
Abstract
Previous work in this laboratory used underwater explosive exposures to isolate the effects of shock-induced principle stress without shear on rat brain aggregate cultures. The current study has utilized simulated air blast to expose aggregates in suspension and enclosed within a spherical shell, enabling the examination of a much more complex biomechanical insult. Culture medium–filled spheres were exposed to single pulse overpressures of 15–30 psi (∼6–7 msec duration) and measurements within the sphere at defined sites showed complex and spatially dependent pressure changes. When brain aggregates were exposed to similar conditions, no cell death was observed and no changes in several commonly used biomarkers of traumatic brain injury (TBI) were noted. However, similarly to underwater blast, immediate and transient increases in the protein kinase B signaling pathway were observed at early time–points (3 days). In contrast, the oligodendrocyte marker 2′,3′-cyclic nucleotide 3′-phosphodiesterase, as well as vascular endothelial growth factor, both displayed markedly delayed (14–28 days) and pressure-dependent responses. The imposition of a spherical shell between the single pulse shock wave and the target brain tissue introduces greatly increased complexity to the insult. This work shows that brain tissue can not only discriminate the nature of the pressure changes it experiences, but that a portion of its response is significantly delayed. These results have mechanistic implications for the study of primary blast-induced TBI and also highlight the importance of rigorously characterizing the actual pressure variations experienced by target tissue in primary blast studies.
Introduction
T
The use of cell and tissue culture systems to investigate primary blast effects on the brain offers approaches that can simplify the questions asked in terms of isolating cause–effect phenomena and the means to answer them. This approach has been well used historically to investigate the biomechanical and injury effects of the secondary and tertiary aspects of blast. 14,15 More recently, several laboratories also have utilized a variety of cell and tissue culture models to study primary blast. 16 –33 While the technical aspects of these systems still remain challenging with respect to the simulation of shock waves relevant to those produced by survivable IEDs, as well as the requirement to maintain both the integrity and sterility of the cultures, the biological complexity of these model systems is substantially reduced compared with animal models. Well-considered studies using tissue culture platforms therefore present the opportunity to investigate the effect of shock waves on the skull and brain tissue without the scaling issues that currently complicate the interpretation of primary blast studies using small rodents. Further, definitive conclusions can be made as to the importance of the interactions of high frequency pressure changes directly on brain tissue. In contrast to whole animal work, the roles of other potential mechanisms of primary blast injury that have been hypothesized as occurring in vivo, such as whole–body systemic effects or the thoracic transfer of kinetic energy through the vasculature to the brain, 34 –37 need not be considered.
Brain cell aggregate cultures exhibit a multi-cell type composition and can be kept in suspension culture for periods of up to several weeks. 38 –40 They are being used in these laboratories as part of a comprehensive and stepwise research effort to understand the effects of primary blast on the brain. In a previous study, this model system was exposed to underwater blast. 41 By utilizing highly hydrolyzed dialysis tubing containment during exposure, suspended aggregates were exposed to single pulse blast waves with minimized boundary effects. Subsequent sampling of the exposed cultures showed that the brain tissue responded in a subtle but reproducible manner to this simplified, well defined, and measurable shock wave–induced principal stress. Although this work demonstrated that the culture model system was sensitive to single pulse overpressures of short duration, it was necessary to establish that it could also discriminate between different types of pressure insult. This report details the effects of imposing a spherical shell between a single pulse air shock wave and the target brain tissue, thus introducing fundamental changes and greatly increased complexity into the nature of the insult, compared with the previous underwater blast studies. The results illustrate the sensitivity of brain tissue to blast/shock waves and also show that this model system can discriminate between different types of pressure insult. This has mechanistic implications for the study of primary blast–induced TBI and importantly, also demonstrates the importance of stringently defining and characterizing the pressure wave exposures actually experienced by target tissue in primary blast studies.
Methods
Materials
Basal medium Eagle's (BME), fetal calf serum (FCS), glutamine, penicillin-streptomycin, and phosphate-buffered saline (PBS) were purchased from Invitrogen (Burlington, Ontario). Routine culture medium consisted of BME supplemented to 10% FCS (v/v), penicillin-streptomycin (10 IU/mL-10 μg/mL), L-glutamine (2 mM) and D-glucose (10 mM). CelLytic MT Lysis Reagent (Sigma Chemical Co., St. Louis, MO) was utilized as a lysis buffer. Other chemicals were of the purest grade available from regular commercial sources. Glass spinner flasks with a culture volume of 125 mL were purchased from Wheaton (Techne, NJ), and siliconized prior to use to prevent adhesion of aggregates to the inner walls. Four-place stirrers and their controllers were obtained from Thermo Fisher Scientific (Burlington, Ontario).
Rat brain cell aggregate culture
In conducting this research, the authors adhered to the Guide to the Care and Use of Experimental Animals and The Ethics of Animal Experimentation published by the Canadian Council on Animal Care. Timed pregnant Sprague-Dawley rats were acquired from Charles River Laboratories (St. Constant, Quebec). The methods to generate rat brain cortical aggregates have been described previously 41 and loosely follow those used by the Honegger and Sa Santos laboratories. 38 –40 Briefly, embryos were isolated at 17 days of gestation and the brains removed. Cells from the cerebral hemispheres were isolated and placed into 125 mL spinner flasks at ∼2.0–2.4 × 108 cells/flask in culture medium supplemented to 15% FCS. The spinner flasks were placed onto stirrers and refed with culture medium (10% FCS) three to four times per week, as required. Cultures were closely followed and the rotational speeds adjusted to ensure a gradual increase in aggregate size to 350–500 μm diameter. Aggregate cultures were exposed to simulated blast at 27–28 days in vitro.
Advanced Blast Simulator
A custom‐built Advanced Blast Simulator (ABS; ∼30.5 cm in diameter and 5.79 m in length; Fig. 1) located at DRDC Suffield was used for producing simulated blast waves. 12, 42 The ABS consists of a “driver” section filled with high-pressure gas separated by a frangible diaphragm from a transition section leading to an ambient pressure test section. Controlled pressurization of the driver causes rupture of the diaphragm at predetermined pressures, abruptly releasing the high-pressure gas and driving a tailored shock wave down the length of the test section. The inclusion of an End Wave Eliminator at the end of the test section prevents rarefaction waves propagating back into the testing area, and also mitigates noise and gas efflux into the laboratory space. By means of these unique design features, the ABS generates highly reproducible single pulse shock waves tailored to replicate those of explosive blast, including sharply defined static and dynamic overpressure rise times, underpressures, and secondary shock waves. 42 The required target pressure and waveform shapes are achieved using compressed helium in the driver and various layering and thicknesses of cellulose acetate sheets for the frangible diaphragm.

Advanced Blast Simulator used for these studies. A divergent driver (foreground) is pressurized with helium and the cellulose acetate diaphragm fails at a predetermined driver pressure, sending a shock wave through the transition section and into the test section. Rarefaction waves are prevented from re-entering the test section through the use of a tunable End Wave Eliminator located in the white dump tank.
ABS pressure data acquisition
Static pressures were measured using PCB 113A28 gauges placed at 2780, 3280, 3780, 4280, and 4780 mm from the diaphragm. All pressure data was recorded using a custom Labview interface and recorded on a GaGe Octopus 8389 CompuScope PCIe digitizer board at a sampling rate of 500,000 samples/sec.
Internal sphere pressure measurements
In studies assessing the pressure changes within the sphere, aggregates were not used. Millar gauges (SPR-524) were placed into the interior of a water-filled polypropylene sphere (50 mm diameter, ∼1.0 mm wall thickness, ∼56 mL volume) and oriented such that the incident air shock wave would transmit through the sphere wall and engulf the transducer. The sphere was mounted on the end of the protruding sting (Fig. 2A) as previously described 43 and longitudinally positioned along the central axis of the tube at 4.28 m downstream of the diaphragm. The gauges were placed inside the sphere along the central axis and located at positions in the center, 16 mm from the center towards the front of the sphere, or 23 mm from the center towards the back of the sphere. Transducer cables extended through the bolt securing the sphere to the sting, out the back of the cross/box assembly (Fig. 2A) and emerged out of one of the shock tube wall instrumentation ports. This cross/box structure was constructed from aluminum sheet and thus provided minimal disturbance to the flow. The assembly was secured into position by the friction of the rails against the ABS inner walls and high speed photography revealed no displacement during testing. It was positioned into the test section end of the ABS so that the leading edge of the sphere closest to the diaphragm was opposite the static pressure gauge at location 4270 mm. This positioning also allowed access to the sphere through a port opposite the gauge.
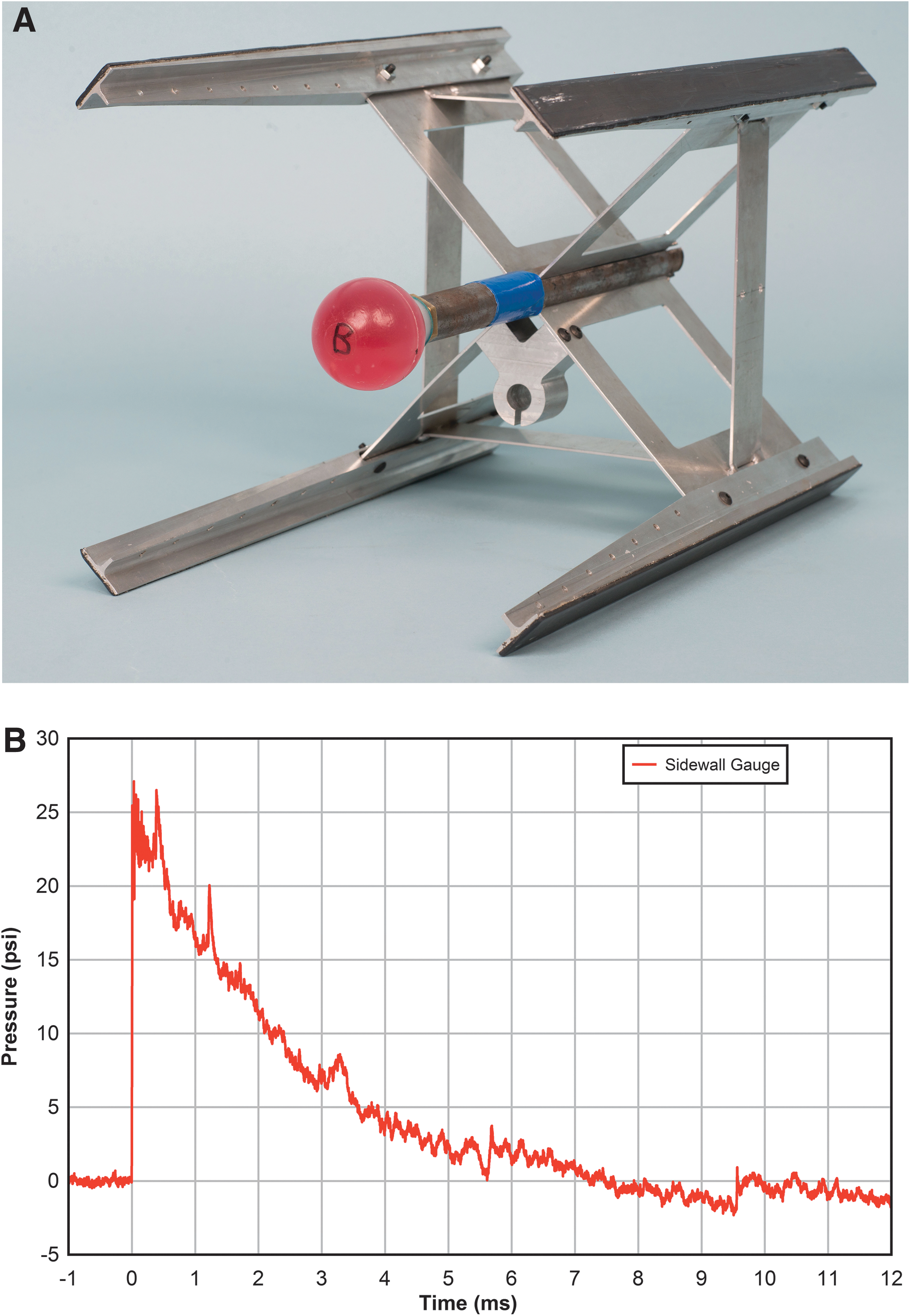
Exposure apparatus for brain cell aggregates and representative simulated blast exposure waveform.
Blast exposure
On the day of exposure, test cultures were isolated from their spinner flasks by sedimentation and placed into sterile polypropylene spherical shells fitted with threaded collars. The aggregates were suspended in complete culture medium and the shells were sealed using threaded bolts. The aggregate-filled spheres were immediately taken to the ABS facility and fitted onto the mounting apparatus described above using the threaded bolt. When the ABS was ready to fire, the sphere was turned 180° by rotating the threaded collar, the port was sealed, and the driver pressurized. The time elapsed from sphere rotation until the shot was less than ∼15 sec and ensured that the aggregates were in suspension during shock wave exposure as they sedimented to the bottom of the sphere after rotation. Immediately after exposure, the shell was removed from the ABS and transported back to the laboratory, where the aggregates were returned to culture until harvested for analysis. On the day of aggregate harvest, the spinner flasks were removed from the incubator and the aggregates allowed to sediment to the bottom of the culture vessels. A portion of the aggregates was removed and the spinner flask containing the balance of the culture was returned to the incubator. The aggregates were homogenized and cell lysates prepared in CelLytic. After protein determination, the lysates were adjusted to 1 mg/mL of protein and aliquots frozen at −80°C until assayed.
Western blot analysis
Protein lysates from brain aggregates (10–15 μg protein) were separated on a 4–20% gradient pre-cast gel (Bio-Rad Laboratories, Mississauga, ON, Canada) and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with 5% skim milk, 0.1% Tween 20 in PBS and then incubated with antibodies against glial fibrillary acidic protein (GFAP; 1:2000; Millipore, Etobicoke, Ontario), neurofilament H (NFH,1:1000; Millipore), phospho-Akt Ser473 or total protein kinase B (Akt; New England Biolabs; 1:1000) overnight at 4°C. The membranes were then washed with PBS plus 0.1% Tween 20 three times for 10 min each. Primary antibodies were probed by incubating membranes with a secondary antibody, donkey anti-rabbit or mouse immunoglobulin G–horseradish peroxidase (GE Health Care Biosciences, Quebec) diluted 1:5000. Detection was carried out by using ECL advanced detection reagents (GE Health Care Biosciences) and imaged using a Molecular Imager VersaDoc MP 4000 system (Bio-Rad Laboratories). To confirm equal protein loading, blots were re-probed with anti-pan-actin antibody (1:1000; Cell Signaling, MA). Band intensities were quantified using Quantity One software and expressed as a percentage of control.
Immunohistochemistry and confocal microscopy
Aggregates were fixed with 4% paraformaldehyde for 2 h at room temperature and then transferred to 30% sucrose in PBS in a 1.5 mL microcentrifuge tube. After the aggregates sedimented to the bottom of the tube, they were embedded in a frozen section compound (Surgipath, FSC 22; Leica Microsystems Canada, Concord, Ontario). Frozen sections (15 μm thickness) of fixed embedded aggregates were prepared with the Cryostat (Leica Microsystems Canada). After rehydration, aggregate sections were permeabilized with 0.25% triton X-100 for 30 min, then blocked in 5% goat serum for 1 h before being stained with primary antibodies against NFH or GFAP. Stained primary antibodies were detected with Alexa Fluor-488 or Alexa Fluor-647 tagged secondary antibodies (Thermo Fisher Scientific). At the end of the staining process, all sections were counter-stained with 4',6-diamidino-2-phenylindole (Invitrogen) to view nuclei. Stained sections were viewed with a Quorum WaveFX laser scanning confocal microscope at 400 × magnification and images were captured with a Hamamatsu EM-CCD camera.
Biochemistry
Rat brain cell aggregate lysates were assessed for glutamine synthetase (GS) and choline acetyltransferase (ChAT) activity using spectrophotometric titerplate assays. Glutamine synthetase activity was measured using a modification of the method of Gawronski and Benson, 44 while the method of Chao and Wolfgram 45 was adapted to measure ChAT. Aggregate homogenate lysates were assayed for 2′,3′-cyclic nucleotide 3′-phosphodiesterase (CNPase) activity kinetically using a modification of the assay developed by Dreiling and Mattson. 46 Acetylcholinesterase (AChE) activity was measured using the radiometric assay of Johnson and Russell. 47 Lactate dehydrogenase (LDH) was determined using a LDH kit (Sigma Chem. Co.). Vascular endothelial growth factor (VEGF) levels were measured according to manufacturer instructions using components contained in the Bio-Plex Pro Rat Cytokine Assay 23-Plex, Group 1 kit (Bio-Rad Laboratories).
Statistical analysis
Statistical differences were analyzed using one-way analysis of variance (ANOVA) and Dunnett's test using GraphPad InStat (San Diego, CA).
Results
Figure 2B depicts a representative 25 psi static pressure trace from the sidewall gauge in line with the leading edge of the test sphere. The simulated blast is single pulse in nature, with an immediate rise in pressure that then decays exponentially. The peaks at 0.5 and 1.2 msec are caused by the reflections of the incident wave off of the sphere. Table 1 shows the high precision and reproducibility of the ABS in simulating single pulse blast wave challenges at target pressures of 15, 20, 25, and 30 psi. Actual static overpressures measured were within 2% of the nominal target pressure, with durations of 6.4–7.6 msec. Peak positive phase impulses were 30.0–66.9 psi msec.
The data represent pressure histories obtained in the ABS studies described in the current report, while the UNDEX values are derived from the data reported by Sawyer and colleagues. 41
ABS, Advanced Blast Simulator; UNDEX, underwater explosion.
The pressure changes inside the sphere were complex and spatially dependent, as illustrated by the pressure histories recorded at forward, center, and back locations of a sphere exposed to 30 psi overpressure (Fig. 3A-3C). The forward gauge first detects the transmission of the reflected pressure off of the leading edge of the sphere (Fig. 3A). This then rapidly decays towards ambient pressure, but in an oscillatory fashion as reflections come off of the rear and stem mount of the vibrating sphere. At the center of the sphere (Fig. 3B), a similar pressure history was observed, but with the amplitude of the oscillations attenuated due to the superimposition of the reflecting waves. In contrast, at the rear of the sphere (Fig. 3C), the wave oscillations were more pronounced compared with the other locations due to their close proximity (2 mm) to the reflecting surface of the stem mount.
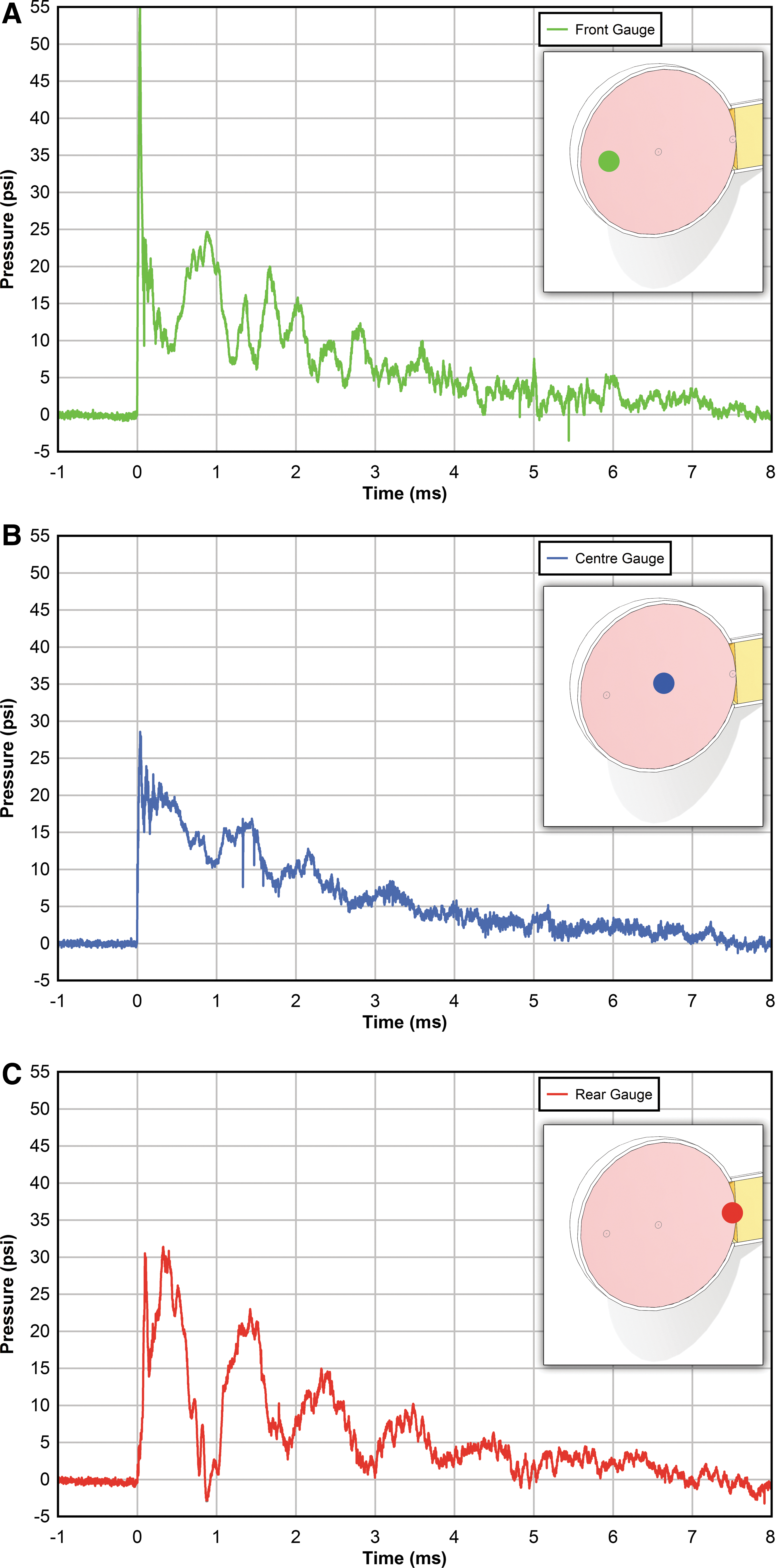
Representative pressure histories from the interior of a 50 mm diameter sphere exposed to 30 psi overpressure.
Aggregate cultures were exposed to air blast on Day 27 or 28 of culture. On the day of treatment, the aggregates from multiple spinner flasks were combined and evenly distributed into a predetermined number of samples so that each pressure response was accomplished using a representative sample from the same population. At this time, the aggregates were spherical with a smooth surface, and no debris and few or no single cells were noted in the cultures. Time from placement of the aggregates into the spheres, exposure to simulated blast, and their return back into routine culture conditions in spinner flasks was generally less than 25 min. No adverse effects due to this interval outside of optimal culture conditions were noted microscopically in the sham controls, although the presence of single cells was not uncommon immediately after shock wave exposure. Sampling from the spinner flasks was carried out on Days 3, 7, 14, and 28 post-exposure, when the samples were either fixed for imaging, or cellular lysates prepared and frozen at −80°C until biochemical/molecular analysis. Exposure of the aggregates to air blast did not have any obvious effects on morphology immediately after exposure or during the next 28 days. The aggregates retained their spherical shape and no or little debris was evident.
No cell death was noted at any time–point after shock wave exposure (Fig. 4A, 4B). Sham control caspase-3 activity was low and no consistent pressure-dependent trends in apoptotic cell death were noted at any time–point (Fig. 4A). Similarly, LDH release into the medium showed no pressure-related changes, compared with sham controls, at any time–point (Fig. 4B). The lower absorbance values of the sham controls as the cultures aged post-treatment are likely due to the sequential harvest of material from the cultures. The effects of air blast on the structural proteins GFAP and NFH are shown in Figure 4C and 4D. No statistically significant changes in either the astroglial marker GFAP (Fig. 4C) or the neuronal marker NFH (Fig. 4D) as a function of pressure or time were observed, and levels remained at or near baseline. This also was confirmed by confocal imaging of aggregates exposed to primary blast and assessed at 28 days post-exposure (Fig. 5). The astroglial GFAP was localized around the perimeter of the aggregates, while NFH was concentrated towards the interior. No changes due to blast exposure were observed in either the levels or distribution of NFH or GFAP, compared with sham controls.
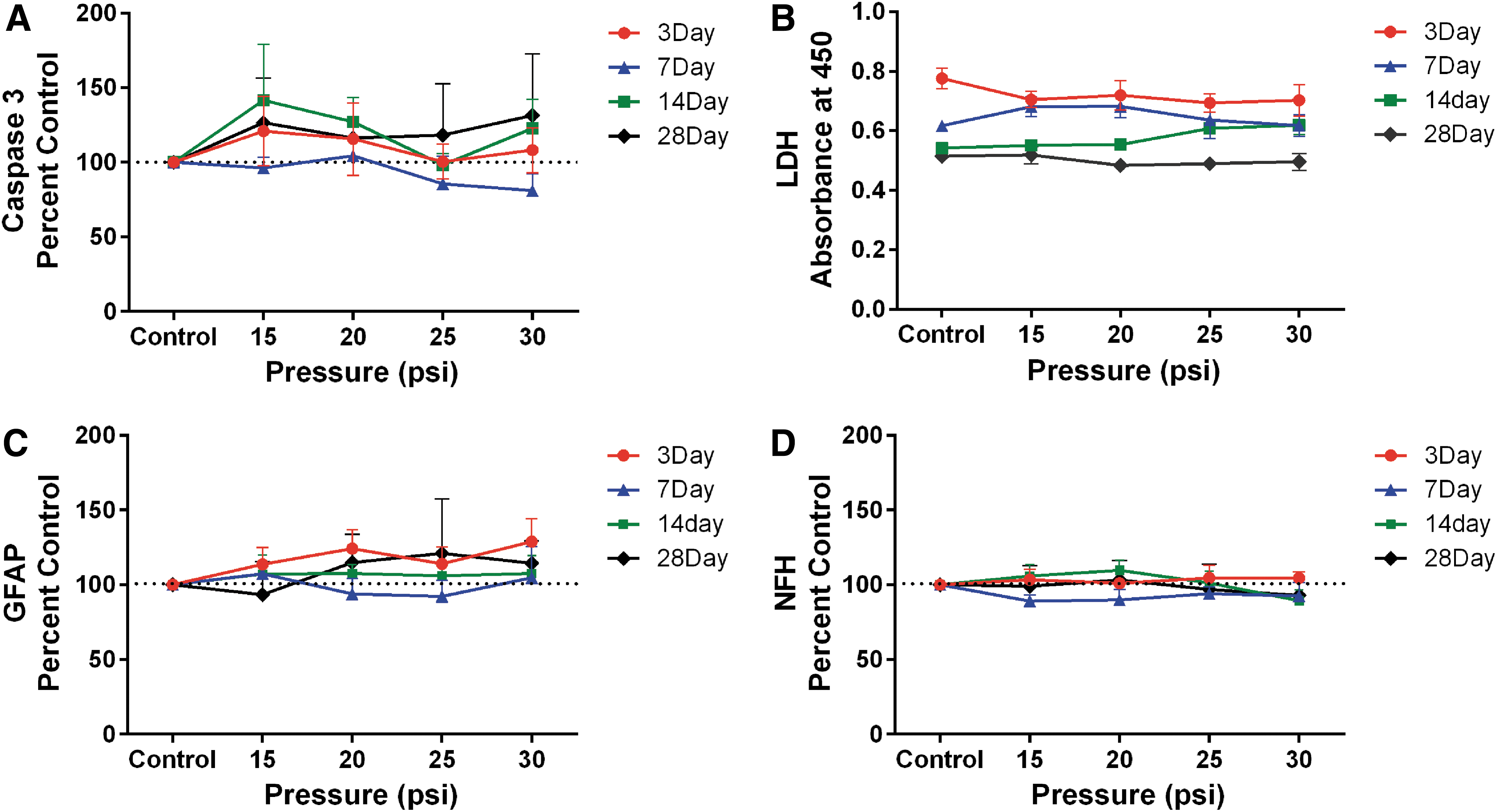
Effect of primary blast on lactate dehydrogenase (LDH) release, caspase-3 activity, neurofilament H (NFH), and glial fibrillary acidic protein (GFAP) levels in rat brain aggregates. Brain cell aggregates were exposed to 15, 20, 25, or 30 psi primary blast, then placed back into culture. Cultures were assessed for either apoptotic cell death using caspase-3 activity
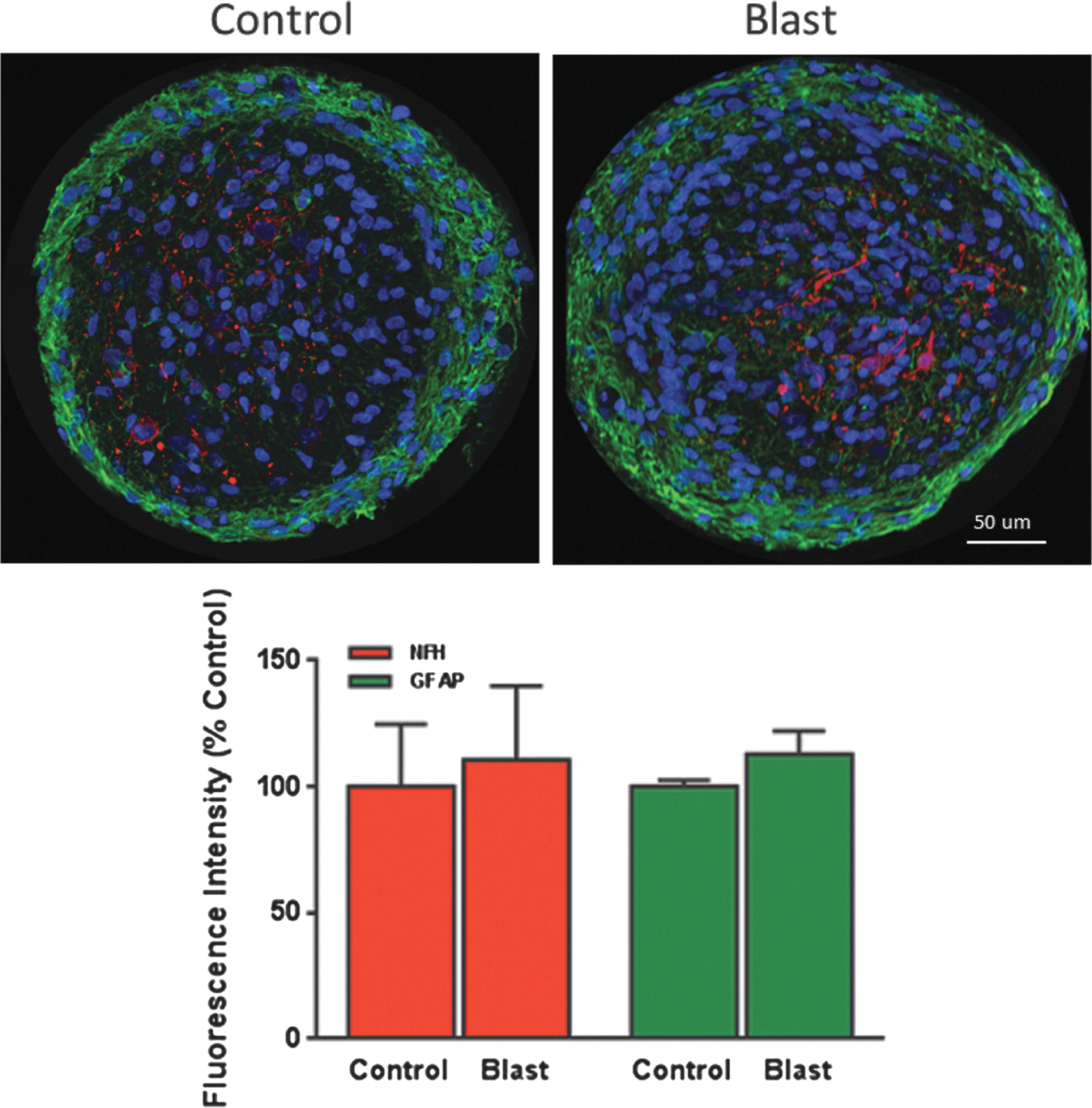
Effect of primary blast on glial fibrillary acidic protein (GFAP) and neurofilament H (NFH) expression in rat brain aggregates. Top panel: Representative confocal images showing the expression of GFAP (green) and neurofilament H (red) in either sham control or aggregates exposed to primary blast (25 psi). Magnification: 400 × . Bottom panel: Quantification of NFH/GFAP fluorescence intensity, expressed as percentage of control. No significant differences in intensity levels between sham control or blast exposed aggregates were observed. Values represent the mean ± standard error of the mean from four separate trials.
The effects of air blast on the activity of the cholinergic neuron enzyme markers AChE and ChAT and the astrocytic marker GS are depicted in Figure 6A-6C. Sham control AChE activities did not change significantly from Day 3 to Day 28 after blast (3.27 ± 0.46 vs. 4.04 ± 1.01 nmoL/mg/min; x ± SD, n = 4), and no significant pressure-related changes were observed at any time–point. No appreciable changes in sham control ChAT values were observed (9.60 ± 1.66 vs. 10.40 ± 0.93 nmoL/mg/min; x ± SD, n = 4) during the time course, and simulated air blast produced no change in enzyme activities at any pressure or time–point. Sham control values for the glial enzyme marker GS were 91.1 ± 33.0 nmoL/mg/min (Day 3), compared with 101.8 ± 39.0 nmoL/mg/min on Day 28 (x ± SD, n = 4), while shock wave exposure did not induce any changes at any time-points after exposure (Fig. 6C).

Effect of primary blast on acetylcholinesterase (AChE), choline acetyltransferase (ChAT), and glutamine synthetase (GS) enzyme activities. Brain cell aggregates were exposed to 15, 20, 25, or 30 psi primary blast, then placed back into culture. Cultures were assessed for changes in AChE
Figure 7 shows the effect of simulated air blast on the Akt cell signaling pathway. Exposure to blast did not significantly change the total levels of Akt at any pressure or time–point (Fig. 7B). In contrast, primary blast induced a significant elevation in the phosphorylated form of this protein at 3 days post-exposure at all pressures (15–30 psi; Fig. 7A). This protein activation was attenuated by Day 7 so that increased levels were observed at only the two highest pressures. By 14 days post-exposure, no increases in Akt phosphorylation were noted and by 28 days, there were indications that phosphorylation was decreased, compared with sham controls. Representative blots of these results are depicted in Figure 7C.

Effect of primary blast on brain aggregate protein kinase B (Akt) levels. Brain cell aggregates were exposed to 15, 20, 25, or 30 psi primary blast, then placed back into culture. Cultures were assessed for changes in phosphorylated Akt (p-Akt;
The effects of simulated blast on the oligodendrocyte enzyme marker CNPase are shown in Figure 8A. Sham values were 393.8 ± 142.8 nmoL/mg/min on Day 3, compared with 291.7 ± 244.1 nmoL/mg/min on Day 28 (x ± SD; n = 4). At 3 and 7 days post-exposure, no elevation in enzyme activity was noted. By 14 days, a slight but statistically insignificant trend of increasing activity was evident. At 28 days post-shock exposure, a pronounced pressure-dependent increase in CNPase activity was observed up to approximately 300% of control values, with the enzyme activities at the two highest pressures being statistically significantly different from control values. Simulated blast exposure also modulated the levels of VEGF in a delayed fashion (Fig. 8B). No significant changes were noted in VEGF levels at 3 or 7 days post-exposure. However, at 14 days, VEGF levels showed a trend towards lower values. By 28 days, pressure-dependent and statistically significant declines in VEGF levels were observed.

Effect of primary blast on brain aggregate 2′,3′-cyclic nucleotide 3′-phosphodiesterase (CNPase) enzyme activity and vascular endothelial growth factor (VEGF) levels. Brain cell aggregates were exposed to 15, 20, 25, or 30 psi primary blast, then placed back into culture. Cultures were assessed for changes in CNPase enzyme activity
Discussion
The results of this study demonstrate that primary blast exposure causes not only immediate changes in brain function endpoints, but also produces much delayed effects in brain tissue. When compared with previous work using this same model system, 41 it also shows that brain tissue can discriminate between different types of pressure insult with different patterns of protein expression, notably with respect to those associated with cell survival and neurodegeneration. This work has implications for the study of primary blast and also highlights the importance of rigorously defining exposure conditions during such studies.
The effect of blast-induced shock waves on the brain has been the subject of considerable controversy due to the difficulties of definitively identifying clinical cases caused by this specific insult, 48 –50 as well as the technical difficulties involved in simulating primary blast in the laboratory. 6,11 –13 While rodents would appear to be a logical route with which to study PbTBI, they present their own unique set of technical challenges. These are significant, notably with respect to the unknown scaling factors that arise due to the dynamic pressure artifacts introduced in experiments using small rodents, compared with those using targets the size of the relatively large and heavy human head. 11 –13 In such rodent studies, the relative contributions of primary blast are difficult to separate from those arising from the exaggerated global head motion produced during either simulated or free-field blast exposure.
Tissue culture platforms offer alternatives that reduce some of the complexity inherent in small animal studies of primary blast, and a number of laboratories have developed tissue culture models utilizing primary cell cultures or established cell lines. 16,17,24,25,28 –30,32,33 In these model systems, the culture vessel (single dish or multi-well) containing the surface-cultured cells is exposed to the blast insult. While the cells in these studies assuredly experience pressure changes as a result of their exposure, the multiple and complex boundary conditions imposed by using monolayer cultures on plastic or glass surfaces and in culture vessels make the assessment of cause–effect relationships difficult. Other laboratories have developed model systems using organotypic hippocampal slice cultures (OHSCs) 18 –20,26,27,31 or endothelial cells 19,21 –23 that are cultured on membranes. The membranes are exposed in instrumented barochambers and appear not to influence the nature of the pressure wave exposure, with similar overpressure measurements obtained both above and below the sample, 19 thus enabling an accurate assessment of exposure. Notably, electrophysiological studies using OHSCs in these experiments have shown significant attenuation of long-term potentiation without accompanying cell death after shock wave exposure. 18,31 However, whether the adherence of brain slices or cells to membranes, plastic, or glass acts to alter the biomechanical and injury response of the tissue to pressure changes is not clear.
Rat brain cell aggregate cultures are composed of the major brain cell types, show many of the same functionalities of the intact brain, and can be cultured for periods of time extending to months. They are suspension cultures and this allows for control of boundary conditions during shock or blast exposure. In previous work, this characteristic was exploited to investigate the effects of underwater blast. 41 Studies were carried out in a large underwater explosion (UNDEX) pond facility that minimized reflections. By encasing the aggregates in dialysis tubing cylinders transparent to the blast insult, the suspended cultures were exposed to highly defined, single pulse blast waves ranging in intensity from those representing a realistic threshold pressure unlikely to cause injury underwater (∼43 psi) to that likely to cause an injurious and lethal exposure to a human (∼2,071 psi). 51 While many of the brain function endpoints assessed were not altered in these experiments, phosphorylation of the cellular survival protein Akt, the oligodendrocytic marker CNPase, and VEGF all showed pressure-related changes indicating the sensitivity of this culture system to blast waves. Although the exposures in this UNDEX pond study were not realistic in the sense that brain tissue never experiences this simplistic type of pressure change due to the presence of the skull, this work did show that the model system was sensitive to a specific pressure insult consisting of a simple and well-defined principal stress without any shear.
It currently is not known which shock wave characteristics (i.e., rise-time, peak amplitude, decay rate, impulse, stress wave duration) are of importance in determining the severity of brain injury in an exposed individual, nor is it well understood how brain/skull interactions impact this damage. While tissue culture models may well represent a simplified platform on which to base studies to answer these unknowns, it is not sufficient that they simply respond to pressure changes; they also must be able to discriminate between different types. As a next step towards validating the use of aggregates as a model system, studies were undertaken to ascertain their response to a much more complex pressure insult, compared with the previous UNDEX work. 41 These studies utilized an ABS and exposed aggregates in suspension and encased within a polypropylene sphere to simulated air blast. Initial experiments assessed the effects of simulated blast on the pressure signatures within the spheres. Although the sphere exposures consisted of highly defined single pulse shock waves, measurements made inside the water-containing spheres showed that the actual pressure changes were highly complex and spatially dependent. Thus, the brain tissue in this study experienced complex pressure changes that were dependent on their position in the sphere. Further, since the aggregates were not segregated in any way after exposure, the measurements of their response was an average of all responses across the sphere. This is in marked contrast to the UNDEX work, where all aggregates were exposed to the same highly defined single pulse shock wave.
The response of the aggregates in the sphere to primary air blast was similar in many respects to that exhibited by aggregates exposed underwater to single pulse shock waves. No cell death was observed using the cell membrane integrity indicator LDH. Although this enzyme's release into medium as a measure of cell death is quantitative when standardized against constants such as total protein, DNA, cell number, or medium volume, this culture model does not facilitate this due to its suspension nature and frequent medium changes. Nevertheless, this assay does provide a reliable indicator of the relative viability of test samples compared with similarly treated control samples, and has been utilized successfully when assessing the time and concentration dependence of the toxicity of agents such as sulphur mustard (unpublished observations). Caspase-3 activation as a marker of apoptotic cell death also was not observed. These findings are largely consistent with several literature reports where cell death was either not detected, was modest, or occurred only at relatively high pressure insults. 18 –20,30 –33 However, several studies, notably those using 96 well culture plates, have reported significant declines in cell survival markers. 16,17,28,29 In one such model system, injury was observed only after decreasing the volume of the culture medium in the well and inducing measurable shear, 28,29 suggesting that gross fluid motion, rather than primary blast effects, may be responsible for the injury. Similarly to the UNDEX study, cell type–specific structural proteins in the aggregates were unaffected by primary blast, with neither NFH nor GFAP showing changes in their levels or their distributions. The astroglial protein GFAP is a prototypical biomarker of acceleration impact–induced TBI 52 –54 and has also been used as a biomarker of PbTBI, 55 –60 although there are reports that this protein may be not be a reliable indicator of this type of brain damage. 12 Ravin and coworkers 28,29 reported elevations in this biomarker in human central nervous system cells exposed to elevated pressure (and shear), as does a recent article describing astrocyte activation in an OHSC model exposed to air blast, 26 findings that were not duplicated in a similar OHSC model system using barochamber exposures. 20 Cell-specific enzyme markers, including the cholinergic neuronal markers AChE and ChAT and the astrocytic marker GS, also showed no alterations in their specific activities in aggregates exposed to either air blast or underwater blast. The findings that primary blast does not cause overt cellular damage in brain cell aggregates is not overly surprising if it is representative of PbTBI. While the role of primary blast in the mild TBI experienced by individuals exposed to blast may be controversial, it seems likely that any such effects would be subtle and not due to rapid structural protein alterations or cytotoxicity.
The activation of protein kinase B or Akt in blast-exposed aggregates occurred in a robust and pressure-dependent manner that was reminiscent of previous findings in the UNDEX studies 41 Three days after exposure to a peak overpressure of 30 psi, Akt phosphorylation levels increased by over 250%, compared with sham controls. This also was comparable to the 150–200% increase observed in head-only exposed rats exposed to similar overpressures (unpublished observations). This activation declined rapidly so that by 14 days, it was no longer statistically significant and by 28 days had fallen below baseline levels. This protein, as part of the phosphatidyl 3-kinase/Akt (PI3K/Akt) pathway, is phosphorylated in response to cellular stress or injury and plays a vital role in a broad range of cellular functions, including cell survival, proliferation, cell growth, metabolism, and migration. Its deregulation has been implicated in the development of numerous pathologies, including cancers, type 2 diabetes mellitus, and cardiovascular and neurodegenerative diseases, 61 –71 and as such, pharmacological modulation of this pathway has been the subject of much interest. 65 –67 Activation of Akt is neuroprotective, 70, 68 –71 and as in the case of aggregates exposed to UNDEX, the absence of cell death in these studies may be a result of this transient activation.
Simulated blast caused a delayed and pressure-dependent increase in the oligodendrocytic marker CNPase of up to almost 300% at 28 days. Enzyme activity prior to this was only slightly increased (14 days) or baseline (3 and 7 days). This was in marked contrast to the effect of underwater single pulse shock waves on brain aggregates, where a modest increase (∼150%) was observed at only 7 days post-exposure at the highest pressure. 41 This protein plays a role in myelination and its catalytic activity also has recently been identified as possibly playing a role in neuroprotection through hydrolysis of the toxin adenosine 2′,3′-cyclic monophosphate (2′,3′-cAMP). 72 –74 Changes in CNPase levels also have been linked to neurodegenerative diseases such as Alzheimer's disease, Down's syndrome, and catatonia-depression syndrome. 72,73
The effect of simulated blast on VEGF levels in these studies was more dramatic than in the previous UNDEX work, where significant inhibition of this protein occurred only at one pressure at 7days and then returned to baseline levels. 41 In contrast, in the sphere work, VEGF levels did not change before 14 days post-exposure, when a pressure-dependent decline was observed. This decrease became much more substantial and statistically significant by 28 days post-exposure, with peak inhibitions of ∼50%. This protein was originally identified as an endothelial growth factor that also increased vascular permeability, 75 leading to it also being known as vascular permeability factor. The significant roles that VEGF plays in angiogenesis, as well as in inflammation and in cancers, has made inhibition of this pathway an extremely attractive target in cancer therapy, 76 –78 as well as in other vascular diseases. 79 –81 Additional research has shown several subtypes of VEGF to occur in a number of other tissue types, notably in the central nervous system. Binding of VEGF to VEGF receptors activates the PI3K/Akt signal transduction pathway and leads to the inhibition of apoptotic cell death. 82 –85 VEGF has been implicated in several neurodegenerative diseases and has been shown to have neuroprotective properties. 85 –89 The effects of blast on Akt and VEGF in these studies shows that the initial activation of Akt is clearly not through its upstream regulator VEGF. The effects of blast on Akt and VEGF warrant further investigation, especially given the great research interest in these two proteins as therapies for a host of neurodegenerative diseases.
In summary, this work has shown that rat brain cell aggregates exposed to complex pressure histories respond in a pressure-dependent fashion with both early, as well as much delayed changes in proteins implicated in cell survival and neurodegeneration. This response was in contrast to that obtained in earlier work using underwater blast where the pressure changes experienced by the aggregates were single pulse in nature. 41 While in both cases Akt activation occurred at early time–points and then returned towards baseline values, the temporal pattern of VEGF inhibition and CNPase enzyme levels were markedly different. Exposure to the UNDEX-induced single pulse blast waves caused CNPase elevation and VEGF inhibition at 7 days that then returned towards baseline levels at longer time-points. In contrast, in response to the complex pressure changes experienced by aggregates encased in a sphere and exposed to simulated air blast, significant CNPase enzyme level increases and VEGF inhibition occurred only after a delay of 14 to 28 days.
The differences in aggregate VEGF and CNPase responses to the UNDEX versus the ABS/sphere-enclosed exposures cannot be directly compared statistically due to the markedly different pressure history exposures. Comparison of the ABS/sphere exposure pressure conditions depicted in Table 1 with those derived from the previously published UNDEX studies (Table 1) 41 reveal no obvious explanations (overpressure, impulse, duration) for the temporal differences in the VEGF and CNPase effects. Variables such as time out of routine culture conditions (∼15–25 min in both studies), changes in temperature, and differences in handling procedures and exposure vessels (dialysis tubing vs. polypropylene spheres) were closely controlled for by using sham controls (which showed no adverse effects). Thus, it seems reasonable to conclude that the temporal differences in VEGF and CNPase responses between the two studies is due to the added complexity of the pressure changes imposed by the sphere after blast exposure.
The exposure of aggregates to UNDEX blast in the previous study represented an artificial situation where brain tissue was directly exposed to highly defined high-pressure (43–2,071 psi) short-duration (0.12–0.43 msec) single pulse blast waves without shear. The current study is much more relevant to what would happen to the head of an individual exposed to low-level blast. In this latter case, the sphere (head) experienced the diffraction of an air-borne shock wave (15–30 psi, ∼6.1–7.7 msec) imparting complex three-dimensional principal and shear stresses to the encased brain tissue. The findings from these two studies have potentially important ramifications. They demonstrate that primary blast can alter cellular proteins that also have been identified as potential targets of pharmacological intervention. The delayed effects on both VEGF and CNPase are also of interest as it is consistent with the delayed mild TBI experienced by some blast-exposed individuals, suggesting a potential role for primary blast in the etiology of this poorly understood disorder. The ability of this model system to discriminate between different types of pressure change is also unique, and is necessary to facilitate future work to elucidate what blast wave characteristics are of importance in determining cellular response and injury. This is a potentially invaluable aid to the development of personal protection, and also highlights the importance of accurately measuring the pressure changes experienced by target tissue in primary blast studies.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.