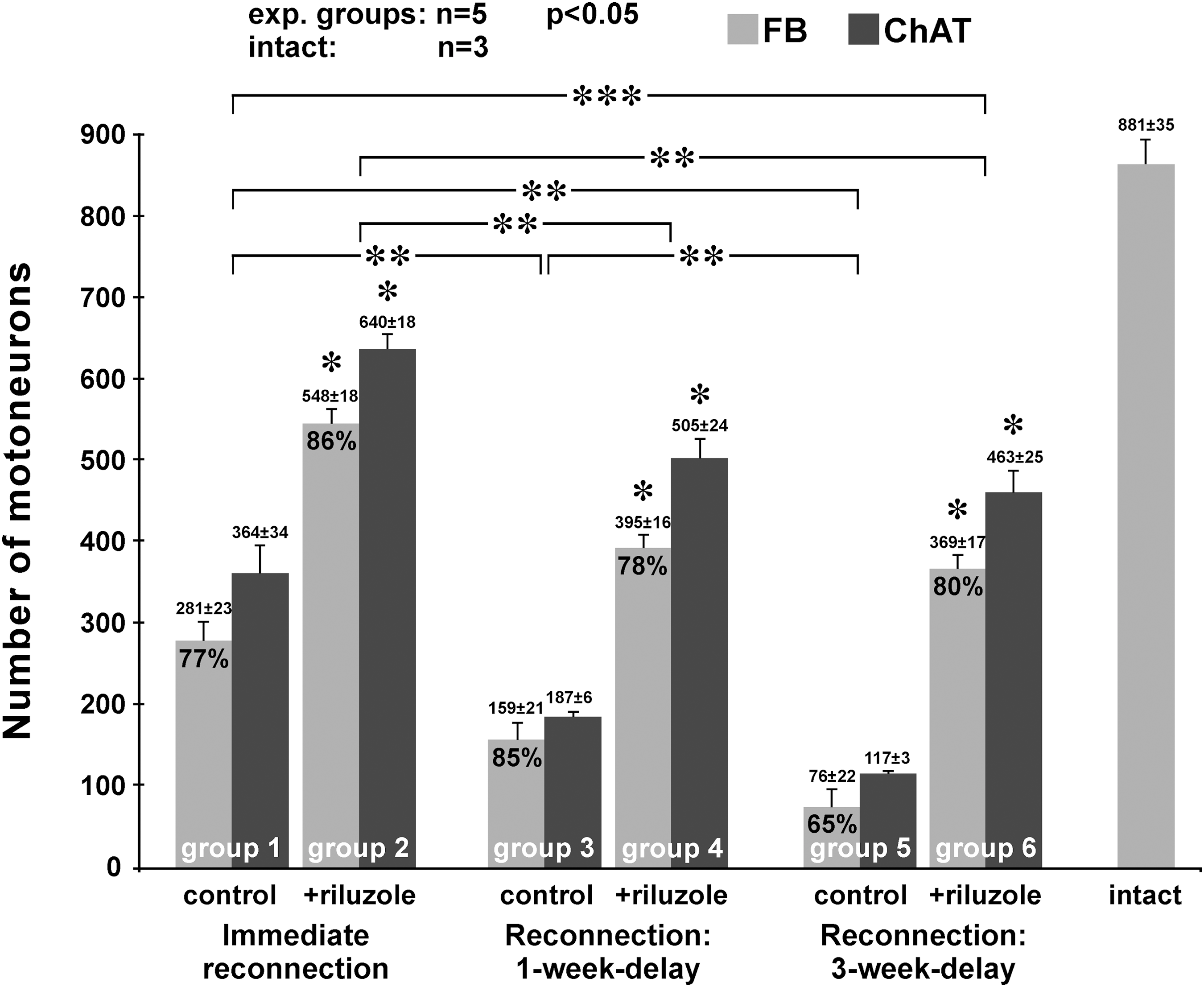
Bar chart shows the number of retrogradely labeled and ChAT-positive motoneurons in various experimental setups. Significantly more retrogradely labeled reinnervating and ChAT-positive surviving motoneurons were found in the riluzole-treated groups compared with their controls (*). ** = significant difference between the various control groups (ctrl to ctrl) and treatment paradigms (riluzole to riluzole). *** indicates a significant difference between the groups of control animals with immediate reconnection and riluzole-treated animals that had their reconnection surgery delayed for 3 weeks, suggesting that even a considerably delayed reconnection strategy results in successful reinnervation when riluzole was applied to preserve the injured motoneuron pool. (ANOVA test computed using Tukey's all pairwise multiple comparison procedures, p ≤ 0.05 for each group). Values are shown as mean ± SEM. ANOVA, analysis of variance; ChAT, choline acetyl transferase; FB, Fast Blue labeling.
In the experiments where the damaged motoneuron pool and the ventral ramus of the spinal nerve was connected after 1 week delay, the procedure resulted in higher numbers of retrogradely labeled motoneurons when riluzole was applied (group 4, 395 ± 16) compared with that of the untreated animals (group 3, 159 ± 21). Although the number of retrogradely labeled motoneurons appeared to be somewhat lower in group 6 (369 ± 17), when the delay of reconnection was 3 weeks, there was no significant difference in the numbers of reinnervating motoneurons between group 4 and 6 animals (1- vs. 3-week delay with riluzole treatment). In contrast, a significant decrease in the number of retrogradely labeled motoneurons was observed in animals whose cord was not treated with riluzole and suffered a 3-week delay in reconnection (group 5, 76 ± 22).
It should be noted that the number of reinnervating motoneurons was higher in group 6 animals (369 ± 17, 3-week delay in reconnection with riluzole treatment), than in animals who underwent immediate reconnection without riluzole treatment (group 1, 281 ± 23). In the spinal cords of animals with delayed nerve grafting, the morphology of the motoneurons was slightly different from those which had a chance to regenerate into freshly reconnected nerve grafts. Many of the motoneurons in the late-reconnected ventral horns had an elongated, sometimes flattened perikaryon and their dendrites appeared to be shorter, than those in intact or immediate-reconnection spinal cords.
Expression of choline acetyltransferase (ChAT) in injured and regenerating motoneurons
Next we compared the number and localization of ChAT immunoreactive surviving motoneurons with that of the FB-labeled reinnervating cells. This way we could determine how many of the surviving (ChAT-positive) motoneurons were able to regenerate (given by the number of FB-labeled motoneurons), that is to extend their axons into the peripheral nerve graft.
Strong colocalization was found in all spinal cords though there were some ChAT immunoreactive cells that were not retrogradely labeled (Fig. 4). This finding suggested that many more motoneurones survived in all cords than were able to regenerate their axons. In the riluzole-treated cords the vast majority of the surviving motoneurons were able to regenerate (86, 78, and 80% for groups 2, 4, and 6, respectively), whereas in the control cords this ratio was lower, except for group 3 animals (77, 85, and 65% for groups 1, 3, and 5).
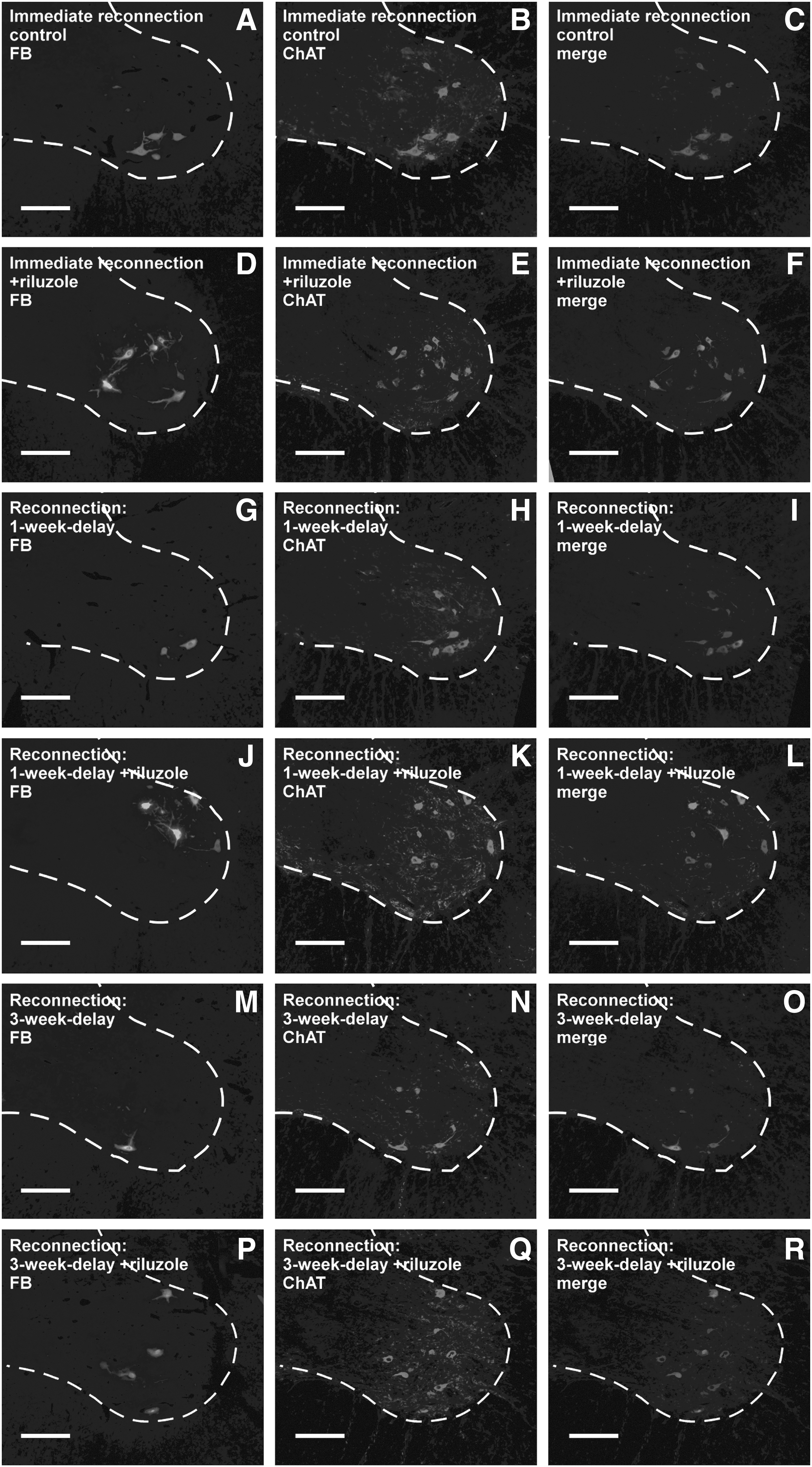
Representative images of retrogradely labeled reinnervating and ChAT-positive surviving motoneurons in the spinal ventral horns of animals in the various experimental groups. The horizontal rows of figures represent the consecutive experimental groups 1–6. Note the relatively higher numbers of FB-labeled and ChAT-positive motoneurons in the riluzole-treated groups. Scale bar: 200 μm. ChAT, choline acetyl transferase; FB, Fast Blue labeling.
Analysis of the nerve grafts
The peripheral nerve grafts used for reconnecting the spinal cord C7 motor pool with the C7 spinal nerve were qualitatively analyzed for the presence of myelinated axons that regenerated via the graft. Considerable numbers of myelinated axons were found in the grafted nerves of all experimental groups, especially in group 1 and group 2, 4, and 6 animals (Fig. 5A,B,D,F, Table 2). Control animals that received their nerve grafts with a 1- or 3-week delay displayed somewhat fewer myelinated axons in the nerve graft (Fig. 5C,E). Counts of myelinated axons in the peripheral nerve conduits (n = 3 in each group) revealed high numbers of fibers in each experimental group. The highest number of myelinated axons (2747 ± 146) was found in group 2 animals, whereas peripheral nerve grafts of group 5 animals displayed the lowest number of myelinated axons (1894 ± 119, Table 2). The g-ratio of the myelinated axons in these nerve grafts ranged between 0.63 and 0.68, without any significant difference among the groups (Table 2).

Microphotographs taken from representative semithins sections of the peripheral nerve grafts in the various experimental groups. Arrows point to myelinated fibers in E. Note the higher numbers of myelinated axons in the riluzole-treated animals. Scale bar: 25 μm.
No statistical difference was found among the g-ratio values of the various groups.
Discussion
In this study we have provided evidence that delayed reconnection of the peripheral target with its rescued motor pool by using a peripheral nerve graft after C7 ventral root avulsion induces satisfactory morphological and functional reinnervation of the denervated forelimb muscles. A fresh conduit applied 1 or 3 weeks after the avulsion injury combined with immediate riluzole treatment appears to be effective to guide considerable numbers of motor axons to their target muscles as proved by double tracer labeling.
Motoneuron survival after avulsion injury: the effect of riluzole
Several studies have shown that affected motoneurons die by the end of the second week after a ventral root avulsion injury 4,10 as the result of negative balance between the various cytotoxic and cytoprotective mechanisms. 37 A number of effective strategies have been worked out for the rescue of the motoneurons otherwise destined to die. These include ventral root reimplantation or peripheral nerve grafting into the spinal cord, 9,26,38 therapy with neurotrophic factors, 39 –43 progenitor and stem cell therapy, 44 –47 and reducing excitatory effects by blocking the presynaptic glutamate release through the use of riluzole, 11,14 Riluzole, the only clinically proven drug that prolongs the life of a patient's suffering from the late onset form of ALS, 20,22 is also neuroprotective in models of Parkinson's disease, spinal cord injury, and ventral root avulsion. 3,48,49
In our earlier study we have suggested that treatment with riluzole not only rescues the majority of injured motoneurons from cell death but maintains these cells in a condition that enables them to regenerate their axons given the right circumstances are available. 9,10 Other authors have shown the promoting effect of riluzole on dendrite outgrowth, 50 and also stimulation of neurotrophic factor (GDNF and BDNF) production in astrocytes. 51
Our earlier findings suggested that riluzole was able to restore the cellular metabolic activity of the injured motoneurons that are ready to regenerate when they are given the appropriate conduit. 9 The present study provides evidence that the vast majority of motoneurons otherwise destined to die can be rescued by riluzole, even if they did not have the opportunity to extend their axons immediately into the implanted conduit nerve and these dormant cells can regenerate provided the proper nerve conduit is established at a later time-point. Moreover, our double retrograde labeling experiments have provided evidence that the vast majority of regenerating axons reach their target muscles and induce morphological and functional reinnervation of them. The 80% overlap between the two levels (spinal nerve and forelimb muscles) of retrograde labeling results is likely to be an underestimate. It is possible that not all motor end plates were able to take up DiY, and/or FB-labeled motor neurons may have projected to other muscles not injected with DiY.
Axonal outgrowth: Root reimplantation and nerve grafts
There are two possible ways for axon regeneration across the central nervous system (CNS) border. The first way is through the remaining nerve stump when axons of regenerating motoneurons grow along the surface of the cord before reaching the nerve graft. 52 The second opportunity is that axons grow through a segment of the CNS along the guidance of Schwann cells of an implanted nerve or ventral root. 53 Reimplantation of the avulsed ventral root or implantation of a peripheral nerve graft not only provides a conduit for axonal regeneration but the denervated Schwann cells produce a number of neurotrophic factors and axon guidance molecules thus promoting axonal regeneration. 54 Earlier we have shown that the reimplanted C7 ventral root or a mixed peripheral nerve graft are equally good conduits for regenerating C7 motoneurons after an avulsion injury. 9 This contrasts with a report by Su and colleagues 55 in which the authors found the reimplanted ventral root to attract more motor axons than a saphenous nerve graft. It can be argued that the peroneal nerve used in this study is a mixed nerve, which under favorable microsurgical conditions is able to attract as many motor axons as the reimplanted ventral root.
Another interesting finding was the high number of myelinated axons found in the peripheral nerve grafts of every experimental paradigm. The number of the wrapped axons always far exceeded and showed only limited correlation with the number of retrogradely labeled motoneurons in the C7 segment in the various experimental groups (Table 2). Several explanations may exist for this phenomenon. First, it is possible that sprouting motoneuron axons persist and become myelinated with time in the nerve graft, as this nerve segment may be considered a proximal nerve stump to the distally coapted C7 spinal nerve. Second, some non-regenerating central axons may return from the nerve graft-C7 spinal nerve coaptation site, thus increasing the number of axon cross sections in the nerve. 56,57 The third opportunity is, in our view, that grafting a relatively strong mixed nerve (the common peroneal nerve is known to contain cc. 1.000 myelinated axons 58 ) may attract numerous collateral axon sprouts from the lateral funiculus pathways mildly injured during the implantation of the nerve graft. In the first two cases only the motoneurons of the C7 motor pool could be detected with retrograde tracers, whereas in the last scenario the supernumerary fibers cannot be detected by retrograde tracing within the spinal cord. It would be, however, not a surprising finding that CNS axons regenerate into peripheral nerve grafts as it has been shown by numerous studies that some injured CNS axons tend to send axon collaterals into such implants. 59
The detailed examination of the myelination (g-ratio) of the axons within the nerve grafts showed that structural design of these myelinated axons tends to be closer to that of the peripheral nerves. This is not surprising as the regenerating central axons were remyelinated by Schwann cells. On the other hand, the statistically non-significant g-ratios of the various groups suggest that riluzole treatment had no detectable effect on the axon/myelin size in these experimental paradigms.
Reimplantation is a feasible surgical procedure in the case of immediate reconstruction of anatomical pathways, or in some cases in experimental models of late reconnections. However, this procedure induces limited scarring and damage to the motor pool. 55 In the cases of human avulsion injuries typically a delayed reconnection of the spinal cord and the brachial plexus can be performed. Under these conditions only peripheral nerve grafts can be used as the avulsed root has already been retracted and cannot be used for reimplantation. In our earlier experience, fresh peripheral nerve grafts were as good conduits for regenerating spinal motor axons as were the ventral root itself, 9 although in the present model the growing axons had to pass two interfaces at both ends of the peripheral nerve graft.
Delayed reconnection provides a good conduit for riluzole-treated injured motoneurons
Considerable evidence has accumulated in the last few years proving that delayed nerve repair performed after an axotomy close to the spinal cord may attenuate the cell loss that becomes significant by 8 and 16 weeks after axotomy in the motoneuron population. Transection of the ventral branch of the C7 spinal nerve followed by a 1- and 8-week delayed nerve repair using a fresh nerve graft prevented retrograde degeneration of the spinal motoneurons. 35 Similar findings were observed in the rat brachial plexus ventral root avulsion model where nerve grafts were implanted into the spinal cord 3 weeks following injury. 60 Others have reported that a 2-week delay in reimplantation of the ventral root of the C6 spinal nerve improved the survival of motoneurons from 12% to 57% and most of the surviving motoneurons were able to regenerate their axons. 61
In our study axons of the preserved C7 motor pool were used to repopulate a reconnected peripheral nerve pathway in different experimental paradigms. It can be stated that the riluzole-treated motor pools were able to induce significant reinnervation in this model and interestingly, riluzole treatment yielded better functional and morphological reinnervation in the animals with 3-week delay in reconnection, than in control animals that had their reconnection surgeries immediately after avulsion.
It has been reported by several authors that the functional loss developed after avulsion of one or two ventral roots may be attenuated by collateral sprouting of intact nerves into the denervated muscles 62 or reorganization of the CNS circuitry. 63,64 We did not find a similar compensatory improvement in our earlier study 9 where the C7 root was avulsed but not reimplanted. This suggests that the compensatory reinnervation after C5–6 avulsion by the adjacent C7 motor pool in neonatal pups found by Korak and associates 62 or the rewiring occurring at CNS levels does not appear to part of the functional amelioration in this adult animal model. Therefore we suggest that the functional improvement found in our study is solely due to the morphological reinnervation by the surviving motoneurons resided within the C7 motor pool.
Clinical aspects
Apart from the numerous animal experiments, 7 significant human clinical findings 36 prove that functional recovery is possible after reimplantation of ventral root or reconnection with a nerve graft. Brachial plexus avulsion injury frequently remains obscured in polytraumatized patients or in patients with a severe spinal cord or head injury. Therefore a delay in diagnosis and treatment may develop in these cases and immediate direct root reimplantation is not possible. The problem lies partly in the progressive and massive reduction of injured motoneurons, a process that likely occurs in humans, too. 2 On the other hand, localization and mobilization of the retracted ventral roots is often impossible. Therefore, connection of injured motoneurons to the lesioned brachial plexus by nerve grafts has been introduced both experimentally and clinically in order to bridge the gap as an alternative treatment. 8
The question is raised about what is the best timing for surgery after plexus injury. 65,66 Our findings indicate that in the case of immediately started riluzole therapy, delayed reconnection with peripheral nerve grafts in patients after brachial plexus injury appears to be a feasible treatment. The time shift between the trauma and the surgery is likely to be further extended: a satisfactory number of motoneurons may survive and remain capable of reinnervating their targets and the peripheral nerve graft serves as an appropriate conduit for the reinnervating fibers. It can therefore be argued that the combination of riluzole treatment followed by a delayed nerve repair is a promising new treatment to restore function after avulsion of the brachial plexus, even after relatively long delays between the time of the injury and surgical intervention.
Footnotes
Acknowledgments
The research was partly supported by the European Union and the State of Hungary, co-financed by the European Social Fund in the framework of TÁMOP-4.2.4.A/2-11/1-2012-0001 National Excellence Program. László Gál was supported by the Innovative Economy–National Cohesion Strategy, Foundation for Polish Science and European Union–European Regional Development Fund. The authors are indebted to Mrs. I. Kovács for her technical assistance. We are grateful to Tocris Cookson for kindly providing us with riluzole.
Author Disclosure Statement
No competing financial interests exist.