
Abstract
The effects of multiple injection of bone marrow mononuclear cells (BMNCs) on spinal cord injury (SCI) were compared with those of single injection in rats. BMNCs separated by density-gradient centrifugation from a bone marrow perfusate were injected three times (once weekly) through the cerebrospinal fluid (CSF) via the fourth ventricle, and the locomotor improvement and tissue recovery, including axonal regeneration, were compared with those of single injection. While the single-injection group showed a steep elevation of the Basso-Beattie-Bresnahan (BBB) score 1 week after transplantation, the multiple-injection group maintained a similar steep elevation for 2 weeks after transplantation, and the BBB scores of the multiple-injection group remained thereafter at a level approximately 2–3 points higher than those of the single-injection group until the end of the experiment. There were significant differences between the single- and multiple-injection groups at 3, 4, and 8 weeks after transplantation. The difference in BBB scores at 8 weeks after transplantation suggested that there was a marked difference in the quality of locomotor behaviors between the single-and multiple-injection groups at this stage. An extensive outgrowth of regenerating axons through the astrocyte-devoid areas and a marked reduction of cavity formation were found in both the single- and multiple-injection groups. There were, however, no significant differences in the density of regenerating axons or volumes of cavities between the single- and multiple-injection groups. These results showed that although tissue recoveries were similar between single and multiple injection, the multiple injection of BMNCs was more beneficial for locomotor improvement than single injection for the treatment of SCI. Considering the technically simple and low-cost procedures for the preparation and injection of BMNCs, multiple injection of BMNCs by lumbar puncture has an advantage over single injection on clinical application.
Introduction
V
Subsequently, we studied the transplantation of BMSCs into the spinal cord lesion in rats with sub-acute SCI, demonstrating that axonal regeneration was enhanced at the spinal cord lesion, and locomotor behaviors were improved as assessed by BBB scores. However, transplanted BMSCs were no longer found in the host spinal cord tissue at 2 weeks post-transplantation. 12 This study showed that BMSCs did not serve as scaffolds for regenerating axons, but acted as the source of some neurotrophic factors effective for the axonal regeneration and locomotor improvements. The subsequent study examined the efficacy of multiple injection (three times) of BMSCs into the CSF in sub-acute and chronic SCI. This study showed that locomotor behaviors were improved and that tissue repair, including axonal regeneration, was promoted even in chronic SCI. 13,14
On the other hand, we examined the effect of transplanting bone marrow mononuclear cells (BMNCs) through the CSF of rats with SCI. 15 BMNCs were separated by density-gradient centrifugation from a bone marrow perfusate and transplanted without cell culture through the CSF via the fourth ventricle. This study demonstrated that the infusion of BMNCs through the CFS improved locomotor functions and enhanced tissue repair, as in the case of BMSC transplantation. Samdani and colleagues reported that the transplantation of human BMNCs to the injured spinal cord of rats was similar in its efficacy to that of BMSC transplantation. 16 Based on these experimental studies, together with those of BMSC transplantation, the BMNC transplantation was advanced to clinical application: BMNCs were transplanted by lumbar puncture into 10 patients with SCI in 2013. 17 Varying degrees of locomotor and somatosensory recoveries were achieved, while there was no adverse effect of the clinical BMNC transplantation on patients with SCI.
It should be noted that BMNC transplantation has a marked advantage over other cell therapies (i.e., BMNCs do not require cell culture before transplantation). They can be transplanted to patients in an operation room immediately after separation from a bone marrow perfusate. This means that the cost of BMNC transplantation is much lower than that of other cells that need a cell-processing facility for cell preparation.
During the study of the clinical application of BMNCs, it was hypothesized that multiple injection might be more effective than single injection. There has been no experimental study comparing the effects of multiple injection with those of single injection. Therefore, we studied the effects, including locomotor improvements and tissue recoveries, of the multiple injection (three times) of BMNCs, compared with those of single injection in rats with SCI.
BMNCs were transplanted into the CSF via the fourth ventricle, as described in our previous studies. Locomotor behaviors were evaluated using the BBB scale every week for 8 weeks after transplantation. The tissue repair, including axonal regeneration, was evaluated at the end of the experiment (8 weeks after transplantation).
Methods
Surgery
Forty Sprague-Dawley rats (6 weeks old, female) supplied by SLC Inc., Shizuoka, Japan, were used in the present study. This study was performed with the approval of the Animal Experiment Committee of Aino University, and all animal experiments were performed in accordance with the Guidelines for Proper Conduct of Animal Experiments and Related Activities formulated by the Ministry of Education, Culture, Sports, Science and Technology. The methods of spinal cord injury were basically the same as those described in the previous paper. 12 Briefly, rats were anesthetized by the inhalation of isoflurane (1–2% Escain; Mylan, Osaka, Japan) administered at a flow rate of 2 L/min. Laminectomy was performed at the Th 8–9 vertebrae to expose the spinal cord. The spinal cord with the dura mater was exposed, and injured by letting a metal weight (10 g) drop from a height of 7.5 cm using an NYU impactor. Bleeding was seen under the dura mater at the site of the crushed spinal cord. The layered muscles and skin were sutured to close the lesion. The animals were given 3 mL of saline subcutaneously immediately after surgery to avoid dehydration, and an antibiotic (gentacin at 3 mg/kg, Schering Plough, Osaka, Japan) daily for 3 days to prevent infection. In addition, rats were given lepetan (0.02 mL/kg, Otsuka, Tokyo, Japan) on the first day of surgery. Small amounts of food were placed on the floor of the cage to enable the rats to access the food easily on the first post-operative day. Later, food and water were provided as usual from the top of the cage. Rats were housed in an atmosphere of 55% humidity at a temperature of 23°C.
The experimental design is shown in Figure 1. BMNC or phosphate-buffered saline (PBS) injection started one week after SCI. BBB scores were examined every week after SCI, and all animals were fixed 8 weeks after the first injection of BMNCs or PBS.
The three experimental groups are shown: (1) one-time transplantation of bone marrow mononuclear cells (BMNCs; single-transplantation group; n = 14), in which BMNCs were transplanted at 1 week after spinal cord injury (SCI) followed by the injection of phosphate-buffered saline (PBS) for 2 weeks; (2) three-time transplantation of BMNCs (multiple-injection group; n = 12), in which BMNCs were transplanted at 1, 2, and 3 weeks after SCI; and (3) the injection of PBS (control group; n = 6), in which PBS, instead of BMNCs, was injected every week for 3 weeks after SCI. The Basso-Beattie-Bresnahan scores were examined every week after the SCI. All rats were fixed 8 weeks after the initial transplantation.
Cell preparation and transplantation
A bone marrow perfusate was obtained from the femurs and tibias of rats, and BMNCs were separated by density-gradient centrifugation using Lymphoprep (Alere Technologies AS, Oslo, Norway). The upper layer of the bone marrow on separation was collected and centrifuged. The precipitated BMNCs were resuspended in PBS at a density of 5 × 106 cells/70 μL. The method of BMNC separation was described in detail in our previous study. 15
BMNCs were transplanted through the CSF via the fourth ventricle as in our previous study. 9 The technique for injection of transplants via the fourth ventricle is reliable. Transplants injected into the CSF via the ventricle flow through the subarachnoid space to the spinal cord. We think that transplants injected via the ventricle have the same effects on the spinal cord as those injected via lumbar puncture in rats.
For cell transplantation, a hole of 1 mm in diameter was drilled in the skull at a site 3.5 mm caudal to the lambda suture on the median line in rats 1 week before spinal cord injury. One week after spinal cord crush-injury, a cell suspension of 30 μL containing 5 × 105 BMNCs was injected using an insulin syringe though the hole into the fourth ventricle. We used the stereological coordinate model SR-6N (Narishige Scientific Instrument Lab, Tokyo, Japan) for cell injection into the fourth ventricle. In a preliminary experiment, we injected Evans blue solution into the fourth ventricle using the same setting of the stereological coordinate, and confirmed that Evans blue solution could be successfully infused into the CSF of the fourth ventricle. The injection time was 5 min, and the syringe needle was kept at the site for 5 min after injection. Rats were separated into three groups depending on the pattern of BMNC transplantation: 1) the multiple-injection group (n = 14) was injected with BMNCs once a week for 3 weeks; 2) the single-injection group (n = 12) was injected with BMNCs once in the first week of the experiment, followed by PBS injection for the 2 subsequent weeks; and 3) the PBS-injection group (n = 6) was injected with the PBS, instead of BMNCs, for 3 weeks in the same manner as in the other two groups (Fig. 1).
Locomotor assessment
The locomotor behaviors were assessed before cell transplantation (i.e., 1 week after spinal cord injury), and every week until 8 weeks after transplantation based on the BBB (Basso, Beattie, and Bresnahan) scale. 18 Rats were observed walking in an open area (ca. 1.0 m2) for 5 min. Locomotion was recorded by video camera, and analyzed by two researchers who were not aware of the experiment. Rats showing BBB scores below 1 point were used for transplantation (0 weeks).
Fixation of rats
All rats were fixed at 8 weeks after transplantation. Rats were initially anesthetized by the inhalation of isoflurane, followed by the injection of 0.1 mL of Somnopentyl (pentobarbital sodium, Sankyo, Tokyo, Japan). For fixation, PBS (50 mL) was first perfused through the heart, followed by 100 mL of the fixative containing 4% paraformaldehyde in 0.1 M PBS (pH 7.5). The injured spinal cord was exposed, excised from the spinal bone, and stored for 1–2 days in the same fixative before proceeding to embedding in O.C.T. compound.
Immunohistochemistry
The spinal cord stored in the fixative as above was immersed for 1–2 days in PBS containing 30% sucrose, and embedded in OCT compound. Specimens embedded in the OCT compound were cut horizontally at 10 μm thick in a cryostat and attached to glass slides. The sections were washed with PBS, immuno-blocked with 3% bovine serum albumin (BSA) in PBS for 30 min at room temperature, and permeated with 0.3% Triton X-100 in 1% BSA-PBS for 30 min prior to incubation with primary antibodies. The sections were then incubated overnight with a solution containing the primary antibodies as follows: mouse anti-β-III-tubulin monoclonal antibody (1:1,000; Covance, NJ) for axons, rabbit anti-S100 monoclonal antibody (1:200; Abcam) for Schwann cells, rabbit-anti collagen type-1 polyclonal antibody (1:200; Abcam) for collagen fibrils, mouse anti-glial fibrillary acidic protein (GFAP) monoclonal antibody (1:500; Sigma–Aldrich, St. Louis, MO), or rabbit anti-GFAP polyclonal antibodies (1:2,000; Novus Biologicals, Littleton, CO) for astrocytes. These primary antibodies were used for single-staining, or the mixtures of any two of these antibodies were used for double-staining. After being washed with PBS, the sections were incubated overnight with CF488A-labeled goat anti-mouse IgG (1:200; Biotium, Hayward, CA), CF488A-labeled anti-rabbit IgG (1:200; Biotium, Hayward, CA), CF555-labeled goat anti-mouse IgG (1:200; Biotium, Hayward, CA), or Alexa Fluor 555 goat anti-rabbit IgG (1:200; Thermo Fisher Scientific, Waltham, MA) antibodies.
Cavity volume
The cavity volume was measured and calculated as described previously. 14 In brief, five, 10, and 12 rats were used for assessment of the cavity volume in the PBS-, single-, and multiple-injection groups, respectively, at 8 weeks after transplantation. Horizontal cryostat sections of the spinal cord were stained by hematoxylin and eosin (HE) on the slide glass. After staining, the sections were embedded in Canada balsam (Wako, Osaka) and used for measurement of the cavity volume. The border of cavities was traced to measure their areas using the Image Filing System (Flovel, Tokyo, Japan) with a microscope (Olympus, CKX41, Tokyo, Japan). As cavities were measured in HE-stained sections, astrocyte-devoid areas were not included in the “cavity volume” of the present study. The areas of the cavities were measured in every third section. Since the sections were 10-μm thick, the total volume of the cavities was calculated by multiplying the average area of the cavities by the depth of the sections examined. The relative volume of the cavities was obtained by dividing the values of the total cavity volume by those of the whole spinal cord volume at the corresponding level.
Axon density
The density of axons extending through the astrocyte-devoid areas was compared among the three groups. Five, 10, and 12 rats were used for assessment of the axon density in the PBS-, single-, and multiple-injection groups, respectively. Three sections at different levels were selected from each spinal cord, and three appropriate fields were randomly selected for measurement of axon density within the astrocyte-devoid area in each section. The total number of pixels for β-tubulin-immunostaining was measured in each field using Image-J software version 1.48v. Pixels of β-tubulin-immunopositive field were represented as a percentage of the total pixels of the astrocyte-devoid areas including cavities in the spinal cord lesion.
Statistical analysis
BBB scores were analyzed by two-way analysis of variance, and post hoc analysis was performed by Tukey's test to assess the significance of differences at each time-point (p < 0.05). Relative volumes of cavities and axon densities were evaluated by Student's t-test.
Results
Axonal regeneration
The spinal cord became narrow at the lesion that extended 3–4 mm rostro-caudally. The structure of the white matter of the spinal cord was disorganized in varying patterns, and the gray matter of the spinal cord was almost completely absent at the lesion. The double-immunostaining at a low magnification stained overwhelmingly for astrocytes (green) in the white matter, and for axons (red) in the gray matter of the spinal cord (Fig. 2 and Fig. 3). However, the higher magnification showed that axons extended in association with astrocytes in the white matter region at the spinal cord lesion (Fig. 3B, 3C).
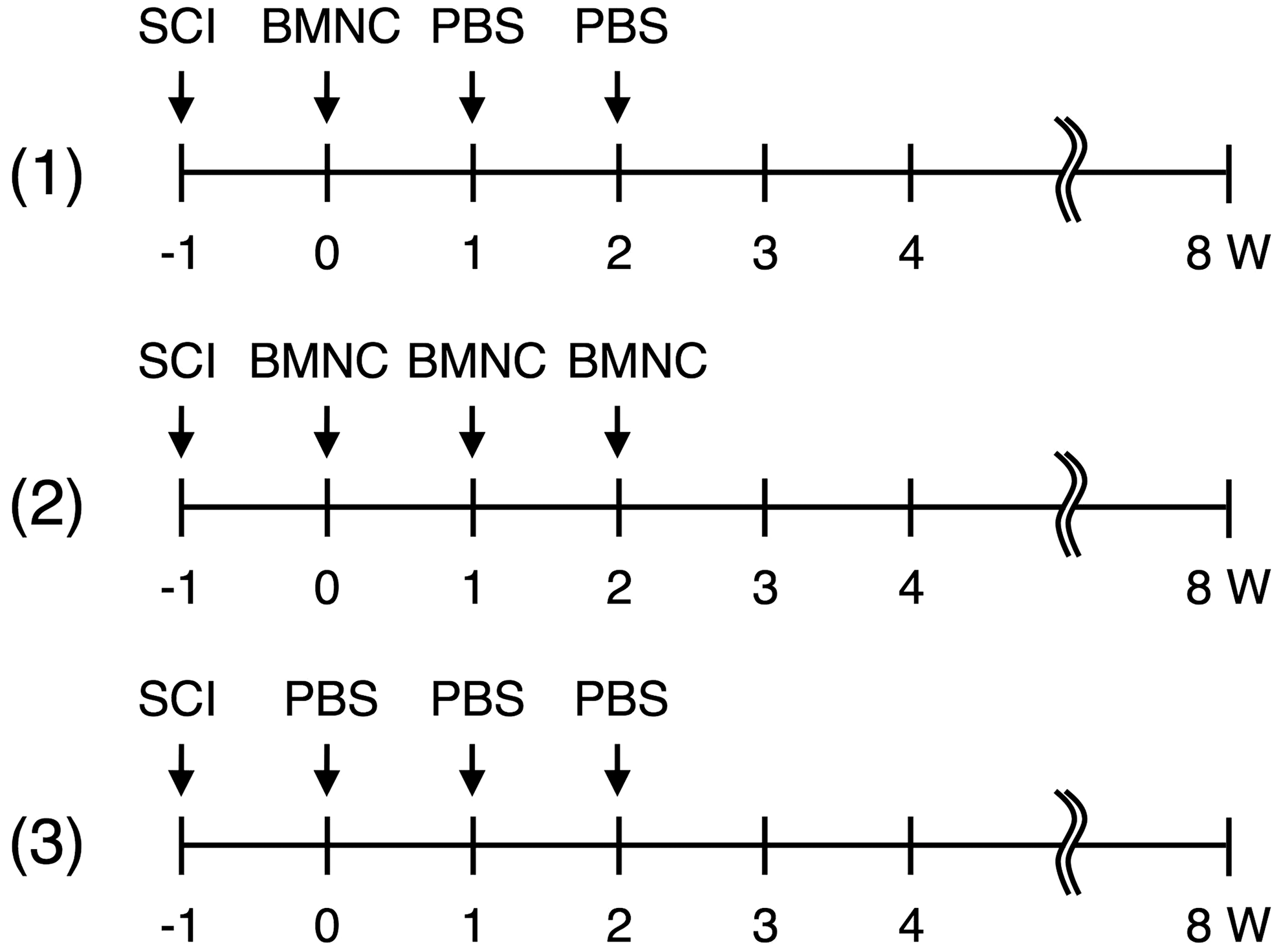
Axonal outgrowth through astrocyte-devoid areas of the single-injection group. The picture shows a representative section of the spinal cord of the single-injection group.
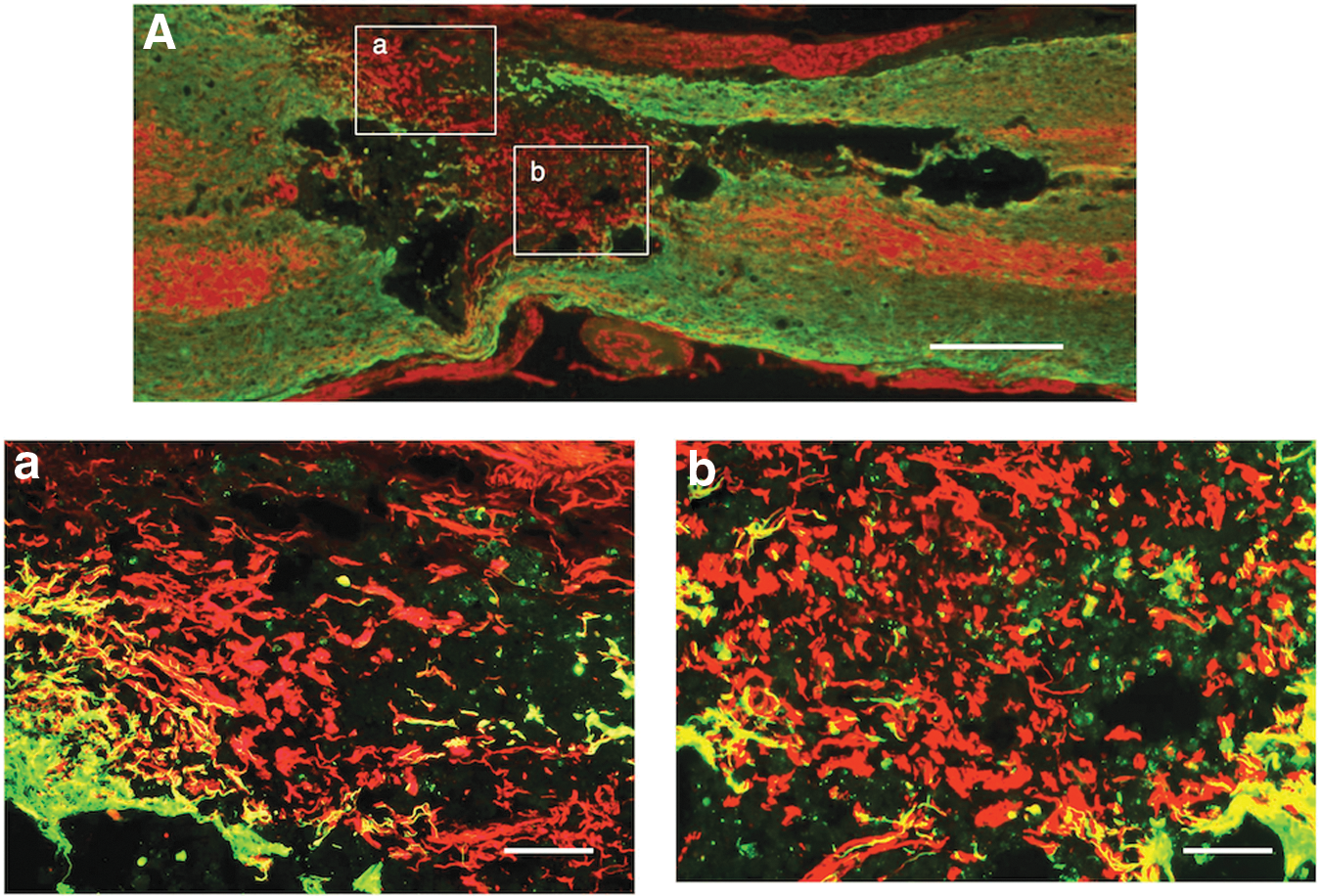
Axonal outgrowth through astrocyte-devoid areas of the multiple-injection group. The picture shows a representative section of the spinal cord of the multiple-injection group.
Immunohistochemistry for astrocytes showed that there were “empty regions” bordered by green-immunostained astrocytes (Fig. 2 –4). Such empty regions with no immunostaining for astrocytes were called “astrocyte-devoid areas” in the previous and present studies. Some astrocyte-devoid areas were actually empty cavities (Fig. 4), but others were not vacant: they contained numerous axons (Fig. 2 and Fig. 3).
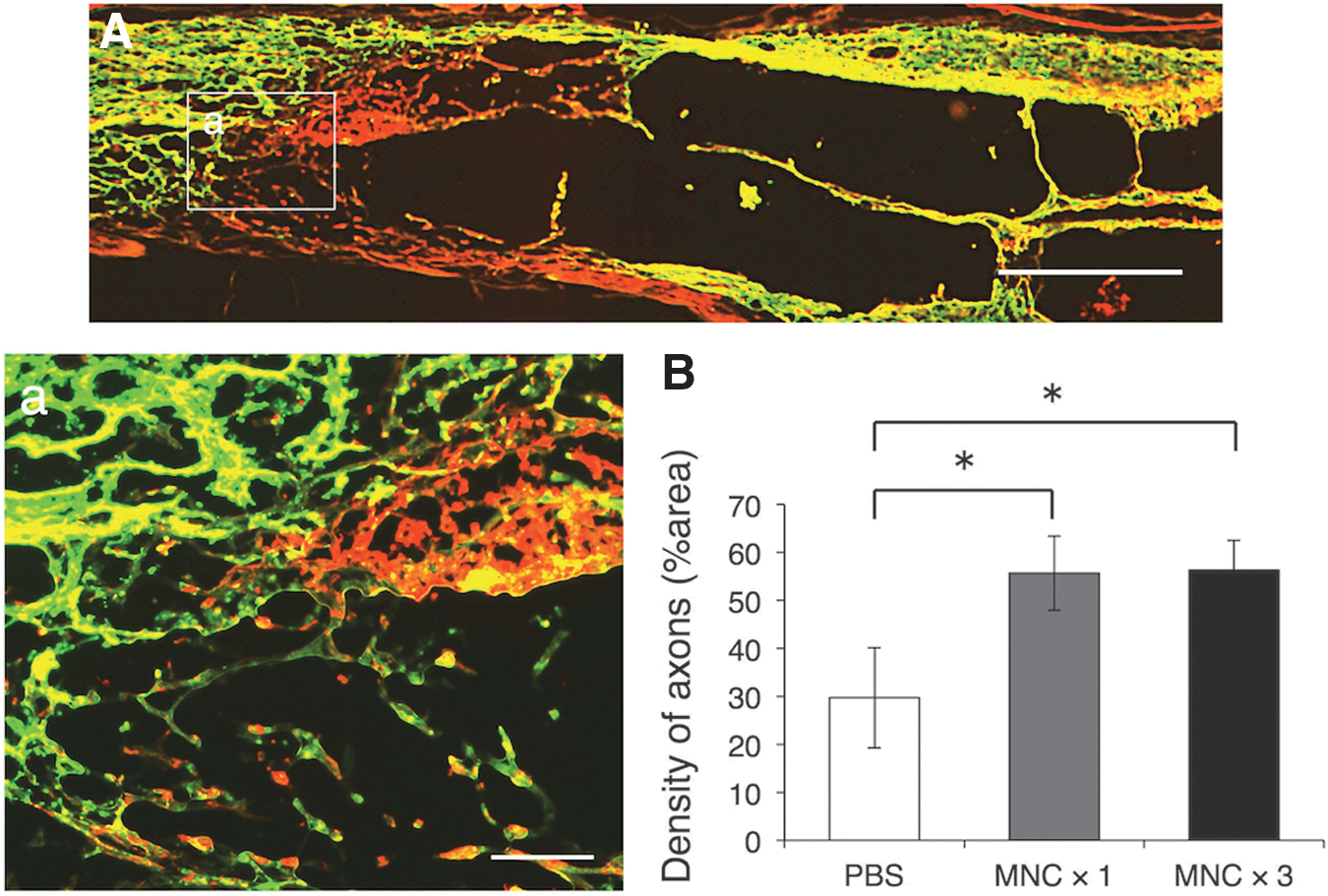
Axonal outgrowth in astrocyte-devoid areas of the phosphate-buffered saline (PBS)-injection group (control), and measurement of axon density.
Numerous axons extended through the astrocyte-devoid areas at the spinal cord lesion in both the single-injection (Fig. 2) and multiple-injection groups (Fig. 3). These axons extended with no association with astrocytes through the astrocyte-devoid areas. The values of axonal density in the astrocyte-devoid areas were 49.0 ± 2.8, 53.5 ± 7.6, and 23.1 ± 4.5% (mean ± standard error of the mean [SEM]) for the multiple-injection, single-injection, and PBS-injection groups, respectively (Fig. 4B). The axon density was much higher in the single/multiple BMNC-injection groups than in the PBS-injection group. There was no significant difference in axon density between single and multiple injection.
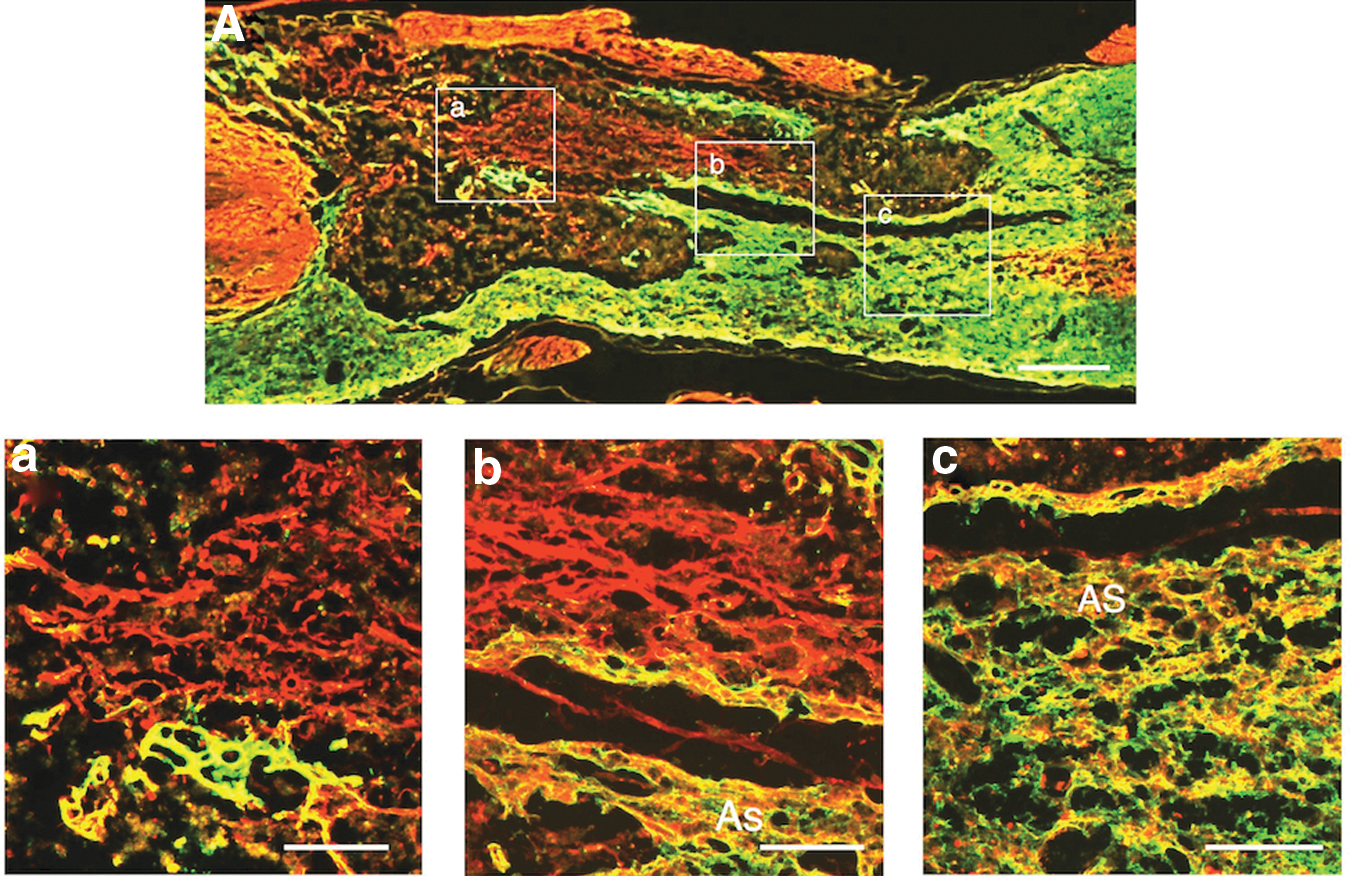
S-100 immunohistochemistry showed that axons extending through the astrocyte-devoid areas were associated with Schwann cells (Fig. 5). S-100 immunohistochemistry usually stained both Schwann cells and astrocytes. Therefore, the present study confirmed the presence of Schwann cells in the astrocyte-devoid areas by comparing the staining patterns of GFAP- and S100-immunohistochemistry in the spinal cord of rats receiving the injection of BMNCs three times (Fig. 5A). In addition, it was demonstrated that axons extending through the astrocyte-devoid areas were associated with Schwann cells by double-staining for axons and Schwann cells (Fig. 5B, 5C).
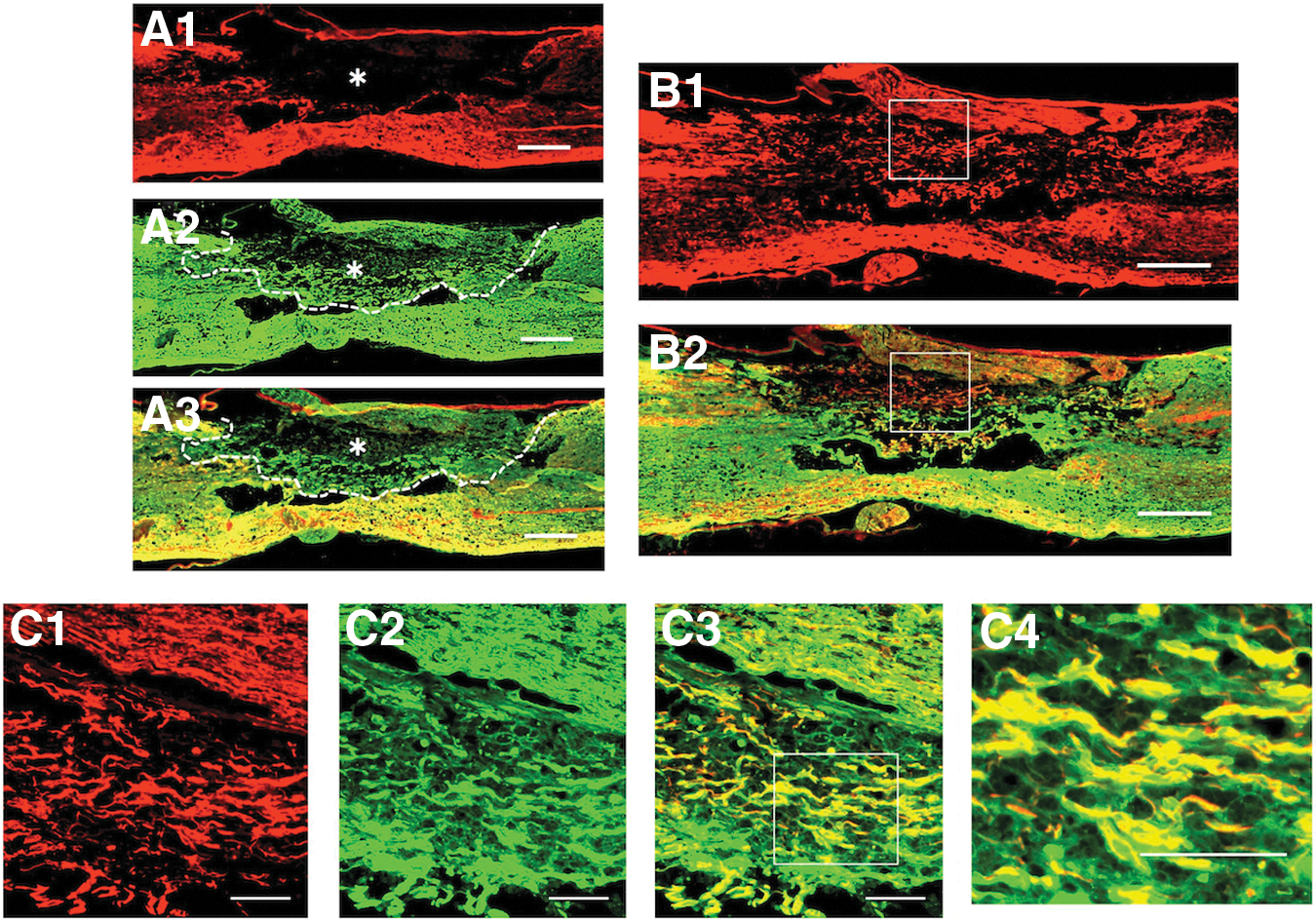
Axons extending through astrocyte-devoid areas are associated with Schwann cells. Specimens were taken from the spinal cord of the multiple-injection group.
We examined the properties of tissue matrices through which axons associated with Schwann cells extended. The previous electron microscopic studies revealed that there were some collagen fibrils in the spaces around axon-associated Schwann cells in the astrocyte-devoid areas. Therefore, the present study examined the presence and distribution of collagen type-1 in the astrocyte-devoid areas of the spinal cord of rats that received the injection of BMNCs three times. It was shown that collagen type-1 was mainly located within the astrocyte-devoid areas (Fig. 6A), and that axons extending through the astrocyte-devoid areas were embedded in such collagen type-1 matrices (Fig. 6B, 6C). These findings indicated that axons associated with Schwann cells were embedded in collagen matrices. These features suggested that axons extending through the astrocyte-devoid area had characteristics of peripheral nerve fibers.
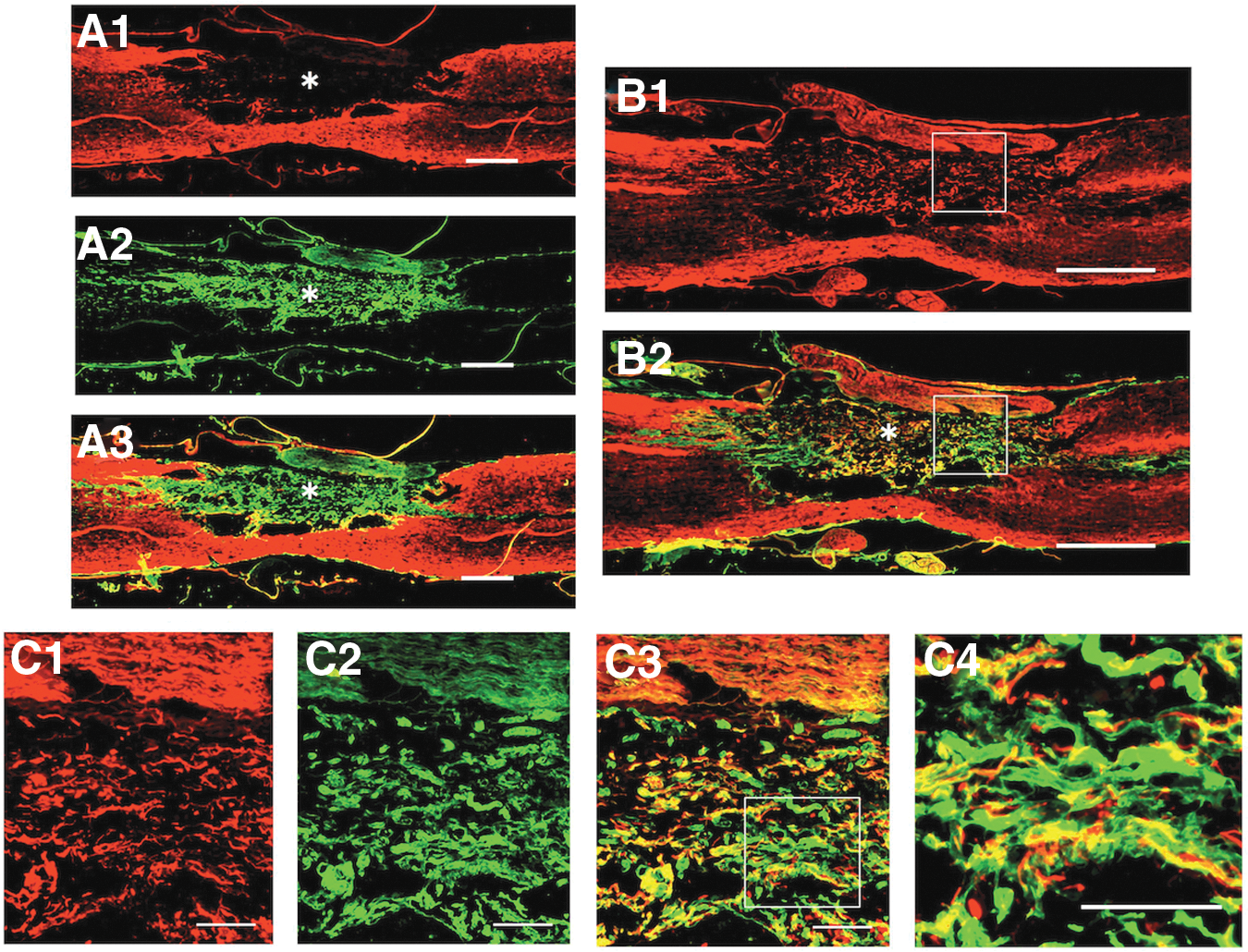
Axons extend through the collagen type-1–containing extracellular matrices in the astrocyte-devoid area. Specimens were taken from the spinal cord of the multiple-injection group.
There was no finding indicating that axons within the astrocyte-devoid areas were prevented from extending at the border of astrocyte-devoid areas (Fig. 3, Fig. 5, and Fig. 6).
Cavity formation
The cavity formation was markedly reduced by BMNC transplantation. Representative sections are presented in Figure 7A-C. The area of vacant cavities was measured in HE-stained sections and is expressed as a percentage of the spinal cord volume at the lesion. The cavity volume was ca. 37–40% in the PBS-injection group, while it was 5–10% in the single-injection group, and 3–5% in the multiple-injection group, with percentages being much smaller in the single/multiple BMNC-injection groups than in the PBS-injection group (Fig. 7D). This indicated that BMNC injection led to much more favorable tissue repair in spinal cord injury. There was no significant difference in cavity formation between the single and multiple injection.
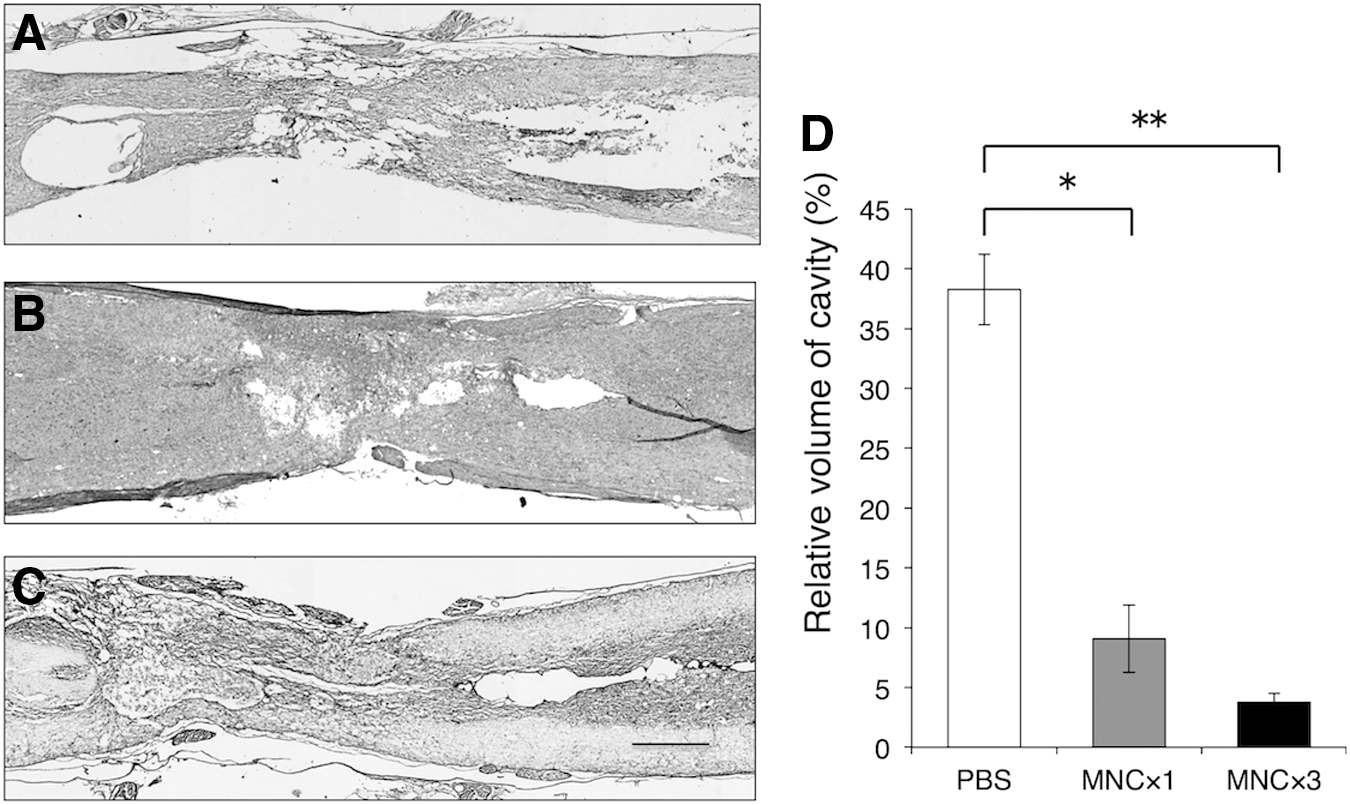
The areas of vacant cavities containing no tissue matrices were measured in several sections using the Image Filing System (Flovel, Tokyo, Japan) with a microscope (Olympus, CKX41, Tokyo, Japan), and the volumes of total cavities were calculated by multiplying the areas with the thickness of the total sections.
Locomotor behaviors
The BBB scores were markedly improved during the first 2 weeks in the multiple-injection group, followed by a moderate elevation of the score until the end of the experiment. On the other hand, the BBB scores were similarly improved in the first week in the single-injection group, followed by a moderate elevation of the score, as in the case of the multiple-injection group. It appeared that the first and second injections of BMNCs strongly promoted the locomotor improvement of rats, and that even after BMNC injection was ceased, rats continued to show a moderate elevation of BBB scores (Fig. 8).
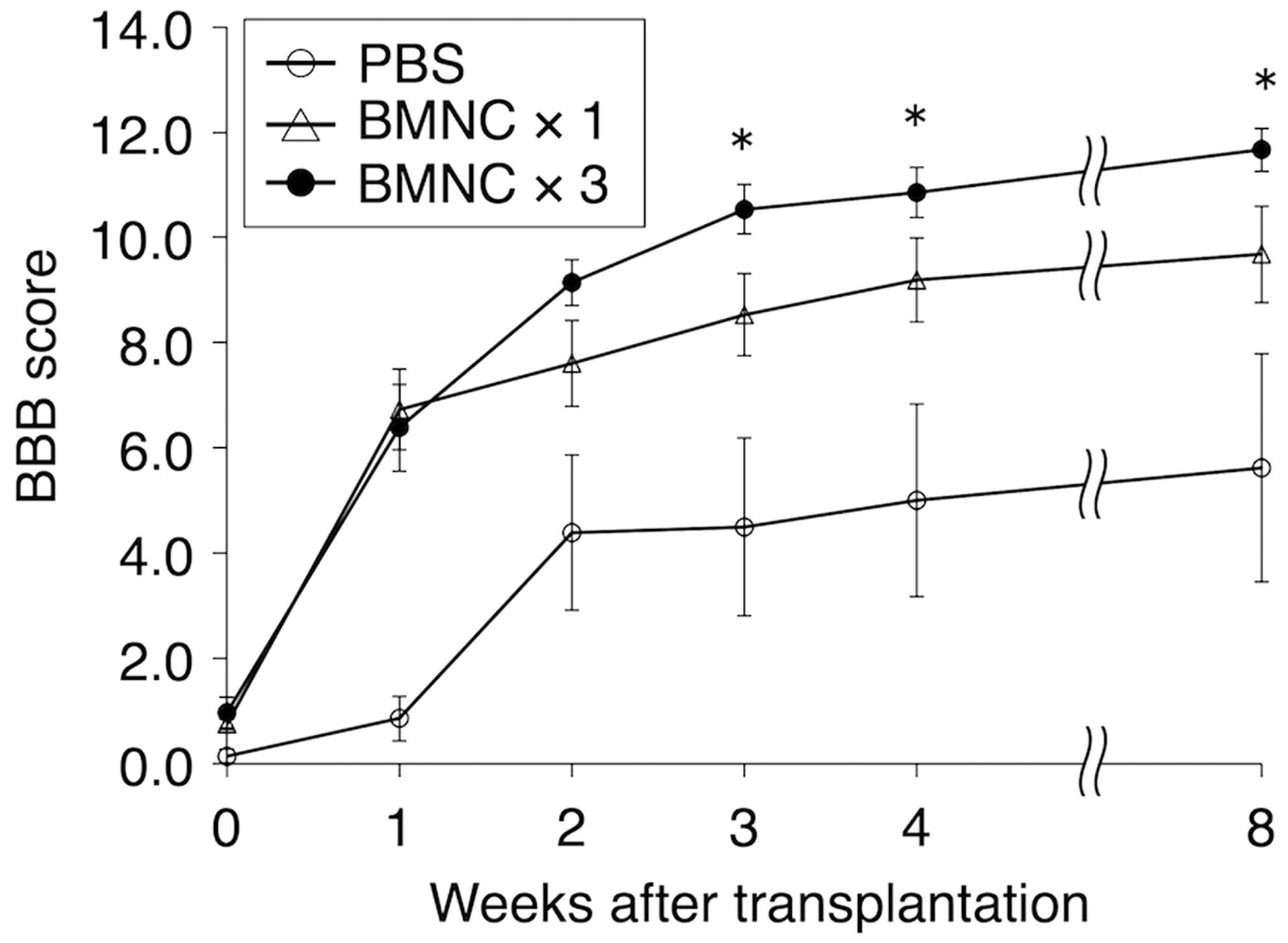
Compared with the phosphate-buffered saline (PBS)-injection group, Basso-Beattie-Bresnahan (BBB) scores elevated steeply at 1 week after bone marrow mononuclear cells (BMNC) transplantation in both the single- and multiple-injection groups, and the multiple-injection group continued to show elevation at 2 and 3 weeks after transplantation. The BBB scores were 9.7 ± 0.9 (n = 14), 11.7 ± 0.4 (n = 12), and 5.7 ± 2.17 points (mean ± standard error of the mean; n = 6) in the single-, multiple-, and PBS-injection groups, respectively. There are significant differences (p < 0.05) between the single- and multiple-injection groups at 3, 4, and 8 weeks after BMNC transplantation. The BBB scores are significantly different (p < 0.005) between the PBS- and multiple-injection groups at 1–8 weeks after transplantation. BMNC × 3, multiple injection; BMNC × 1, single injection; PBS, PBS injection (control).
The PBS-injection group showed a low level of locomotor improvement throughout the experiment. There were significant differences between the single-/multiple-injection and PBS-injection groups at 2–8 weeks after transplantation. There were significant differences between the single- and multiple-injection groups at 3, 4, and 8 weeks after BMNC transplantation. The multiple BMNC-injection group continued to maintain a score of 2–3 points higher than the single-injection group at 2–8 weeks after transplantation. BBB scores at 8 weeks post-transplantation were 9.7 ± 0.9 (n = 14), 11.7 ± 0.4 (n = 12), and 5.7 ± 2.17 (mean ± SEM; n = 6) for multiple-, single-, and PBS-injection groups, respectively. The BBB score of 9 to 10 points in the single-injection group indicated that rats walked only occasionally supporting their body weight on their hind limbs, while the BBB score of 10- to 11-points in the multiple-injection group indicated that rats could walk in coordination with the forelimbs, constantly keeping their body weight on their hind limbs. These differences in locomotor behavior are critical for actual life. Overall values of BBB scores suggested that the multiple BMNC injection more effectively contributed to the elevation of BBB scores of rats with SCI than single injection.
Discussion
In the present study, BMNCs were separated by density-gradient centrifugation from the bone marrow perfusate, and transplanted into the CSF via the fourth ventricle without cell culture. The present study demonstrated that the multiple injection of BMNCs was more effective than single injection for locomotor improvement. This indicates that multiple transplantations of BMNCs by lumbar puncture can enhance the effects on locomotor improvement clinically in patients with SCI. Since the procedures for the preparation and injection of BMNCs by lumbar puncture are technically simple, the multiple injection of BMNCs by lumbar puncture would be acceptable for patients.
Fate of transplanted BMNCs
It was shown that transplanted BMNCs did not survive long-term in the spinal cord, regardless of whether cells were injected into the spinal cord lesion or into the CSF. 12,14,15 They were not integrated into the host spinal cord to serve as scaffolds for regenerating axons in the spinal cord. Since BMNCs were not green fluorescent protein labeled, the fate of transplanted BMNCs was not traced in the present study. However, it is reasonable to consider that BMNCs disappeared within 1–3 weeks after transplantation, as in the case of a previous study. 15 The previous studies showed no finding indicating that BMNCs trans-differentiated into other cell types, such as astrocytes, oligodendrocytes, neurons, or Schwann cells, after transplantation. These considerations suggest that the effects of BMNC transplantation will be due to neurotrophic factors secreted by BMNCs. As discussed in our previous studies, 8,9,12,14,15 BMNCs are considered to release many kinds of molecules/factors, including exosomes, effective for locomotor improvement and tissue repair, including the outgrowth of regenerating axons, after transplantation. It is not known what kinds of molecules/factors are responsible for these beneficial effects. The precise mechanisms whereby BMNCs mediate functional and histological recovery remain elusive. Transplanted BMNCs are considered to survive within 1–3 weeks after transplantation. 12,14,15 These data suggest that multiple injection of BMNCs at approximately 1- to 3-week intervals will be more effective than single injection. The present study showed that multiple injection (once a week for 3 weeks) of BMNCs is more effective than single injection for locomotor improvements following spinal cord injury.
Axonal regeneration, Schwann cells, and collagen matrices
Numerous axons extended through the astrocyte-devoid areas of the spinal cord lesion in both the single- and multiple-injection groups. This finding was the same as that in our previous studies of BMSC transplantation. 12,14 The axon density was much higher in the single/multiple BMNC-injection groups than the PBS-injection group. There was no significant difference between the single- and multiple-injection groups. Both single and multiple injection of BMNCs can strongly promote axonal regeneration through the spinal cord lesion.
In the previous electron microscopic studies, we showed that axons were associated with Schwann cells surrounded by basal laminae in collagen matrices. The same findings also were observed by electron microscopy, in which choroid plexus epithelial cells were transplanted into a spinal cord lesion. 7 The present study confirmed by immunohistochemistry that regenerating axons extending through the astrocyte-devoid areas are associated with Schwann cells embedded in collagen-1–containing connective tissue matrices. There are several reports concerning collagen and laminin matrices in spinal cord injury. 19 –21 The outgrowth of numerous regenerating axons associated with Schwann cells in the collagen matrices through the astrocyte-devoid areas was unexpected as a pattern of axonal regeneration in the spinal cord. However, this pattern of axonal regeneration promoted by BMSC-transplantation should be regarded as an intrinsic ability of the spinal cord to regenerate following spinal cord injuries. 22 The appearance of Schwann cells on axons surviving after traumatic demyelination in the spinal cord lesion has been reported. 23 –25 In such a case, diameters of remyelinated axons are almost the same as those before demyelination. On the other hand, axons associated with Schwann cells in astrocyte-devoid areas included myelinated and unmyelinated axons with diameters of various sizes, as demonstrated in our previous electron microscopic studies. 7,12,14 The finding of the outgrowth of axons associated with Schwann cells through newly generated connective tissue matrices indicates that those axons are regenerating ones. 26 Sources of Schwann cells have been subjects of controversy.
Cavity formation
Cavity formation is markedly reduced by BMNC transplantation. The improvement in the single-/multiple-injection groups might be due to the production of extracellular matrices, extension of regenerating axons, and/or proliferation of glial as well as non-neural cells, such as fibroblasts, in the spinal cord lesion. By BMNC transplantation, the astrocyte-devoid areas are filled with regenerating axons and collagen matrices. It is suggested that some trophic factors secreted by BMNCs have effects on the regeneration of axons and proliferation of extracellular matrices, glial cells, and connective tissue cells at the lesion of the spinal cord, leading to the promotion of the intrinsic regeneration ability of the spinal cord following spinal cord injuries.
Clinical aspects
Our previous studies demonstrated that BMSCs and BMNCs secreted some trophic factors promoting the repair of spinal cord injury. The present study showed that such effects can continue longer-term, and may be amplified by the multiple injection of BMNCs.
The multiple injection of BMNCs is technically simple for clinical application. BMNCs can be separated from a bone marrow perfusate of the iliac bone and injected by lumbar puncture into the CSF. The interval of each cell injection can be varied depending on the conditions of patients. A 1- to 3-week interval can be regarded as reasonable, since BMNCs disappear from the spinal cord within 1–3 weeks after transplantation in rats.
BMNCs have been transplanted through various routes in clinical cases: BMNCs are applied by lumbar puncture, 17 subarachnoid space injection, 27 or intravenous (or intra-arterial) injection. 28 There is a report of many clinical cases (297 patients) treated by BMNC transplantation via lumbar puncture with good results. 29 Geffner and colleagues transplanted CD34(+) cells separated from a bone marrow perfusate into patients via multiple routes, such as direct injection into the lesion, and injection into the subarachnoid space or venous system. 30 They concluded that the transplantation of BMNCs was feasible, safe, and improved the quality of life of patients. Macrophage responses, tissue sparing, and scar formation in BMNC transplantation were comparable with those of BMSC transplantation. Jarocha and colleagues reported that repeated applications of BMNCs by lumbar puncture to patients with SCI produced clinically meaningful improvements for SCI patients. 31
Autologous BMNCs can be obtained from a bone marrow perfusate of the iliac bone, and injections of BMNCs can be safely performed by lumbar puncture in an operation room in a hospital. Such BMNC transplantation is safe and simple, and the cost of transplantation is much lower than that of other types of transplants such as BMSCs and neural stem cells that need expensive cell processing facilities for cell culture. Thus, BMNCs have a marked advantage.
A regeneration study is of significance only when the results of the study can be applied for clinical purposes. BMNCs are an appropriate candidate for clinical application: they are not genetically manipulated, but ordinary somatic cells. BMNCs are safe, since they disappear within 1–3 weeks after transplantation. They are suitable for multiple injections to promote the efficacy of transplantation.
Footnotes
Acknowledgments
This work was supported in part by grants from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (No. 230012 to CI).
Author Disclosure Statement
No competing financial interests exist.