
Abstract
Myelination in the central nervous system (CNS) is a dynamic process that includes birth of oligodendrocyte progenitor cells (OPCs), their differentiation into oligodendrocytes, and ensheathment of axons. Regulation of myelination by neuronal activity has emerged as a new mechanism of CNS plasticity. Activity-dependent myelination has been shown to regulate sensory, motor, and cognitive functions. In this work, we aimed to employ this mechanism of CNS plasticity by utilizing induced neuronal activity to promote remyelination and functional recovery in a subchronic model of spinal cord injury (SCI). We used a mild contusive SCI at T10, which demyelinates surviving axons of the dorsal corticospinal tract (dCST), to investigate the effects of induced neuronal activity on oligodendrogenesis, remyelination, and motor function after SCI. Neuronal activity was induced through epidural electrodes that were implanted over the primary motor (M1) cortex. Induced neuronal activity increased the number of proliferating OPCs. Additionally, induced neuronal activity in the subchronic stages of SCI increased the number of oligodendrocytes, and enhanced myelin basic protein (MBP) expression and myelin sheath formation in dCST. The oligodendroglia regenerative response could have been mediated by axon-OPC synapses, the number of which increased after induced neuronal activity. Further, M1-induced neuronal activation promoted recovery of hindlimb motor function after SCI. Our work is a proof of principle demonstration that epidural electrical stimulation as a mode of inducing neuronal activity throughout white matter tracts of the CNS could be used to promote remyelination and functional recovery after CNS injuries and demyelination disorders.
Introduction
M
In the injured spinal cord, demyelination occurs as a result of oligodendrocyte death. 7 Spontaneous remyelination occurs even in chronic stages of spinal cord injury (SCI), 8 but it is insufficient to prevent long-term neurological disability. Therefore, augmenting the intrinsic remyelination response remains an important therapeutic direction for treatment of SCI and other myelin-related disorders. 9 Augmentation of oligodendrogenesis that leads to myelin remodeling by modulation of neuronal activity, where neuronal activation was induced through electrical stimulation, was first shown in an in vitro model. 10 Induction of activity through electrical stimulation of the medullary pyramid increased oligodendrogenesis in the dorsal corticospinal tract (dCST) throughout the spinal cord. 11 In addition, optogenentic activation of M2 layer V projection neurons promoted oligodendrogenesis and remodeled myelin in subcortical white matter tracts; moreover, this mode of induced activation was accompanied by changes in motor function. 12
Here, we induced neuronal activity through electrical stimulation of the M1 cortex after a mild thoracic contusion SCI that spared but demyelinated dCST axons. We show that induced neuronal activation in subchronic SCI promotes oligodendrogenesis and remyelination, and generates motor function recovery. We provide a novel strategy for promoting remyelination in CNS injury and other demyelination disorders.
Methods
Animals and experimental timeline
Twenty-seven female Sprague–Dawley rats (250 ± 25 g) were used. The Johns Hopkins University Institutional Animal Care and Use Committee approved all the procedures performed in this study. Twenty-four animals were divided into stimulated and sham-stimulated cohorts (n = 12 per cohort). Three additional rats were used to confirm the severity of the mild SCI model, primarily showing that it causes demyelination in the injury epicenter while preserving dCST axons (one was naïve; one underwent an SCI; one underwent an SCI and received an injection of biotinylated dextran amine [BDA]). Day 0 denotes the day that animals received the SCI. On day 28, BDA was injected into the M1 cortex, and electrodes were implanted over the same region. Electrical stimulation was performed daily starting on day 28 and continued until the experimental end-point on day 49. On days 42–49, 5-bromo-2’-deoxyuridine (BrdU, Roche) was administered daily. All animals were euthanized at day 49 (Fig. 1).
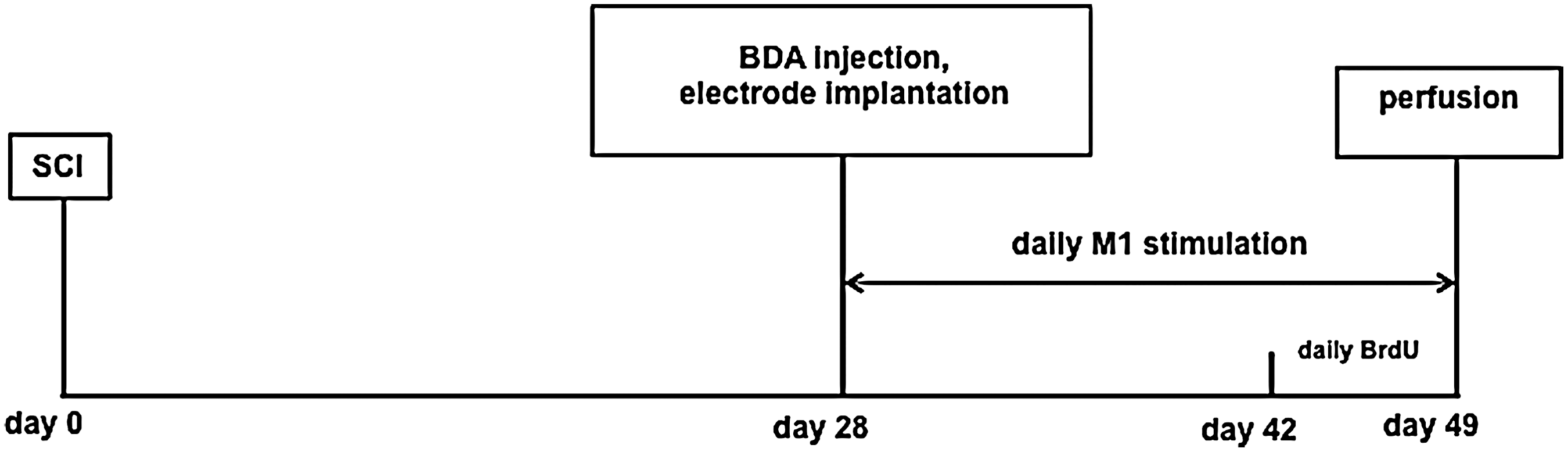
Experimental timeline. Animals received mild contusive spinal cord injury (SCI) on day 0. Biotinylated dextran amine (BDA) was injected unilaterally in the M1 cortex, and electrodes were implanted over M1 on day 28. Electrical stimulation was performed for 4 h daily from day 28 to day 49. During the last week of stimulation (days 42–49), 5-bromo-2’-deoxyuridine (BrdU) (50 mg/kg) was administered intraperitonally (i.p.) daily. All animals were euthanized at day 49.
Mild contusive SCI
Animals were anesthetized with ketamine/medetomidine cocktail (75:0.5 mg/kg), and a dorsal laminectomy was performed. A midline incision was made, and the paravertebral muscles were dissected to expose the spinal column at the level of T9–T11. Clamping the T9 and T11 spinous processes stabilized vertebral column. The T10 dorsal lamina was then removed. The dorsal surface of the spinal cord at T10 with intact dura was exposed, and hit with a 40 kdyn force using an Infinite Horizon Impactor (Precision System). Following the impact, muscles were sutured, and the skin was stapled. Animals received intramuscular (i.m.) injections of 0.5 mL atipamezole (Antisedan, Pfizer), and were kept in a 37°C incubator until they recovered from anesthesia. Subsequently, animals were hydrated through subcutaneous (s.c.) injections of 10 mL saline, and were administrated 0.12 mg/kg Enrofloxacin (Baytril, Bayer) i.m. for 3 consecutive days. Bladders of the rats were expressed manually three times daily until reflex bladder emptying was established, which occurred the following day.
Anterograde tracing of CST
On day 28, anterograde tracer BDA (Molecular Probes) was injected into the cortex to label the CST. Briefly, animals were anesthetized again as described, and the head was fixed in a stereotactic frame (Kopf Instruments). Unilateral craniotomy was performed over the hindlimb representative area of the M1 cortex, and the intact dura was exposed. The glass pipette (tip diameter: 20 μm) containing 5 μL 10% BDA in saline was lowered from the surface of the dura to a depth of 1 mm (cortical layer V). BDA was iontophoresed for 20 min (7 sec on, 7 sec off; 5 μA). Four injections were made within the lateral portion of the hindlimb representation. After injections were completed, we waited for 2 min before removing the pipette.
Electrode implantation and electrical stimulation
Immediately after BDA injection, animals received electrode implantation as previously described. 13 Two 127 μm diameter stainless steel electrodes (MS333/1-A, Plastics One) were bent into an “L” shape, and the deinsulated bottom of the “L” was placed over the dura of the hindlimb representative and the BDA injection area (1.5 mm apart). To confirm proper placement of the electrodes, we stimulated the M1 cortex to provoke a motor response in the contralateral hindlimb. A constant current stimulator (Neopraxis) was used to deliver trains of stimuli (190 msec on, 1500 msec off; 1.0–1.5 mA; 200 Hz). Once proper placement was confirmed, electrodes were secured with dental acrylic cement supported by three microscrews (Small Parts). The skin on the head was subsequently sutured.
Staring on day 28, animals were stimulated for 4 h daily for 3 weeks (Fig. 1). The connector on the head of the animals that contained the implanted electrode was attached to a commutator (Plastics One) mounted to the top of the cage, and trains of stimuli (described previously) were applied, to provoke a motor response limited to the hindlimb that was contralateral to stimulation. Animals were able to eat, drink, and walk during the stimulation. Animals did not paw at their heads or show signs of pain or distress. On the days when both behavior testing and stimulation were performed, behavior testing was done first. Sham-stimulated animals received electrode implantation only but were not stimulated.
BrdU administration
BrdU was dissolved in saline (0.1M) with a brief ultrasonic treatment at 37°C. During days 42–49, BrdU was administered through intraperitoneal (i.p.) injections at a 50 mg/kg concentration. The last injection was administered on day 49, 3 h before the animals were perfused (Fig. 1).
Tissue processing, epon embedding, and anatomical staining
Three animals were perfused with 2%-glutaraldehyde and 2%-paraformaldehyde in phosphate buffered saline (PBS) on day 49. Spinal cords were dissected, and 1 mm blocks at T10 (injury epicenter, the black segment in Fig. 2) from the naïve (Fig. 3A,B) and SCI only (Fig. 3 C,D) animals were cut, and post-fixed overnight in the same fixation solution. The blocks were exposed to 1% OsO4 for 1 h, stained in 0.5% uranyl acetate overnight, and dehydrated in a series of alcohols (15 min each) followed by propylene oxide for 3 h. After being infiltrated with a 1:1 mixture of propylene oxide and EMBed-812 embedding resin (Electron Microscopy Sciences) for 3 h, the blocks were embedded with same resin at 60°C overnight. Transverse semi-thin sections were cut on a microtome at 1 μm thickness and stained with 1% Toluidine blue. The semi-thin sections were analyzed with a light microscope (Zeiss ApoTome) (Fig. 3A–D).
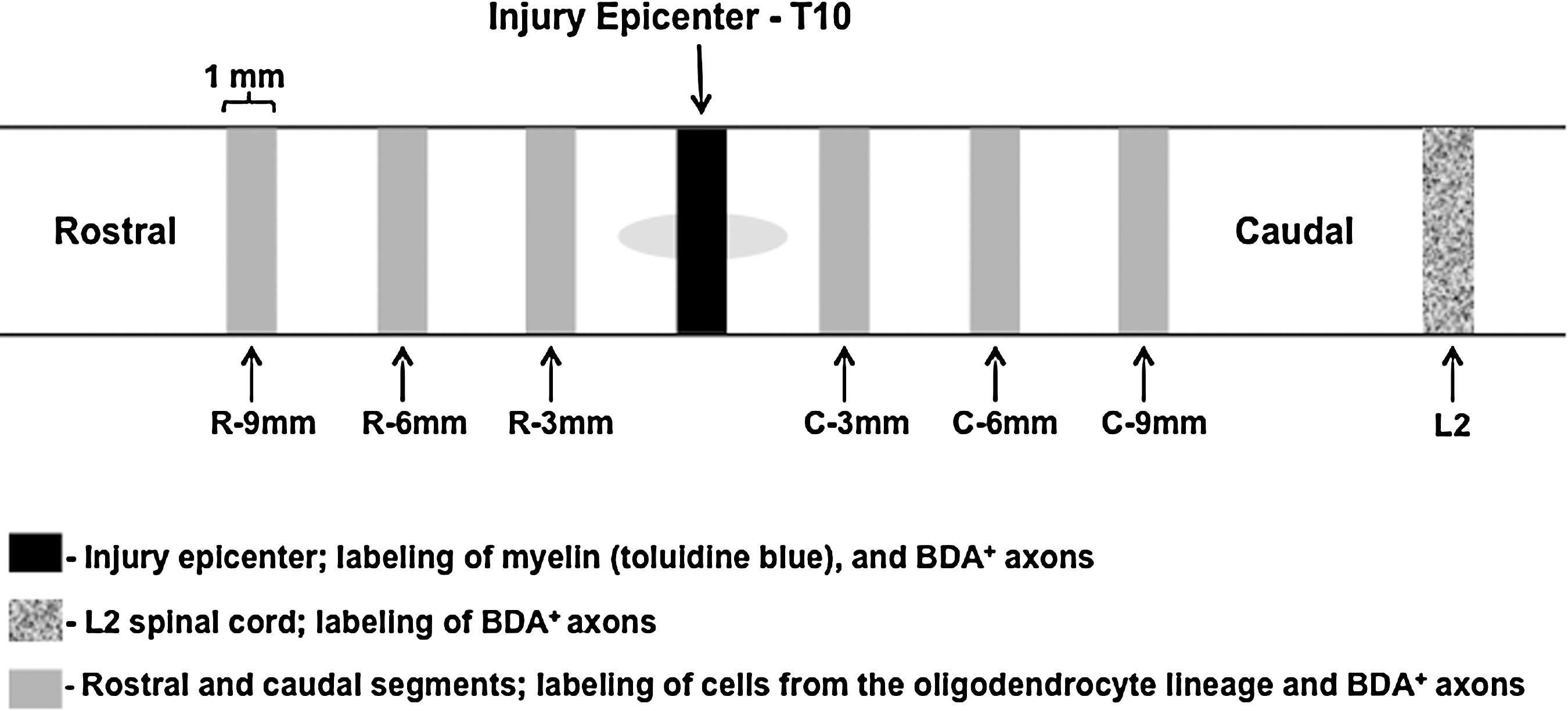
Schematic diagram of tissue sampling. After perfusion at day 49, spinal cords were dissected and cut into 1 mm segments for morphological analysis. In three animals (naïve; spinal cord injury [SCI] only; SCI and biotinylated dextran amine [BDA] injection), tissue from the epicenter of the lesion (T10) was processed for epon embedding and stained with Toluidine blue (black segment; for sample images see Fig. 3A–F). A 1 mm segment at L2 from the animal that received SCI and BDA injection was further cut into 40 μm sections and stained with Cy2-streptavidin to detect BDA-labeled dorsal corticospinal tract (dCST) axons (shadowed segment; for sample images see Fig. 3G). For the experimental animals (six stimulated and six sham animals), 1 mm segments located 3 mm, 6 mm, and 9 mm rostral and caudal from the lesion epicenter were further cut into 40 μm sections and immunostained (gray segments).
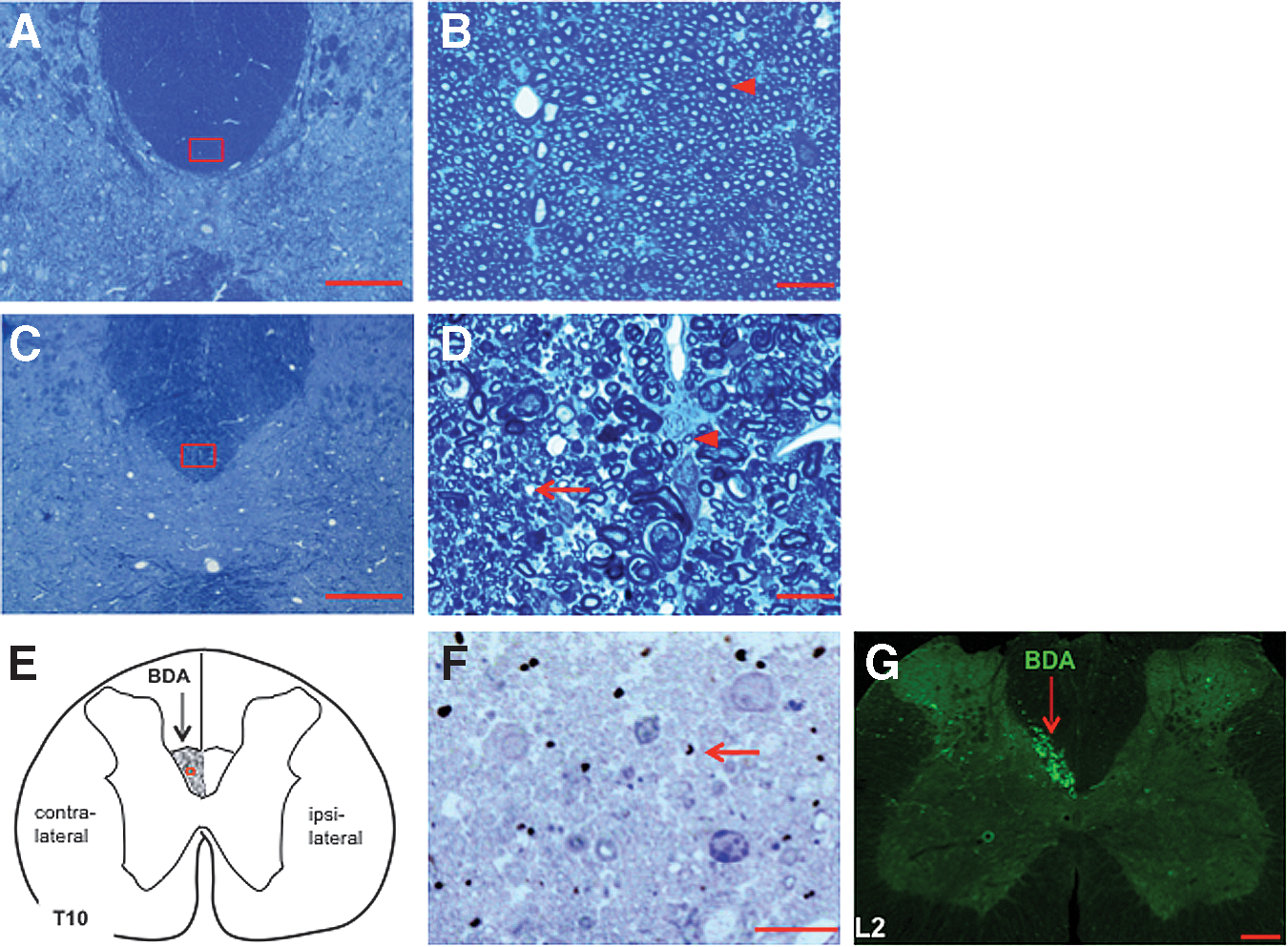
Mild contusion at T10 causes demyelination of surviving axons in the dorsal corticospinal tract (dCST) of injured spinal cord.
The animal that received both SCI and BDA injections (Fig. 3F, G) underwent the same procedures for perfusion, post-fixation, and dissection as described. However, here, the 1 mm block from the injury epicenter at T10 (Fig. 2) was first sectioned at 60 μm using a vibratome. Sections were incubated in 1% avidin-biotin complex (ABC) reagent (Vector Laboratories) for 1 h at room temperature. After rinsing twice in PBS, the tissue was incubated with the chromogen (diaminobenzidine [DAB]) for 6–10 min in: 1 DAB tablet (10 mg; Sigma) plus 0.01% H2O2 in 15 mL PBS. After the dehydration and osmium fixation procedures, sections were flat-embedded in EMBed-812 resin between two sheets of Aclar in 60°C overnight. The plastic sections were dissected, and the area of dCST was trimmed and affixed on blank beam capsules. Tissue was further micro-sectioned in 0.5 μm slices, and stained with toluidine blue (Fig. 3F). L2 spinal cord tissue caudal from the injury epicenter (shadowed segment in Fig. 2) from the same animal was freeze-sectioned at 40 μm, stained with Cy2-streptavidin, and imaged with an Olympus BX61 fluorescence microscope (Fig. 3G).
Immunohistochemistry (IHC)
Twelve experimental animals (six stimulated and six sham-stimulated) were used for IHC. On day 49, animals were perfused with 4% paraformaldehyde in 0.1M PBS, and spinal cords were dissected. For each animal we cut six blocks of 1 mm long spinal cord tissue; specifically, these blocks were cut 3 mm, 6 mm, and 9 mm rostral from the injury epicenter, and 3 mm, 6 mm, and 9 mm caudal from injury epicenter (gray segments in Fig. 2). Blocks were post-fixed at 4°C overnight, then transferred into 20% sucrose solution until the tissue sank to bottom of the vials. The spinal cords were freeze-cut transversely in 40 μm sections. For each spinal cord block (1 mm in length), ∼24–25 sections were collected and they were divided into four groups (usually six sections/group) in a rotating order. These four groups of tissue sections were immunostained for: (1) neural/glial antigen (NG2)/BrdU/BDA; (2) APC-CC1/BDA; (3) MBP/neurofilament (NF); and (4) NG2/synaptophysin/BDA. For BrdU staining, sections were pretreated with 2N HCl to denature DNA (37°C; 45 min), and with two 15 min borate buffer treatments (pH 8.5) to neutralize the HCl. Following blocking for 60 min in 10% normal goat serum (NGS) and 0.1% triton X-100 in PBS, sections were incubated in primary antibodies at 4°C for 48 h. Primary antibodies used in this study were: mouse anti-BrdU (1:600; Serotec), rabbit anti-NG2 (1:200; Chemicon), mouse anti-APC-CC1 (1:400; Oncogene), rabbit anti-MBP (1:1000; Millipore), mouse anti-NF (1:1000; Abcam), and mouse anti-synaptophysin (1:400; Chemicon). After three 10 min washes in PBS, sections were incubated with the following secondary antibodies: Alexa 488 conjugated goat anti-rabbit IgG (1:200; Life technologies) and Cy3 conjugated goat anti-mouse IgG (1:200; Jackson labs) or Alexa 488-goat anti-mouse IgG (1:200; Life technologies) at room temperature for 2 h. Cy3- or Cy5-conjugated streptavidin (1:1000: Jackson labs) was mixed with secondary antibodies to stain BDA-labeled CST axons. Sections were mounted onto slides, air-dried, cover-slipped, and observed with an Olympus BX61fluorescence microscope. 11
Cell counting
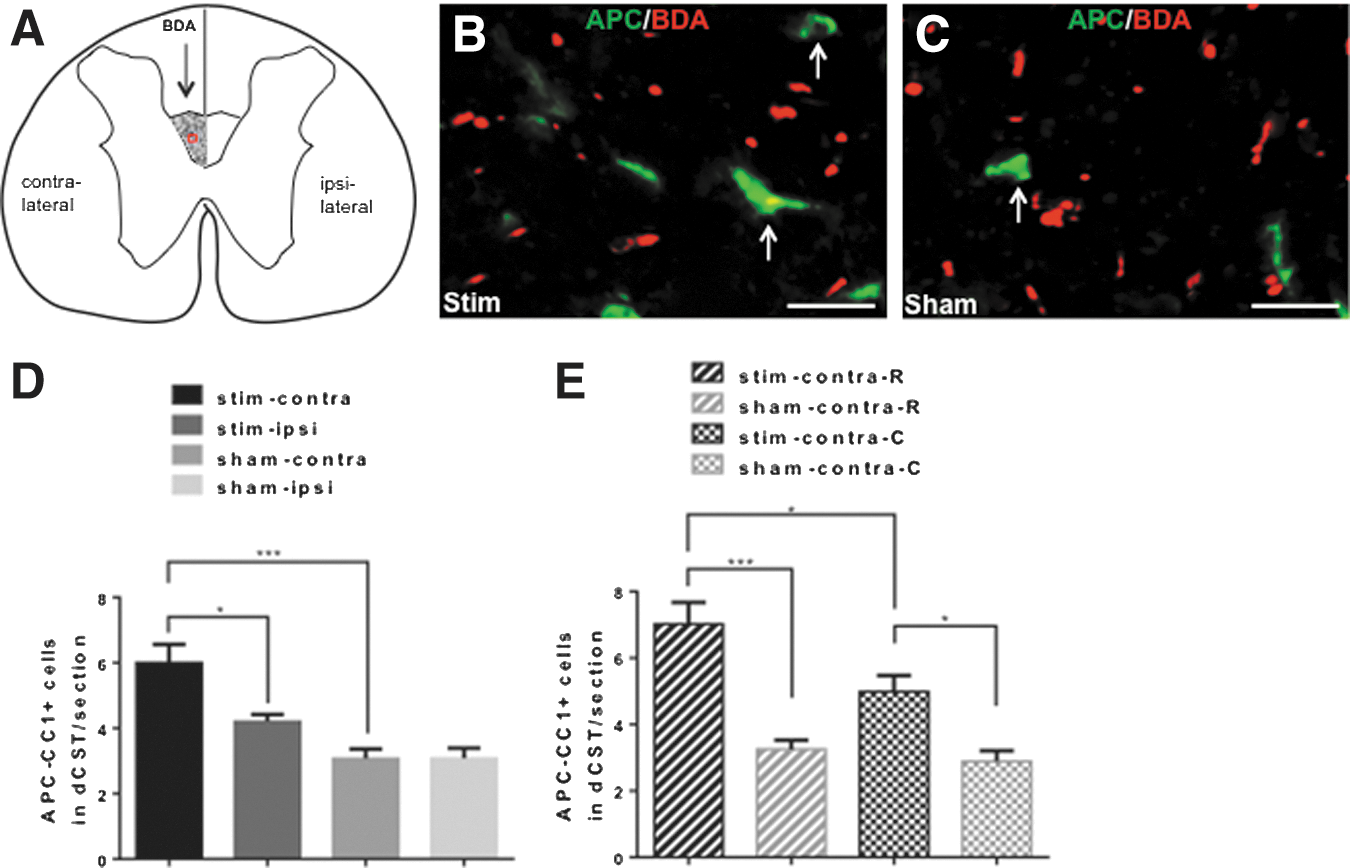
BrdU+/NG2+ and APC-CC1+ cells within the dCST area were counted using ImageJ. The criteria for counting proliferating OPCs were for a cell to have BrdU+ nuclei and NG2+ cytoplasm (Fig. 4B, C). Mature oligodendrocytes were counted for cells that had APC-CC1-labeled cell bodies (Fig. 5B,C). Images of the dCST area from all the immunostained transverse sections were taken at 40 × magnification on an Olympus BX61 fluorescence microscope. Images were opened and initialized in ImageJ. The dCST was outlined using the “Freehand Selection” tool. The contralateral dCST was identified by the presence of BDA-labeled axons, whereas the symmetric area across the midline was identified as the ipsilateral dCST (Figs. 4A and 5A). The “Cell Counter” tool was then used to count each immune-labeled cell, with which each counted cell is marked preventing the same cell from being counted twice. The program recorded the number of counted cells for each section automatically. In each 1 mm spinal cord segment the total number of labeled cells from six 40 μm sections were added and divided by 6 (the number of sections per each segment). In Figure 4, because of the low number of proliferating OPCs within the dCST in one section, we averaged the number of BrdU+/NG2+ cells from all six 1 mm spinal cord segments. The average number of BrdU+/NG2+ cells within dCST per 40 μm section ± SEM was compared for four groups as described under Data analysis and statistics (Fig. 4D). In Figure 5, because the number of labeled oligodendrocytes was higher, in addition to comparing the average number of labeled oligodendrocytes in all six, 1 mm spinal cord segments for four experimental groups (Fig. 5D), we compared the average number of labeled oligodendrocytes within the contralateral dCST per 40 μm section ± SEM in the rostral (R-3mm, R-6mm, and R-9mm) versus caudal (C-3mm, C-6mm, and C-9mm) segments of the spinal cords. (Fig. 5E).
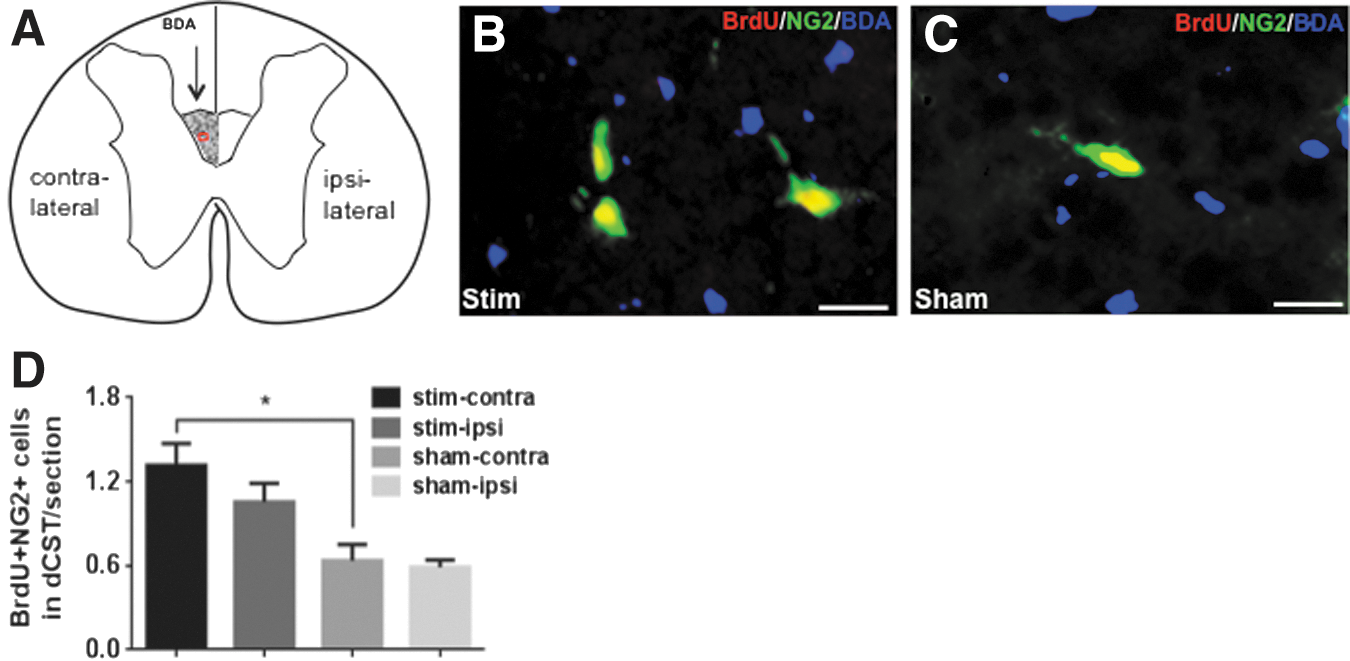
Induced neuronal activity elicits oligodendrocyte progenitor cells' (OPCs') mitogenic response in the dorsal corticospinal tract (dCST) in subchronic spinal cord injury (SCI).
Induced neuronal activation increases oligodendrogenesis in the dorsal corticospinal tract (dCST) in subchronic spinal cord injury (SCI).
Western blotting
Twelve animals (n = 6 per group) were quickly perfused with cold saline on day 49. A 10 mm long spinal cord section centered at the injury epicenter (R-5-mm to C-5-mm) was removed, and put on dry ice. Spinal cord tissue was separated along the midline into the ipsilateral half and the contralateral half. Each half was cut into the ventral and dorsal part. The dorsal spinal cord tissue was then lysed in RIPA buffer (Sigma). Samples were prepared in denaturing sample buffer (Bio-Rad), run on 4–15% Tris-HCl SDS polyacrylamide gels (Bio-Rad), and transferred to PVDF-membranes (Bio-Rad). Blots were probed with anti-MBP (Oncogene) and anti-β-actin antibodies. Blots were visualized using SuperSignal chemiluminescence kits (Pierce Biotechnology), and images were acquired and quantitated using FluorChem Q (AlphaInnotech) (Fig. 6A).
Induced neuronal activation increases remyelination in the dorsal corticospinal tract (dCST) in subchronic spinal cord injury (SCI).
Quantitative image analysis
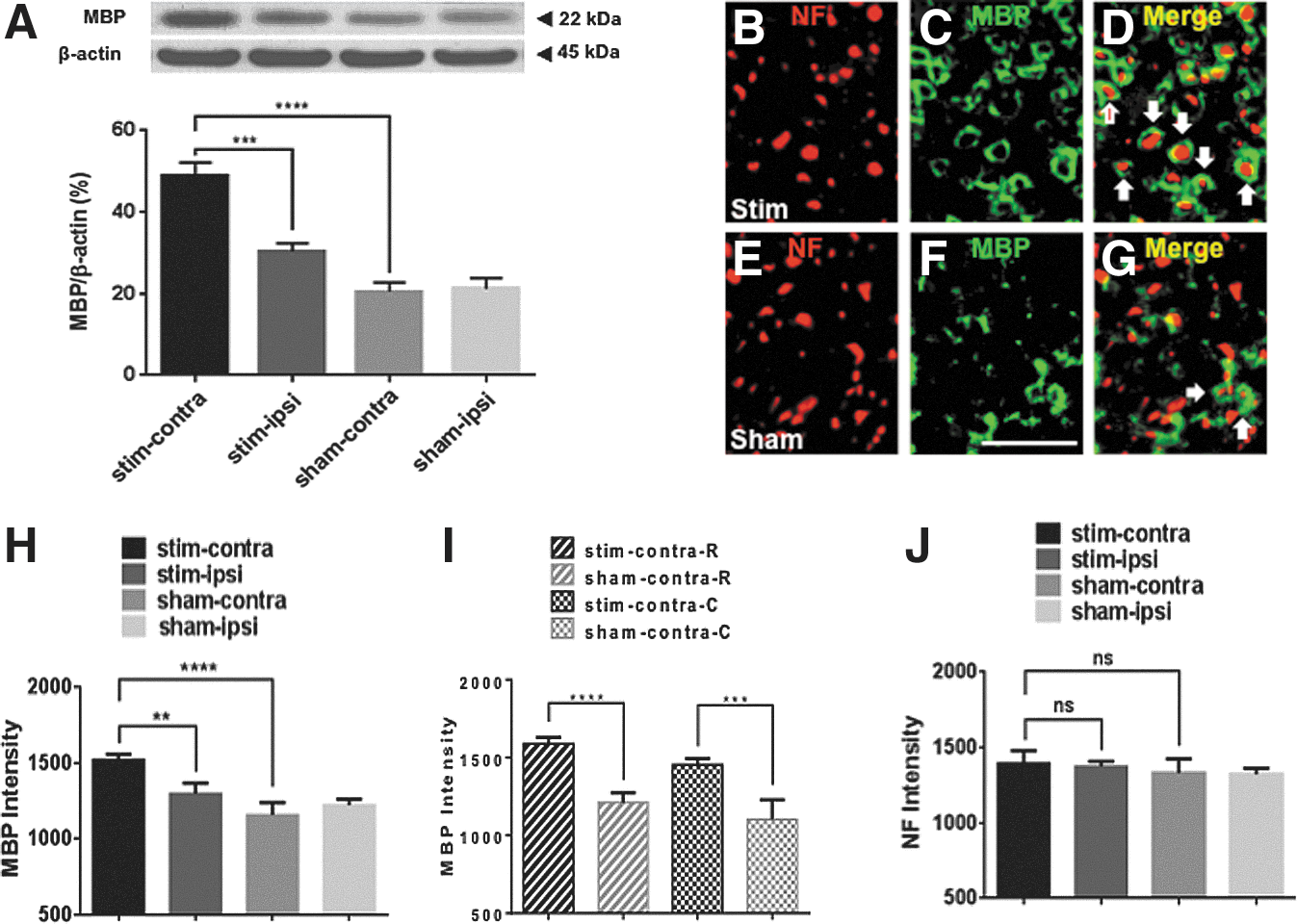
Mean fluorescence intensities for NF+ axons and MBP+ myelin sheaths within the dCST were obtained by using Slide Book 5.0 (Olympus) (Fig. 6B–J). Images of the dCST area from NF- and MBP-immunostained transverse sections were also taken at 40 × magnification on an Olympus BX61 fluorescence microscope. For quantification, images were opened in Slidebook 5.0. The mean fluorescence intensities from a region of interest (ROI) that covered the dCST were obtained for each channel separately using the Intensity Statistics function within the Statistics menu. In Figure 6, mean fluorescence intensities are reported per section as well; therefore, the same averaging across rostral and caudal regions as reported for cell counting of oligodendrocytes was repeated here (Fig. 6H–J).
Confocal microscopy and axon-OPC synapse counting
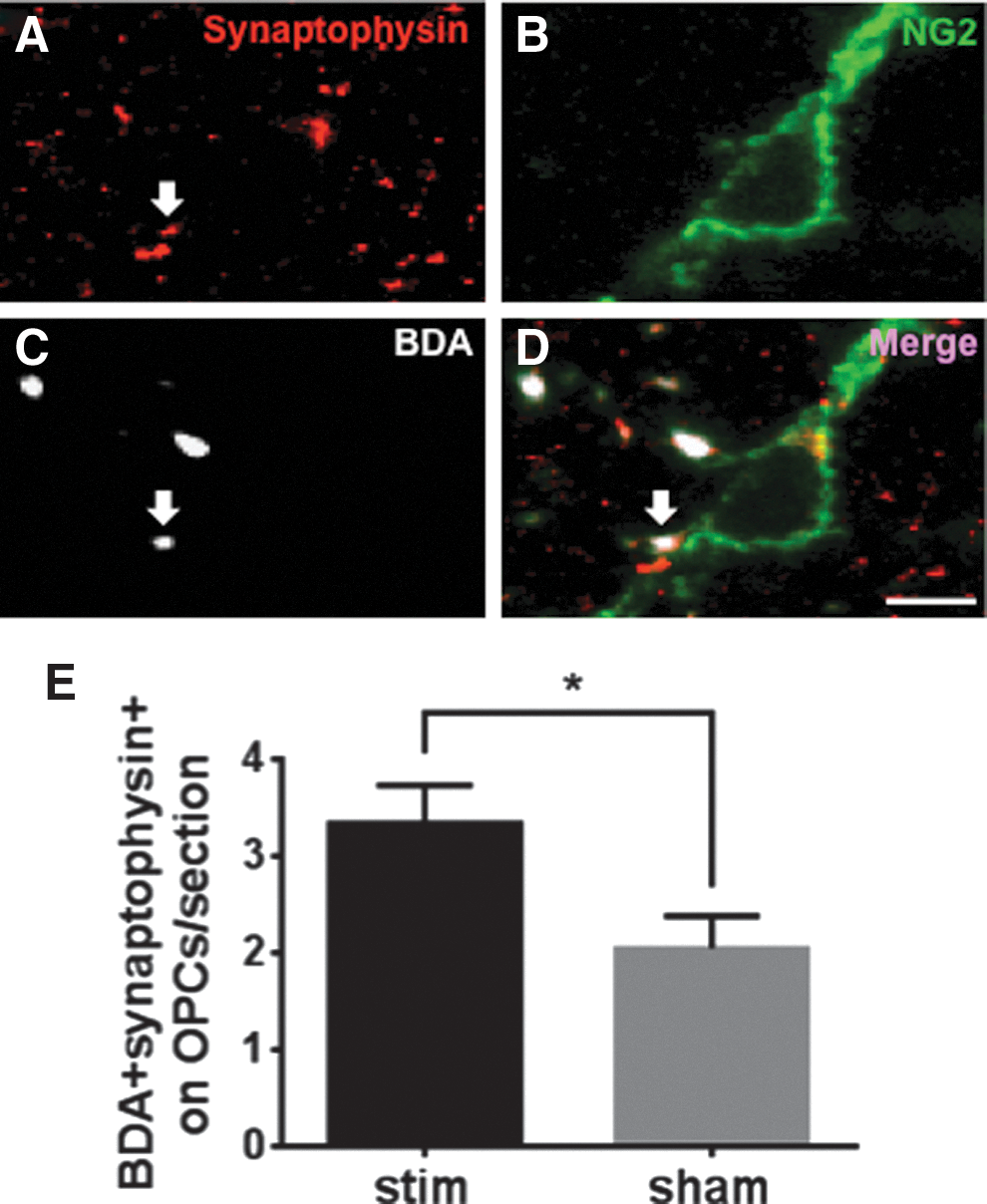
Confocal microscopy (Olympus FluoView1000) was used to image axon-OPC synapses within the dCST. The criterion for axon-OPC synapses were that within a 1 μm optical image, BDA+ dCST axons had to colocalize with synaptophysin, and this double-labeled component had to be adjacent to the cell body or proximal process of a NG2+ OPC (Fig. 7A–D). Axon-OPC synapses were counted in the contralateral side of stimulated and sham animals (Fig. 7E). The counting for synapses from 1 μm optical images of the dCST was done using ImageJ as described.
Induced neuronal activity promotes formation of axon-oligodendrocyte progenitor cell (OPC) synapses in the contralateral dorsal corticospinal tract (dCST) in subchronic spinal cord injury (SCI). (
Hindlimb motor function assay
Grid-walk assay was performed weekly from day 28 to day 49. Animals were pre-trained for 3 days before the first testing day. Animals were required to run two round trips across a 120 cm horizontal ladder beam that had 40 irregularly spaced (1–3 mm) metal bars (0.8 cm diameter). All hindlimb steps were videotaped, and analyzed in slow motion. The criteria for a correct step were that all digits held the bar and that the animal placed the palm of the paw directly onto the bar. Steps with slips and misplacements were incorrect steps. The correct-step rate was calculated as number of correct steps over number of total steps. The data from all time points were normalized to the pre-stimulation (day 28) correct-step rate. Therefore, the normalized correct-step rate was calculated as the correct-step rate at a particular time point over the correct-step rate on day 28 times 100% (Fig. 8).
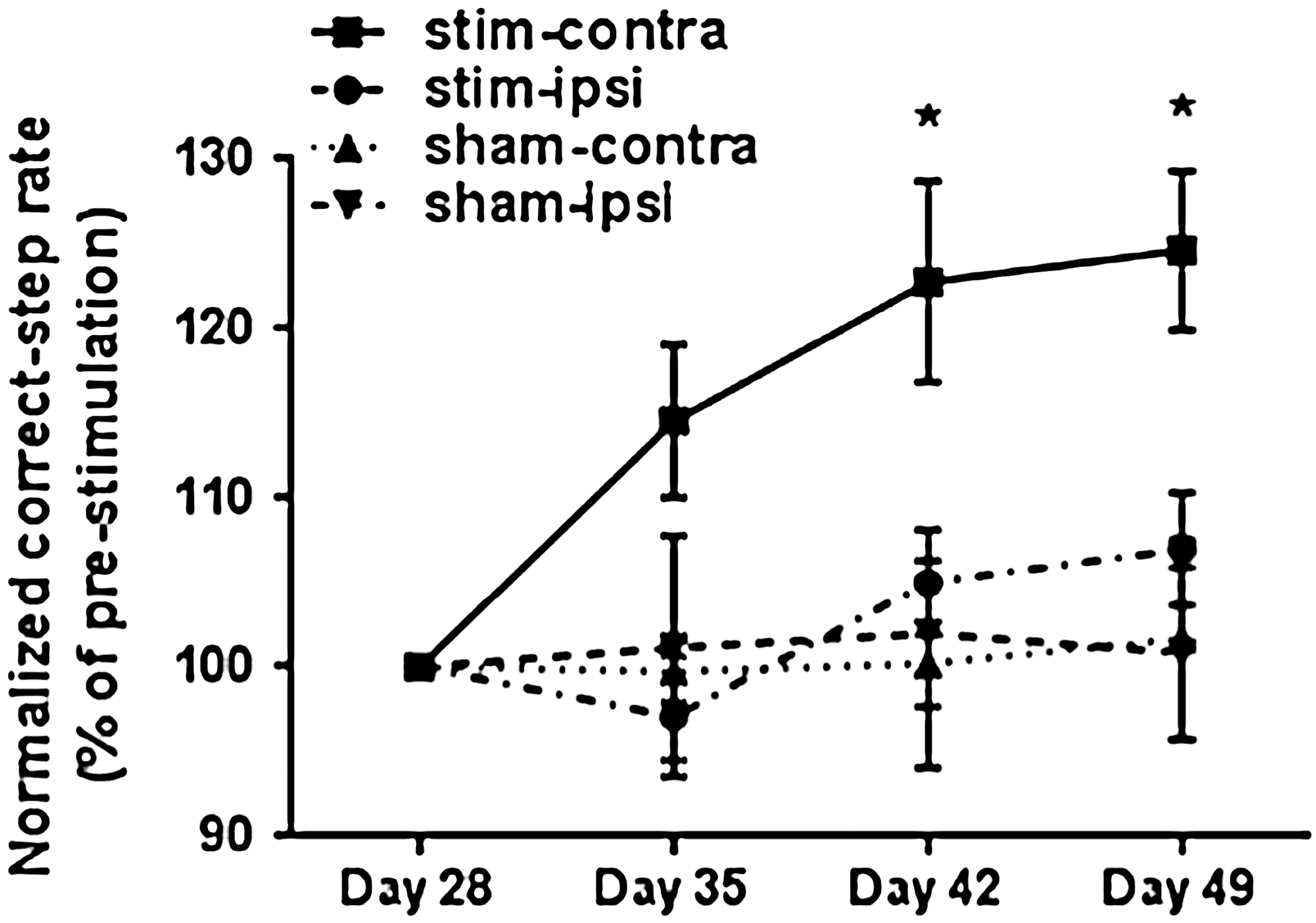
Induction of neuronal activation through the M1 cortex in subchronic spinal cord injury (SCI) improves hindlimb motor function. Hindlimb motor function was assessed weekly after induction of neuronal activation using a grid-walk assay. The normalized correct-step rate represents the correct-step rate (number of correct steps over number of total steps) from all time points normalized to the pre-stimulation rate (day 28). Note that the normalized correct-step rate in the stim-contralateral hindlimb was significantly higher on days 42 and 49 than that in the stim-ipsilateral hindlimb and both hindlimbs of sham-stimulated animals. Two way analysis of variance (ANOVA) was conducted with significance indicated as *p < 0.05; errors bars indicate SEM.
Statistical analysis
Data were analyzed from the following groups: (1) stim-contralateral (stim-contra), contralateral to the implanted electrode in stimulated animals; (2) stim-ipsilateral (stim-ipsi), ipsilateral to the implanted electrode in stimulated animals; (3) sham-contralateral (sham-contra), contralateral to the implanted electrode in sham-stimulated animals, and (4) sham-ipsilateral (sham-ipsi), ipsilateral to the implanted electrode in sham-stimulated animals. Comparisons were made between stim-contra and sham-contra, or between stim-contra and stim-ipsi. IHC and Western blot data were analyzed using one way analysis of variance (ANOVA); grid-walk assay data were analyzed with two-way ANOVA. The t test was used for comparison of axon-OPC synapses.
Results
Mild SCI causes demyelination within the dCST at the injury epicenter, but preserves dCST axons
We aimed to determine if induced neuronal activation affected oligodendrogenesis in the spinal cord after SCI. However, current rodent models of SCI induce loss of oligodendrocytes and axons and cause a contusive cyst at the injury epicenter. Therefore, for this study, we utilized a mild model of SCI that was inflicted by a 40 kdyn impact at T10. In the subchronic stage (49 days after injury; Fig. 1), the mild contusive SCI did not create a contusive cyst at the injury epicenter, and there was no scar tissue surrounding the injured tissue at the epicenter of the lesion (Fig. 3A and C). This mild contusive SCI did result in partial demyelination of dCST axons at the lesion epicenter (Fig. 3B and D). To assess axonal preservation of the dCST axons that project to the contralateral spinal cord, on day 28 we injected BDA unilaterally into the M1 cortex. Three weeks after injection, surviving BDA-labeled axons within the dCST were seen at the epicenter of the SCI (Fig. 3F). Despite demyelination within the dCST at the SCI epicenter, BDA-labeled axons survived caudal (L2) from the T10 injury (Figs. 2 and 3G). These data established a mild thoracic contusive SCI as a good model to study potential effects of induced neuronal activity on oligodendrogenesis and remyelination in the injured spinal cord.
M1-induced neuronal activity promotes proliferation of OPCs, oligodendrogenesis, and remyelination in subchronic SCI
Toward the aim of determining if induced neuronal activation affected oligodendrogenesis after SCI, we induced neuronal activation through electrical stimulation of the M1 cortex in a subchronic model of mild contusive SCI at T10 (experimental timeline is described in Fig. 1). Four weeks after SCI, we implanted unilateral electrodes over the M1 cortex and induced neuronal activation through electrical stimulation for a period of 3 weeks. In the last week of stimulation, BrdU was injected to label proliferating cells (Fig. 1). All animals were euthanized on day 49, and effects on oligodendrogenesis were analyzed in spinal cord transverse sections that were cut 3 mm, 6 mm, and 9 mm rostral and caudal from the lesion epicenter (Fig. 2). First, we examined proliferation of OPCs in the dCST after stimulation by counting BrdU+/NG2+ cells (proliferating OPCs) using ImageJ (Fig. 4). The contralateral dCST was identified by the presence of BDA-labeled axons, and the symmetric area across the midline represented the ipsilateral dCST (Fig. 4A). We counted 0–4 proliferating OPCs within the dCST in each 40 μm section (Fig. 4B,C); therefore, the number of proliferating OPCs from all the sections was averaged. The number of proliferating OPCs was higher in stim-contra dCST than in sham-contra dCST (p < 0.05). In stim-ipsi dCST, the trend for proliferating OPCs was higher than in sham-ipsi dCST, and lower than in stim-contra; however, these differences did not reach statistical significance (Fig. 4D). These data show that in subchronic SCI, induced neuronal activity promotes proliferation of OPCs.
Next, we wanted to determine if the increased proliferation of OPCs that was generated by induced neuronal activity leads to a higher number of oligodendrocytes (APC-CC1+ cells; Fig. 5). Cell counting was also performed with ImageJ in dCST from the transverse section. Using Image J, we counted 1–12 oligodendrocytes within the dCST in each 40 μm section (representative images in Fig. 5B, C). The number of oligodendrocytes in the stim-contra dCST was higher than in sham-contra (p < 0.001) and stim-ipsi (p < 0.05) dCST, and there was no difference between sham-contra and sham-ipsi dCST (Fig. 5D). Within the stim-contra dCST, the number of oligodendrocytes in segments rostral from the SCI epicenter was higher than in caudal segments (p < 0.05). The number of oligodendrocytes in rostral stim-contra dCST was also higher than that in rostral sham-contra dCST (p < 0.001). However, this affect of neuronal activity was not restricted to the rostral region, as the number of oligodendrocytes in caudal stim-contra dCST was higher than that in caudal sham-contra dCST (p < 0.05). In sham animals, the number of oligodendrocytes did not differ between the rostral and caudal dCST (Fig. 5E). These data show that in subchronic SCI, induced neuronal activity promotes oligodendrogenesis.
To determine if the increase in oligodendrocytes resulted in remyelination of demyelinated dCST axons, we first assessed total MBP protein levels in the injured dorsal spinal cord following induced neuronal activation using Western blotting. MBP protein levels were higher in the stim-contra dorsal spinal cord than in the same region in stim-ipsi (p < 0.001) and sham-contra dorsal spinal cord (p < 0.0001). MBP protein levels did not differ between the dorsal sham-contra and sham-ipsi (Fig. 6A). To further examine the status of myelination within the dCST following induced neuronal activation, we assessed MBP+ sheath formation by immunofluorescence (representative images are presented in Fig. 6C, D, F, and G). 13 MBP+ sheath formation was increased in stim-contra dCST compared with sham-contra dCST (p < 0.0001) and stim-ipsi dCST (p < 0.01), and it did not differ between the sham-contra and sham-ipsi dCST (Fig. 6H). Both rostral and caudal segments of stim-contra dCST had more MBP+ sheaths than the rostral (p < 0.0001) and caudal (p < 0.001) dCST, respectively, of sham-stimulated animals. However, within the stim-dCST there was no difference in MBP+ sheath formation in rostral and caudal regions (Fig. 6I). Therefore, MBP+ sheath formation corresponds to the number of oligodendrocytes as long as a critical number of oligodendrocytes is reached. The possibility that the increase in MBP+ sheaths in response to electrical stimulation of M1 cortex occurs as a result of a higher loss of axons in sham-stimulated animals or because of myelination of regenerated axons of the dCST remains, considering that this form of stimulation has shown to induce axonal regeneration. 14 –16 To account for these possibilities, we assessed the number of NF+ axons by immunofluorescence (representative images are presented in Fig. 6B, E, D, and G). 13 However, we found that the fluorescent intensity of NF+ axons within the dCST did not differ in the four experimental groups; therefore, differences in the number of axons are not responsible for the observed increase in remyelination. Taken together, these data indicate that in subchronic SCI, induced neuronal activity enhances remyelination of surviving axons.
M1-induced neuronal activity increases the number of axon-OPC synapses in subchronic SCI
Having established that induced neuronal activity promotes proliferation of OPCs, oligodendrogenesis, and remyelination in sub-chronically injured spinal cord, we aimed to determine a mechanism through which these effects are mediated. In EB-induced demyelination, endogenous neuronal activity within demyelinated axons promotes formation of generate de novo synapses with OPCs, which go on to remyelinate. 6 Therefore, we examined if the number of axon-OPC synapses in the injured spinal cord changed in response to induced neuronal activity. Thus, we compared the number of axon-OPC synaptic interactions from stimulated animals versus sham-stimulated animals where we defined such synapses as direct contact between BDA+/synaptophysin+ axonal terminals and NG2+ OPCs (representative images are presented in Fig. 7A–D). We found that the number of axon-OPC synapses in stim-contra dCST was higher than in sham-contra dCST (p < 0.05; Fig. 7E). These data show that in subchronic SCI, induced neuronal activity increases the number of axon-OPC synapses, indicating that the oligodendrocyte mitogenic response could be mediated through excitatory axon-OPC synapses.
M1 neuronal activity promotes recovery of motor function
Finally, we determined if neuronal activity induced through electrical stimulation of M1 cortex generated recovery of fine hindlimb motor function. Using a grid-walk assay, we compared the normalized correct-step rate (correct steps/total steps normalized to the pre-stimulation rate) of the contralateral and ipsilateral hindpaw in stimulated and sham-stimulated animals. This normalization step was done to account for the variability that could be introduced as a result of the difference in the SCI between each animal. We found that normalized correct-step rate for stim-contra hindlimbs significantly increased 2 weeks after stimulation (day 42) as compared with the stim-ipsi (p < 0.05), sham-contra (p < 0.05), and sham-ipsi (p < 0.05) hindlimbs. The increase persisted until the experimental end-point (day 49) (p < 0.05) (Fig. 8). These data demonstrate that in subchronic SCI, neuronal activity promotes recovery of motor function.
Discussion
This study demonstrates that neuronal activation induced through electrical stimulation of the M1 motor cortex elicits proliferation of OPCs in stimulated white matter tracts of the injured spinal cord, which in turn generates a higher number of oligodendrocytes and enhances remyelination within these tracts. Neuronal activation also increased the number of axon-OPC synapses in stimulated white matter tracts of the injured spinal cord. This increase in axon-OPC synapses indicates that such synapses could mediate these effects. The abovementioned effects in cells of the oligodendrocyte lineage and their interactions with axons are associated with an improvement in motor function. Collectively, our findings confirm that activity-dependent myelination is a mechanism of nervous system plasticity in the injured spinal cord. In addition, our work is proof of concept that neuronal activity induced through cortical stimulation is a valid therapeutic strategy for treatment of SCI and other demyelination disorders.
Evidence of continued remyelination after SCI has cast doubt on whether remyelination is a valid therapeutic target in treatment of SCI. 8,17 However, several lines of evidence confirm that remyelination is a feasible target for inducing neurological recovery after SCI. First, 3 months after SCI, evidence of demyelination and dysmyelination remain in the injured spinal cord both at the epicenter and distal from the injury. 8 Demyelination (lack of myelin) or dysmyelination (abnormal myelin) reduces efficacy of axonal signal transduction, and leads to axonal degeneration. Loss of oligodendrocytes is noted within the 1st h after SCI, 18 and persists for at least 3 weeks. 19,20 However, SCI also causes an oligodendroglia regenerative response, which consists of extensive proliferation of OPCs, 8,21 –24 and their differentiation into oligodendrocytes. 8,18,21,22 Partial demyelination and dysmyelination that persists 8 weeks after SCI is estimated to cause a 21% decrease in conduction velocity through remyelinated axons, 25 and such alterations in conduction velocities can lead to paralysis. Second, minimal sparing of myelinated axons is sufficient to restore locomotion. Survival of 5–10% of myelinated white matter axons concentrated towards the pial surface is sufficient to restore walking in cats after SCI. 26 Finally, interventions that enhance remyelination improve neurological function after SCI. Transplanted shiverer-derived neural progenitor cells (NPCs) survive in the injured spinal cord; generate neurons, astrocytes and oligodendrocytes; promote axonal growth; and prevent atrophy at an equal rate to that of wild type NPCs. However, the inability of shiverer-derived NPCs to form compact myelin because of a deletion of the myelin basic protein (MBP) gene renders them incapable of restoring motor function after transplantation in the injured spinal cord, whereas wildtype NSCs that form compact myelin are able to restore motor function. 27
Myelination of axons in the CNS is necessary for integrated motor, sensory, and cognitive functions; this is a dynamic process that is regulated by endogenous neuronal activity. Activity-dependent myelination is emerging as a mechanism of CNS plasticity. 2 Experimentation in zebrafish highlights subtle differences in mechanisms through which endogenous neuronal activity regulates myelination in the spinal cord. 28,29 One study finds that neuronal activity-dependent synaptic release regulates myelination first by altering the number of OPCs, where induction of neuronal activity increases the number of OPCs whereas inhibition decreases the number of OPCs, and second, by increasing the initial formation of myelin sheaths, where myelin sheaths are defined as being >5 μm in length. 29 Another study, where the myelin sheath is not defined by size, finds that neuronal activity-dependent secretion regulates myelination by extending and stabilizing myelin sheaths formed on select axons rather then affecting the initial step of myelin sheath formation. 28 Interestingly, myelin sheaths form on inactive axons, but they are shorter in length; when activity of neighboring axons is suppressed, inhibition of sheath growth is relieved. 28 This suggests that desynchronized activity in axons (a characteristic of axonal tracts after SCI) can lead to dysmyelination, insufficient or structurally altered myelin formation.
Effects of induced neuronal activity on oligodendrogenesis and myelin remodeling are similar to the effects of endogenous neuronal activity, that is induction of neuronal activity through cortical stimulation promotes oligodendrogenesis and myelin remodeling in subcortical white matter tracks of the brain, 12 and in the uninjured spinal cord. 11 Our work illustrates that the effects of induced neuronal activation persist in the injured spinal cord, where we first noted an increase in OPC proliferation that surpassed the endogenous OPC proliferative response after SCI. 8 Interestingly, we observed a trend of increased proliferation of OPCs in the ipsilateral dCST, which was not stimulated. Such an observation can be explained by the introduction of activity through sprouting of CST tracts. The mild SCI model used in this study is designed to minimize axonal damage; however, some axonal damage can occur. Axonal outgrowth from damaged contralateral dCST axons, promoted by cortical stimulation, could be responsible for stimulation of OPC proliferation in the ipsilateral dCST, albeit at a lower rate. Concomitantly to the increase in proliferating OPCs, induced neuronal activity increases the number of oligodendrocytes and promotes remyelination within the injured spinal cord. The parallel increase in OPCs and oligodendrocytes described here and as previously reported 11,12 is a subtle differences compared with endogenous neuronal activity, which depletes the number of OPCs as the number of oligodendrocytes increases. 6
Mechanisms through which neuronal activity modulates oligodendrogenesis and myelination are under investigation. In our SCI model, induced neuronal activation increases axon-OPC synapses in the dCST, a white matter tract that is devoid of neuronal spines, dendrites, and somata. Induced neuronal activation has been reported to promote glutamate release in white matter tracts.
30
Axon-OPC synapses, which exhibit similar plasticity as neuronal synapses,
31
could mediate the effects of glutamate in OPCs. In EB-induced demyelination lesions, AMPA receptors in OPCs mediate effects of neuronal activity exerted through glutamate on OPC proliferation, differentiation, and remyelination.
6
Further work is needed to ascertain the role of axon-OPC synapses in activity-dependent regulation of oligodendrogenesis and remyelination, as these could be mediated by other mechanisms. For example, vesicular release at non-synaptic axon-OPC junctions can also induces myelination,
32
potentially through sodium-dependent glutamate transporters.
33
Additionally, in vitro AMPA receptors do not enhance myelin formation even though they upregulate differentiation of OPCs into oligodendrocytes in response to endogenous neuronal activity.
34
Further, in the presence of neuregulin and brain-derived neurotrophic factor (BDNF), it is N-methyl-
Induced neuronal activity modulates motor function in uninjured animals, and pharmacological agents that block oligodendrogenesis inhibit such modulation. 12 Additionally, motor skill learning is hindered when OPC differentiation into oligodendrocytes is blocked. 4 In accordance with these reports, our work demonstrates that induced neuronal activation improves motor function in subchronic SCI. Specifically, we demonstrate that M1 induction of neuronal activity improves fine hindpaw motor function exclusively in the treated limb. Hindpaw motor function was assessed while the animals were not being stimulated, affirming that this mode of clinically applicable stimulation is therapeutic rather than neuroprosthetic.
In addition to improvements in oligodendrogenesis and remyelination, the stimulation paradigm described in the current study was previously shown to promote recovery of motor function after unilateral pyramidotomy that coincides with augmented sprouting of dCST. 15,16,36 However, regeneration of the CST axons alone might not be sufficient for recovery of motor function. Optogenetic activation of the CST terminals shows that SOX11-expressing regenerated CST axons form functional synapses in the newly innervated gray matter of the spinal cord, yet fail to improve motor function. 37 It is possible that the regenerated axons do not form a sufficient number of synapses to generate a functional output, or that the newly formed synapses are not properly targeted. However, a recent study in the visual system introduced the possibility that regenerated axons need to be myelinated to generate functional recovery. 38
Conclusion
In conclusion, our work illustrates that epidural cortical stimulation is a valid therapeutic approach for treatment of patients with spinal cord injury. Cortical stimulation results in neuronal activation that induces axonal regeneration. In addition, our work has shown that neuronal activation also induces an oligodendroglial regenerative effect. The mechanism through which this occurs remains to be delineated; however, we show that this could occur through axon-OPC synapses. Other studies have shown that enhancement of remyelination or myelin remodeling through induced neuronal activation is a feasible therapeutic strategy in treatment of demyelination disorders such multiple sclerosis or enhancement of cognitive and motor functions. 4,6,12
Footnotes
Acknowledgments
We thank Michelle D. Harran for assistance in preparing the figures and the manuscript. This work was supported by the Office of the Assistant Secretary of Defense for Health Affairs under Award No. W81XWH-14-1-0069. Opinions, interpretations, conclusions, and recommendations are those of the author and are not necessarily endorsed by the Department of Defense.
Author Disclosure Statement
No competing financial interests exist.