
Abstract
Unilateral high cervical spinal hemisection (i.e., C2Hx) interrupts the respiratory bulbospinal pathway and results in paralysis of the hemidiaphragm. The ipsilateral diaphragmatic activity can partially recover over weeks to months; however, its contribution to the tidal volume generation is less than 20%. Accordingly, we hypothesized that the contralateral diaphragm exerts a compensatory function to maintain the essential ventilation following C2Hx. The cardiorespiratory pattern and bilateral diaphragm electromyogram (EMG) signals were measured in urethane-anesthetized and spontaneously breathing adult rats at 1 day, and 2 or 8 weeks post-C2Hx or C2 laminectomy. The functional contribution of the diaphragm was assessed by measuring immediate changes of the tidal volume following phrenic nerve section. At 1 day post-injury, the tidal volume was significantly reduced after contralateral phrenicotomy in C2Hx animals (54 ± 3% decline) compared with uninjured controls (20 ± 2% decline). Moreover, the arterial carbon dioxide partial pressure was significantly elevated in C2Hx animals (from 76 ± 8 mmHg to 117 ± 5 mmHg) but not in uninjured animals (from 51 ± 4 mmHg to 55 ± 3 mmHg). By 2 and 8 weeks post-injury, contralateral phrenicotomy still caused a greater reduction in the tidal volume in C2Hx than in uninjured animals, and the percentage decline of the tidal volume was similar to the response at 1 day post-injury. These data suggested that unilateral cervical spinal cord injury induced a persistent compensatory plasticity in the contralateral diaphragm, which plays a critical role in maintenance of essential ventilation.
Introduction
H
These results suggested that compensatory response of the contralateral diaphragm plays a predominant role in maintenance of the tidal volume following cervical spinal cord injury, and functional improvement of the respiratory function was also dependent on neuroplasticity of the contralateral diaphragm. However, there is no study directly assessing the functional role of the contralateral diaphragm following cervical spinal cord injury. Accordingly, the present study was designed to quantitatively measure the contribution of the contralateral diaphragm to the tidal volume by examining the immediate changes of the tidal volume following contralateral phrenicotomy at different time-points post-injury.
Methods
Animals
All experimental procedures were approved by the Institutional Animal Care and Use Committee at National Sun Yat-sen University. Adult (7–8 weeks of age) male Sprague-Dawley rats (purchased from BioLasco Taiwan Co., Ltd.) were divided into the uninjured (C2 laminectomy only, n = 24) and the C2Hx (n = 24) group, and subgrouped into three time-points post-injury (1 day post-injury [uninjured, n = 8; C2Hx, n = 8], 2 weeks post-injury [uninjured, n = 8; C2Hx, n = 8] and 8 weeks post-injury [uninjured, n = 8; C2Hx, n = 8)]).
Cervical spinal cord hemisection
Cervical spinal cord hemisection surgery was performed as previously described at 9–10 weeks of age. 4,13,14 Briefly, all animals were anesthetized with xylazine (10 mg/kg, s.c., Rompun®, Bayer) and ketamine (140 mg/kg, i.p., Ketalar®, Pfizer) and received a C2 laminectomy following an incision of dorsal skin and muscles. A unilateral hemilesion cavity was created at the left second cervical spinal cord by incision of the microscalpel and aspiration using a micropipette connected to a suction pump (TOMIN) in the C2Hx group. The dura, overlying muscles, and skin were sutured with 10-0 nylon (UNIK), 4-0 chromic (UNIK), and 4-0 nylon sutures (UNIK), respectively. The rat received subcutaneous injections of yohimbine (1.2 mg/kg, Tocris) to reverse the effect of xylazine, lactated Ringer's solution (5 mL, Nang Kuang Pharmaceutical Co., Ltd.) to prevent dehydration. After the animal awoke, a subcutaneous injection of buprenorphine (0.03 mg/kg, Shinlin Sinseng Pharmaceutical Co., Ltd.) was provided for analgesia. An oral supply of Nutri-cal (1–3 mL, EVSCO Pharmaceuticals) and lactated Ringer's solution injection (5 mL, s.c.) were applied daily until recovery of adequate eating and drinking.
Cardiorespiratory patterns and diaphragm EMG activity
Animals were anesthetized with urethane (1.6 g/kg, i.p., Sigma) at 1 day, and 2 or 8 weeks post-injury and placed in a supine position. The rectal temperature was monitored by a temperature probe and maintained at 37 ± 1°C by a temperature controller (model TC-1000, CWE Inc.). The depth of anesthesia was examined by pinching the toe after the initial anesthesia and surgical procedure; the surgical procedure would be executed until absence of toe-pinch reflex and alteration of breathing and/or blood pressure. A tracheotomy was performed at the cervical region and an endotracheal tube (PE-240, Clay Adams) was inserted into the trachea below the larynx. A hyperoxic gas (50% O2, 50% N2; flow rate: 2 L/min) was provided to the animal via a T-piece connector with the endotracheal tube, which was connected with a flow head (MLT1L, AD Instruments) coupled with a spirometer (FE141, AD Instruments) for respiratory flow measurement. Additional PE-50 catheters (Clay Adams) were inserted into the femoral artery and vein for blood pressure measurement (Transducer: DTX-1; Amplifier: TA-100, CWE Inc.) and drug administration, respectively. The ventral surface of the diaphragm was exposed by a laparotomy and two pairs of bipolar electrodes (coated stainless-steel wire with 1–2 mm exposed tips, #791500, A-M Systems) were inserted into the medial costal region of the bilateral diaphragm with an inter-electrode distance of ∼3 mm. The diaphragm EMG signals were amplified (1000 × ) and band-pass filtered (0.3–10 kHz) by a differential A/C amplifier (Model 1700, A-M Systems). The raw EMG signals were rectified and smoothed (time constant: 25 msec) by the Spike2 software (Cambridge Electronic Design Ltd.). This setting can appropriately reflect the discharge pattern (e.g., discharge onset and waveform) of diaphragm EMG signals for quantification of the burst amplitude. The physiological signals were digitized using CED Power 1401 (Cambridge Electronic Design Ltd.) at sampling rate of 100 Hz (e.g., airflow and blood pressure) or 10 kHz (e.g., diaphragm EMG activity) and stored in a computer by the Spike 2 software.
Experimental protocol
The bilateral phrenic nerve was exposed via a ventral approach at the cervical region. 13 –15 The baseline recording was established for 10 min under hyperoxic breathing condition. The right phrenic nerve was carefully dissected by #5 forceps and then sectioned (i.e., contralateral phrenicotomy) using microscissor to evaluate the contribution of the right diaphragm to the tidal volume generation. 9 The left phrenic nerve was subsequently separated and sectioned (i.e., ipsilateral phrenicotomy) at 10 min following contralateral phrenicotomy. The arterial blood sample (0.2 mL) was drawn to measure the arterial oxygen partial pressure (PaO2) and arterial carbon dioxide partial pressure (PaCO2) during the beseline and 10 min post contralateral phrenicotomy. At 10 min after ipsilateral phrenicotomy, the uninjured animals were euthanized by overdoses of KCl (150 mg/kg) and C2Hx animals were systemically perfused with heparin-saline followed by 4% paraformaldehyde (Alfa Aesar) and then 10 % sucrose (Sigma) in 4% paraformaldehyde.
Spinal cord histology
The cervical spinal cord tissue of C2Hx animals was removed and placed in 30% sucrose (Sigma) in phosphate buffered saline (UniRegion Bio-Tech). The cervical spinal cord was transversely sectioned (40 μm) via a cryostat (CM 1850, Leica) after the tissue sank. The cervical spinal sections were mounted on a glass slide (S7441, MATSUNAMI) and stained with cresyl violet (Acros Organics). The lesion was examined using an upright microscope (DM750, Leica) connected to a digital camera (EOS 600D, Canon). A histological example of cervical spinal cord section is shown in Figure 1. The extent of spared white matter ipsilateral to the hemisection (i.e., left) at the lesion epicenter was calculated by Image J software, and the data are expressed as a percentage of the contralateral (i.e., right) lateral and ventral white matter as previously described. 4,13,16 The spared white matter in C2Hx animals was 1.4 ± 1.0% of the contralateral white matter.
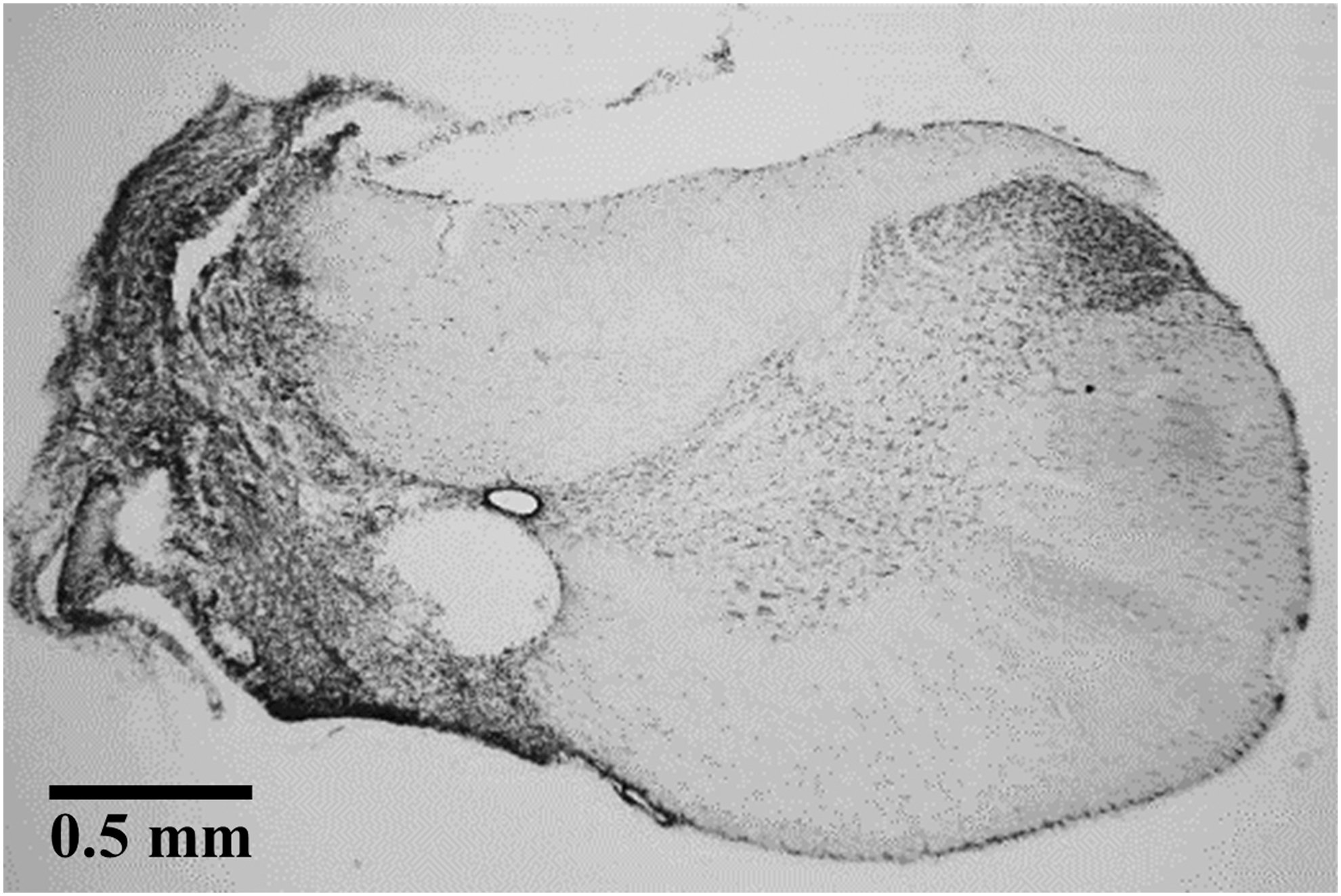
A representative histological C2 spinal cord section in a cervical spinal hemisection (C2Hx) animal at 8 weeks post-injury.
Statistical analysis
The cardiorespiratory parameters and diaphragm EMG activity were analyzed by the Spike 2 script. The respiratory cycle was calculated from the respiratory airflow trace recorded by the flow head and spirometer. The tidal volume was derived from the integration of the inspiratory flow and presented as an absolute value (i.e., mL) and normalized by the weight (i.e., mL/100 g). The diaphragm EMG burst amplitude was quantified as the difference between the maximal and minimal value of the rectified and smoothed EMG signals within an individual breath, which was detected using Spike 2 script. When there was an electrocardiogram artifact in EMG signals, the maximal value of the EMG signal would be manually detected by a horizontal cursor function of Spike 2 software.
All physiological parameters were averaged over 10 sec during the baseline (i.e., before phrenicotomy). The respiratory cycle, tidal volume, and diaphragm EMG activity were analyzed individually for the first breaths and averaged over 10 sec at 1 and 10 min post-phrenicotomy. The baseline cardiorespiratory pattern was analyzed by a two-way measures analysis of variance (ANOVA) followed by Student-Newman-Keuls (SNK) post hoc test (factor one: animal group [uninjured vs. C2Hx]; factor two: time-point [1 day, 2 and 8 weeks post-injury]). The blood gas parameters before and after contralateral phrenicotomy and time course of respiratory cycle and tidal volume following phrenicotomy was compared using a two-way mixed-design ANOVA followed by SNK post hoc test (factor one: animal group [uninjured vs. C2Hx]; factor two: before vs. after phrenicotomy). The time course of diaphragm EMG activity following phrenicotomy were analyzed by a one-way ANOVA followed by SNK post hoc test. A two-way mixed-design ANOVA followed by SNK post hoc test was used to compare the changes of tidal volume and blood gas parameters after phrenicotomy between uninjured and C2Hx animals at 1 day, 2 and 8 weeks post-injury.
All data are expressed as the mean ± standard error of the mean. A p value lower than 0.05 was considered statistically significant.
Results
The weight of the animal at three time-points post-injury is shown in Table 1. The weight of uninjured and C2Hx animals were similar at 1 day post-injury (Table 1). Both groups showed a gradual increase in body weight at 8 weeks post-injury (p < 0.01, Table 1); however, the weight of C2Hx animals was significantly lower than uninjured animals at 2 or 8 weeks post-injury (p < 0.01, Table 1).
p < 0.01 significance from the uninjured animals; † p < 0.01 compared with the 1 day group; θ p < 0.01 significance from 1 day and 2 week groups.
C2Hx, cervical spinal hemisection.
Cardiorespiratory pattern and diaphragm EMG activity following cervical spinal hemisection
The representative examples of cardiorespiratory pattern following C2Hx are shown in Figures 2 –4. The respiratory frequency was similar between uninjured and C2Hx animals at 1 day and 2 weeks post-injury, but C2Hx animals exhibited a lower respiratory frequency than uninjured animals at 8 weeks post-injury (p < 0.01, Fig. 5A). The tidal volume of C2Hx animals was significantly lower than that of uninjured animals at all time-points post-injury. Specifically, the tidal volume of C2Hx animals was 0.28 ± 0.03 mL/100 g at 1 day post-injury, which was significantly lower than that of uninjured animals (0.41 ± 0.01 mL/100 g, p < 0.01; Fig. 5B). The tidal volume of C2Hx animals was 0.33 ± 0.01 mL/100 g at 2 weeks and 0.29 ± 0.02 mL/100 g at 8 weeks post-injury; however, it remained lower than uninjured animals (0.40 ± 0.03 mL at 2 weeks and 0.37 ± 0.02 mL at 8 weeks post-laminectomy, p < 0.01; Fig. 5B). The minute ventilation was significantly lower in C2Hx animals than uninjured animals at 1 day and 8 weeks post-injury (p < 0.05, Fig. 5C).
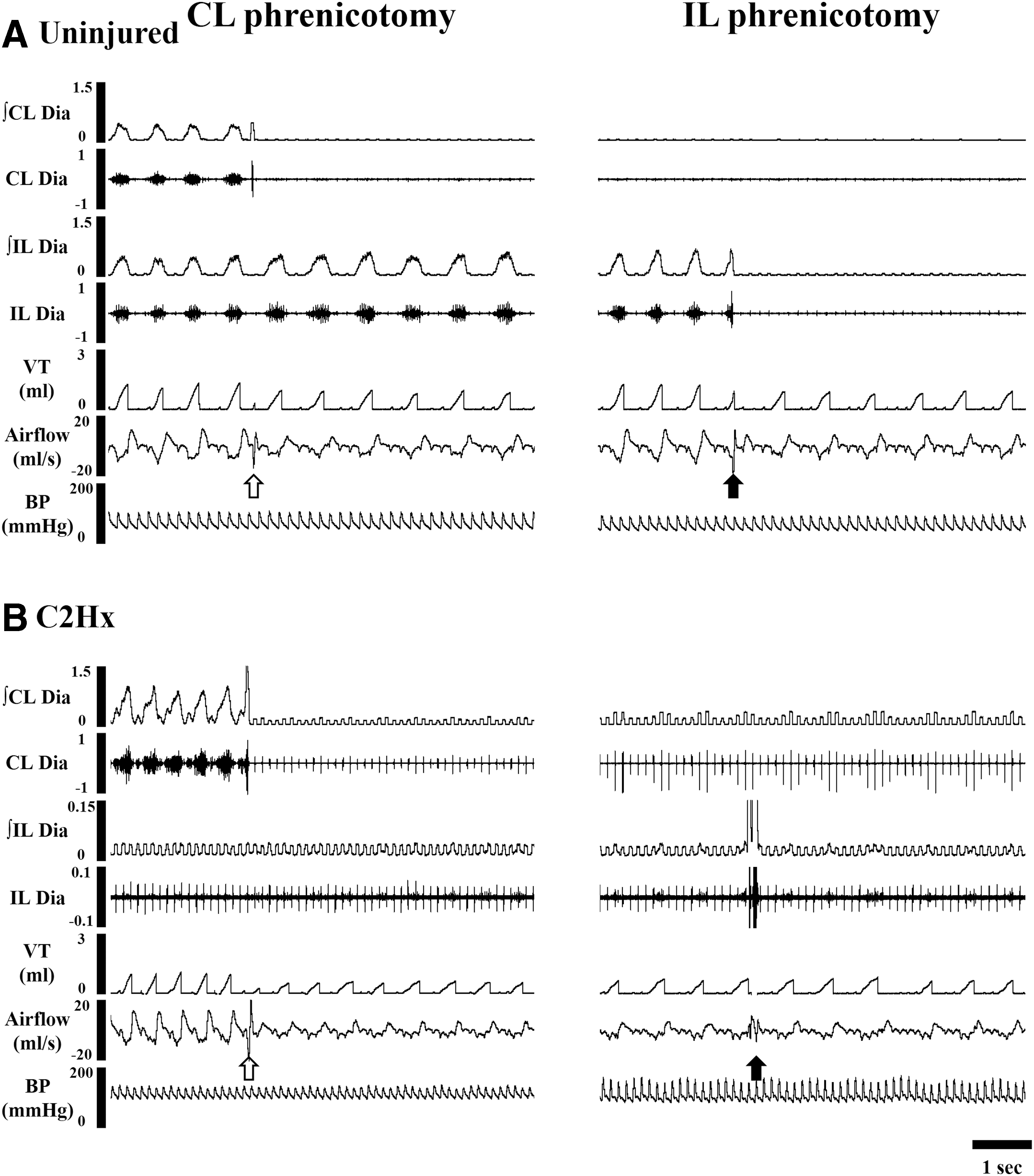
Representative examples of cardiorespiratory pattern and bilateral diaphragm electromyogram (EMG) activity before and after phrenicotomy in an uninjured

Representative examples of cardiorespiratory pattern and bilateral diaphragm electromyogram (EMG) activity before and after phrenicotomy in an uninjured

Representative examples of cardiorespiratory pattern and bilateral diaphragm electromyogram (EMG) activity before and after phrenicotomy in an uninjured
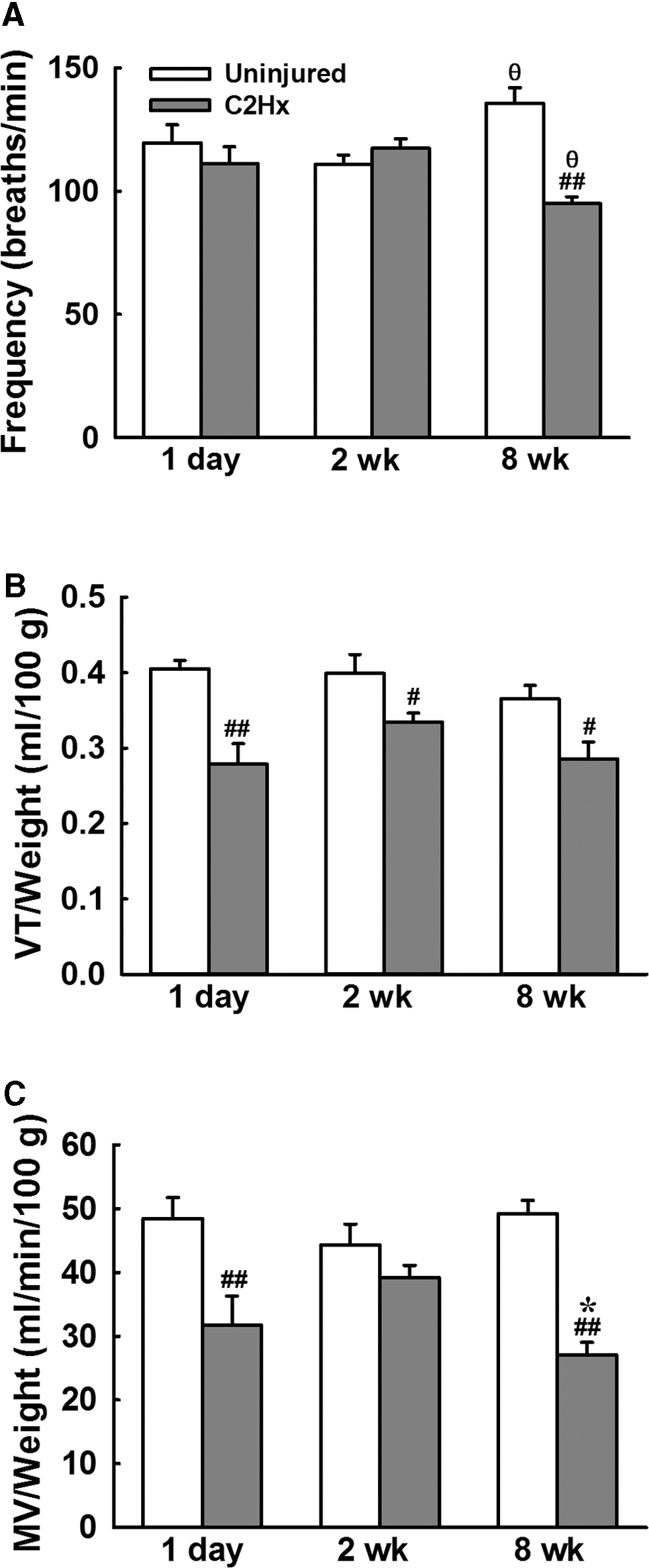
The baseline cardiorespiratory pattern of uninjured and C2Hx animals. The respiratory frequency
The mean arterial blood pressure and heart rate was similar between uninjured and C2Hx animals regardless of the time post-injury. But we noticed that the mean arterial blood pressure was relatively lower at 2 and 8 weeks post-injury than at 1 day post-injury in C2Hx animals (p < 0.05, Table 2).
p < 0.05 compared with the 1 day group.
C2Hx, cervical spinal hemisection; HR, heart rate; MAP, mean arterial blood pressure.
The representative examples of bilateral diaphragm EMG activity in uninjured and C2Hx animals are shown in Figures 2 –4. Bilateral diaphragm of uninjured animals exhibited a clear inspiratory rhythmic activity during the baseline at all time-points post-laminectomy. C2Hx animals retained the contralateral (i.e., right side) diaphragm EMG activity but lost their ipsilateral diaphragmatic bursting at 1 day post-injury (Fig. 2). A weak inspiratory bursting of the ipsilateral diaphragm on EMG could be observed in a few C2Hx animals at 2 (n = 3 in 8 rats) and 8 weeks (n = 2 in 8 rats) post-injury (Fig. 4), suggesting the crossed phrenic pathway was activated from the subchronic to chronic injury stage.
Because the animal breathed hyperoxic gas during the baseline condition, PaO2 was maintained above 100 mmHg in both uninjured and C2Hx animals at all-time points post-injury (Fig. 6). However, PaO2 of C2Hx animals was still significantly lower than that of uninjured animals at 1 day (uninjured: 181 ± 9 mmHg; C2Hx: 137 ± 7 mmHg) and 8 weeks (uninjured: 178 ± 4 mmHg; C2Hx: 131 ± 9 mmHg) post-injury (p < 0.05, Figs. 6A,C). Moreover, PaCO2 was higher in C2Hx animals than uninjured animals regardless of time-point post-injury (p < 0.05, Figs. 6A–C).

The blood gas parameters of uninjured and cervical spinal hemisection (C2Hx) animals before and after contralateral phrenicotomy. The arterial oxygen partial pressure (PaO2) and arterial carbon dioxide partial pressure (PaCO2) were quantified at 1 day
The influence of contralateral phrenicotomy on respiratory motor outputs
The representative examples of cardiorespiratory pattern and bilateral diaphragm EMG activity following contralateral phrenicotomy are shown in Figures 2 –4. The tidal volume was immediately reduced following contralateral phrenicotomy in uninjured and C2Hx animals. Specifically, the tidal volume significantly reduced from 1.31 ± 0.04 mL to 1.05 ± 0.04 mL in uninjured animals, and 0.90 ± 0.08 mL to 0.42 ± 0.06 mL in C2Hx animals following contralateral phrenicotomy at 1 day post-injury (p < 0.05, Fig. 7A). The tidal volume was persistently lower than the baseline for at least 10 breaths post-phrenicotomy and it could partially recover at 1–10 min post-phrenicotomy (Fig. 7A) in both groups. The respiratory cycle duration immediately increased associated with the reduced tidal volume in acute C2Hx animals; however, the respiratory cycle duration remained longer than the baseline value (0.55 ± 0.03 sec) at 10 min post-phrenicotomy (0.80 ± 0.05 sec) even as the tidal volume gradually returned toward the baseline value. Notably, there were three C2Hx animals (n = 3/8, 37.5%) that died within 10 min after contralateral phrenicotomy, suggesting the contralateral phrenic nerve was essential for the survival of acute C2Hx animals. At 2 and 8 weeks post-injury, contralateral phrenicotomy still induced a substantial reduction in the tidal volume in both uninjured and C2Hx animals (Figs. 7B,C). However, most C2Hx animals (2 weeks: 7/8, 87.5%; 8 weeks: 7/8, 87.5%) were still surviving after section of the contralateral phrenic nerve. The respiratory cycle duration was not influenced by contralateral phrenicotomy in uninjured animals, but C2Hx animals increased the respiratory cycle duration following section of the contralateral phrenic nerve (p < 0.05).

Alteration of the tidal volume following contralateral and ipsilateral phrenicotomy in uninjured and cervical spinal hemisection (C2Hx) animals at 1 day
To evaluate the functional impact of the contralateral phrenic nerve on tidal volume generation, we specifically compared the alteration of tidal volume immediately after contralateral phrenicotomy (i.e., first breath) between uninjured and C2Hx animals. The data demonstrated that the reduction of tidal volume was significantly greater in C2Hx animals at 1 day (uninjured: 0.26 ± 0.03 mL vs. C2Hx: 0.48 ± 0.04 mL) and 8 weeks (uninjured: 0.49 ± 0.06 mL vs. C2Hx: 0.63 ± 0.03 mL) post-injury (p < 0.05). When the data were presented as a percentage of the baseline value, the tidal volume of C2Hx animals (1 day: 46 ± 3%; 2 weeks: 57 ± 4 %; 8 weeks: 51 ± 4%) was significantly lower than that of uninjured animals (1 day: 80 ± 2%; 2 weeks: 75 ± 3%; 8 weeks: 78 ± 2%; p < 0.05, Fig. 7). These findings suggest that the contralateral phrenic nerve was responsible for ∼50% of the tidal volume generation in the C2Hx animals but only contributed to 20–25% of the tidal volume in the uninjured animals.
The contralateral diaphragm EMG activity immediately ceased after contralateral phrenicotomy in both uninjured and C2Hx animals in all groups (Fig. 8), suggesting this approach can eliminate phrenic motor outputs. The ipsilateral diaphragm EMG activity in uninjured animals was not significantly changed; however, previous silent diaphragmatic bursting could be recruited by contralateral phrenicotomy in a subset of C2Hx animals (1 day post-injury, n = 1; 2 weeks post-injury, n = 2; 8 weeks post-injury, n = 3). Moreover, the burst amplitude of ipsilateral diaphragm could be significantly increased from 0.006 ± 0.004 mV to 0.054 ± 0.012 mV at 1 min post-phrenicotomy in C2Hx animals at 8 weeks post-injury (Fig. 8C).
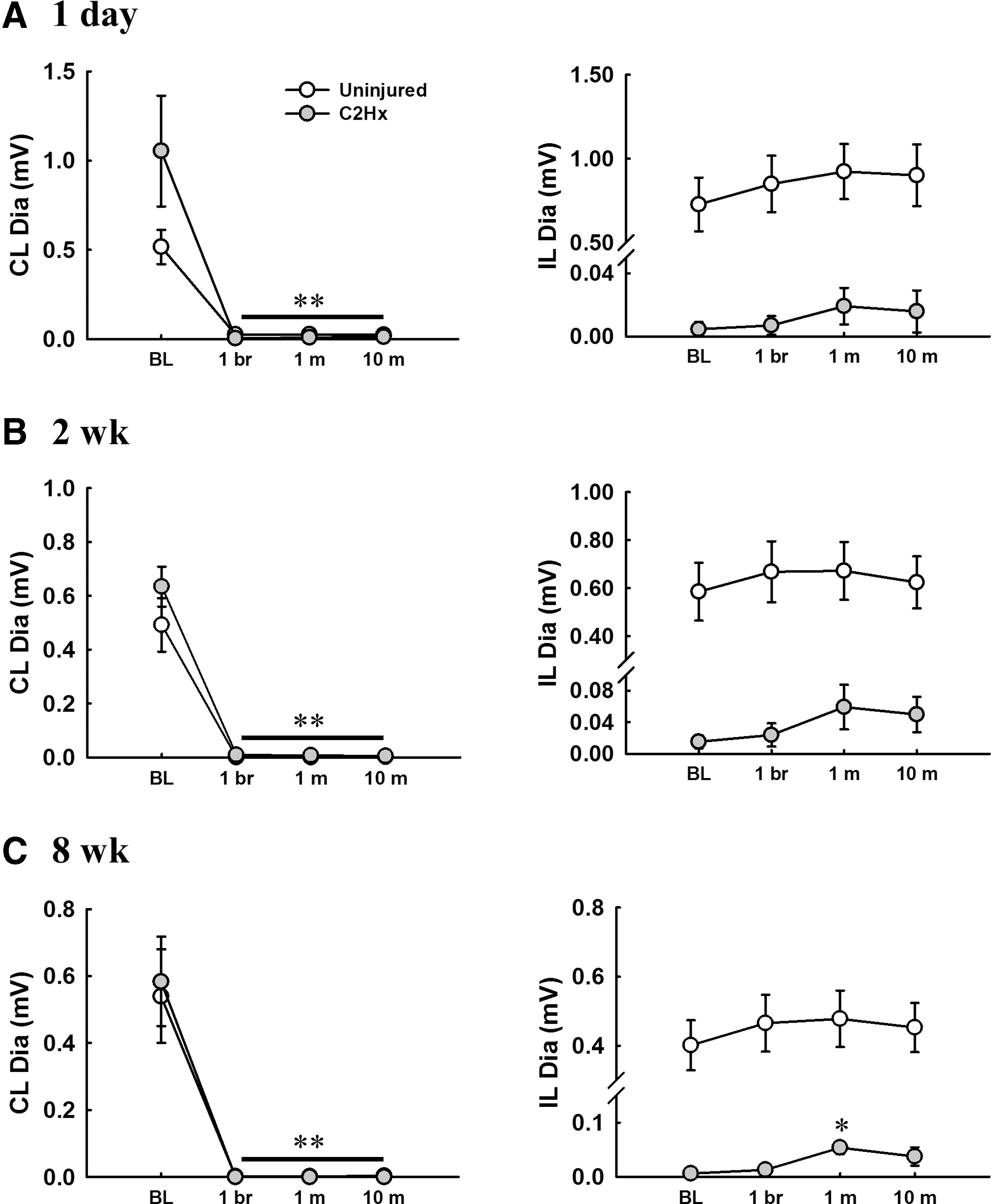
Alteration of the contralateral and ipsilateral diaphragm electromyogram (EMG) activity following contralateral phrenicotomy in uninjured and cervical spinal hemisection (C2Hx) animals at 1 day
The changes of blood gas parameters following contralateral phrenicotomy was also quantified to evaluate the functional role of contralateral phrenic motor outputs on blood gas homeostasis. The results demonstrated that contralateral phrenicotomy did not significantly modulate the PaO2 and PaCO2 in uninjured animals (Fig. 6); however, PaO2 was significantly reduced from 137 ± 7 mmHg to 83 ± 16 mmHg, and PaCO2 was significantly increased from 76 ± 8 mmHg to 117 ± 5 mmHg in response to contralateral phrenicotomy in C2Hx animals at 1 day post-injury (p < 0.01, Fig. 6A,D). At 2 and 8 weeks post-injury, PaO2 could be maintained but PaCO2 was still elevated following contralateral phrenicotomy. Moreover, the degree of increased PaCO2 was greater in C2Hx animals at 1 day (51 ± 5 mmHg) post-injury than at 2 (25 ± 3 mmHg) and 8 (34 ± 7 mmHg) weeks post-injury (p < 0.05, Fig. 6D).
The influence of ipsilateral phrenicotomy on respiratory motor outputs
The ipsilateral diaphragm activity was immediately eliminated by ipsilateral phrenicotomy in both uninjured and C2Hx animals (Figs. 2 –4). Although the respiratory cycle duration was not significantly changed, the tidal volume of uninjured animals was significantly reduced following ipsilateral phrenicotomy. Specifically, the tidal volume was reduced from 1.25 ± 0.04 mL to 0.85 ± 0.05 mL immediately following ipsilateral phrenicotomy at 1 day post-laminectomy (p < 0.05, Fig. 7A). At 2 and 8 weeks, the tidal volume also significantly declined after ipsilateral phrenicotomy (p < 0.05, Figs. 7B,C). Notably, ipsilateral phrenicotomy did not significantly alter the tidal volume in C2Hx animals at all time-points post-injury (Fig. 7).
The maintenance of the tidal volume following phrenicotomy
To evaluate the contribution of residual respiratory motor outputs following phrenicotomy to the tidal volume maintenance, we specifically compared the tidal volume under phrenic-intact condition (i.e., baseline) with the contralateral phrenicotomized (i.e., 10 min after contralateral phrenicotomy) and bilateral phrenicotomized condition (i.e., 10 min after ipsilateral phrenicotomy in contralateral phrenicotomized rats). In uninjured animals, the stable tidal volume generated by the contralateral phrenicotomized rats was 96 ± 2%, 95 ± 2%, and 95 ± 3% at 1 day, 2 weeks, and 8 weeks post-laminectomy, respectively (Fig. 7). The subsequent ipsilateral phrenicotomy also caused a further decrease in the tidal volume; however, the tidal volume could still maintain above 80% of the baseline in these bilateral phrenicotomized rats (1 day: 89 ± 1%; 2 weeks: 85 ± 3%; 8 weeks: 87 ± 4%; Fig.7). The stable tidal volume generated by the contralateral phrenicotomized C2Hx rat (1 day: 79 ± 5%; 2 weeks: 81 ± 5%; 8 weeks: 84 ± 4%) was significantly lower than that of uninjured animals at three time-points post-injury (p < 0.05, Fig. 7), but the subsequent ipsilateral phrenicotomy did not significantly influence the tidal volume in C2Hx animals. Interestingly, we noticed that the tidal volume can also maintain above 75% of the baseline in C2Hx animals at 2 (86 ± 2%) and 8 (78 ± 6%) weeks post-injury after bilateral phrenicotomy.
Discussion
The present study demonstrated that contralateral phrenicotomy caused a robust reduction in the tidal volume and resulted in a disruption of blood gas homeostasis at acute injury state. During subchronic to chronic injury state, contralateral phrenicotomy still induced a greater reduction in the tidal volume and elevation of PaCO2 in C2Hx animals than in uninjured animals. Notably, subsequent ipsilateral phrenicotomy following contralateral phrenic nerve section did not significantly alter the ventilation. These data suggest that compensatory function of the contralateral phrenic motor outputs exerts a critical role in maintenance of the essential ventilation following unilateral cervical spinal cord injury.
Critique of methods
The present study evaluated the functional contribution of phrenic motor outputs on the tidal volume by assessing the immediate changes of tidal volume following section of the phrenic nerve. The diaphragm EMG activity was abolished following phrenicotomy, suggesting this approach can eliminate the phrenic motor output and cause paralysis of the diaphragm. However, the phrenic nerve is composed of both sensory afferents and motor efferents. Previous studies already have shown that phrenic nerve activity ipsilateral to the lateral cervical spinal cord injury is differentially modulated by contralateral and ipsilateral phrenic afferent. 17,18 Therefore, we cannot exclude that the removal of phrenic afferent inputs may also have an impact on the tidal volume generation. Accordingly, the alteration of tidal volume following phrenicotomy in our model should be considered as a combinatorial effect of elimination of both phrenic sensory and motor signals. In addition, previous reports have indicated that there is an accessory phrenic branch innervating the diaphragm in the rat. 19,20 Because the purpose of the current study was to evaluate the immediate change of the tidal volume following phrenicotomy, the phrenicotomy procedure had to be as quick as possible (e.g., within 1 breath). Accordingly, it was not feasible to cut both the main phrenic nerve and the accessory phrenic nerve at the same time. Thus, the alteration of the respiratory parameters following phrenicotomy in the present study mainly resulted from the contribution of the main phrenic nerve.
The potential mechanism underlying compensation of the contralateral diaphragm
Our current data demonstrate that the tidal volume was immediately and substantially reduced in C2Hx animals (approximately 42–52% decline) compared with uninjured animals (approximately 21–27% decline) following contralateral phrenicotomy, suggesting there is a compensatory function of the contralateral diaphragm on the tidal volume generation after unilateral cervical spinal cord injury. Several factors may contribute to this phenomenon. First, we noticed that PaCO2 was greater in C2Hx animals than in uninjured animals. The elevated PaCO2 can activate central chemoreceptors and then enhance the respiratory motor outputs. 21 Accordingly, the greater contribution of the contralateral diaphragm to the tidal volume generation could be partially contributed by higher respiratory drives in C2Hx animals. Second, unilateral cervical hemisection not only interrupts the ipsilateral bulbospinal respiratory pathway to the ipsilateral phrenic motoneuron pool but it also damages the projections from the brainstem to the contralateral phrenic nucleus. 22 In addition to the excitatory drives from the rostral ventral respiratory group, there are inhibitory inputs from the Bötzinger complex to the phrenic motoneurons. 23 Therefore, C2Hx may remove this inhibitory pathway and augment contralateral diaphragmatic activity. Third, the phrenic motor activity is primarily driven by the bulbospinal respiratory pathways under normal conditions, but several studies already have demonstrated that spinal interneuron may be involved in modulation of respiratory motor activity after spinal cord injury. 24,25 Specifically, Lane and colleagues have shown that there is an increase in interneuron innervations of spared phrenic motoneurons after spinal cord injury. 26 Accordingly, re-organization of the spinal circuit may provide additional excitatory inputs to the phrenic motoneurons and enhance its contribution to the tidal volume. Fourth, the tidal volume generation depends on coordination of the phrenic motor activity and diaphragmatic contraction. It is established that cervical spinal cord injury can induce alterations in neuromuscular junction and fiber size of the diaphragm. 27 –29 Although these studies focus the diaphragm properties ipsilateral to the lesion, we speculated that neuromuscular junction, fiber size, and/or composition of the contralateral diaphragm may also undergo a remodeling process and influence the function of the diaphragm. Lastly, the above-mentioned factors may be all involved in the compensatory response of the contralateral diaphragm, but there may be a differential mechanism underlying the compensatory response of the contralateral diaphragm during acute versus chronic injury state.
The functional impact of compensatory response following cervical spinal cord injury
The current data demonstrated that some C2Hx animals (3/8, 37.5%) died after contralateral phrenicotomy during the acute injury phase. During subchronic to chronic injury phase, the blood homeostasis was still disrupted (e.g., PaCO2 increased) after section of the contralateral phrenic nerve in C2Hx animals. These data suggest that the contralateral phrenic nerve is critical to the maintenance of the adequate ventilation after cervical spinal cord injury. Interestingly, ipsilateral phrenicotomy following contralateral phenicotomy only had a slight influence on the ventilation. In addition, our recent report demonstrated that augmentation of ipsilateral diaphragm EMG activity following exogenous application of 5-HT agents was not associated with improvement of the tidal volume in C2Hx animals. 4 Moreover, Navarrete-Opazo and associates showed that daily mild acute intermittent hypoxia-induced recovery of the tidal volume in C2Hx animals was correlated with an increase of contralateral but not ipsilateral diaphragm activity. 12 Together, these findings suggest that maintenance of ventilation during the baseline and improvement of the tidal volume induced by the pharmacological intervention or rehabilitative training mainly depends on the function of the contralateral but not ipsilateral diaphragm.
In addition to the phrenic motor system, the intercostal muscle and other accessory inspiratory muscles also play a crucial role in the maintenance of the tidal volume following phrenicotomy and C2Hx. Our current data demonstrate that uninjured animals can still retain >80% of baseline tidal volume after the bilateral phrenic nerve is sectioned. Interestingly, C2Hx animals can also have ∼75% of baseline tidal volume after bilateral phrenicotomy. The absolute value of the tidal volume in bilateral phrenicotomized C2Hx animals was ∼50–60% of the tidal volume in phrenic-intact uninjured animals. In other words, both contralateral uninjured and ipsilateral injured side intercostal and other accessory muscles can enable the C2Hx animals to survive after loss of bilateral phrenic innervation.
Physiological significance
Previous studies have shown the functional contribution of the phrenic motor outputs ipsilateral to the cervical spinal cord injury. For example, Golder and co-workers demonstrated that animals had a significant lower tidal volume at 2 months following dual injury (e.g., C2Hx and ipsilateral phrenicotomy) than did C2Hx only animals during high respiratory drives (e.g., after vagotomy and during sigh). 30 Dougherty and colleagues noticed that acute ipsilateral phrenicotomy induced an ∼16 % reduction in the tidal volume at 2 and 8 weeks post-C2Hx. 9 The current study extended these results and demonstrated that the contralateral phrenic nerve plays a predominant role in maintenance of the tidal volume during baseline breathing from the acute to chronic injury state. These data indicate that there is a differential functional role of ipsilateral versus contralateral phrenic motor outputs on the tidal volume during different time-points after injury.
Although several approaches have been utilized to enhance inspiratory activity of the ipsilateral phrenic nerve or diaphragm following unilateral cervical spinal cord injury, 4,31 –34 ; however, recovery of ipsilateral phrenic motor outputs may not be sufficient to induce functional improvements in the tidal volume due to limitation of recovery status and/or atrophy of the diaphragm. 4,35 The current study demonstrates that the contralateral diaphragm plays a predominant role in the maintenance of the tidal volume during baseline breathing. Although compensatory response of the contralateral phrenic motor outputs following cervical spinal cord injury may prevent further expression of respiratory neuroplasticity. 36 It has been shown that daily acute intermittent hypoxia can restore the capacity of the contralateral phrenic nerve to express intermittent hypoxia-induced long-term facilitation. 37 Moreover, augmentation of contralateral diaphragm activity has been associated with an increase of the tidal volume induced after daily acute intermittent hypoxia training in awake rats. 12 Accordingly, it is critical to understand the process underlying the compensatory mechanism after spinal cord injury, and the contralateral respiratory motoneurons and/or muscles may be potential targets for a future therapeutic strategy.
Footnotes
Acknowledgments
Support for this work was provided by grants from the National Health Research Institutes (NHRI-EX105-10223NC), Ministry of Science and Technology (Most 102-2320-B-110-004-MY3 & 105-2628-B-110-002-MY3), and NSYSU-KMU Joint Research Project (105-I004 & 106-I009).
Author Disclosure Statement
No competing financial interests exist.