
Abstract
The present study was designed to tackle two notorious features of mild traumatic brain injury (mTBI)—heterogeneity and invisibility—by characterizing the full scope of mTBI symptoms. Mice were exposed to brain injuries of different intensities utilizing a weight-drop model (10, 30, 50, and 70 g) and subsequently subjected to a comprehensive battery of behavioral tests at different time points and immunohistochemical examination of cortical slices. Whereas the physiological, neurological, emotional, and motor function of mTBI mice (i.e., their well-being) remained largely intact, cognitive deficits were identified by the y-maze and novel object recognition. Results from these two cognitive tests were combined and a dose-response relationship was established between injury intensity and cognitive impairment, ranging from an 85% decline after a 70-g impact (p < 0.001) to a 20% decline after a 10-g impact (essentially no effect). In addition, higher intensities of injury were accompanied by decreased expression of axonal and synaptic markers. Thus, our mTBI mice showed a clear discrepancy between performance (poor cognitive function) and appearance (healthy demeanor). This is of major concern given that diagnosis of mTBI is established on the presence of clinical symptoms and emphasizes the need for an alternative diagnostic modality.
Introduction
E
TBI, defined as an alteration in brain function caused by an external force, 3 is an extremely versatile disorder in terms of both etiology: the “external force” includes, for example, an object striking the head, a foreign body penetrating the brain, an acceleration-deceleration movement of the head, or forces generated from an explosion 3,4 ; and symptomatology: the “alteration in brain function” comprises, for example, changes in cognition (e.g., memory and executive functioning), mood (e.g., depression and anxiety), and personality (e.g., poor impulse control and apathy). 5 However, in spite of such heterogeneity in the cause and outcome of TBI, TBIs, specifically mTBIs, are usually not accompanied by any change in physical appearance. Hence, unlike a broken leg that is easily recognized by the cast, crutches, or clear signs of pain, the long-term symptoms of mTBI are notoriously “invisible.”
These two properties of mTBI, heterogeneity and invisibility, are considered among the most significant barriers to mTBI research not only in terms of funding 2 and public awareness, 6 but also with respect to: 1) diagnosis; mTBI is remarkably difficult to identify, especially given that the patients themselves frequently fail to notice and/or properly articulate the problem; 2) quantification and classification; efforts to evaluate the magnitude of mTBI are hindered because of various limitations of epidemiological studies, such as under-reporting of mild injuries and unsettled or ambiguous measurements of injury 7 ; and 3) treatment; despite all that we have learned about the underlying pathophysiology, the overall mortality of TBI patients has not decreased since 1990, 8 nor has the outcome been substantially improved. 9 In sum, it seems as though TBI fully justifies its epithet—“the silent epidemic.” 10
Given the pathophysiological heterogeneity of TBI, no single animal model can fully recapitulate the entire spectrum of findings observed in humans. 11 Still, as opposed to the “classical” models of TBI, which recruit foreign injury mechanisms such as cryogenic, 12 suction, 13 or fluid percussion, 14 and/or require a pre-injury operation to remove part of the animal's scalp 15 and skull, 16 the following weight-drop model, which is an adaptation of Feeney's procedure 16 modified in our lab, 17,18 utilizes a guided, free-falling weight to induce a diffused mechanical impact on the animal's head, which is otherwise untouched. Thus, the presented weight-drop model is a very high-throughput method, given that it is quick and convenient, and, at the same time, the most faithful imitation of traffic accidents and falls—the lion's share of TBI. 7,19,20
The present study was designed to tackle mTBI's heterogeneity and invisibility by characterizing the full scope of TBI symptoms. To do so, mice were exposed to brain injuries of different intensities (10, 30, 50, and 70g), utilizing the weight-drop model described, and subsequently subjected to a comprehensive battery of behavioral tests at different time points and immunohistochemical (IHC) examination of cortical slices.
Methods
Animal subjects and experimental design
All procedures were approved by the Sackler Faculty of Medicine Ethics Committee (01-16-021) and were conducted in strict accord with the guidelines for animal care provided by the NIH. 21 Adult ICR male mice (6–8 weeks old), weighing 25–35 g, were kept at room temperature under a constant 12-h light/dark cycle. Mice were held 5 per cage with food and water ad libitum. Each animal was used for only one experiment, with the exception of neurological assessment that was carried out on the same group of mice at several time points (Fig. 1). The minimal possible number of animals was sacrificed while all efforts were made to reduce their suffering. Without exception, all subjects survived until the end of the experiment, with no signs of illness such as weight loss, increased temperature, fur changes, or trembling.
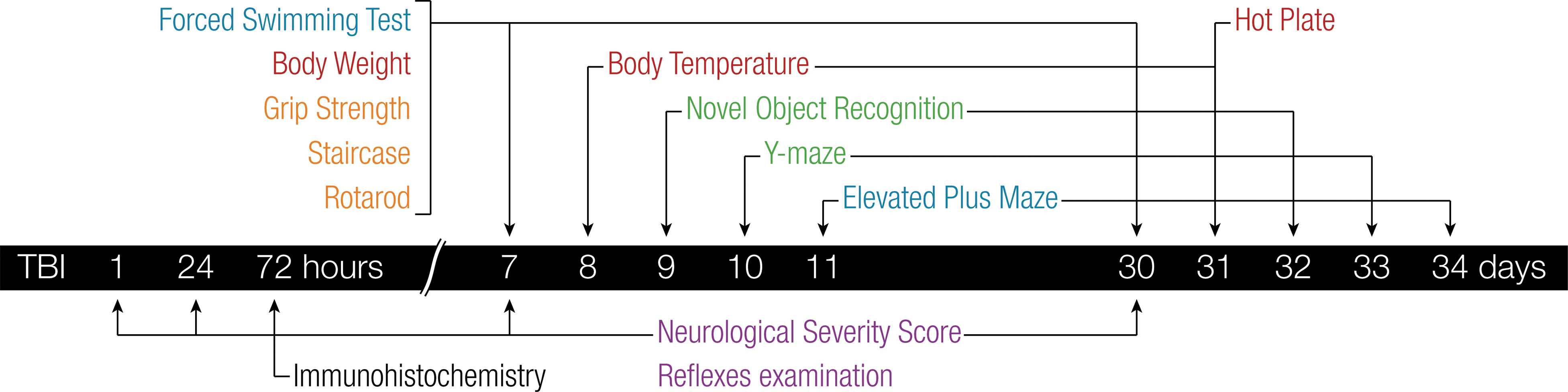
Time course of experimental analyses post-TBI. For convenience, behavioral tests were classified into five categories according to their main target of interest: neurological (purple); physiological (red); motor (orange); emotional (blue); and cognitive (green). Each animal was used for only one experiment, with the exception of neurological assessment which was carried out on the same group of mice at several time points. TBI, traumatic brain injury.
Physical and behavioral assessments were carried out approximately 1 week and 1 month after brain trauma, with the exception of neurological examination, which was also performed 1, 24, and 72 h post-impact (Fig. 1). All experiments were performed between 8:00
Traumatic brain injury procedure
TBI was induced using a weight-drop device as previously described. 18 Each mouse was lightly anesthetized with isoflurane and fixed to a sponge (10 × 15 × 6 cm), such that its right temporal bone, located between the ear and eye, was positioned directly below the opening of a metal tube (80 cm height). A rod-shaped metal weight (13 mm diameter, varying length) of 10, 30, 50, or 70 g was dropped through the tube to strike the head. Control mice were similarly anesthetized and placed beneath the metal tube without receiving any trauma. Weight mass was determined with an Ohaus CS2000 electronic scale (Ohaus, Parsippany, NJ).
Neurological tests
In complement to the Glasgow Coma Scale (GCS) that is commonly used to assess neurological impairment in TBI patients,
22
a standardized Neurological Severity Score (NSS) was used on the same group of mice (n = 5–10 per experimental group) 1 h, 24 h, 72 h, 7 days, and 30 days after brain injury. The NSS, first developed as a 25-point scale
23
and later shortened to 10 “essential” parameters,
24
includes a series of tasks designed to evaluate the motor and sensory abilities of mice. Failure to perform a given task awards the mouse 1 point, such that a higher score indicates a more severe injury (Supplementary Table 1) (see online supplementary material at
The neurological status of mice was further evaluated by testing their hindleg flexion reflex (spinal cord), corneal reflex (medulla), righting reflex (pons), and secretory signs (autonomic function), as previously described (Supplementary Table 1) (see online supplementary material at
Physiological tests
Overall health of mice (n = 5–10 per experimental group) was addressed by measuring their body temperature, using a RET-3 rectal thermometer (Kent Scientific Corporation, Torrington, CT), and weight, using an Ohaus CS2000 electronic scale (Ohaus).
Further, the hot-plate test was used to evaluate thermal nociception and pain sensitivity as an indication for peripheral integrity. 25 The apparatus consists of a transparent Plexiglas wall (28 cm height) surrounding a metal platform (30 × 30 cm), which is uniformly heated to a temperature of 50°C (±1°C). Each mouse (n = 5–10 per experimental group) was individually placed on the hot plate and the latency to the first nociceptive reaction is measured. The cut-off time was set at 20 sec to avoid tissue damage.
Motor tests
The staircase was used to evaluate exploratory and locomotor activity. 26 The apparatus consists of five identical steps (6.5 × 10 × 2 cm) enclosed by walls with a constant height (12.5 cm) along the entire length of the staircase. Each mouse (n = 9–10 per experimental group) was placed on the floor of the staircase and allowed to climb the steps repetitively for 3 min. Two parameters were measured: the number of steps ascended (NSA) and the number of rearings (NR). A step was considered climbed only if the mouse placed all four paws on it, and rearing was recorded when the mouse raised on its hind legs.
In addition, a novel grip strength procedure was developed for the purpose of evaluating neuromuscular fitness and fatigue. Each mouse (n = 10 per experimental group) was suspended on a metal bar (15 mm diameter) elevated 20 cm above a foam surface. The test consisted of two variations: once the mouse was suspended with all four limbs and once with just the two forelimbs. The latency until falling down was measured and analyzed.
Further, a four-lane accelerating rotarod (San Diego Instruments, San Diego, CA) was used to evaluate motor learning 27 (n = 8–10 per experimental group). The apparatus consists of a 3.2-cm-diameter horizontal rod, elevated 45 cm above a foam surface to intensify the subject's fear of falling without ever risking damage. The test procedure was comprised of 8 consecutive days. During the first 3 days, mice were habituated to the apparatus by placing them on the motionless rod for 2 min three times a day. During the next 5 days, mice were subjected to five trials a day, of which the best three trials were considered and averaged. A trial begins with the rod spinning at 4 rpm and gradually accelerating by a factor of 0.5 cm/sec every 5 sec, for a maximum duration of 5 min. The latency until falling from the rod was measured and analyzed such that an increased capability to stay on the spinning rod was considered an exhibition of motor learning.
Emotional tests
The Forced Swimming Test was used to evaluate depression on the basis of induced “behavioral despair.” 28 The apparatus consists of a clear Plexiglas cylinder (24 cm height, 19 cm diameter) filled with 16 cm of water at 21°C. Each mouse (n = 10 per experimental group) was placed in the cylinder for 6 min, during which it gradually stopped trying to swim. Immobility time, defined as the percent of time each mouse remained floating motionless in the water, was measured throughout the last 4 min as a manifestation of depression.
The Elevated Plus Maze was used to evaluate anxiety-related behavior on the basis of rodents' natural uneasiness toward open, elevated fields. 29 The apparatus consists of a four-armed platform resembling a “+” shape positioned 50 cm above the ground. The arms are confined by either low (“open”; 30 × 5 × 1 cm) or high (“closed”; 30 × 5 × 15 cm) walls, such that similar arms face one another. Each mouse (n = 6–16 per experimental group) was placed in the center of the maze and permitted to move freely for 5 min. The percent of time spent exploring the open arms served as a contraindication to anxiety.
Cognitive tests
The Novel Object Recognition (NOR) task was used to evaluate visual memory on the basis of rodents' natural tendency to explore new objects.
30
The apparatus consists of a square arena (60 × 60 × 20 cm). Each mouse (n = 6–15 per experimental group) was placed in the center of the arena for 5 min on three different occasions separated by a 24-h interval. During the first trial, the arena was empty. During the second trial, two identical objects were placed within the arena. During the third trial, one of the former objects was randomly replaced by a novel object, and the time spent near each object was measured. Visual memory was assessed by comparing the time spent exploring the “novel” and “familiar” (i.e., previously present) objects, normalized by the Aggleton Discrimination Index (ADI) as shown by Equation 1.
31
The Y-Maze was used to evaluate spatial memory on the basis of rodents' natural tendency to explore new environments. 30 The apparatus consists of three identical arms (8 × 30 × 15 cm) separated by a 120-degree angle. Each mouse (n = 7–15 per experimental group) was placed at the edge of the “start” arm on two occasions separated by a 2-min interval. During the first trial, lasting 5 min, one of the two remaining arms was randomly blocked, whereas during the second trial, lasting 2 min, all the arms were open and the time spent in each arm was measured. Spatial memory was assessed by comparing the time spent exploring the novel (i.e., previously blocked) and familiar arms, normalized by the ADI. 31
Immunohistochemistry
Post-injury (72 h), mice were anesthetized with a combination of ketamine (100 mg/kg) and xylazine (10 mg/kg) and perfused transcardially with phosphate-buffered saline (PBS) and then with 4% paraformaldehyde (PFA) in 0.1 M of phosphate buffer (pH 7.4). Subjects' brains were removed, fixed overnight in 4% PFA, and placed in 30% sucrose for 48 h. Frozen coronal sections (30 μm) of the temporal lobe (the site of weight impact) were cut on a sliding microtome and collected serially. Free-floating sections were first blocked by 0.1% Triton X-100 in PBS and 10% normal horse serum (NHS) for 1 h at 25°C, and then incubated for 24 h at 4°C with mouse primary antibodies raised against phosphorylated neurofilament H (pNF-H; SM1-31P; Covance Laboratories, Dedham, MA) and rabbit primary antibodies raised against Synaptophysin (b32594; Abcam, Cambridge, UK), diluted 1:500 in PBS with Tween 20 and 2% NHS. Later, sections were incubated for 1 h at 25°C with donkey anti-mouse immunoglobulin G (IgG) H&L (Alexa Flour® 488 ab150109; Abcam) and donkey anti-rabbit IgG H&L (Alexa Flour 594 ab150076; Abcam), and mounted on dry gelatin-coated slides for fluorescence visualization using a Leica SP5 confocal microscope (Leica, Wetzlar, Germany). Excitation light was provided by the 488-nm line of argon lasers for the DyLight-488 fluorophore, the 543-nm line of HeNe lasers for the DyLight-594 fluorophore, and the 405-nm line for 4′,6-diamidino-2-phenylindole (DAPI). Scanning parameters were fixed for all images. For each section, images were taken from three fields at the post-central cortex. Overall, we had two to four coronal sections from each brain along the anterior-posterior axes of the temporal lobe.
Statistical analysis
Statistical analysis was performed using IBM SPSS 23 software (Genius Systems, Petah Tikva, Israel). Behavioral tests were analyzed by one-way analysis of variance (ANOVA) with a Bonferroni correction applied for post-hoc comparisons. Three tests were analyzed differently: 1) The 1-cm beam walking task was analyzed as a binary variable utilizing a generalized linear mixed model of logistic regression, with time as the repeated measure; 2) rotarod data were analyzed with a mixed ANOVA model that included one between-subjects variable (TBI intensity) and one within-subjects variable (time); and 3) comparison of the two cognitive tests (the NOR and Y-maze) at the two time points examined was performed with a three-way ANOVA (TBI-intensity × test × time). All results are given as mean ± standard error of the mean, excluding 1-cm beam walking, which is presented as percent failure. Nominal significance levels were set at 0.05 (“*”), 0.01 (“**”), and 0.001 (“***”).
Results
Neurological function
Mice were subjected to neurological assessment 1 h, 24 h, 72 h, 7 day, and 30 days post-injury. None of the injured mice showed abnormalities in any of the neurological tests used, with the exception of one item. Hence, looking at the NSS scores as a whole, all mice received ≤1 points (Fig. 2A). The NSS, which was developed as an injury severity scale parallel to the GCS in humans, classifies injuries according to their score 1-h post-trauma: >9 = fatal; 7–8 = severe; 5–6 = moderate; and <5 = mild. 32 –34 Thus, the brain injuries induced in this study should be all classified as mild.
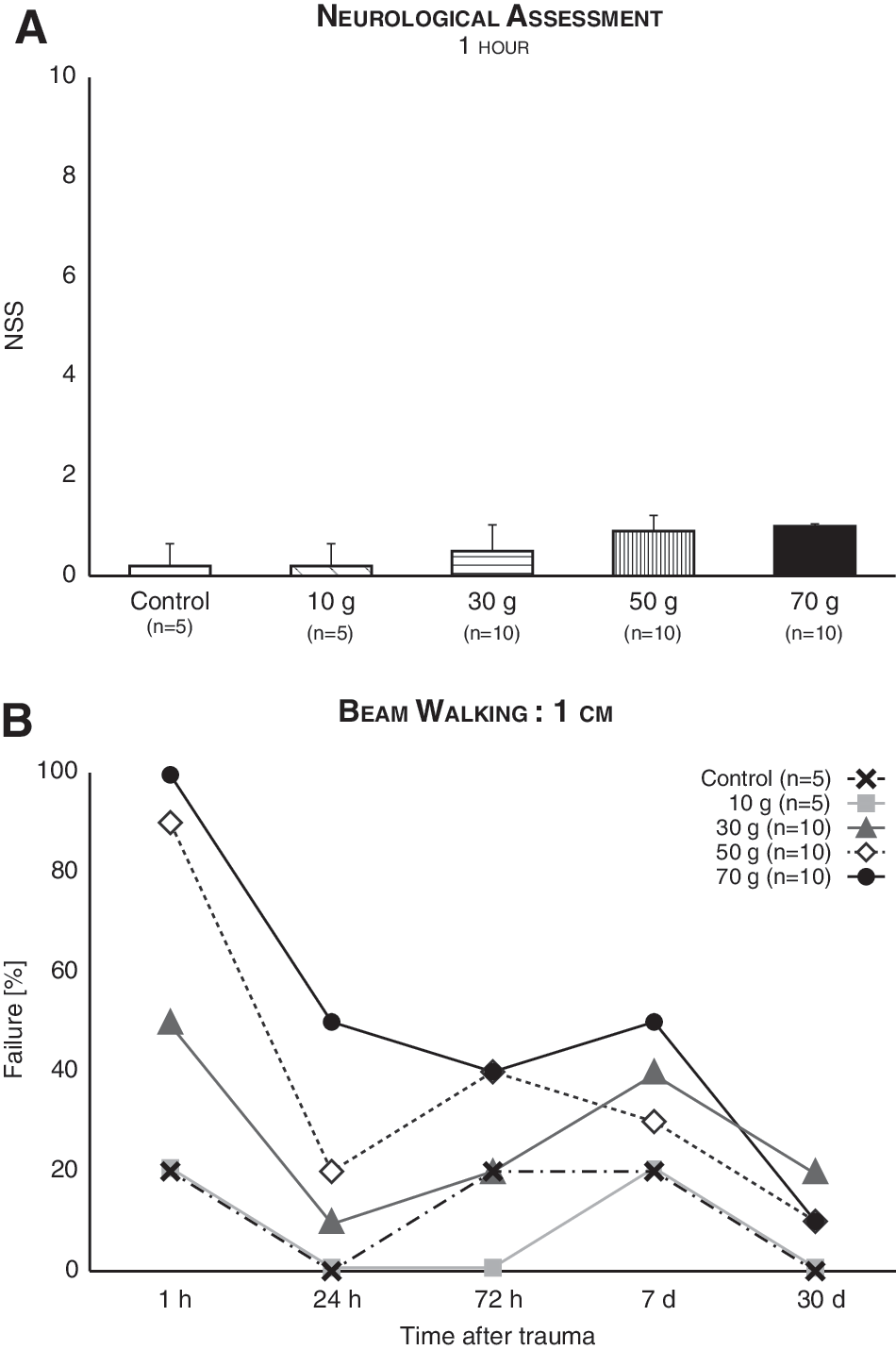
Neurological function post-mTBI remained largely intact. (
The single item that did demonstrate variance between mice, the 1-cm beam walking task, was isolated from the NSS and analyzed separately as a binary variable (1 = failure; 0 = success). Logistic regression of repeated measures revealed a significant main effect for TBI intensity (F (4,190) = 3.82; p = 0.005) and time (F (1,190) = 5.21; p = 0.02), with a nonsignificant interaction (F (4,190) = 1.93; n.s.; Fig 2B). Thus, the ability to walk on a 1-cm-wide beam was affected by TBI, but improved over time.
Physiological function
mTBI at all intensities did not affect the physiological function of mice: neither the animal's body temperature 8 and 31 days post-injury (F
(4,40) = 0.51; not significant [n.s.], and F
(4,19) = 0.60; n.s., respectively; Supplementary Fig. 1A,B) (see online supplementary material at
Motor function
Motor function remained intact post-mTBI: In the staircase test, all experimental groups demonstrated equal exploratory and locomotor activity 7 and 30 days post-impact, as evident by the NSA (F
(4,44) = 0.47; n.s., F
(4,45 = 0.37; n.s., respectively; Supplementary Fig. 2A,B) (see online supplementary material at
Similarly, no differences in neuromuscular fitness and fatigue were found 7 and 30 days post-impact, as evident by the time mice remained suspended on the grip strength bar using two (F
(4,45) = 2.27; n.s., F
(4,45) = 0.56; n.s., respectively; Supplementary Fig. 2E,F) (see online supplementary material at
The accelerating rotarod test performed 7 (Fig. 3A) and 30 (Fig. 3B) days post-injury was analyzed with a mixed ANOVA model that included one between-subjects variable (TBI intensity) and one within-subjects variable (time). Whereas the performance of mice improved over time (7 days, F (4,164) = 27.19; p < 0.001; 30 days, F (4,172) = 22.04; p < 0.001), experimental groups did not differ from one another (7 days, F (4,41) = 0.31; n.s.; 30 days, F (4,43) = 0.19; n.s), nor was the interaction between time and TBI intensity significant (7 days, F (16,164) = 0.95; n.s.; 30 days, F (16,172) = 1.16; n.s.). These results contradict previous findings in mice 35,36 and rats 37 of a significant effect of TBI on rotarod performance, a discrepancy that may be explained by the dissimilar rotarod protocol and/or animal model used in our study, which was designed to mimic the most subtle of TBIs.
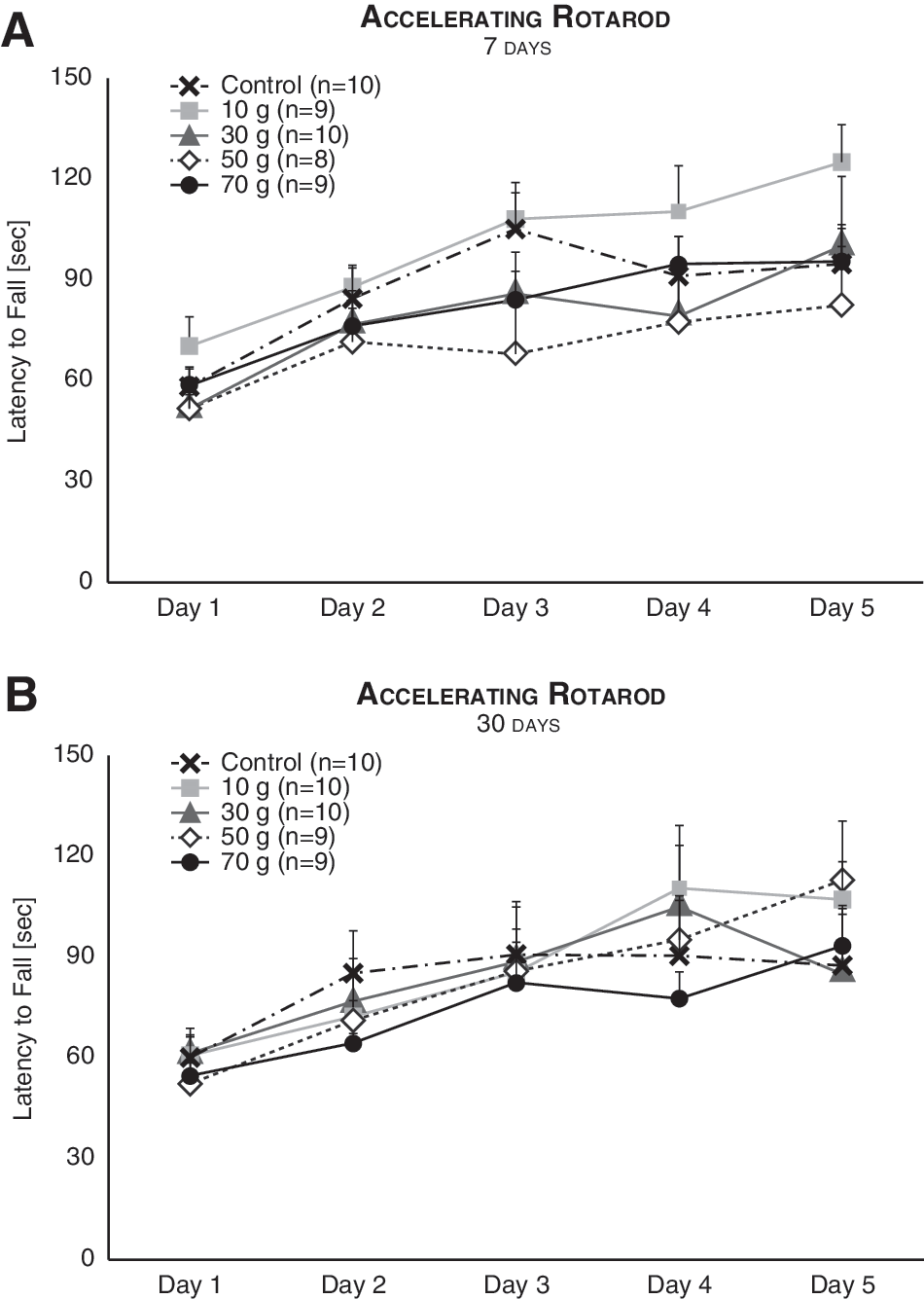
Motor learning was unaffected by mTBI. The performance of mice in the accelerating rotarod improved with time both 7 (
Emotional function
In the Forced Swimming Test, experimental groups differed 7 days post-injury (F (4,55) = 3; p = 0.03), such that mice inflicted with a 70-g weight drop demonstrated greater immobility time compared to control mice (p = 0.003; Fig 4A). Similarly, immobility time after 30 days increased (F (4,45) = 3.31; p = 0.02) subsequent to a 50- and 70-g impact (p < 0.01; Fig 4B). These results implicate, in congruence with previous findings in mice 38,39 and humans, 40 that high-intensity mTBI may lead to depressive-like behavior.
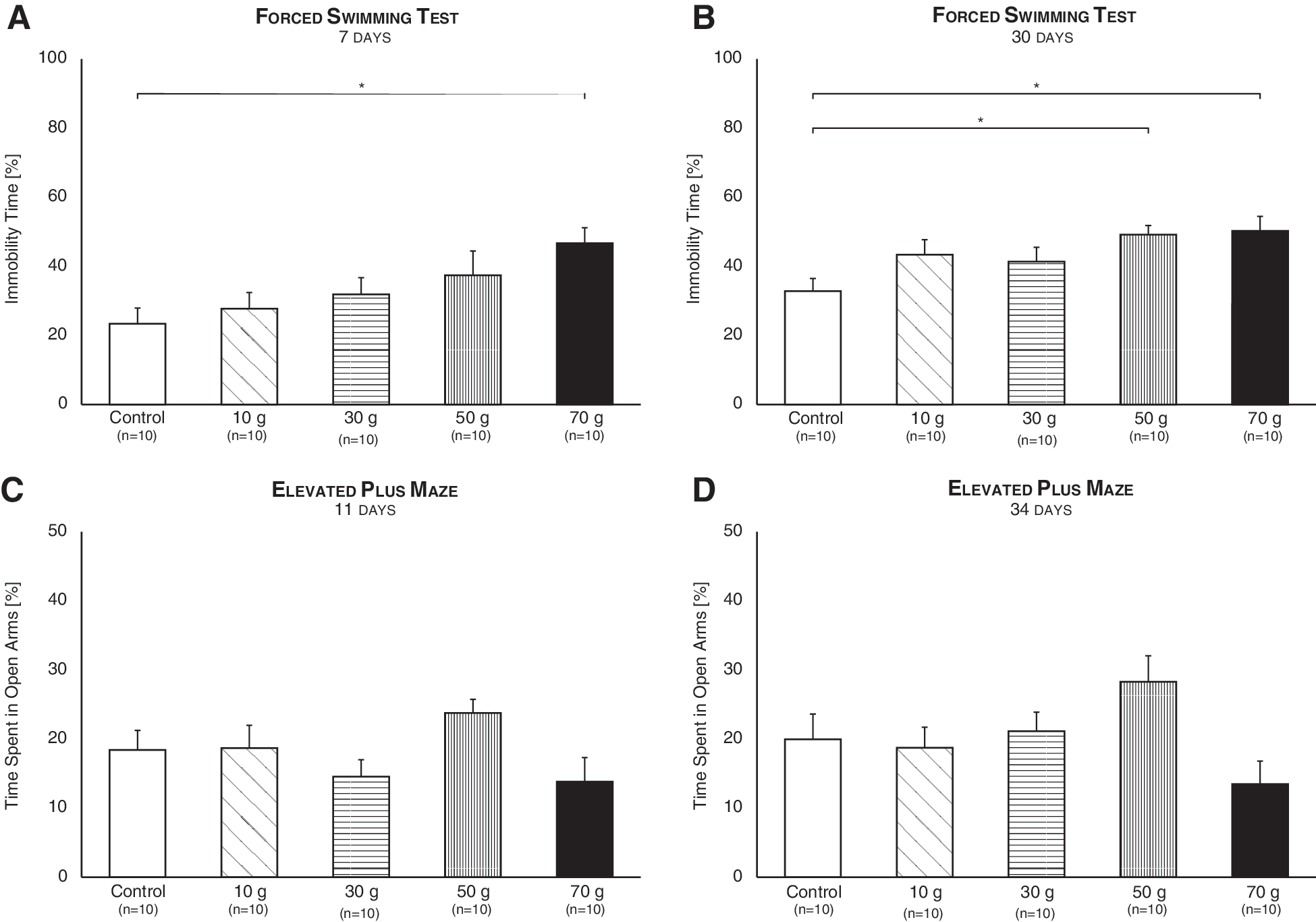
Emotional function post-mTBI. The Forced Swimming Test performed 7 (
In the Elevated Plus Maze, all experimental groups spent an equal amount of time in the open arms, and thus exhibited normal anxiety-like behavior, 11 and 34 days post-impact (F (4,61) = 2.12; n.s., F (4,57) = 1.60; n.s., respectively; Fig. 4C,D). This is in congruence with several publications, 29,41,42 but in contrast with others that demonstrate that TBI may increase 33,43 or decrease anxiety. 44,45
Cognitive function
NOR performance differed between experimental groups 9 days post-injury (F (4,55) = 3; p = 0.03), such that TBI mice functioned poorly compared to control mice subsequent to 30- (p < 0.05), 50- (p < 0.05), and 70-g (p < 0.01) weight drops (Fig. 5A). An equivalent difference was found 32 days after brain trauma (F (4,55) = 7.20; p < 0.001). Specifically, 30-, 50-, and 70-g weight drops led mice to function poorly compared to control mice (p < 0.001, p < 0.01, and p < 0.001, respectively), and 10-g-inflicted mice (p < 0.01, p < 0.05, and p < 0.001; respectively; Fig. 5B).
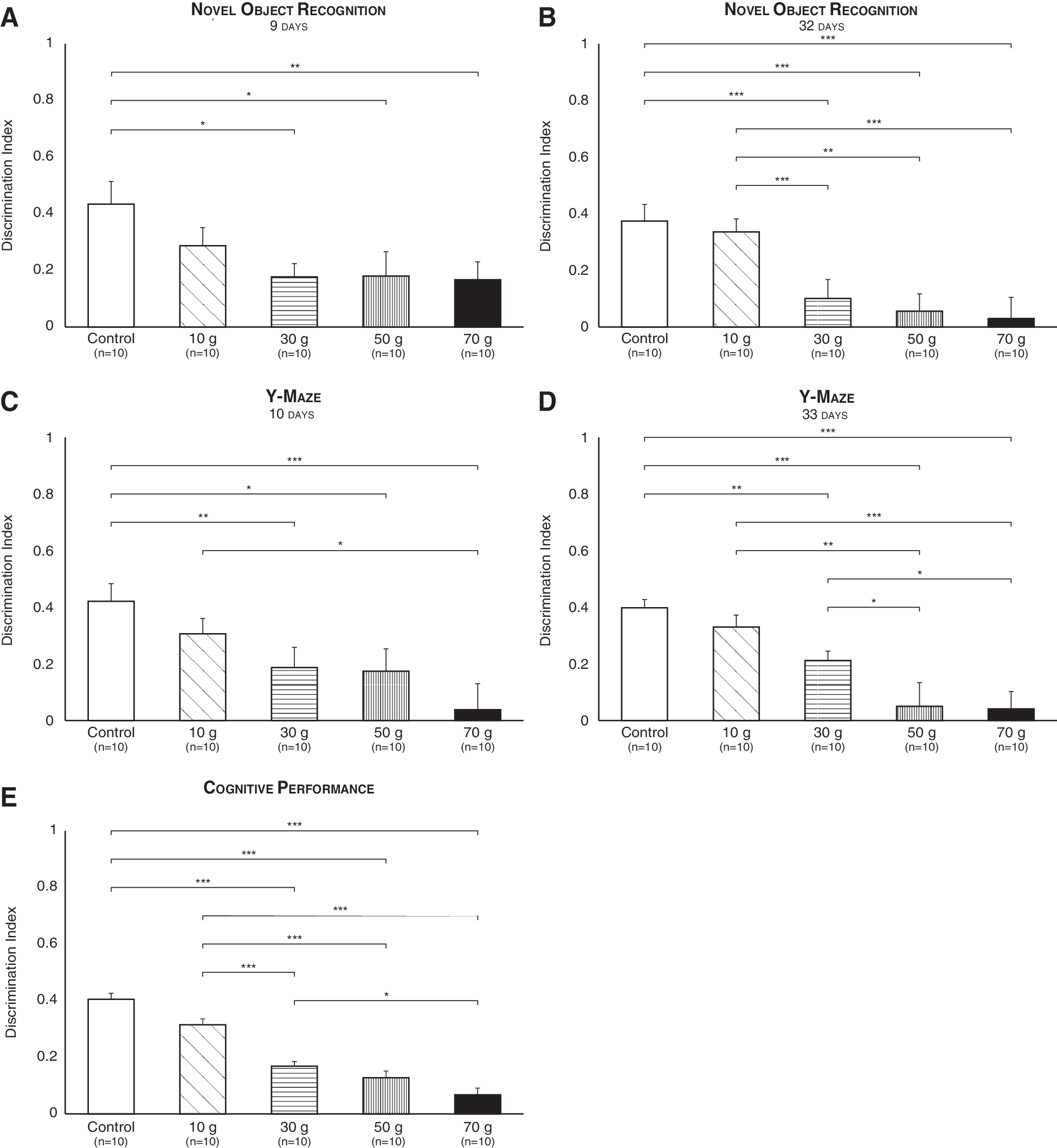
Dose-response relationship between mTBI intensity and cognitive function. The NOR submitted 9 (
Similarly, Y-maze performance differed between experimental groups 10 days post-injury (F (4,55) = 4.50; p = 0.003). Compared to control mice, TBI mice functioned poorly following 30- (p < 0.01), 50- (p < 0.05), and 70-g (p < 0.001) weight drops. Further, a 70-g impact induced a significantly robust effect compared to a 10-g impact (p < 0.05; Fig. 5C). An equivalent difference was found 33 days after brain trauma (F (4,55) = 10.20; p = 0.000), demonstrating that TBI mice functioned poorly compared to control mice subsequent to 30- (p < 0.01), 50- (p < 0.001), and 70-g (p < 0.001) weight drops, and that 50- and 70-g impacts induced a significantly robust effect compared to 10- (p < 0.01 and p < 0.001, respectively) and 30-g (p < 0.05) impacts (Fig. 5D).
Because results from both the NOR and Y-maze are given as ADI scores, 31 we could compare them by means of a three-way ANOVA (TBI-intensity × test × time), which returned a nonsignificant probability for the effect of test (F (1) = 0.03, n.s.), time (F (1) = 2.33, n.s.), and test × time (F (1) = 0.28, n.s.). This allowed us to combine data from the two tests at the two time points examined and create a novel variable: cognitive performance (Fig. 5E). Cognitive performance was analyzed by one-way ANOVA, which revealed a significant effect of TBI (F (4,230) = 21.12; p < 0.001). Specifically, 30-, 50-, and 70-g weight drops led mice to function poorly compared to control and 10-g-inflicted mice (p < 0.001). Further, the effect of a 70-g impact was significantly robust compared to a 30-g impact (p < 0.05; Fig. 5E).
Thus, a dose-response effect of TBI on cognitive performance was established, this despite the fact that all injury intensities were classified as mild according to their NSS score. 32 –34
Immunohistochemistry
Cortical neurons were assessed by double staining coronal sections of the temporal lobe (the site of weight impact) with antibodies raised against pNF-H, a marker for axonal neurofilaments and neuronal regeneration, 46 and Synaptophysin, a marker for synaptic density. 47 In addition, DAPI was used to stain DNA and mark cell nuclei. As qualitatively shown in Figure 6, TBI reduced axonal and synaptic markers expression in a dose-dependent manner, that is, higher impact intensities led to a stronger decline in fluorescence. No notifiable alternation was observed in the number of cells.
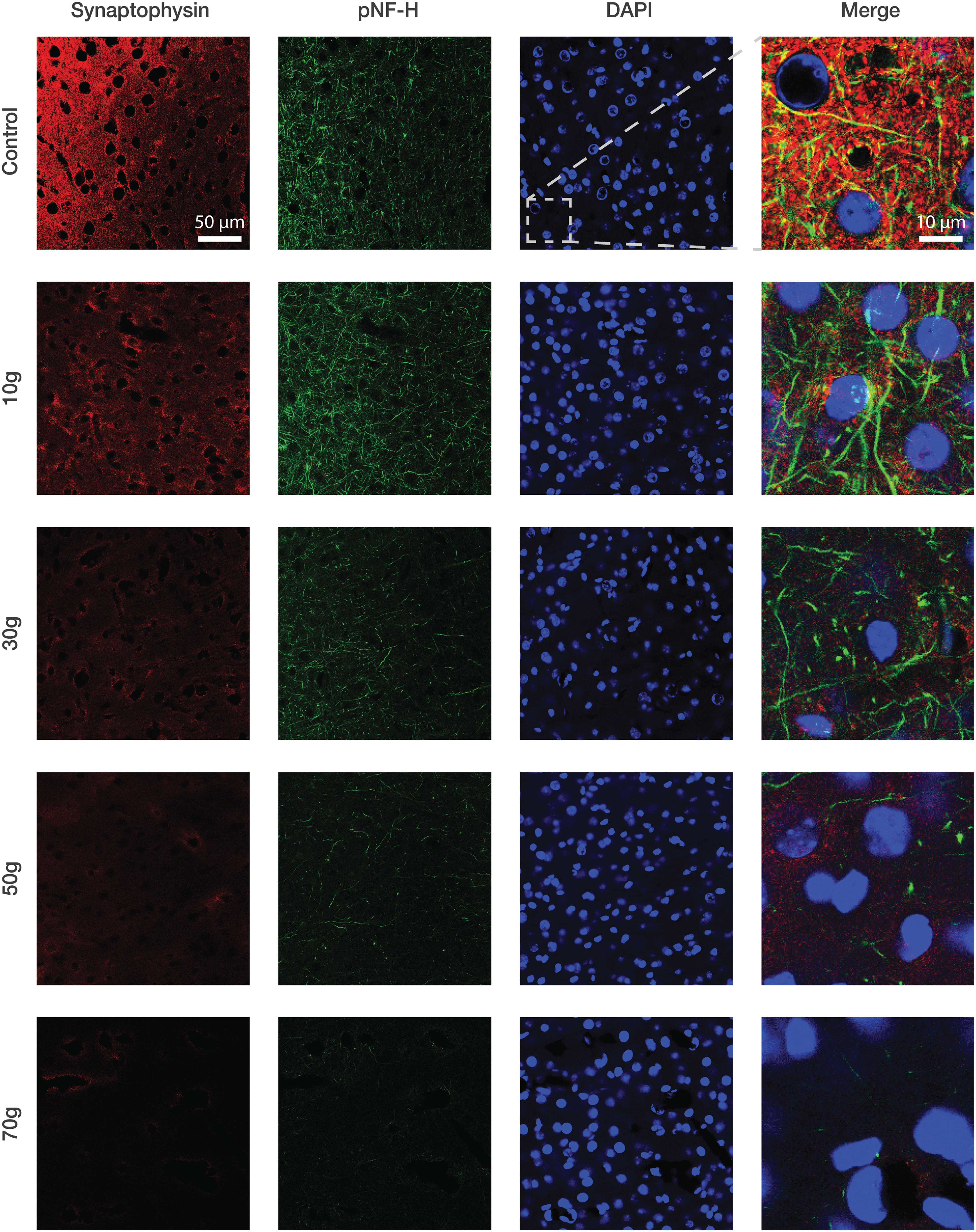
mTBI reduced expression of axonal and synaptic markers in cortical slices of the temporal lobe. Representative images of cortical sections taken 72 h post-injury and immuno-stained with phosphorylated neurofilament H (pNF-H; green), Synaptophysin (red), and 4′,6-diamidino-2-phenylindole (DAPI; blue). Merge images are × 5 magnifications of the area framed in white. The same contrast and brightness was applied to all images. mTBI, mild traumatic brain injury.
Discussion
In an attempt to characterize the profile of TBI symptoms, mice were inflicted with TBI of different severities and subjected to a comprehensive battery of behavioral tests at different time points, and IHC examination of cortical slices (Fig. 1). A dose-response relationship was established between the severity of injury, that is, the mass of the weight dropped on the animal's head, and the deterioration of cognitive abilities, assessed by the NOR and Y-maze (Fig. 5). This dose-response effect is congruent with findings in human patients 48 –51 and provides important validation to our TBI model. 52
Inspection of neuronal damage post-mTBI was achieved by staining cortical slices with antibodies raised against pNF-H and Synaptophysin, markers for axonal neurofilaments and synaptic density, respectively. In parallel to the cognitive deficits displayed by injured mice, expression of pNF-H and Synaptophsin decreased in an intensity-dependent manner (Fig. 6). The resemblance between these two findings—deteriorating cognitive abilities and decaying immunofluorescence of axonal and synaptic markers—might suggest that a decline in intercellular communication participates in the underlying pathophysiology of mTBI cognitive sequelae.
No differences between control and injured mice were found in neurological status (NSS; Fig. 2a), anxiety (Fig. 4C,D), rectal temperature (Supplementary Fig. 1A,B) (see online supplementary material at
Thus, even though the appearance of TBI mice was essentially indistinguishable from healthy mice, the NOR and Y-maze revealed that they exhibited significant cognitive impairments, specifically spatial and visual memory. Such a discrepancy should be of major concern given that diagnosis of TBI is established on the basis of clinical symptoms, mainly the GCS. 22,56 Our results indicate that even when brain-injured mice present the same neurological status as healthy mice, they may still experience an 85% cognitive impairment (cognitive performance of 70-g TBI vs. control; Fig. 5E).
Noticeably, 10-g-inflicted mice were consistently indistinguishable from control mice, implying that certain low-intensity mTBIs, when they occur as a singular event, may have no discernable impact. However, such seemingly innocuous assaults may be destructive if repeated. Repetitive mTBI has recently received much attention as the underlying cause of chronic traumatic encephalopathy (previously known as punch-drunk syndrome), a progressive degenerative disease prevalent among contact sports participants and war veterans. 5,57 Thus, the military, scientific, and medical communities might benefit from a detailed characterization of the frequency-dependent effect of mTBI, alongside the dose-dependent effect established in this study. For example, if one knows the minimum latency between two 10-g weight drops that produces no effect on mice, one can begin to estimate the recovery time needed to prevent multiple injuries from interacting.
The single neurological task that showed a significant effect of TBI also showed a significant effect of time (beam walking, 1 cm; Fig. 2B), corresponding to the well-documented phenomenon of spontaneous recovery after brain trauma. 24,32,58 –60 It may be tempting to interpret the restoration of neurological function as a sign of health, albeit here we demonstrated that in spite of intact neurological status, cognitive testing may still reveal persisting abnormalities 30 days post-TBI (Fig. 5B,D), perhaps even more. 18,38,61 This should be kept in mind when deciding whether to allow clinically asymptomatic individuals to return and serve in combat zones, or play in high-risk sports such as boxing or American football. Further, this might also encourage health care and insurance providers to recognize TBI as a possibly chronic disorder rather than a single acute event. 62
Severity of TBI is unevenly distributed within the population, with 70–90% of all treated brain injuries classified as mild. 63 Nonetheless, these are the injuries most difficult to diagnose. 7 Our results suggest that despite mTBI's reputation as a heterogeneous disorder, certain symptoms may have a stronger, longer-lasting impact than others and thus possess greater diagnostic value. Specifically, cognitive performance has been proven sensitive enough to differentiate mTBI severities that were otherwise, according to the NSS, clustered together. 32 –34 In analogy, perhaps the GCS, which has evolved into a universal classification system for the severity of TBI, 56 should be supplemented with a more-sensitive tool. Accumulating evidence suggests that a holistic approach, namely neuropsychological assessment, may assist with diagnostic issues by relying on multiple, complex, and interactive sources of data. 64 –66 Notwithstanding, such programs demand a clinical interview followed by a variety of tests performed by a trained neuropsychologist, 65 and given the rising incidence of TBI worldwide, which is already estimated at 50 to 500 injuries per 100,000 people, 7 such higher order analysis may not always be practical, especially in developing countries. Alternatively, imaging techniques, such as computed tomography, magnetic resonance imaging, and single-photon emission computerized tomography, have already been proved useful when formulating TBI diagnoses. 67,68 However, when applied to mild injuries, conventional structural and/or functional scans fail to detect subtle changes in the brain such as diffuse axonal injury, the most common abnormality present in mTBI. 69,70
The results of our study highly recommend cognitive testing, for example, memory evaluation, 71 as a diagnostic modality, which is both sensitive and feasible and therefore suitable to complement the GCS in detecting mTBI.
Footnotes
Acknowledgments
We are grateful to Ms. Ilana Gelernter (M.A.) from the Statistical Consulting Laboratory, School of Mathematical Sciences, Tel-Aviv University, and Mr. Alex Smolar (M.A.) from Prof. Daniel Michaelson Laboratory, Department of Neurobiochemistry, Tel-Aviv University, for their help.
This work was supported, in part, by the Ari and Regine Aprijaskis Fund, Tel-Aviv University, Tel-Aviv, Israel.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.