
Abstract
In mature neurons, low intracellular chloride level required for inhibition is maintained by the potassium-chloride cotransporter, KCC2. Impairment of Cl− extrusion after KCC2 dysfunction has been involved in many central nervous system disorders, such as seizures, neuropathic pain, or spasticity, after a spinal cord injury (SCI). This makes KCC2 an appealing drug target for restoring Cl− homeostasis and inhibition in pathological conditions. In the present study, we screen the Prestwick Chemical Library® and identify conventional antipsychotics phenothiazine derivatives as enhancers of KCC2 activity. Among them, prochlorperazine hyperpolarizes the Cl− equilibrium potential in motoneurons of neonatal rats and restores the reciprocal inhibition post-SCI. The compound alleviates spasticity in chronic adult SCI rats with an efficacy equivalent to the antispastic agent, baclofen, and rescues the SCI-induced downregulation of KCC2 in motoneurons below the lesion. These pre-clinical data support prochlorperazine for a new therapeutic indication in the treatment of spasticity post-SCI and neurological disorders involving a KCC2 dysfunction.
Introduction
G
A recent high-throughput screening assay identified CLP257 as a positive modulator of KCC2 able to restore impaired Cl− homeostasis in dorsal horn neurons derived from a rat model of neuropathic pain. 20 Likewise, activation of 5-hydroxytryptamine type 2A (5-HT2A) receptors with (4-bromo-3,6-dimethoxybenzocyclobuten-1-yl)methylamine hydrobromide (TCB-2) repairs alterations of Cl− homeostasis in motoneurons of spastic rats post-SCI. 21 Although promising, transfer of CLP257 or TCB-2 from research discovery to patients is a costly and arduous undertaking. The strategy of repurposing approved drugs for another indication reduces time and effort in bringing a new chemical entity from bench to bedside. To this end, we have screened molecules from the Prestwick Chemical Library® consisting of off-patent approved drugs to identify a new KCC2 enhancer in a bioavailable and safe formulation. We identified piperazine phenothiazine derivatives as pharmacologically active agents for activating KCC2. We focused on prochlorperazine and found that, in addition to activating KCC2 function, it reduces spasticity in rats with SCI.
Methods
Animals
Wistar rats were housed under a 12-h light/dark cycle in a temperature-controlled area with ad libitum access to water and food. All animal care and use conformed to the French regulations (Décret 2010-118) and were approved by the local ethics committee CEEA 71–Comité d'éthique en neurosciences–INT Marseille (authorization Nb A9 01 13).
Surgery
Surgery and postoperative care were described previously. 18 Briefly, we anesthetized adult female Wistar rats (225/250 g; Charles River, Burlington, MA) with a mixture of ketamine (50 mg/kg, intraperitoneal [i.p.], Imalgen®; Merial, Duluth, GA) and medetomidine (0.25 mg/kg, i.p., Domitor®; Janssen Pharmaceutica, Beerse, Belgium) and neonates by hypothermia. Amoxicillin was administered after anesthesia (150 mg/kg, subcutaneous [s.c.]; Duphamox LA®; Pfizer Inc, New York, NY). After laminectomy, the spinal cord was transected at the T8 thoracic level (for neonates at the day of birth). In some adult rats, a catheter was inserted with the distal end in proximity to the lumbar enlargement. Sham-operated animals were subjected to all procedures except the spinal cord transection.
Drugs
We purchased perphenazine, prochlorperazine dimaleate (PCPZ), fluphenazine, trifluoperazine dihydrochloride, thioproperazine dimesylate, thiethylperazine from Prestwick Chemical® (Illkirch, France) or Tocris (Bristol, UK); ouabaine, bumetanide, acetophenazine, baclofen, DIOA [(dihydroindenyl)oxy]alkanoic acid], and NEM (N-ethylmaleimide) from Sigma-Aldrich (St. Louis, MO); perazine from Santa Cruz Biotechnology (Dallas, TX, USA); and 2-amino-5-phosphonovaleric acid (AP5) and 6-cyano-7-nitroquin-oxaline-2,3-dione (CNQX) from Abcam (Cambridge, UK). AP5 and CNQX were prepared in artificial cerebrospinal fluid (aCSF), ouabaine in H2O, and bumetanide, phenothiazines piperazine, baclofen, and DIOA in dimethyl sulfoxide (DMSO).
Drug administration and assessment of spasticity in adult spinal rats
We used the rate-dependent depression (RDD) of the H-reflex as a measurement of spastic symptoms. 15 A pair of stainless steel stimulating needle electrodes were inserted transcutaneously into the surroundings of the tibial nerve, a recording electrode into the flexor digitorum brevis muscle (flexor of the lateral four toes), and the reference electrode was placed s.c. into the foot. We stimulated the tibial nerve for 0.2 ms at 0.2 Hz with increasing current intensities and determined the intensity necessary to get a maximal H response. Then, we used this intensity for trains of 12 stimulations at 0.2, 1, 2, and 5 Hz. Normalization of the amplitude of responses (the first three discarded) to the controls evoked at 30-sec intervals (mean of five pulses) determined the level of the RDD. Chronic spastic rats were treated with a single intravenous (i.v.) administration of PCPZ (10 μg/kg in saline +0.1% DMSO), baclofen (2 mg/kg in saline +0.1% DMSO), or vehicle (saline +0.1% DMSO). DIOA was intrathecally (i.t.) delivered (40 μg in Dulbecco's phosphate-buffered saline, calcium, and magnesium +10% DMSO). The RDD was evaluated twice before drug injection and at intervals of 20, 40, 60, and 80 min after the drug injection.
In vitro electrophysiological recordings
The spinal cord below T8 was isolated from neonatal rats (postnatal day [P]4–P6), as previously described, 22 and transferred to the recording chamber perfused with an oxygenated (95% O2/5% CO2) aCSF composed of the following (in mM): 130 NaCl, 4 KCl, 3.75 CaCl2, 1.3 MgSO4, 0.58 NaH2PO4, 25 NaHCO3, and 10 glucose (pH 7.4; 24–26°C). Extracellular recording/stimulation were made from lumbar L3–L5 ventral (VR) and dorsal (DR) roots by contact stainless steel electrodes insulated with Vaseline. A glass suction electrode was also used to stimulate the ipsilateral ventral funiculus at the L2–L3 level. AC recordings from VR were amplified ( × 2000) and bandpass filtered from 70 Hz to 3 kHz. Current clamp recordings from L4 to L5 motoneurons were performed in DCC mode with an Axoclamp 2B amplifier (Molecular Devices, Sunnyvale, CA). Motoneurons were identified by an antidromic response to VR stimulation. Microelectrodes were filled with 2 M of K-acetate (90–150 MΩ). Intracellular signals were sampled at 10 kHz (Digidata 1440a; Clampex 10 software; Molecular Devices).
To estimate the ionic gradient of Cl− efflux in motoneurons, we measured the reversal potential of the inhibitory postsynaptic potential (EIPSP) evoked by ipsilateral ventral funiculus stimuli and pharmacologically isolated with CNQX (10 μM) and AP5 (50–100 μM). The amplitudes of inhibitory post-synaptic potentials (IPSPs) measured at various holding potentials (500-ms-long current pulses) were plotted to obtain EIPSP from the regression line.
Reciprocal inhibition was assessed by using a variation of the monosynaptic model technique. 23 We evoked monosynaptic reflexes in VR L5 by a supramaximal stimulation of the DR L5 (0.3-ms duration). When this stimulation was preceded by a stimulation of the DR L3 (2 × times the threshold [T] for the monosynaptic reflex) with a delay ranging from 0 to 40 ms, the amplitude of the L5 monosynaptic reflex was reduced, indicative of the reciprocal inhibition. Normalization of the amplitude of the L5 monosynaptic reflex recorded before and after the conditioning DR L3 stimulation were used to assess the reciprocal inhibition.
Stable mammalian cell line expressing KCC2
HEK-293 cells stably expressing KCC2 were grown to confluence at 37°C with 5% CO2 in Dulbecco's modified Eagle's medium/Nutrient mixture F-12 supplemented with fetal bovine serum (10%), penicillin (50 U/m), and streptomycin (50 pg/mL; Life Technologies, Carlsbad, CA). Puromycin (20 μg/mL; Life Technologies) was added to select KCC2-expressing cells.
Thallium influx assay
The FluxOR™ Thallium detection kit from Life Technologies was used. Briefly, HEK-293 cells were loaded with the thallium (Tl+)-sensitive fluorescent dye, FluxOR, for 90 min. Once the unloaded dye was removed, cells were supplemented with assay buffer containing ouabain (200 μM) and bumetanide (10 μM). Cells were then plated (100,000 cells/75 μL/well) in clear-bottomed, black-walled, 96-well plates (Greiner Bio-One, Monroe, NC). Compounds to be assayed from the Prestwick library collection were added (5 μL/well) by using the Biomek® NX Laboratory automation workstation (Beckman Coulter, Villepinte, France) and incubated for 15 min before measuring an initial baseline fluorescence (490 nm excitation and 520 nm emission) onto the screening system POLARstar omega (BMG Labtech, Ortenberg, Allemagne). The fluorescence signal was initiated by adding 20 μL of a 5 × Tl2SO4 solution (final concentration of 2 mM of Tl2SO4) and recorded 30 min after. The test was conducted for each plate using two columns on which cells on half were treated with 33 μM of NEM, an accepted activator of KCC2, 24 as positive control and the other half with no compounds as negative control. After subtracting each well's fluorescence obtained after compound addition (baseline fluorescence), activity was expressed as percentage change of Tl+ influx in compound-treated cells versus influx in vehicle-treated cells. Hits were selected as compounds that cause an increase >20% without apparent effect on parental untransfected HEK cells. Hits were retested in triplicate. Positive compounds were further evaluated by testing them at varying concentrations. To evaluate the quality of the Tl+ flux assay, the value of Z’ factor was calculated using the following equation: Z’ = 1-[3 SD of sample +3 SD of control]/[mean of sample-mean of control]. At the end of the fluorescent assay, cell viability was checked by the PrestoBlue® Cell Viability Reagent (Life Technologies) protocol. Briefly, cells viability in 96-well plates were expressed as the 570-nm absorbance ratio of drug-treated cells compared with cells incubated in the absence of drug (medium). The result was expressed as percentage of the control (defined as 100%). The initial screen of 880 compounds was performed at 2 μg/mL (3–10 μM), 12 μg/mL (18–60 μm), and 20 μg/mL (30–100 μM).
Immunostaining KCC2
Adult rats were perfused transcardially with 4% paraformaldehyde (PFA; Sigma-Aldrich) in phosphate-buffered saline (PBS; bioMérieux SA, Marcy l'Etoile, France) at 4°C. Tissues were post-fixed overnight (16 h) in 4% PFA and washed 3 times with PBS. Lumbar spinal cords (L4–L5) were then embedded in 4% low-melting agarose and sectioned (30 μm) with a vibratome. Sections from all samples were mounted on the same poly-lysine-coated slides and processed for immunohistochemistry. Slices were thus 1) dried overnight, 2) permeated (45 min) in PBS with 3% bovine serum albumin (BSA; Sigma Aldrich), and 0.2% Triton X-100, 3) washed in PBS (2 × 5 min), 4) preincubated (1 h) in PBS with normal goat serum (3% BSA, 1/100 normal goat serum), 5) washed and incubated overnight at room temperature in the affinity-purified rabbit anti-KCC2 polyclonal antibody (diluted 1:500; catalog no.: 07-432, RRID:AB_310611; Merck-Millipore Billerica, MA), 6) washed in PBS (3 × 5 min), 7) incubated (1 h) with a goat antirabbit immunoglobulin (Ig) G conjugated to Alexa-546 (diluted 1:500; Life Technologies), 7) washed in PBS 3 × 5 min, and finally 9) cover-slipped with a gelatinous aqueous medium. In control experiments, the primary antiserum was replaced with rabbit Ig fraction during the staining protocol. Images were taken using the confocal microscope (LSM500; Carl Zeiss, Jena, Germany) at 60 × magnification, digitized in stacks of 0.5-μm-thick optical sections and processed with the Zen (Carl Zeiss) software.
We quantified KCC2 immunolabeling of motoneuron (identified as the biggest cells in the ventral horn). Line scans show peaks of intensity at the periphery of the cell body, likely plasma membrane. We drew a region of interest within the presumed plasma membrane and measured the mean pixel intensities in this delimited area. 15,18,21 Values from sham-, SCI-, and PCPZ-treated rats were then normalized to the mean value measured from sections of SCI rats on the same slide.
Statistical analysis
Group measurements are expressed as means ± standard error of the mean (SEM). We used Mann-Whitney U tests, Kruskal-Wallis tests, and one- or two-way analyses of variance (ANOVAs) with or without repeated measures, as appropriate (GraphPad Prism 5 software; GraphPad Software Inc., Cary, NC). For all statistical analyses, the data met the assumptions of the test and the variance between the statistically compared groups was similar. The significance was set at p < 0.05.
Results
Phenothiazine piperazine derivatives upregulate KCC2 function
To identify KCC2 activators from the Prestwick Chemical Library, we used a standard Tl+ flux assay reported as suitable for high-throughput screening in the HEK-293 cell line expressing KCC2. 25 –27 In principle, upon the activation of KCC2 in reverse mode, the uptake of Tl+ into cells releases a fluorescent signal from the pre-loaded FluxOR dye, which reports the activity of KCC2. The initial screen of 880 compounds performed at the concentration of 12 μg/mL (18–60 μM) yielded 121 hits with more than 20% increase in the fluorescent signal (Fig. 1A). The Z factor with an average of 0.83 ± 0.004 (range, 0.75–0.92) conformed to the assay quality requirement (Fig. 1B). We then selected 27 compounds among these 121 hits and that were ineffective on naïve HEK293 cells. A secondary screen tested each compound in triplicate and confirmed the activity of approximately 60% of them (17 of 27). A classification of positive hits revealed four molecules (perphenazine, prochlorperazine dimaleate, trifluoperazine dihydrochloride, and thioproperazine dimesylate) that belong to antipsychotic piperazine phenothiazine derivatives characterized by a three-ring structure and a piperazine substituent on nitrogen (Fig. 1C). Based on their commercial availability, eight piperazine phenothiazines were purchased for further evaluation (chemical structure of each compound is illustrated in Fig. 1C). To determine their potency, dose-response curves (range, 0.1–50 μM; example for prochlorperazine in Fig. 1D) were performed in parallel to a counter screen to evaluate their cytotoxicity. All compounds were effective with the minimal effective concentration ranging from 3 to 12 μM (see Fig. 1C). Prochlorperazine, thiethylperazine, fluphenazine, and perazine were the most potent activators (∼140% of basal activity). Among these molecules, prochlorperazine dimaleate (termed henceforth as PCPZ) exhibited the most potent effect with the lowest minimal effective concentration (3 μM) without apparent cytotoxic effect at concentration below 200 μM (see Fig. 1C). Therefore, from a drug repurposing point of view, PCPZ was selected for further experimental investigations of the molecule.
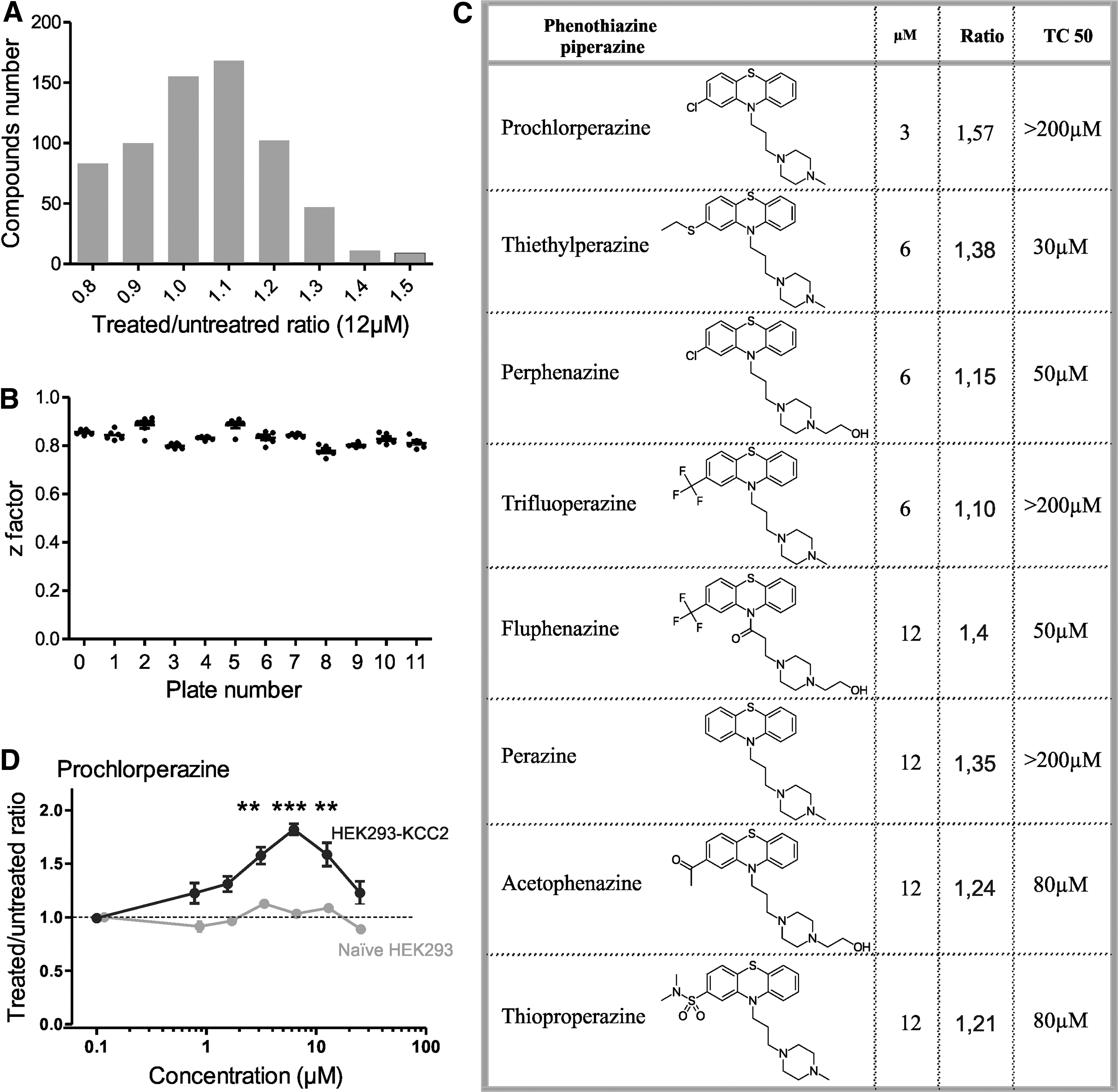
Prestwick library screening on KCC2 using the thallium flux assay in HEK-293 cells. (
Prochlorperazine dimaleate enhances Cl− extrusion capacity in motoneurons
By means of intracellular recordings, we investigated whether PCPZ could enhance Cl− extrusion capacity of lumbar motoneurons recorded in in vitro isolated whole spinal cord preparation from intact neonatal rats (postnatal day [P]4–P6). The effect of PCPZ was examined by determining the reversal potential of inhibitory post-synaptic potentials (EIPSP) as a measure of the KCC2 function. 28 We found that PCPZ (10 μM) hyperpolarized EIPSP within 20–25 min by −3.3 ± 0.66 mV (Fig. 2A; n = 6; *p < 0.05, Wilcoxon's test). Given that there was no change of the resting membrane potential (Vrest; n = 6; Fig. 2a; p > 0.05, Wilcoxon's test), the net driving force for Cl− (EIPSP-Vrest) increased significantly (n = 6; Fig. 2A; *p < 0.05, Wilcoxon's test), potentially strengthening the inhibitory synaptic transmission.

Prochlorperazine hyperpolarizes EIPSP and increases the strength reciprocal inhibition. (
Prochlorperazine dimaleate strengthens inhibition and decreases spasticity in rats with spinal cord injury
To test the ability of PCPZ to enhance the post-synaptic inhibition, we took advantage of the recent in vitro demonstration that the reciprocal inhibition in the spinal cord is markedly reduced post-SCI. 29 Typically, flexor-related Ia interneurons (recruited by stimulation of the DR L3) inhibits L5 extensor-related motoneurons, 30 such as when the DR L5 stimulation was preceded by DR L3 stimulation, there was a reduction of the DR L5-evoked monosynaptic response (Fig. 2B). This reduction almost disappeared in spinal cord isolated from neonatal rats that underwent SCI at birth (Fig. 2B). In those rats, the bath application of PCPZ at a concentration as low as 10 μM significantly reduced the amplitude of the L5 monosynaptic response to values observed in intact animals (Fig. 2B, n = 6; ### p < 0.001; two-way repeated-measures ANOVA). Altogether, these results indicate that PCPZ is able to restore the reciprocal inhibition after neonatal SCI.
The Hoffmann reflex (H-reflex), resulting from the monosynaptic activation of motoneurons by Ia afferents, declines in amplitude over repetitive stimulation and becomes more depressed as the frequency of stimulation increases. 31,32 The lower rate-dependent depression (RDD) of the H-reflex in spastic patients and animals identifies RDD as a reliable assessment of spasticity. 15,33,34 Because the low RDD is indicative of a spinal disinhibition and a KCC2 dysfunction, 15 we tested whether the hyperpolarizing shift of EIPSP in motoneurons by PCPZ would restore the RDD in adult rats with chronic SCI (21 days post-SCI). The i.v. administration of PCPZ (10 μg/kg) or vehicle (DMSO) did not affect the maximal amplitude of H-reflex and M-wave (Table 1), but significantly increased the RDD to values observed in SCI animals (Fig. 3A). At frequencies of 1 Hz and higher, the amplitude of the H wave over repetitive stimulation was smaller than after vehicle injection, with a maximal effect 80 min after PCPZ administration (n = 9 vehicle, n = 8 PCPZ; Fig. 3A; two-way ANOVA test with Bonferroni's post-test; not significant [ns] p > 0.5; *p < 0.5; **p < 0.01; ***p < 0.001). To identify the mechanisms underlying this effect, we i.t. administered DIOA (40 μg), a highly specific KCC2 blocker, 25 50 min after PCPZ injection. KCC2 blockade reversed the PCPZ effect (Fig. 3B; n = 6 in each group), which is consistent with the hypothesis of KCC2 mediating the EIPSP shift induced by PCPZ. Taken together, these results indicate that PCPZ reduces spasticity essentially through a modulation of KCC2.
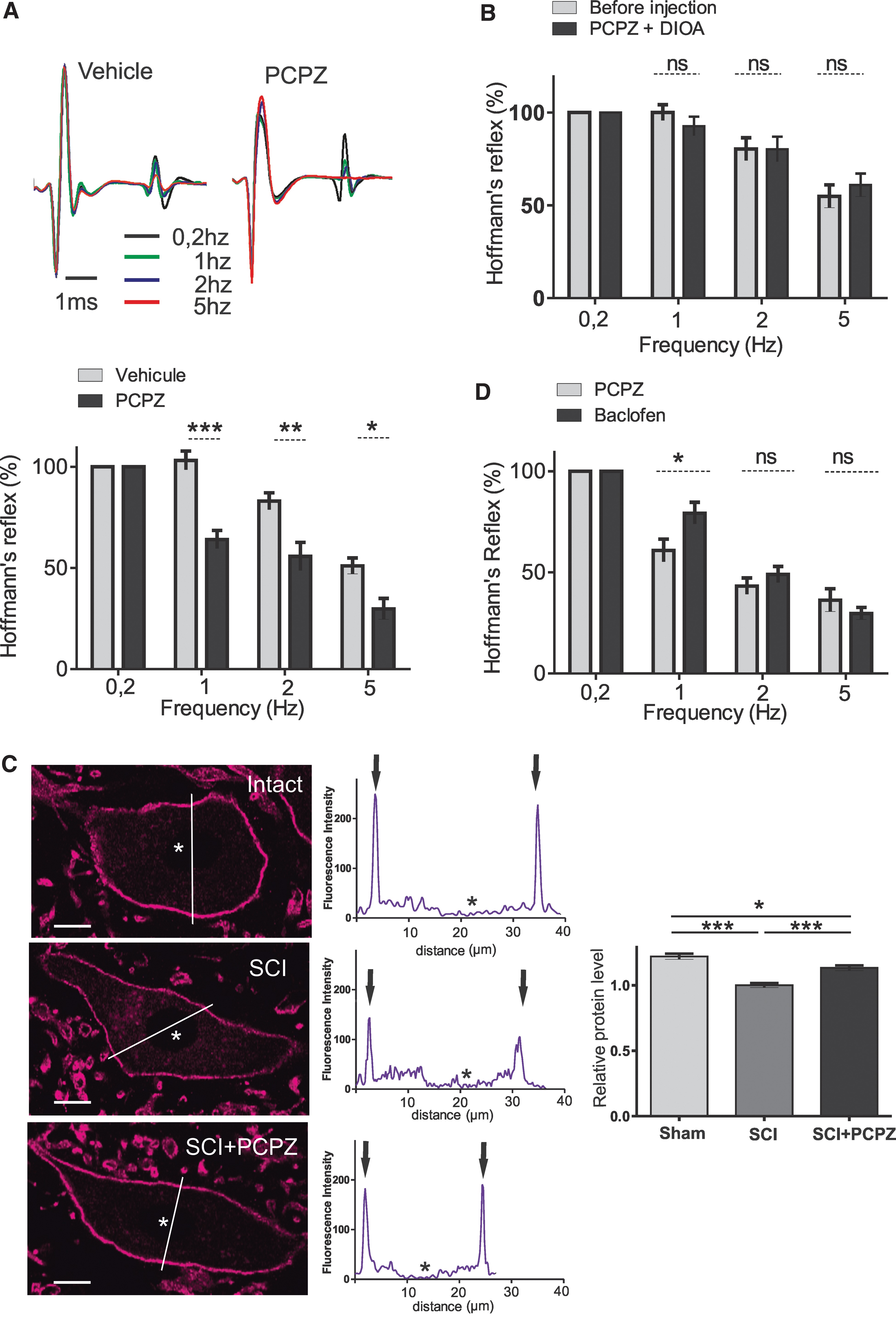
PCPZ decreases spasticity in rats with SCI through KCC2. (a) Prochlorperazine increases the RDD of the H reflex in chronic SCI adult rats. Top: M and H waves evoked in control (vehicle) and 80 min after PCPZ intravenous (i.v.) injection (10 μg/kg). Each trace is the mean response to three last consecutive stimulations at 0.2, 1, 2, or 5 Hz. Bottom: mean relative amplitudes of the H reflex after (80 min) the injection of vehicle (gray) or PCPZ (black, n = 9 rats with vehicle; n = 8 rats with PCPZ). (
PCPZ, prochlorperazine dimaleate; DMSO, dimethyl sulfoxide; DIOA, [(dihydroindenyl)oxy]alkanoic acid]; SEM, standard error of the mean; inject., injection.
Prochlorperazine dimaleate increases the expression of KCC2 in motoneurons below the spinal cord injury
We used immunohistochemistry to quantify the effect of PCPZ on the expression of KCC2 on lumbar motoneurons post-SCI. Peaks of KCC2 labeling were detected in the periphery of cell bodies of sham-, SCI-, and PCPZ-treated rats showing a membrane labeling (Fig. 3C). Levels of KCC2 labeling surrounding motoneurons were higher in PCPZ-treated compared to vehicle-treated animals (10 μg/kg; i.v. 80 min), but did not return to the level found in sham-operated rats (Fig. 3C, sham vs. SCI + prochlorperazine *p < 0.05; SCI vs. SCI + prochlorperazine ***p < 0.001; Kruskal-Wallis' test, Dunn's post-tests; n = 231 motoneurons in each group).
Prochlorperazine dimaleate is as effective in reducing spasticity as baclofen
From a translational perspective, the therapeutic profile of a new antispastic drug cannot be defined solely on the basis a comparison to vehicle-treated rats. We thereby assessed PCPZ potential advantages with respect to existing antispastic drug. The GABAB receptor agonist, baclofen, is the most effective and widely used drug for the treatment of spasticity post-SCI. 35 –40 We compared the relative efficacy of PCPZ to that of the “gold standard,” baclofen (2 mg/kg, i.v.), in restoring the RDD. The reduction of the RDD was more pronounced in PCPZ-treated animals at 1 Hz and remained comparable to that of baclofen-treated rats at higher frequencies of stimulation (Fig. 3D, two-way ANOVA test with Bonferroni's post-test; ns p > 0.5; *p < 0.5, n = 6 in each group). These results suggest that antispastic effects of PCPZ are at least similar to that of baclofen.
Discussion
We identified PCPZ as a positive modulator of KCC2 capable to restore endogenous inhibition of motoneurons post-SCI. PCPZ leads to a hyperpolarizing shift of EIPSP and reduces SCI-induced spasticity. The observed effect of PCPZ on spasticity likely results from an enhancement of KCC2 function. In line with this assumption, PCPZ increases cell-surface expression of KCC2 in motoneurons post-SCI and blocking KCC2 prevents the effect of PCPZ to alleviate spasticity. Likewise, the reciprocal inhibition, downregulated post-SCI and determined by the level of KCC2 function, 29 is restored by PCPZ.
Deficits in KCC2 function have been documented in brain and spinal cord injuries, 15,41 in temporal lobe epilepsy, 42,43 and in neuropathic pain. 44 –47 Accordingly, KCC2 appears to be an attractive target to restore endogenous inhibition in pathological conditions. Previous investigations showed KCC2 as a druggable target in modulating [Cl−]i for the development of therapeutics. The 5-HT2A high-affinity agonist, TCB-2, reduces spasticity post-SCI by boosting the KCC2 function, 21 whereas CLP257, a new KCC2 enhancer, alleviates hypersensitivity in a rat model of neuropathic pain. 20 Unfortunately, these two compounds are not approved human drugs. Conversely, PCPZ is currently licensed for its neuroleptic actions with an adult daily dosage ranging from 15 to 150 mg. 48 Typical antipsychotic drugs such as PCPZ may produce extrapyramidal side effects at clinically effective doses against schizophrenia. 48 However, the antispastic effect of PCPZ, equivalent to that of baclofen, was achievable with a low human equivalent dose estimated at 1.6 μg/kg for a 70-kg human [HED = Animal Dose (mg/kg) × (animal wt/human wt in kg)0.33. 49 The phenothiazine ring possesses a high degree of lipophilicity, and these drugs easily cross the blood–brain barrier. 50 –52 Chlorine substitution of C-2 atom of prochlorperazine confers less antipsychotic activity than other substituents (X = -SO2NR2 > -CF3 > -CO-CH3 > -Cl). 52 Collectively, the use of PCPZ at lower doses represents an effective and tolerable therapeutic strategy for reducing spasticity, likely better than the common antispastic agent, baclofen, which often causes sedation and dizziness. 38,39
The mechanism by which PCPZ modulates KCC2 function in motoneurons has not been investigated in the present study. The therapeutic potential of PCPZ in reducing psychosis has been mainly attributed to its antidopaminergic action. 53,54 However, the main source of dopamine in the mammalian spinal cord arises from the diencephalon and disappears post-SCI. 55,56 Further, dopamine appears to be antispastic by restoring the RDD in chronic SCI rats, 57 thereby suggesting that inhibition of dopaminergic receptors unlikely accounts for the upregulation of KCC2. Antipsychotic drugs have been also found to display antiserotoninergic actions, most notably through an inverse agonist activity of 5-HT2C receptors. 58 Interestingly, 5-HT2C receptors in motoneurons become constitutively active post-SCI and contribute to muscle spasms. 59 We recently showed that blocking 5-HT2BRs and 5-HT2CRs hyperpolarized EIPSP in rats with SCI. 21 However, PCPZ is one of the typical antipsychotic devoided of inverse agonist activity 58 and thus unlikely modify KCC2 function through 5-HT2C receptor-mediated mechanisms.
PCPZ may also interact indirectly through some intermediate signaling molecules, as suggested by the potent inhibitory action of phenothiazines on Ca2+-dependent protein kinase C (PKC). 60 The phosphorylation of KCC2 by PKC constitutes a powerful and dynamic mechanism to influence KCC2 function. 61 Post-SCI, a tonic activation of a Ca2+-dependent PKC signaling pathway alters motoneuronal Cl− homeostasis by depolarizing shift EIPSP. 21 A similar relationship between Ca2+-dependent PKC isozymes and the positive shift of ECl has been pointed out in hippocampal neurons. 62 Therefore, the potency of PCPZ in restoring KCC2 function may be connected with its inhibitory action on Ca2+-dependent PKC. 60
In sum, we provide strong pre-clinical evidence for translation to chronic SCI subjects, a process that will be facilitated because PCPZ is already an approved compound and has been recently patented for treating spasticity (WO 2015135947 A1). 63
Footnotes
Acknowledgments
We dedicate this article in memory of our colleague and friend, Laurent Vinay, who passed away on March 26, 2015. The work was funded by the Agence National de la Recherche Scientifique (to L.V., 2010-BLAN-1407-01), the Fondation pour la Recherche Médicale (to L.V. and F.B., DEQ20130326540), the French Institut pour la Recherche sur la Moelle épinière et l'Encéphale (to L.V. and F.B.), and SATT Sud-Est (Sociétés d'Accélération du Transfert de Technologies, to L.V.). We thank Jean Claude Guillemot and Bruno Canard for advice and facility of the screening platform Marseille-Luminy (PCLM). HEK-293 cells stably expressing KCC2 were generously provided by Dr. Eric Delpire (Vanderbilt University School of Medicine, Nashville, TN).
Author Disclosure Statement
No competing financial interests exist.