
Abstract
Spinal cord injury (SCI) results in marked atrophy and dysfunction of skeletal muscle. There are currently no effective treatments for SCI-induced muscle atrophy or the dysfunction of the remaining muscle tissue. Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase-4 (Nox4) produces reactive oxygen species (ROS) in sarcoplasmic reticulum (SR) and has been identified as an important O2 sensor in skeletal muscle. Ryanodine receptors (RyRs) are calcium (Ca2+) channels that are responsible for Ca2+ release from SR. In skeletal muscle, type1 RyR (RyR1) is predominantly functional. RyR1 is regulated by multiple proteins, including calstabin1, which assures that they close appropriately once contraction has ceased. RyR1 function is also regulated by oxidation and redox-dependent cysteine nitrosylation. Excessive oxidation/nitrosylation of RyR1 is associated with dissociation of calstabin1 and reduced muscle force generation. However, whether Nox4 levels in skeletal muscle are elevated or whether RyR1 is oxidized or nitrosylated after SCI has not been determined. In this study, we examined Nox4 expression, oxidation/nitrolysation status, and association of calstabin1 with RyR1 in skeletal muscle derived from rats that were subjected to T4 complete transection (SCI), and observed elevated expression of Nox4 messenger RNA and protein in muscle after SCI associated with enhanced binding of Nox4 to RyR1, increased oxidation and nitrosylation of RyR1, and dissociation of calstabin1 from RyR1 in SCI rat muscle. Our data suggest that RyR1 dysfunction resulting from excessive oxidation/nitrosylation may contribute to reduced specific force after SCI and suggest that Nox4 may be the source of ROS responsible for increased oxidation and nitrosylation of RyR1.
Introduction
S
One plausible mechanism for reduced specific force production after SCI is ryanodine receptor (RyR) dysfunction. RyRs are calcium-activated Ca2+ channels that are responsible for release of Ca2+ from the SR. 4,5 In skeletal muscle, the rise in cytosolic Ca2+ necessary for EC coupling occurs predominantly through type 1 RyR (RyR1). 4 The RyR1 is a homotetrameric protein complex to which other proteins bind, including protein phosphatase 1 (PP1), phosphodiesterase (PDE4D3), calmodulin, and the RyR1 channel-stabilizing protein calstabin1 (also called FK506 binding protein 12, [Fkbp12]). 9 –11 Binding of calstabin1 to RyR1 stabilizes the channel and prevents Ca2+ leaks through the closed RyR1. 9,11 In several physiological and pathological conditions such as age-related sarcopenia and cancer-induced cachexia, intracellular Ca2+ leakage resulting from dissociation of calstabin1 from RyR1 has been observed. 6 –8 In these conditions, administration of small molecules that promote binding of calstabin1 to RyR1 reduces the probability of RyR1 channels leaking and improves specific force. 6 –8
RyR1 is an established redox-sensitive channel. Alterations in RyR1 redox state can result in either activation 12 or inactivation 13,14 of the receptor. Redox status affects skeletal muscle RyR1 function and Ca2+ signaling. 15 –17 It has been demonstrated that skeletal muscle acutely couples changes in pO2 to altered Ca2+ release through the RyR1. 14 At relatively low concentrations of pO2, endogenously generated nitric oxide (NO) regulates RyR1 activity by S-nitrosylation of a single Cys thiol. 18,19 With increasing levels of pO2, RyR1 activity is enhanced in association with the oxidation of additional Cys thiols. 14,18 RyR1 contains multiple cysteine residues 16 that can be modified by S-nitrosylation. 20 –22 Exogenous S-nitrosylation of RyR1 has been shown to reduce the affinity of calstabin1 binding to purified SR vesicles. 20 It also has been reported that protein S-nitrosylation can be triggered by free radical nitric oxide derivatives such as peroxynitrite. 21,22 Moreover, many pathological conditions are associated with increased levels of reactive oxygen species (ROS) in skeletal muscle, as well as with altered cellular Ca2+ handling. 23 The source of ROS in these conditions may include mitochondrial respiration and nicotinamide adenine dinucleotide phosphate (NADPH) oxidases (Nox). Nox is a family of specialized enzymes with their primary function to generate ROS. 24,25 In addition, it has been reported that ROS production in the SR is primarily catalyzed by Nox4. 14 Among the seven NADPH oxidases, Nox4 is unique in that it is present in the SR and is constitutively active. 14,24,25 In addition, Nox4 has been reported to catalyze ROS production in the SR. 14 In the heart, oxidation of RyR by Nox4 reduces the binding of calstabin, resulting in impaired Ca2+ handling and contractile dysfunction. 12,26 In the present study, the effects of SCI on Nox4 expression, oxidation/nitrosylation status of RyR1, and association of calstabin1 with RyR1 was examined in rat skeletal muscle below the level of injury.
Methods
Animal studies
Procedures with experimental animals were approved by the Institutional Animal Care and Use Committee at the James J. Peters VA Medical Center and were conducted in according with accepted standards of humane animal care. Male Wistar rats (8–10 weeks of age) weighing approximately 250 g were obtained from Taconic Farms (Hudson, NY). The animals were anesthetized and a complete transection of the spinal cord at T4 level was performed. Some animals underwent a sham SCI in which the spinal cord was exposed but not transected. At Day 56 after surgery, the animals were euthanized. The gastrocnemius muscles were carefully removed, weighted, and immediately frozen in liquid nitrogen. The effects of SCI on body and muscle weights for this cohort of animals have been reported elsewhere. 27
Reagents
Primers and probes used for real-time polymerase chain reaction (PCR) were obtained from Applied Biosystems (Foster City, CA). Antibodies against Nox4 were a product of Santa Cruz Inc. (Santa Cruz, CA). Anti-RyR1 antibody was purchased from Thermo-Fisher (Rockford, IL). Anti- 2,4-dinitrophenylhydrazine (DNPH) and calstabin1 antibodies were obtained from AbCam (Cambridge, MA). Antibody against Cysteine-SNO was purchased from LifeSpan BioSci. (Seattle, WA).
Reverse transcriptase quantitative real-time PCR (RT-qPCR)
Real time PCR was performed using Assay on Demand probes and primers and TaqMan 2 × PCR mix (Applied Biosystems). Each measurement was performed in triplicate. The mean of crossing points for the three technical replicates was used in subsequent calculations. Expression levels of target messenger RNAs (mRNAs) were expressed relative to values for the sham-SCI group using the 2−ΔΔCt method 28 ; 18s RNA was used as the internal control.
Preparation of tissue lysates, Western blotting, and immunoprecipitation (IP)
Gastrocnemius muscle (25 mg) was homogenized in 500 μL of lysate buffer (150 mM sodium chloride, 3.2 mM Na2PO4, 0.8 mM K2PO4 [pH 7.4], 1% NP-40, 0.5% sodium deoxycholate, 0.5% sodium dodecylsulfate) using a Polytron. Homogenates were cleared by centrifugation in a microcentrifuge at 14,000 rpm for 5 min. Proteins from the cytosolic and nuclear fractions were isolated using a commercial kit according to the manufacturer's instructions (Thermo-Fisher; Rockford, IL). For Western blot analysis, glyceraldehyde 3-phosphate dehydrogenase, and proliferating cell nuclear antigen were used as internal controls for the analyses of total tissue lysates and nuclear protein, respectively. IP was performed using a kit from Thermo-Fisher.
Statistical analysis
Data are expressed as mean values ± standard error of the mean. The significance of differences between sham and SCI groups was determined using an unpaired, two-tailed Student's t-test. Statistical calculations were performed using Prism 4.0 (GraphPad Software, San Diego, California). A p < 0.05 was considered statistically significant.
Results
Upregulation of Nox4 expression after SCI
The mRNA levels of Nox4 and Nox2 in rat gastrocnemius muscle were determined in SCI and Sham groups using RT-qPCR analysis. While Nox2 mRNA levels were unchanged after SCI, Nox4 mRNA was significantly upregulated by approximately 2-fold in the tissue from SCI rats, compared with that from Sham rats (Fig. 1A). Nox4 protein also was significantly upregulated in total tissue lysates after SCI. To gain initial insights as to the localization of Nox4, immunoblots were repeated after subcellular fractionation and the fraction enriched in nuclei was subjected to Western blotting. A more than 15-fold increase in Nox4 protein levels were observed in the nuclear fraction from SCI rats (Fig.1B, 1C).
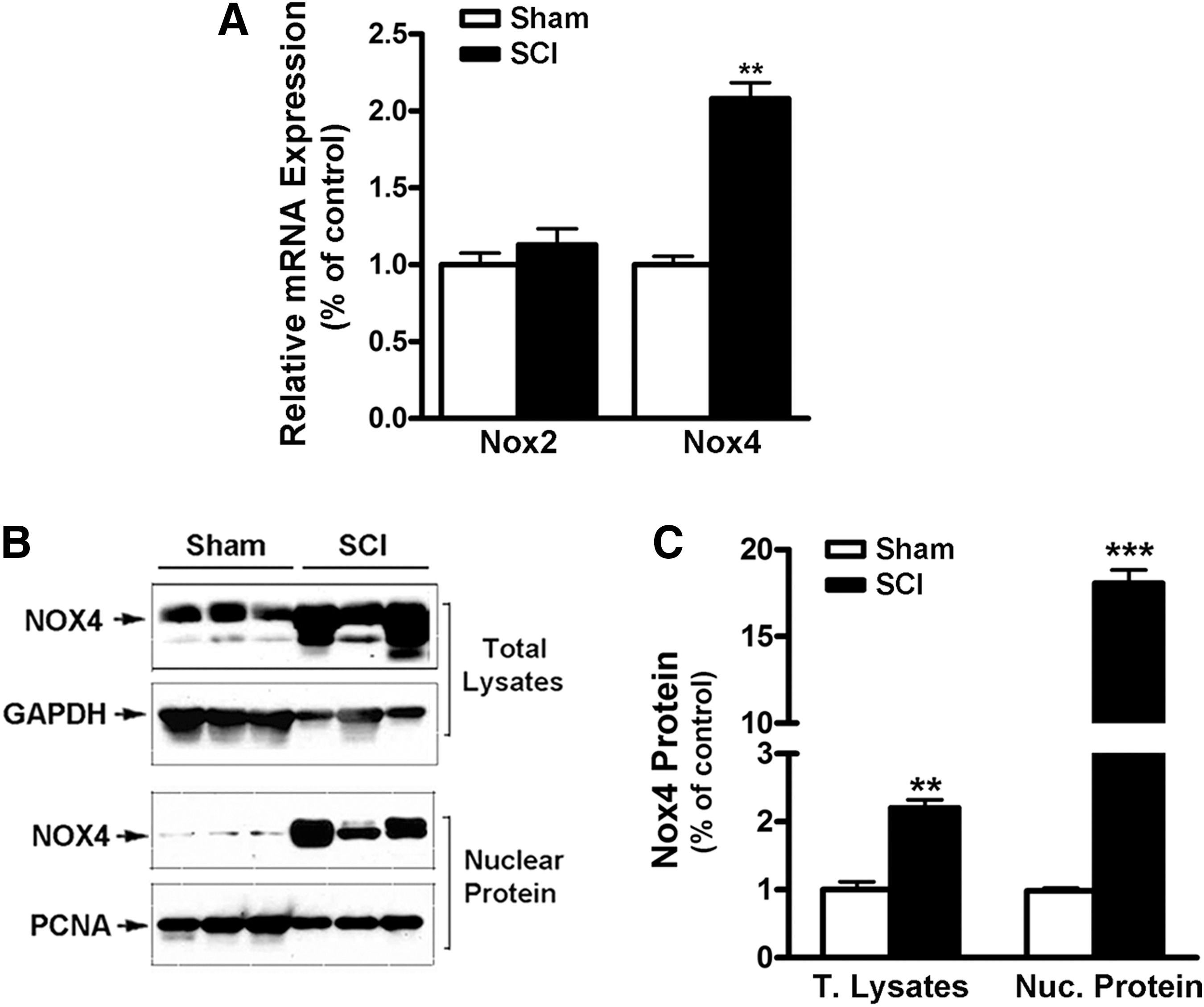
Upregulation of Nox4 messenger RNA (mRNA) and protein expression in muscle after spinal cord injury (SCI).
Increased binding of Nox4 to RyR1 after SCI
Nox4 has been reported to directly bind to RyR1. 8 To determine whether SCI promoted binding of Nox4 to RyR1 in gastrocnemius muscle, immunoprecipitation assays were performed using an antibody against RyR1 followed by Western blotting using anti-Nox4 antibody. We observed similar levels of RyR1 protein in the muscle from sham control and SCI rats. By contrast, the amount of Nox4 that was co-immunoprecipitated with RyR1 was increased 2-fold in tissues from SCI rats than that in Sham rats (Fig. 2A, 2B).
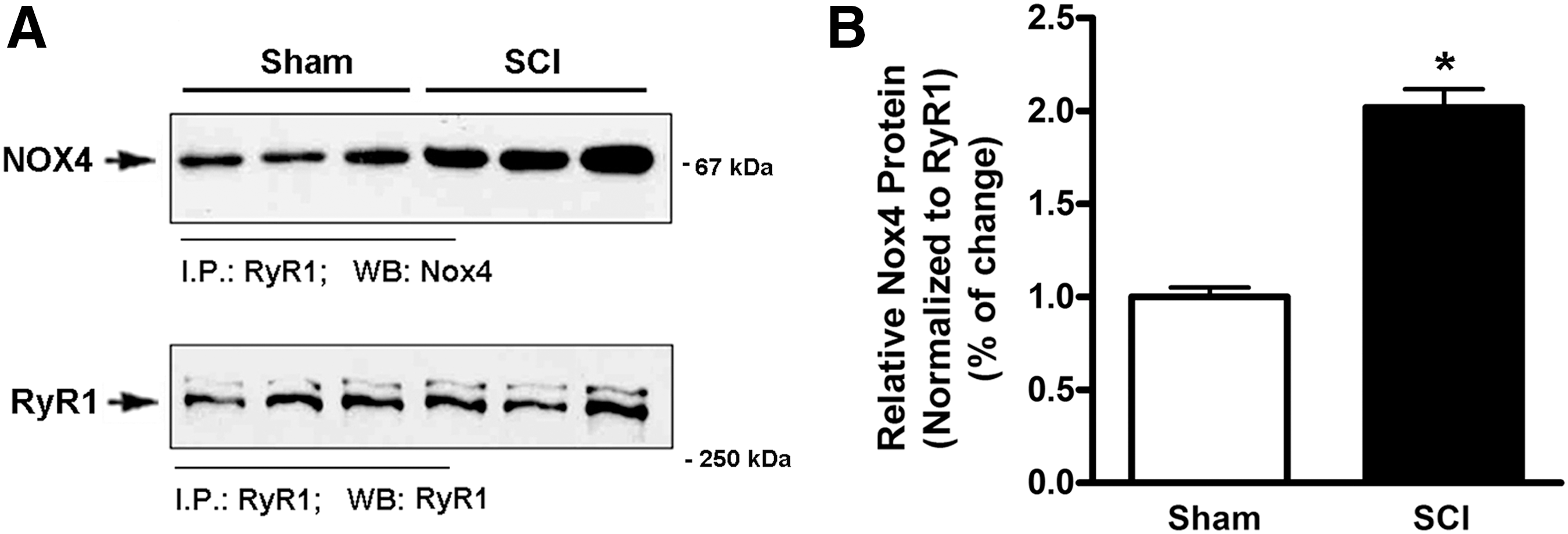
Increased levels of Nox4 facilitate the binding of Nox4 to type1 ryanodine receptor (RyR1).
Hyperoxidation and hypernitrosylation of RyR1 after SCI
The effects of SCI on oxidation and nitrosylation of RyR1 in gastrocnemius muscle were next determined. To detect RyR1 oxidation, the immunoprecipitated products of RyR1 were derivatized to DNPH and the signal associated with RyR1 was assayed using an anti-DNPH antibody. S-nitrosylation status of RyR1 was measured by Western blotting of immunoprecipitated RyR1 using an anti-Cysteine-SNO antibody. Oxidation and nitrosylation of the RyR1 were greatly increased in muscle from SCI rats, compared with that of sham rats (Fig. 3). To obtain further evidence regarding oxidative stress, lipid peroxidation was evaluated in muscle homogenates from SCI rats and compared with that for sham animals. This analysis revealed a significant elevation of lipid peroxidation in muscles from SCI animals (Supplementary Fig. 1; see online supplementary material at

Spinal cord injury (SCI) causes hyperoxidation and hypernitrosylation of the type1 ryanodine receptor (RyR1) in skeletal muscle.
Dissociation of calstabin1 from RyR1 after SCI
The possibility that hyperoxidation and hypernitrosylation of the RyR1 after SCI influenced the binding of calstabin1 to RyR1 was tested. The expression of calstabin1 mRNA and protein were not altered in gastrocnemius muscle after SCI (Fig. 4). Immunoprecipitation using antibodies against RyR1 revealed that calstabin1 was co-immunoprepitated from gastrocnemius muscle from sham rats but had almost totally dissociated from RyR1 in SCI rats.
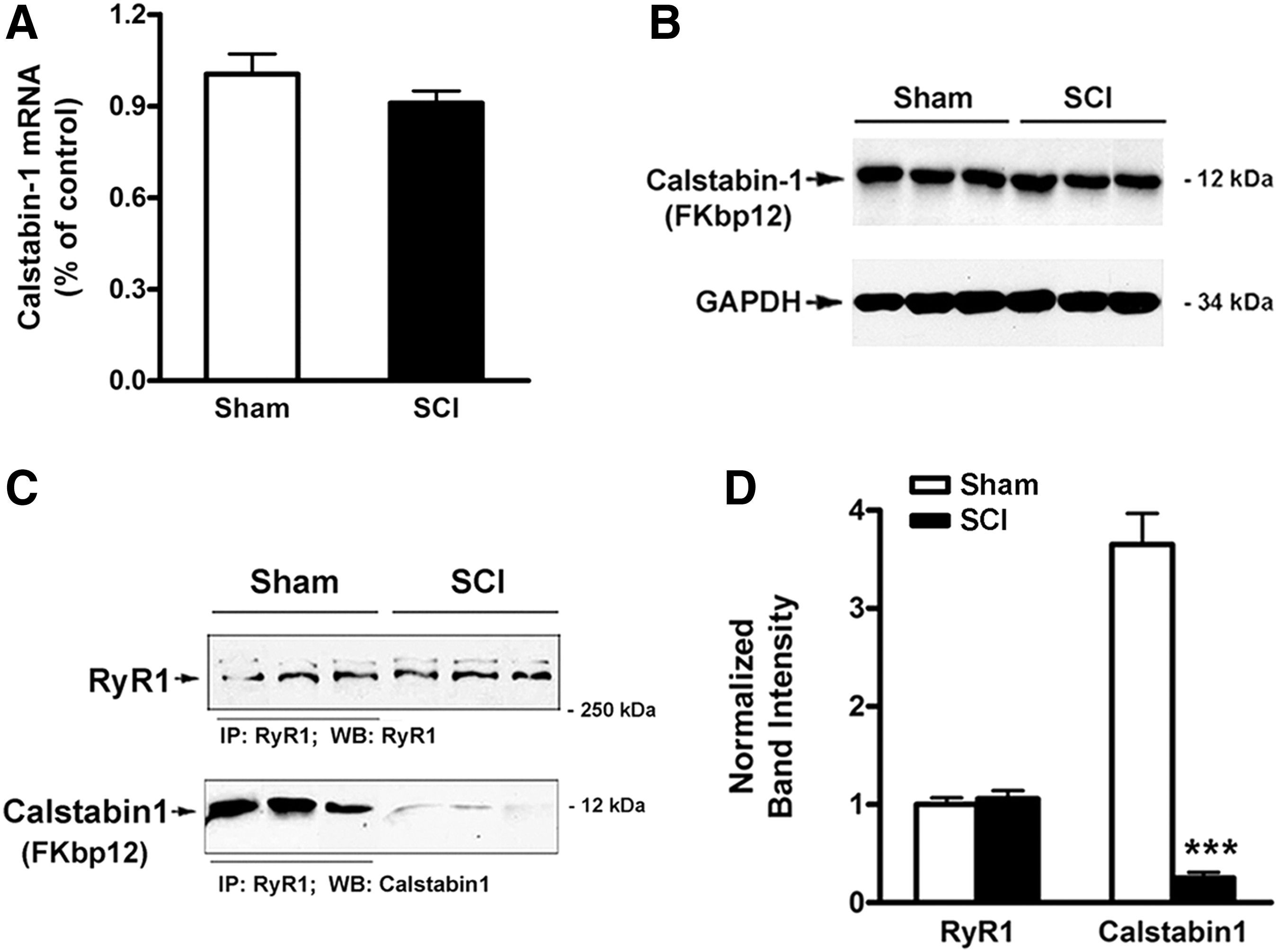
Dissociation of calstabin1 from type1 ryanodine receptor (RyR1) after spinal cord injury (SCI).
Discussion
Our findings in a rat model of motor-complete SCI support several conclusions. An elevation of Nox4 mRNA and protein expression associated with increased binding of Nox4 to RyR1 occurs in paralyzed muscles. Our findings provide biochemical evidence of RyR1 hyperoxidation and hypernitrosylation at 56 days after SCI and demonstrate that these modifications are associated with virtually complete dissociation of calstabin1 from RyR1. As noted above, dissociation of calstabin1 from RyR1 has been linked to an increased open probability of RyR1 and a drug that normalized calstabin1 binding to oxidized, nitrosylated RyR1 increased specific force of contraction. 4,6 –8 Taken together, these findings support the view that RyR1 dysfunction, as a consequence of excessive oxidation and nitrosylation and resultant calstabin1 dissociation from RyR1, may be a significant cause of skeletal muscle weakness after SCI. Studies regarding the source of ROS in skeletal muscle after SCI have not been performed. The physical association of Nox4 with RyR1, elevation of Nox4 binding to RyR1 after SCI, and constitutive activity of Nox4 suggest a role for Nox4 in oxidation of RyR1 in skeletal muscle after SCI. The identification of the source of ROS responsible for oxidation and nitrosylation of RyR1 and functional consequences oxidized and nitrosylated RyR1 channels to bind calstabin1 are important directions for future studies.
How Nox4 expression is elevated after SCI remains unclear. Expression of Nox4 is upregulated by angiotensin II, 29 leptin, 30 nitric oxide synthase 1, 31 and transforming growth factor β (TGFβ). 8,32 In a study of the pathogenesis of weakness due to breast cancer metastasis to bone, release from bone matrix of TGFβ was found to be responsible for upregulation of Nox4, increased phosphorylation of RyR1, dissociation of calstabin1 from RyR1, and weakness. 8 Whether TGFβ, leptin, or angiotensin II contributes to the elevated Nox4 expression in muscle paralyzed by SCI is a subject of obvious future investigation.
Small molecules have been identified that promote RyR1-calstabin1-binding interactions in animal models of muscular dystrophy and aging. 6,7 These drugs promote the binding of calstabin1 to oxidized and/or nitrosylated RyR1 leading to stabilization of the receptor, and prevent Ca2+ leaks from the channel. As consequence, muscle-specific force production is increased by such agents by up to 50%. 6 The above findings support further studies regarding the possible functional benefits of these drugs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.