
Abstract
More than half of all spinal cord injuries (SCIs) occur at the cervical level, often resulting in impaired respiration. Despite this devastating outcome, there is substantial evidence for endogenous neuroplasticity after cervical SCI. Spinal interneurons are widely recognized as being an essential anatomical component of this plasticity by contributing to novel neuronal pathways that can result in functional improvement. The identity of spinal interneurons involved with respiratory plasticity post-SCI, however, has remained largely unknown. Using a transgenic Chx10-eGFP mouse line (Strain 011391-UCD), the present study is the first to demonstrate the recruitment of excitatory interneurons into injured phrenic circuitry after a high cervical SCI. Diaphragm electromyography and anatomical analysis were used to confirm lesion-induced functional deficits and document extent of the lesion, respectively. Transneuronal tracing with pseudorabies virus (PRV) was used to identify interneurons within the phrenic circuitry. There was a robust increase in the number of PRV-labeled V2a interneurons ipsilateral to the C2 hemisection, demonstrating that significant numbers of these excitatory spinal interneurons were anatomically recruited into the phrenic motor pathway two weeks after injury, a time known to correspond with functional phrenic plasticity. Understanding this anatomical spinal plasticity and the neural substrates associated with functional compensation or recovery post-SCI in a controlled, experimental setting may help shed light onto possible cellular therapeutic candidates that can be targeted to enhance spontaneous recovery.
Introduction
T
One of the best examples of respiratory neuroplasticity has been demonstrated after high cervical (C2) lateral hemisection (Hx), with the return of ipsilateral phrenic motoneuron (PhMN) and diaphragm activity either induced 1,3 under terminal neurophysiological conditions shortly after injury (crossed phrenic phenomenon, CPP) or occurring spontaneously weeks post-injury (spontaneous CPP, sCPP). While both types of plasticity have been attributed to spared (uninjured) mono- and polysynaptic bulbospinal pathways within the spinal cord contralateral to the injury, the contribution of spinal interneurons to recovery remains poorly defined.
One potential interneuron aiding in respiratory plasticity is the V2a class of spinal interneurons (identifiable by transcription factor Chx10 expression). The V2a interneurons located in the medial reticular formation directly project onto respiratory neurons in the pre-Bötzinger complex, and genetic ablation of these interneurons results in irregular breathing. 4 In addition, V2a interneurons in the brainstem and cervical spinal cord have been demonstrated to have a crucial role in the recruitment of inspiratory accessory respiratory muscles as a form of compensatory plasticity that maintains ventilation at early stages of amyotrophic lateral sclerosis (ALS). 5 Previous studies have demonstrated anatomical recruitment, albeit not statistically significant when looking at a heterogeneous interneuronal population, of cervical interneurons after a C2 SCI. 6,26 Cross-correlation analyses from phrenic nerve recordings have supported this, suggesting that spinal interneurons could be functionally recruited into injured phrenic pathways weeks post-C2Hx.
The objective of the present study was to examine the distribution and potential anatomical recruitment of a subset of spinal interneurons only recently identified as playing a role in respiratory plasticity. 5 Our study reveals the involvement of spinal Chx10-V2a interneurons in anatomical plasticity after a C2Hx. Using a retrograde, transneuronal tracer (pseudorabies virus), we demonstrate anatomical recruitment of Chx10 interneurons into the phrenic circuit two weeks after injury. These experiments provide the first evidence for the involvement of a specific population of spinal interneurons in anatomical plasticity post-SCI, capable of modulating phrenic motor output and functional plasticity after SCI.
Methods
All experimental procedures were conducted in strict accordance with the guidelines set by the Drexel University Institutional Animal Use and Care Committee and the Institute of Laboratory Animal Resources, U.S. National Academy of Sciences. Adult Chx10-eGFP (green fluorescent protein) transgenic mice (Strain 011391-UCD) obtained from Mutant Mouse Regional Research Centers (MMRRC University of California, Davis, CA) were created using a bacterial artificial chromosome on a FVB/N- Swiss Webster hybrid background strain and used in all experiments. A total of 22 mice (naïve, uninjured n = 8; C2Hx, n = 14) were used for this study. Of those, three mice were used for terminal electrophysiological recordings immediately after C2Hx and five mice survived the C2Hx injury to receive terminal electrophysiological recordings two weeks post-injury.
All operations were performed under isoflurane anesthesia (induction 5%; maintenance 2%, in 100% O2 at 2 L/min) on a water-circulating heating pad. Upon recovery, animals received buprenorphine analgesic (subcutaneous [sc] at 0.03 mg/kg) and warmed lactated Ringer's solution (1 mL sc to prevent dehydration). In those animals receiving C2Hx (n = 14), an incision was made extending from the base of the skull to the third cervical segment (C3), and a laminectomy was made at the second cervical segment (C2). A dural incision was then made, and a lateral Hx was performed on the left side of the spinal cord by inserting a No. 11 scalpel blade through the midline of the dorsal columns ventrally to the underlying vertebral column, drawing the blade laterally and angling the tip outward so that it followed the underlying vertebrae, transecting the lateral funiculus. This approach was repeated three times on the left side of the spinal cord. Unlike our previous experience with C2Hx in the adult rat, 7 –9 the dura was not sutured immediately post-injury. Overlying muscle was sutured in layers, and the skin was closed with wound clips. Extent of the lesion was confirmed via post-hoc histochemical analysis.
Injured (n = 11) and naïve Chx10-eGFP (n = 8) mice were traced with a transsynaptic, retrograde pseudorabies virus tracer (PRV614; a PRV Bartha promoting mRFP expression). Animals were anesthetized as described above, a laparotomy was performed to expose the diaphragm, the liver gently moved from the muscle surface, and PRV614 was topically applied to the left hemidiaphragm (50 μl, 2.0 × 108 plaque-forming units [pfu]/mL) as described previously for both rat 6 and mouse. 10 Animals were allowed to survive for 72 h before undergoing terminal, bilateral, diaphragm electromyography (dEMG).
Terminal bilateral dEMG recordings were made immediately (n = 3) or two weeks post-C2Hx (n = 5), or 72 h post-PRV (n = 8) in age-matched naïve control animals, just before being euthanized and perfusion-fixed for histological analyses. Animals were anesthetized with a mixture of xylazine hydrochloride and ketamine hydrochloride (9 mg/kg sc and 72 mg/kg intraperitoneally, respectively). A laparotomy was then performed along the incision made during PRV-tracing to re-expose the abdominal surface of the diaphragm. Bipolar hook electrodes (paraformaldehyde [PFA]-coated tungsten wire with exposed tips, A-M Systems, Cat # 796500) were then placed into the medial costal region of the left and right hemidiaphragm as described previously. 11 Activity was recorded during spontaneous breathing, and baseline was counted as a minimum of 10 min of stable activity. The dEMG signals were amplified (1,000 × ) and band pass filtered (0.3-10KHz) using differential A/C amplifier (Model 1700, A-M Systems) and digitized (Power 1401, Cambridge Electronic Design). Recordings were analyzed using Spike 2 and OriginLab 9 software as described previously. 11
At the end of each experiment, all animals were deeply anesthetized by an overdose of ketamine, intracardially perfused with saline (0.9% NaCl in water) and paraformaldehyde (PFA; 4% w/v in 0.1 M phosphate buffered saline [PBS], pH 7.4), and the spinal cord immediately dissected and stored in PFA. For cell counts, tissue was sectioned in the horizontal plane (20 μm, on slide), and all sections were stained for chicken-GFP (1:750, Abcam) and Rabbit-PRV (1:10,000, courtesy of Lynn Enquist, Princeton University) with anti-chicken Alexa Fluor 488 (1:200; Jackson ImmunoResearch Laboratories, Inc.) and anti-rabbit Alexa Fluor 594 (1:200, Jackson ImmunoResearch Laboratories, Inc.). For lesion reconstruction, tissue (n = 3) was sectioned in the horizontal plane (20 μm, on slide), histologically processed with cresyl violet, and every section was used to create an average overlay of the lesion created by the injury. For characterization of the transgenic line, tissue was sectioned in the transverse plane (20 μm, on slide) and stained for sheep-Chx10 (1:200, Millipore), with anti-sheep Alexa Fluor 594 (1:200, Jackson ImmunoResearch Laboratories, Inc.). Sections were photographed and examined using a Zeiss M2 AxioImager microscope.
PRV-positive PhMNs, PRV positive spinal interneurons (SpINs), and PRV/GFP double-labeled cells with visible nuclei were counted at 10 × magnification in every longitudinal cervical spinal cord section from each animal sectioned longitudinally as described previously. 6,11 The corrected number (Abercrombie correction factor) of PhMNs and PhINs are reported as averages for post-injury and naïve groups ± standard deviation.
Anatomical and electrophysiological results were quantified using Microsoft Office Excel 2016, Spike 2 (version 8, Cambridge Electronic Design, UK) and OriginLab9 software (Northampton, MA). Average numbers are shown ± standard deviation. Statistical analyses were performed using SPSS (IBM SPSS Statistics 23). Comparison between injured and naïve groups was made using analysis of variance and Student t test (IBM SPSS Statistics 23).
Results
Histochemical analysis using cross sections allowed for the average reconstruction of the lesion site as depicted in Figure 1A. The extent of the lesion included severed dorsal funiculus (Fig. 1Ai), intermediate gray (Fig. 1Aii), and ventral gray and white matter (Fig. 1Aiii, Aiv) on one side of the spinal cord. The C2Hx resulted in the silencing of diaphragm activity on the side of the injury, immediately after injury (Fig. 1C) compared with activity before injury (Fig. 1B) as measured via bilateral diaphragm EMG. Some activity appears at two weeks post-injury on the previously silenced ipsilateral hemidiaphragm (Fig. 1D). Output on the contralateral hemidiaphragm remained unchanged within 10 min of C2Hx (Fig. 1C'), compared with before injury (Fig. 1B'), but was altered two weeks post-injury, displaying a slight increase during late-inspiratory bursting (Fig. 1D').
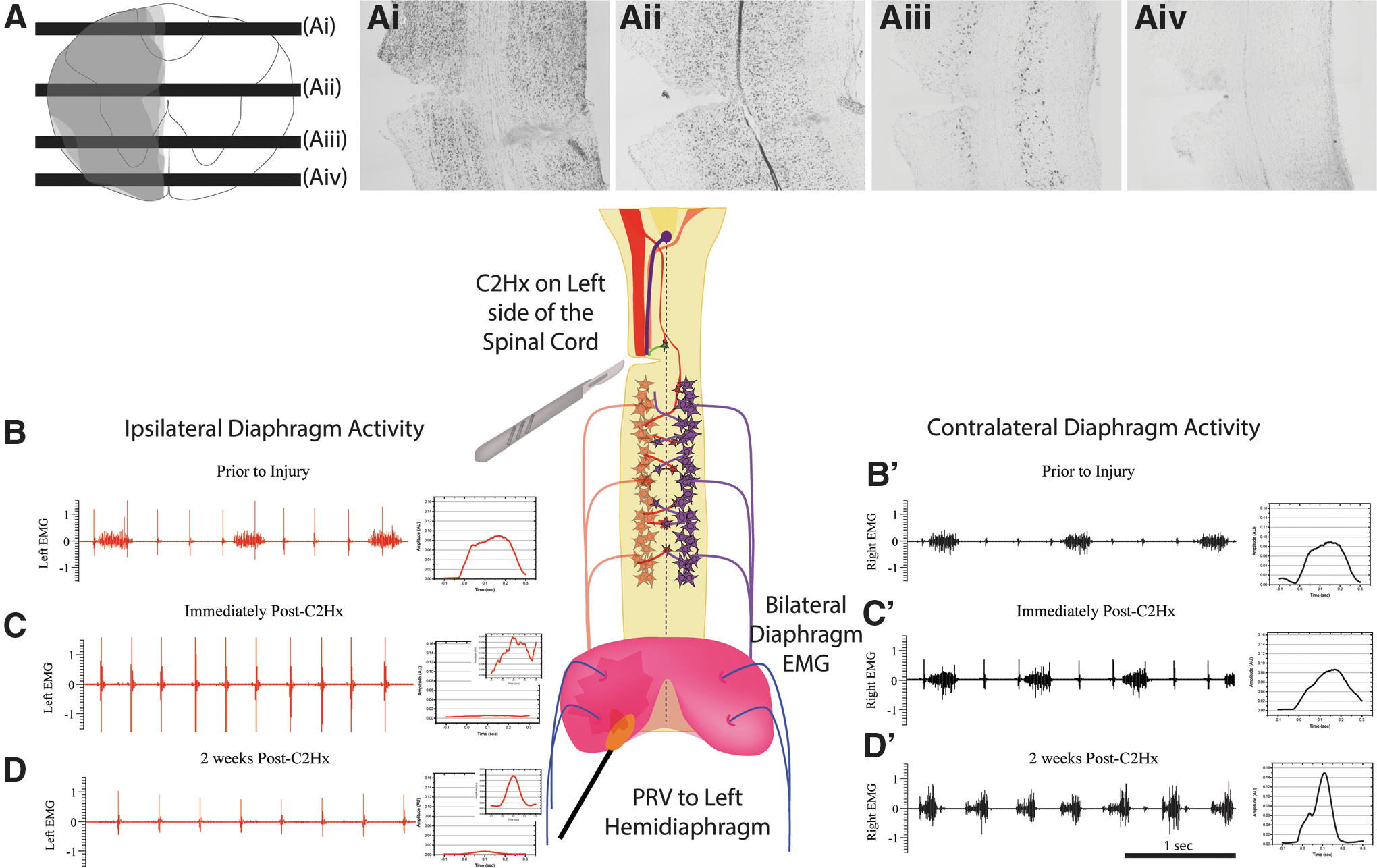
Schematic diagram depicting the experimental design and representative diaphragm electromyography (EMG) recordings from before and after injury. Cresyl violet staining was used to assess lesion severity using the described surgical technique, with an average overlaid trace depicted in (
Two weeks after C2Hx, burst frequency (168 ± 8 breaths per min) increased compared with naïve mice (98 ± 7 breaths per min). Average amplitude of diaphragm EMGs was not significantly different between left and right sides in naïve animals (0.10 ± 0.05 AU; 0.10 ± 0.05 AU left and right, respectively), whereas injury-induced deficits were observed two weeks post-injury (0.0050 ± 0.0021 AU; 0.062 ± 0.0052 AU left and right, respectively). No statistically significant difference in ipsilateral-to-injury diaphragm activity was observed in C2Hx animals two weeks after injury compared with activity immediately post-injury. Average contralateral output was not significantly different from that seen in uninjured animals.
Immunohistochemical analysis revealed the distribution of Chx10-eGFP interneurons in the naïve transgenic mouse to be within the brainstem, dorsal horn, and medial and ventral intermediate gray of the cervical spinal cord, similar to that which has been demonstrated in other (lumbar) regions of the spinal cord (Fig. 2A,B). 4,12 The GFP-positive cells in the dorsal horn were tightly packed and had similar morphology to cells found in substantia gelatinosa with small (10–15 microns in diameter) cell bodies (Fig. 2E). Triple labeling for Chx10, GFP, and PRV was performed to confirm the distribution of Chx10 positive cells, revealing that the GFP positive cells in the dorsal horn were negative for Chx10 immunolabeling (Fig. 2F,G) as reported previously with this transgenic model. 12 GFP cells in intermediate gray were larger (20–30 microns in diameter) located primarily throughout laminae VII and X (Fig. 2). In contrast to the GFP cells found in the dorsal horn, GFP-positive cells located in intermediate gray also are co-labeled with Chx10 antibody and thus are Chx10 interneurons (Fig. 2A,B,D). These cells have bipolar morphology with rostrocaudally projecting dendrites in regions closer to laminae VII and decussating neurites in cells closer to laminae X.
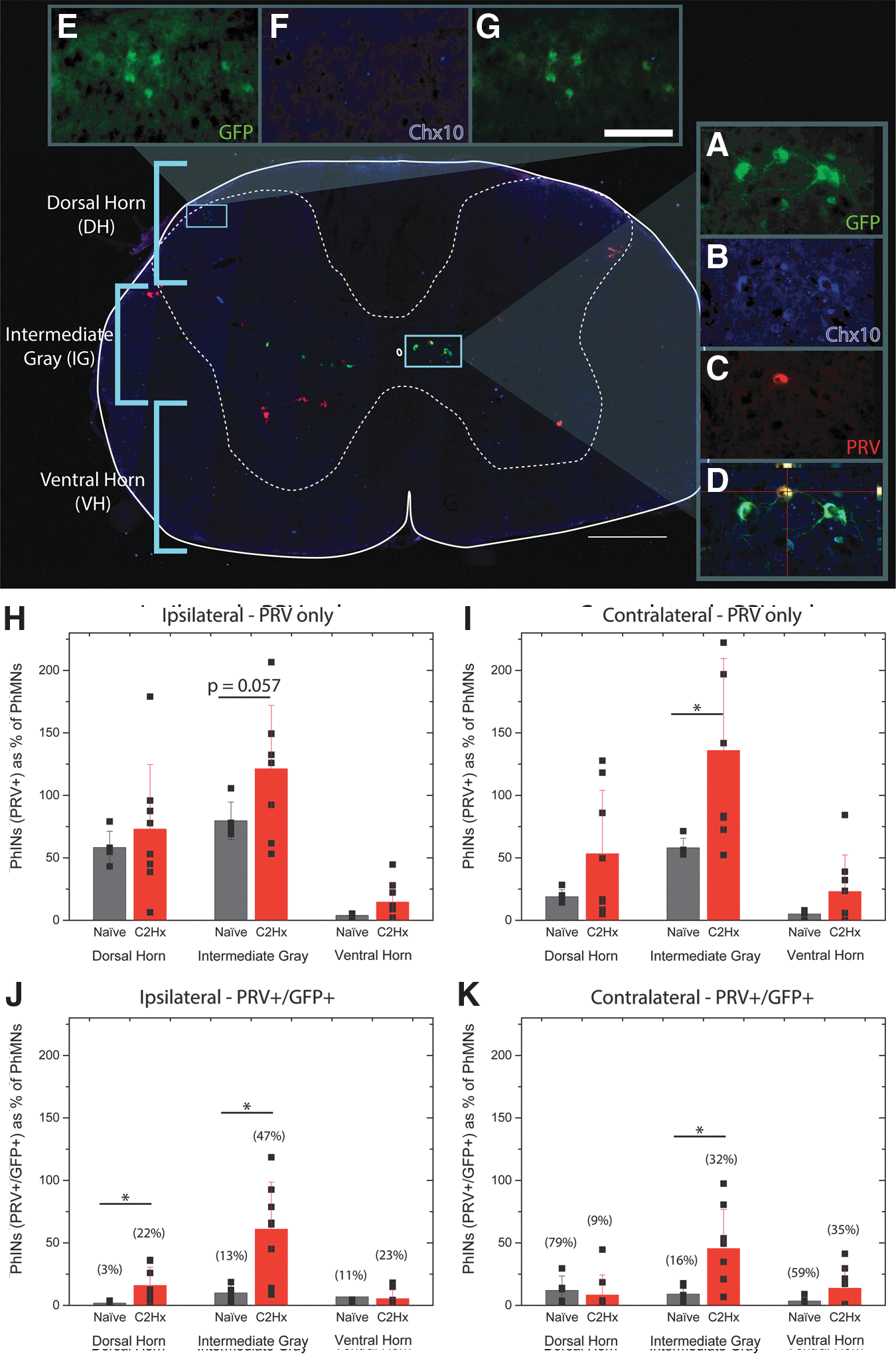
Immunohistochemical analysis revealed the recruitment of Chx10-eGFP (green fluorescent protein) interneurons into phrenic circuitry, two weeks after high lateralized spinal cord injury. An example of a cross section labeled with GFP (green), Chx10 (blue), and transneuronal pseudorabies virus (PRV) (red), showing the distribution of GFP/Chx10 cells (located in intermediate gray,
The PRV tracing of the phrenic circuit associated with ipsilateral hemidiaphragm in the naïve animal revealed a distribution of PhMNs and interneurons similar to that which has been demonstrated by Qui and associates in 2010, 10 in which robust labeling of cell bodies was seen in the phrenic motor nucleus (C3–C6), interneurons localized to lamina VII and X, and brainstem neurons throughout the caudal medulla. PRV-positive interneurons were found in the dorsal horn and intermediate gray on both sides (ipsi- and contralateral to the left hemidiaphragm) of the naive spinal cord (Fig. 2H,I, black bars). Similar to the naïve spinal cord, PRV labeling was observed ipsi- (Fig. 2H, red bars) and contralateral (Fig. 2I, red bars) to C2Hx injury.
A significantly greater number of PRV positive interneurons were seen in the contralateral intermediate gray matter, both rostral and caudal to injury (Fig. 3B,D, respectively). It is important to note that as has been shown in other models post-SCI, 6,11,13 less PhMN labeling was observed in the C2Hx animals (73 ± 24 PhMNs) compared with uninjured mice (195 ± 12 PhMNs). To account for this, all phrenic interneuron counts are represented as a percentage of PRV positive PhMNs (Fig. 2H–K, 3A–H).
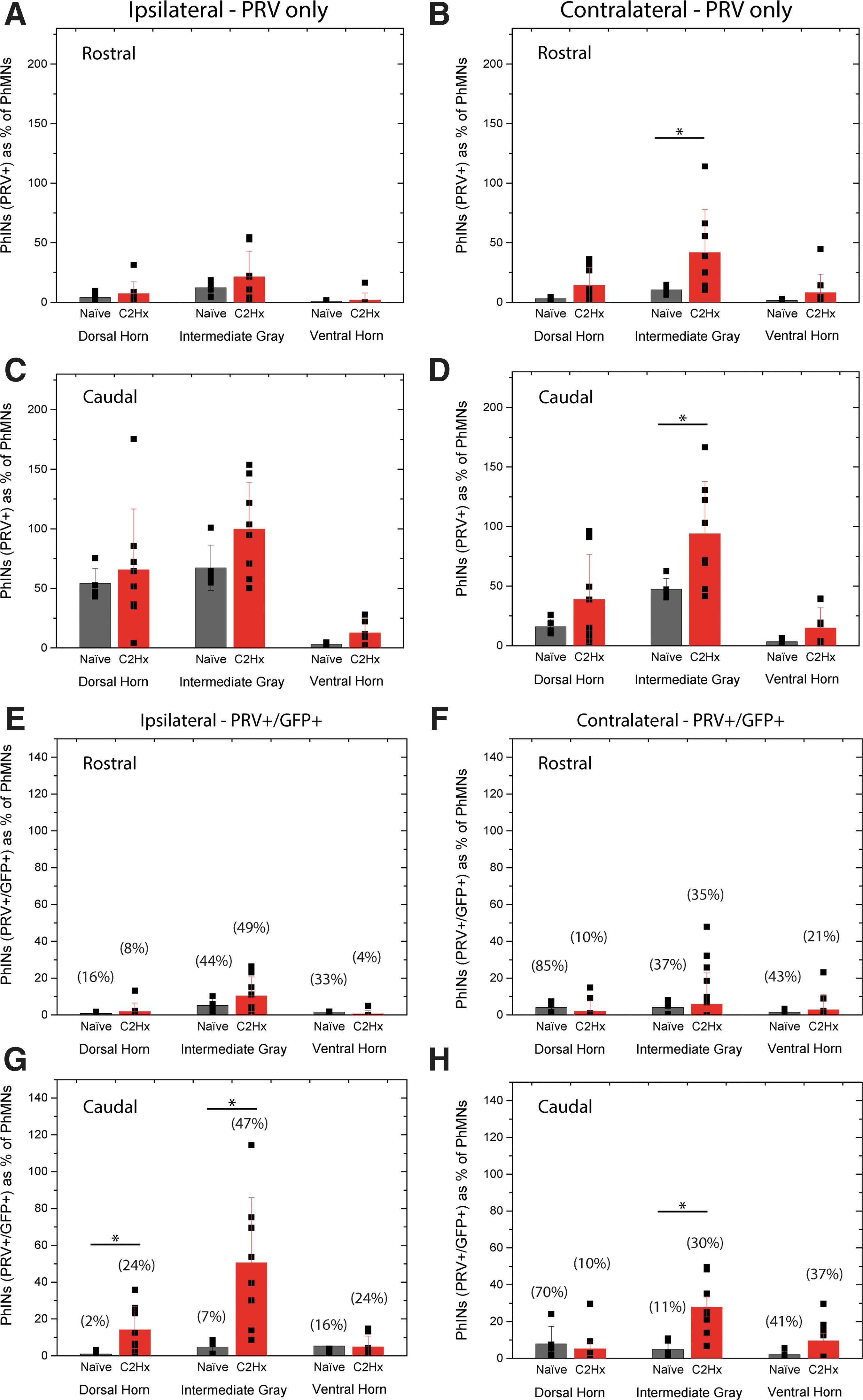
Analysis of transneuronal pseudorabies virus (PRV) positive and PRV/GFP (green fluorescent protein) dual-labeled cells revealed differences in the rostrocaudal distribution of recruited V2a interneurons two-weeks after cervical hemisection (C2Hx). The number of PRV positive interneurons significantly (p < 0.05) increased on the contralateral side of the spinal cord, both rostral (
Double-labeled spinal interneurons (SpINs) were counted and separated based on their rostrocaudal and dorsal-ventral location in the spinal cord. The separation criteria are visually represented in the cross section of the transgenic mouse spinal cord (Fig. 2). For quantification purposes, “dorsal” region was defined encompassing laminae I–V, to the level of the lateral spinal nucleus. “Intermediate gray” comprised laminae VI, VII, and X, determined by tissue containing gray matter immediately below the lateral spinal nucleus, to the presence of thin ventromedial white matter but absence of larger (40–50 microns in diameter) neurons (e.g., putative motoneurons), and absence of a distinct PhMN pool. “Ventral” was defined as remaining gray matter (and laminae VIII and IX, which includes the motoneuron pools (thus PhMNs).
Surprisingly, few PRV/GFP double-labeled cells were observed throughout the cervical spinal cord in naïve animals (ipsilateral: dorsal horn = 2 ± 1 cells, intermediate = 10 ± 6 cells and ventral gray = 7 ± 13 cells; contralateral: dorsal horn = 12 ± 11 cells, intermediate = 9 ± 7 cells and ventral gray = 4 ± 3 cells; Fig. 2 J,K, black bars). In C2Hx injured animals, however, after PRV-delivery to the side of injury, robust double labeling was observed bilaterally (ipsilateral: dorsal horn = 16 ± 14 cells, intermediate = 61 ± 37 cells, and ventral gray = 5 ± 3 cells; contralateral: dorsal horn = 8 ± 16 cells, intermediate = 46 ± 31 cells, and ventral gray = 14 ± 16 cells;) (Fig. 2J,K). Analysis of the rostrocaudal distribution of PRV/GFP double-labeled cells revealed significant increase in the ratio of interneurons caudal to C2Hx on both the ipsilateral (Fig. 3G) and contralateral (Fig. 3H) sides of the spinal cord. The number of PRV+/GFP+ cells as a percent of PRV+ cells are reported in Figures 2 (J,K) and Figure 3 (E–H) above their respective bar graphs.
Discussion
With an increasing appreciation for the neuroplastic potential of the injured spinal cord, there is a greater need for identifying the neural components that contribute to spontaneous change. The identified anatomical components may also represent targets for harnessing and amplifying functional plasticity and promoting lasting improvement in outcome post-SCI. The goal of the present work was to study the contribution of a specific subset of excitatory spinal interneurons—the V2a interneurons—to the uninjured and injured spinal phrenic motor circuitry.
C2Hx interrupts descending bulbospinal projections to the PhMNs located from C3 to C5/C6, resulting in paralysis of the ipsilateral hemidiaphragm. While paralysis of the ipsilateral hemidiaphragm was first described after a complete lateral C2Hx, 3 partial C2Hx may exhibit some similarities. Vinit and colleagues 14 –16 revealed that ipsilateral paralysis occurs after partial lateral C2Hx and persists for seven days. Three months later, significant recovery was detected ipsilateral to C2Hx ( 2,14 for discussion). Similarly, we observed very limited activity from the hemidiaphragm ipsilateral to injury two weeks after injury. Despite variability in the extent of the lesion (e.g., not all injuries were complete because we observed minor medial anatomical sparing), we found no significant difference in diaphragm activity ipsilateral to injury in C2Hx animals.
Despite initial paralysis, recovery of ipsilateral phrenic activity can be elicited acutely (minutes to hours post-C2Hx) by transection of the contralateral phrenic nerve (phrenicotomy)—which unmasks the CPP (reviewed in 17 ). Recovery of ipsilateral phrenic nerve and diaphragm activity can also occur spontaneously over the course of weeks 8,9,16,18,19 to months 20,21 after C2Hx in the adult rat. Consistent with the time course of detectable diaphragm activity in the present study, this delayed recovery has been termed the sCPP. 22 It is not known at present whether different anatomical substrates contribute to the CPP and sCPP, or how the pathways recruited differ between complete and incomplete C2Hx. 7 Although still under investigation, recovery via either the CPP or sCPP has been attributed to activation of normally silent contralateral bulbospinal projections that cross the spinal midline below the level of injury. 1,2 These pathways monosynaptically 23,24 or polysynaptically (via spinal interneurons 6,22,25,26 ) innervate PhMNs ipsilateral to injury, restoring inspiratory drive and phrenic motor output. Several studies have also shown that this plasticity is regulated by serotonin, 27,28 adenosine, 29 and glutamate. 28,30,31
Although acute activation of the CPP supports the presence of an existing latent pathway, previous studies have also revealed that ongoing neuroanatomical reorganization and recruitment of polysynaptic pathways associated with the sCPP likely contribute to further changes within crossed-phrenic pathways and long-term recovery. 25,26,32 Much like that described in other motor systems, the apparent recruitment of spinal interneurons into injured phrenic motor pathways 33 –36 may be an integral part of the lasting functional change seen after SCI. The V2a interneurons are an attractive candidate population for new circuit interneurons after SCI, because they are ipsilaterally projecting glutamatergic neurons 37,38 and glutamate and receptors are reported to play an important role in recovery after C2Hx. 28,30,31 Further, they are known to receive serotonergic input, which is also closely tied to phrenic plasticity post-SCI.
The V2a interneurons within the lumbar spinal cord have been reported to regulate left-right limb alternation in locomotion. It has been shown that subsets of these spinal neurons project to commissural interneurons and motor neurons within the lumbar spinal cord, 39 having both local and long range projections. Their function in the cervical spinal cord, however, has not been well defined. Recently, studies using genetic ablation of V2a neurons revealed their involvement in respiration, although this may also occur at the brainstem level. 4 Brainstem V2a neurons are located in the medial reticular formation, sending projections to neurons in the pre-Bötzinger complex. Likely because of these brainstem and lumbar spinal cord findings, V2a neurons have also been implicated as having a crucial role for recruitment of accessory respiratory muscles (compensatory plasticity) in a mouse model of ALS. 5 The contribution of spinal V2a interneurons to the phrenic motor circuitry, however, has not been previously explored.
The present work demonstrates that V2a interneurons are not integrated (or weakly integrated) with the phrenic motor circuit within the naïve animal. They are recruited into injured pathways, however, after high cervical SCI, and thus contribute to anatomical plasticity. Given that V2a interneurons are reported to project ipsilaterally in the lumbar spinal cord, 37 the bilateral distribution of these cells associated with the PhMN pool ipsilateral to injury is of some surprise. It should be noted, however, that the transneuronal tracing method used here does not distinguish between interneurons projecting directly onto the PhMNs, or indirectly via other spinal interneurons. Thus, those neurons integrated with the phrenic circuit that are found contralaterally could project onto other interneurons with commissural projections (e.g., V0 or V3) as reported within the lumbar spinal cord (for review 39 ).
While the CPP/sCPP has been studied across a range of species for more than 100 years, 3,40 it has been characterized only recently in the adult mouse. 41 This has enabled the use of transgenic mouse models as a means for dissecting mechanisms contributing to the CPP, and breathing post-SCI, in more detail. 42 –45 The present study capitalizes on this by using the Chx10-eGFP transgenic mouse to investigate the distribution of V2a SpINs within the cervical spinal cord associated with the phrenic motor system, demonstrating a statistically significant recruitment of this interneuronal subtype into the phrenic circuitry after a high cervical SCI.
Conclusion
These data are among the first to begin to classify the subpopulations of interneurons involved in respiratory plasticity after SCI. 46 Identifying that these specific glutamatergic interneurons could be involved with respiratory plasticity post-SCI helps to define therapeutic targets for enhancing spontaneous recovery processes, but also reveals neuronal candidates that could be beneficial for therapeutically driven repair of the injured spinal cord (e.g., transplantation of interneuronal precursor subtypes, pharmacological treatment). Future studies should examine the role of V2a in post-SCI function in more detail and across additional experimental models—specifically, exploring whether diaphragm function is enhanced with greater recruitment of these cells at more chronic times post-SCI, or with increased connectivity and entrainment of these cells.
Footnotes
Acknowledgments
This work was funded by the NINDS, NIH R01 NS081112 (MAL), R01 NS095366 (KJD), P01 NS 055976, Craig H. Neilsen (338432, MAL) and the Spinal Cord Research Center at Drexel University, College of Medicine (MAL). PRV614 was produced and supplied by Dr David Bloom (University of Florida). Primary antibodies to PRV were supplied by Lynn Enquist (Princeton University) as part of Virus Center funding (P40 RR018604) awarded to Dr. Enquist.
Author Disclosure Statement
No competing financial interests exist.