
Abstract
Persistent inflammation, mediated in part by increases in cytokines, is a hallmark of traumatlc brain injury (TBI). Minocycline has been shown to inhibit post-TBI neuroinflammation in male rats and mice, but has not been tested in females. Here, we studied sex differences in thermal, stress, and inflammatory responses to TBI and minocycline. Female rats were ovariectomized under isoflurane anesthesia at 33–36 days of age. At 45–55 days of age, male and female rats were implanted intraperitoneally (i.p.) with calibrated transmitters for monitoring body temperature. Moderate cortical contusion injury (CCI) or sham surgery was performed when the rats attained 60–70 days of age. One hour after surgery, rats were injected i.p. with minocycline (50 mg/kg) or saline (0.3 mL); injections were repeated once daily for the next 3 days. At 28 days after CCI or sham surgery, 30 min restraint stress was initiated and blood samples were obtained by tail venipuncture before the onset of restraint and at 30, 60, and 90 min after stress onset. At 35 days after CCI or sham surgery, rats were decapitated and blood was collected for corticosterone (CORT) and cytokine analysis. The brains were removed and ipsilateral cortical tissue and hippocampus were dissected and subsequently assayed for interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α. Hyperthermia occurred during days 1–6 post-CCI in male rats, but only on the day of CCI in female rats, and minocycline prevented its occurrence in both sexes. Minocycline facilitated suppression of the CORT response to restraint stress in both sexes. In females, but not males, hippocampal IL-6 content increased post-CCI compared with sham-injured controls, whereas IL-1β content was augmented by minocycline. Hippocampal TNF-α was unaffected by CCI and minocycline. These results demonstrate sex differences in immediate thermal and long-lasting stress and cytokine responses to CCI, and only short-term protective effects of minocycline on hyperthermia.
Introduction
P
Various anti-inflammatory drug treatment strategies have been shown to inhibit post-TBI neuroinflammation, as reviewed by Kumar and Loane. 6 Minocycline is one such anti-inflammatory drug that was shown to prevent the development of neurobehavioral abnormalities and to normalize serum and tissue levels of selected inflammatory markers as late as 45–51 days after mild blast injury in male rats. 7 Minocycline also suppressed the elevated serum levels of corticosterone (CORT) at day 51 in these brain-injured rats. 7 In our prior work using adult male rats, moderate cortical contusion injury (CCI) induced elevations of body temperature for at least the first 5 days post-injury, 8 and at 28 days after CCI did not affect basal levels of CORT, but did suppress the CORT response to restraint stress 9,10
As we discussed previously, 9 TBI causes a generalized activation of the hypothalamus–pituitary–adrenal (HPA) axis during the early post-injury period and induces a constellation of early, acute-phase responses that includes fever. These responses are mediated at least in part by pro-inflammatory cytokines, such as interleukin (IL)-1β, IL-6 and tumor necrosis factor-α (TNF-α), which are released within minutes of the injury. 11,12 However, beyond the early effects of TBI on neuroendocrine-immune function, the responses of this system at later time points remain essentially unknown. Given the known sex differences in human inflammatory responses, 13 the present experiments were designed to determine the effects of minocycline treatment, according to the protocol of Kovesdi and colleagues, 7 in male and female rats with moderate CCI. We examined the short-term effects of minocycline treatment on body temperature during the first 6 days after moderate CCI. Long-term effects of the drug were studied using the CORT response to restraint stress at 28 days post-CCI, and cytokine levels in blood and brain tissue at 35 days after CCI in male and ovariectomized female rats.
Methods
Subjects
In two separate experiments, male (n = 30) and female (n = 30) Sprague–Dawley rats, received from Charles River Breeding Labs (Hollister, CA) as weanlings, were maintained in our standard temperature and lighting conditions (22 ± 1°C, 14 h/10 h lighting cycle, lights on: 0400–1800 h). Rats of each sex were group housed until temperature recording commenced 1 week prior to CCI/sham surgery, when they were individually housed to enable temperature recording from each rat. All animals were randomly assigned to injury (sham versus CCI) and drug treatment (saline versus minocycline) conditions. All experimental procedures were approved by the UCLA Institutional Animal Care and Use Committee.
Surgical procedures
At 33–36 days of age, all female rats were ovariectomized under isoflurane anesthesia, thereby alleviating estrous cyclicity of their gonadal hormones. At 45–55 days of age, all male and ovariectomized female rats were implanted intraperitoneally (i.p.) under isoflurane anesthesia with calibrated transmitters (Model VM-FH, Phillips Respironics, Sunriver, OR) for monitoring body temperature.
Moderate CCI (n = 16) or sham (n = 14) surgery was performed when all rats attained 60–70 days of age. Rats were anesthetized with isoflurane (2.0–2.5% in 100% O2, 2.0 mL/min flow rate) and placed into a stereotaxic frame (Kopf Instruments, Tujunga, CA) with the head positioned in a horizontal plane with respect to the interaural line. During all surgical procedures, body temperature was maintained at 37–38°C using a thermostatically controlled heating pad (Harvard Apparatus, Holliston, MA). All surgical procedures were performed under aseptic conditions and have been previously described in detail. 8 –10,14 In brief, after a midline incision, the skin, fascia, and temporal muscle were reflected. Animals receiving CCI were subjected to a 6 mm diameter craniotomy over the left parietal cortex centered at 3 mm posterior and 4 mm lateral to bregma. An electronically controlled, small bore, dual-stroke, pneumatic piston cylinder with a 40 mm stroke (Hydraulics Control, Inc., Emeryville, CA) was mounted onto a stereotaxic micromanipulator, allowing for precise control of the impact site and depth of tissue compression. The piston cylinder was angled 22.0 degrees away from vertical, enabling the flat, circular impactor tip (5 mm diameter) to be perpendicular to the surface of the brain at the site of injury. After induction of a moderate CCI (30 psi or ∼2.8 m/sec, 2.0 mm depth), the fascia and scalp were sutured closed and triple antibiotic was applied to the wound margins. Sham-operated control rats underwent similar procedures to control for surgical stress and duration of anesthesia, but did not receive craniotomy or any impact. Animals were returned to their individual cages with free access to food and water after recovery from anesthesia.
Biotelemetric measurement of core body temperature
Core temperature was monitored by a telemetric system (model VM-FH; Phillips Respironics, Sunriver, OR) at an ambient temperature of 22
Temperature recording from those rats with active transmitters commenced on the day prior to CCI/sham surgery; that is, Day 0. To avoid fluctuations in the temperature recordings caused by manipulation of the rats during their 1000–1200 h injections, analyses of variance (ANOVAs) of the daily averages of the 10 min body temperature data were performed for the 1300–1800 h light-phase period (Fig. 1). A comparable 5 h period commencing 1 h after lights out was selected for the dark phase daily averages (1900–2400 h, Fig. 2).
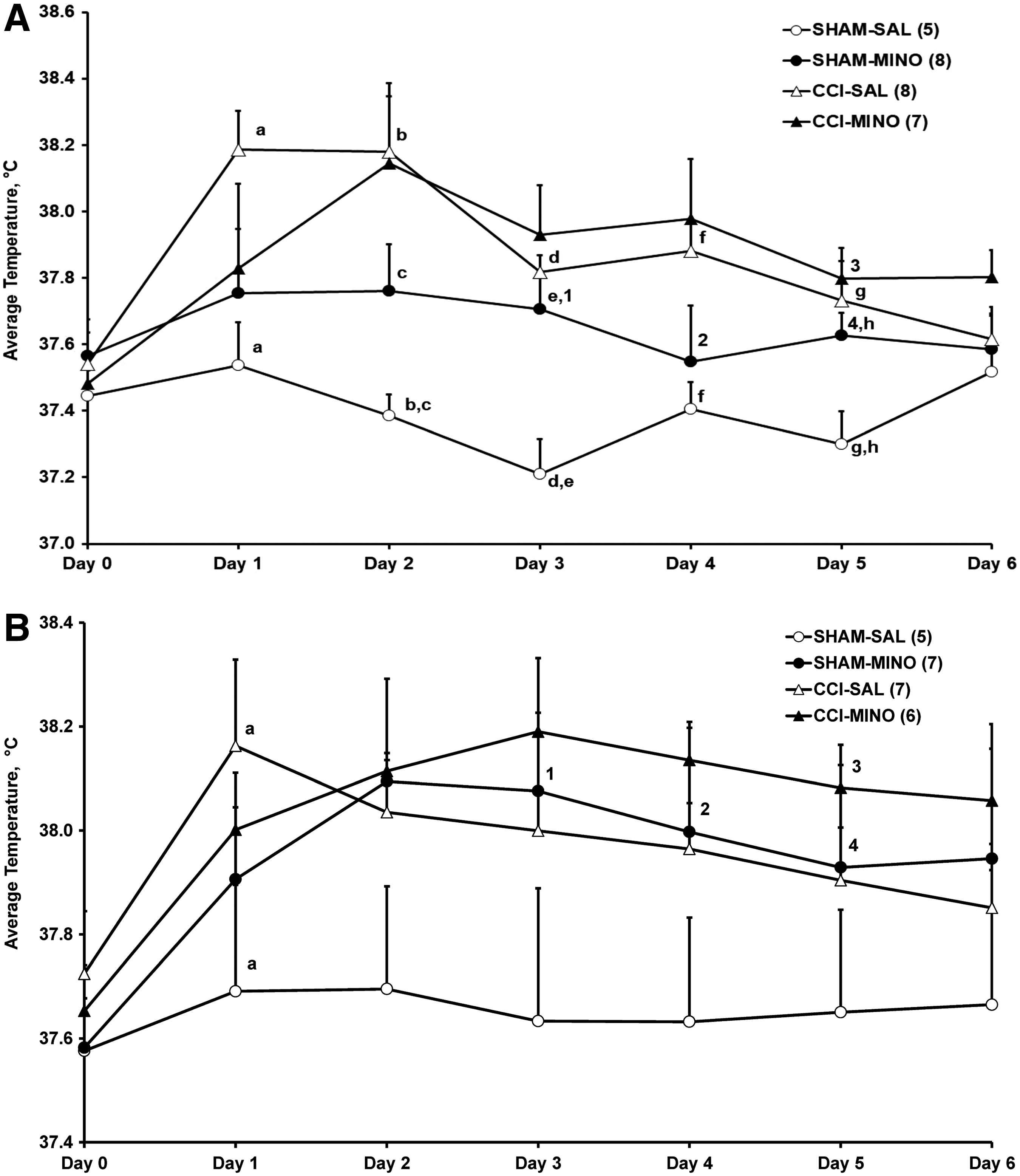
Average body temperature (mean ± SEM) during the light phase (1300–1800 h) in Sprague–Dawley male
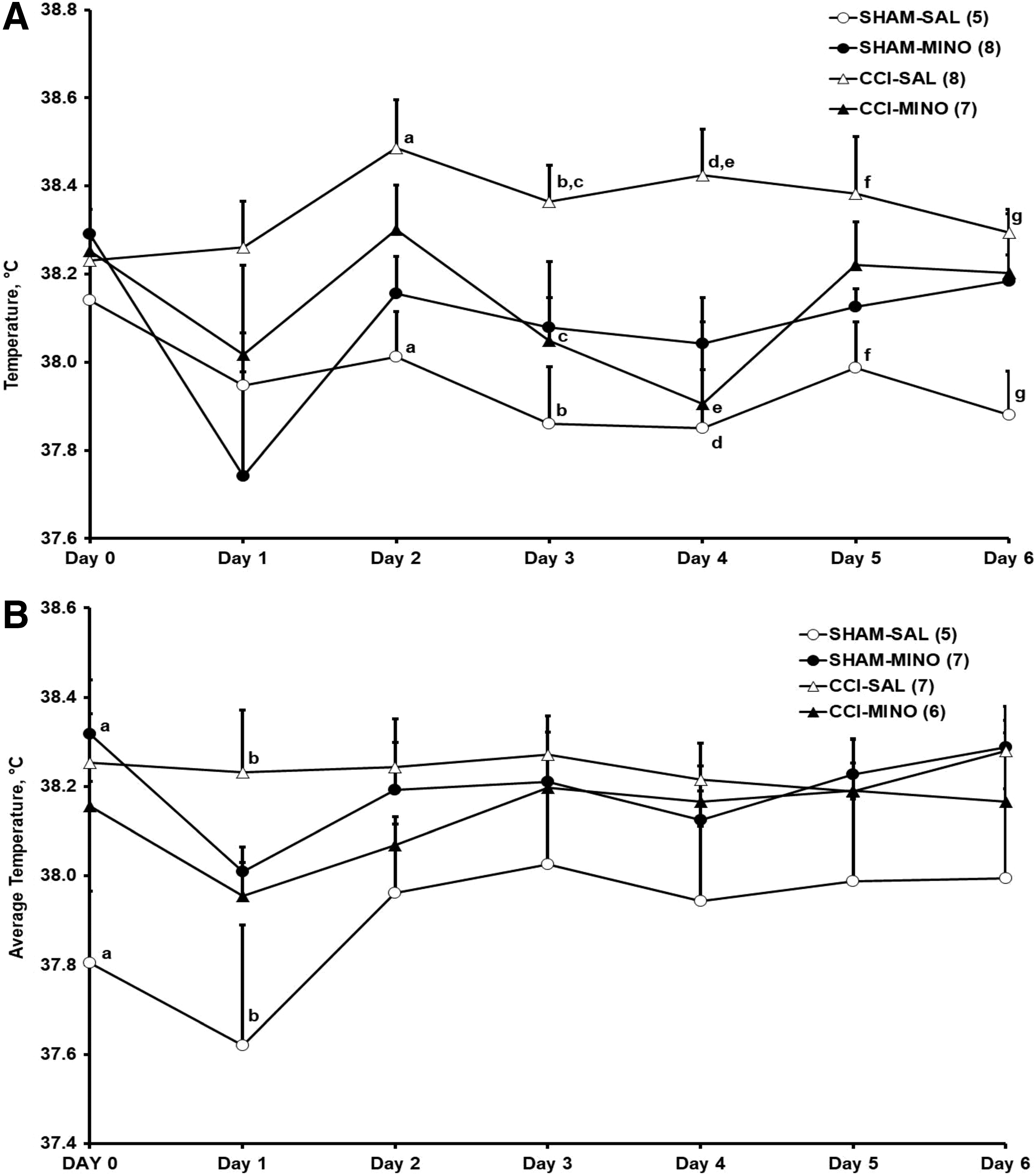
Average body temperature (mean ± SEM) during the dark phase (1900–2400 h) in Sprague–Dawley male
Drug treatment
One hour after injury/surgery all rats were injected i.p. with minocycline hydrochloride (Sigma) suspended in saline at a dose of 50 mg/kg bwt or saline (0.3 mL). The injections were repeated once daily for the next 3 days at the identical time of day (1000–1200 h), according to the procedure of Kovesdi and colleagues. 7
Restraint
At 28 days after CCI or sham surgery, restraint stress was initiated as we have done previously. 8 –10 Animals were placed into flat-bottom Plexiglas restraining tubes (13 cm length × 6 cm width × 8 cm height, Harvard Apparatus Co.) and tightly restrained for 30 min followed by light restraint for an additional 60 min. A blood sample was obtained by tail venipuncture immediately before onset of restraint. Additional blood samples were obtained at 30, 60, and 90 min after stress onset. All testing occurred between 0900 and 1300 h.
Assay of CORT in plasma
Blood samples (300 μL) were collected at the time points described, in microcapillary blood collection tubes that contained ethylenediaminetetraacetic acid (EDTA) with added aprotonin (200 KIU/mL). After centrifugation at 2000 rpm for 20 min, plasma was separated and stored at −80°C until analysis for CORT. Plasma CORT was assessed with a rat CORT 125I radioimmunoassay kit (MP Biomedicals, Irvine, CA), as we have done previously. 8 The reported detection limit of this assay was 8 ng/mL, the recovery of exogenous CORT was 100%, and intra- and interassay coefficients of variation were <10.3% and <7.2%, respectively. The results are expressed as nanograms per milliliter of plasma.
Assay of IL-1β, IL-6, and TNF-α in blood, cortex, and hippocampus
Allowing 1 week for recovery from restraint stress, at 35 days after injury or sham surgery, the rats were decapitated. Blood was collected for CORT and cytokine analysis, and brains were removed and immediately dissected on ice. Ipsilateral cortical tissue that contained the injury site with a 1 mm surround and the entire ipsilateral hippocampus were dissected separately, weighed, and placed on dry ice. Dissected cortical and hippocampal tissues were frozen at −80°C until assayed for cytokines with enzyme-linked immunosorbent assays (ELISA) specific for IL-1β, IL-6, and TNF-α.
For total protein extraction, tissues were homogenized in lysis buffer (50 mM Tris/HCL [pH 7], 250 mM NaCl, 5 mM EDTA, 2 mM Na2VO4, 1 mM NaF, 20 mM Na4P2O7, 0.02% NaN3). A proprietary detergent and protease inhibitor cocktail tablets (Roche Applied Science, Indianapolis, IN) were added according to the manufacturer's guidelines for protecting proteins from degradation by endogenous proteases released during protein extraction and purification procedures. Homogenates were centrifuged at 14,000g for 30 min at 4°C. The resulting supernatants were collected, aliquoted for cytokine assays, and immediately processed for total protein concentration according to the Micro BCA procedure (Pierce Biotechnology, Rockford, IL) using bovine serum albumin as the standard. All chemicals were obtained from Sigma-Aldrich unless otherwise noted.
IL-1β and TNF-α in plasma and IL-6 in serum and in cortical and hippocampal tissue supernatants were measured using specific commercially available ELISA kits from Thermo Fisher Scientific, Inc. (Rockford, IL) for IL-1β, and from Invitrogen Corporation (Camarillo, CA) for IL-6 and TNF-α. Standards and test samples in duplicate were pipetted to specific antibody pre-coated microplate wells and assayed according to the manufacturers' instructions. Optical density was measured at 450 nm using an automated microplate reader (Model 3550; BioRad Laboratories Inc.).
The assay range of the IL-1β kit was 25.6–2500 pg/mL, with a lower limit of detection of <12 pg/mL. The intra-assay and interassay precision (% coefficient of variability [CV]) range was 6.6–8.0 and 6.2–7.0, respectively. The minimum detectable amount of IL-6 was < 5 pg/mL. Its intra-assay precision (%CV) range was 2.8–5.8, whereas interassay precision (%CV) was 5.7–8.8. The minimum detectable amount of TNF-α was < 4 pg/mL. Intra-assay precision (%CV) range was 4.3–6.9%, whereas interassay precision (%CV) was 7.8–9.0. Results are expressed as pg/mg protein.
Statistical analysis
Two-way ANOVAs were performed on the temperature, CORT, and cytokine data for the factors of group (injury condition [CCI/sham] and drug/vehicle treatment) and sex. Given the differences in basal CORT levels, all results are expressed as the CORT responses; that is, difference from basal or 0-time at each subsequent time point. Individual group means were compared using t tests under the Tukey–Fisher least significant difference (LSD) criterion with α set at ≤0.05 (two sided).
Results
Minocycline and body temperature
During the light phase (Fig. 1A), CCI in male rats treated with saline induced significant (p ≤ 0.01) hyperthermia compared with the sham-saline (SHAM-SAL) controls in the first 5 days posti-injury, confirming our previous report. 8 Minocycline-treated CCI males showed hyperthermia on treatment Days 2–6, which was not significantly different from that of the CCI-SAL or the minocycline-treated sham males. On Days 2, 3, and 5, average body temperature was significantly (p < 0.05) higher in the minocycline-treated than in the saline-treated male sham rats (Fig 1A). During the dark phase (Fig. 2A), significant (p < 0.05) hyperthermia compared with the temperature of the SHAM-SAL controls persisted in the CCI saline-injected male rats during Days 1–6, and on Days 3 and 4, temperature was significantly (p < 0.05) higher in the CCI-SAL group than in the CCI- minocycline males (Fig. 2A).
In the females during the light phase (Fig. 1B), CCI had a significant (p = 0.05) hyperthermic effect on the day of CCI (Day 1) after the first treatment with saline, which did not persist to the subsequent 3 days of treatment or beyond. In the light phase (Fig. 1B), but not in the dark phase (Fig. 2B), average temperature in the CCI-minocycline females tended to be higher than that of the CCI-SAL females on Days 2–6 after injury. In the dark phase (Fig. 2 B) on Day 0, temperature in the sham-minocycline females was significantly (p < 0.05) higher than in SHAM-SAL females. The greater variability on Day 0 suggests that the shift from lights on to lights off exerts a stronger effect on females than on males. As in the males, the diurnal increase in body temperature is evident from light and dark phase comparisons of all groups on Day 0 (Figs. 1 and 2). On Day 1, temperature in the CCI-SAL females was significantly (p < 0.01) higher than that of the SHAM-SAL females.
On Days 3, 4, and 5, average temperatures of the sham-minocycline treated females in the light phase were significantly (p < 0.05) higher than those of their male counterparts (Fig. 1 A and B). On Day 5, light phase temperatures of the CCI-minocycline treated females were also significantly (p < 0.01) higher than those of their male counterparts (Fig. 1 A and B).
Minocycline and restraint stress
At 28 days post-CCI, basal levels of CORT were unaffected by minocycline treatment in CCI and sham control males (Table 1). CCI-minocycline treated females had significantly (p = 0.05) higher basal plasma CORT levels than their sham controls, and basal CORT was significantly (p < 0.01) higher in all CCI females irrespective of treatment than in their respective males (Table 1).
In both sexes, there were eight rats per group except in the Sham-Saline groups, in which there were six rats per group.
p ≤ 0.01 for groups with similar superscripts.
Day 28: Analyses of variance (ANOVA) indicated significance in sex (F 1,52 = 21.28, p < 0.0001), and a trend in interaction with group (F 3,52 = 2.77, p = 0.051) but not in group. The CCI-Mino and CCI-Saline females had significantly (a p = 0.01) higher plasma corticosterone (CORT) than males, and the CCI-Mino females had significantly (c p = 0.01) higher plasma CORT than sham-Mino females. Day 35: ANOVA indicated significance in group (F 3,52 = 15.44, p < 0.0001) but not in sex or their interaction. Because all shams were killed later in the day than the CCIs, CORT was significantly higher in the male Sham-Saline group than in the male CCI-Salines (d p < 0.01) and in the male Sham-Minos than in the male CCI-Minos (e p < 0.01). The same pattern held in the females (f p < 0.01 and g p < 0.01).
Minocycline enhanced CCI blunting of the CORT response to 30 min restraint stress in both males and females at 30 min after restraint onset (Fig. 3), as we have repeatedly shown to occur after moderate CCI in male rats. 9,10 In the males, although the 30 min CORT response to restraint stress tended to be lower in the CCI-saline treated animals than their sham controls, minocycline enhanced this difference (p < 0.05). In the females, the CORT response to 30 min restraint stress was significantly suppressed in the CCI females whether treated with minocycline (p = 0.0001) or saline (p < 0.01) at 30 min after stress onset (Fig. 3). Blunting of the CORT response (p < 0.05) persisted in the CCI-SAL females up to 90 min after stress onset (Fig. 3).
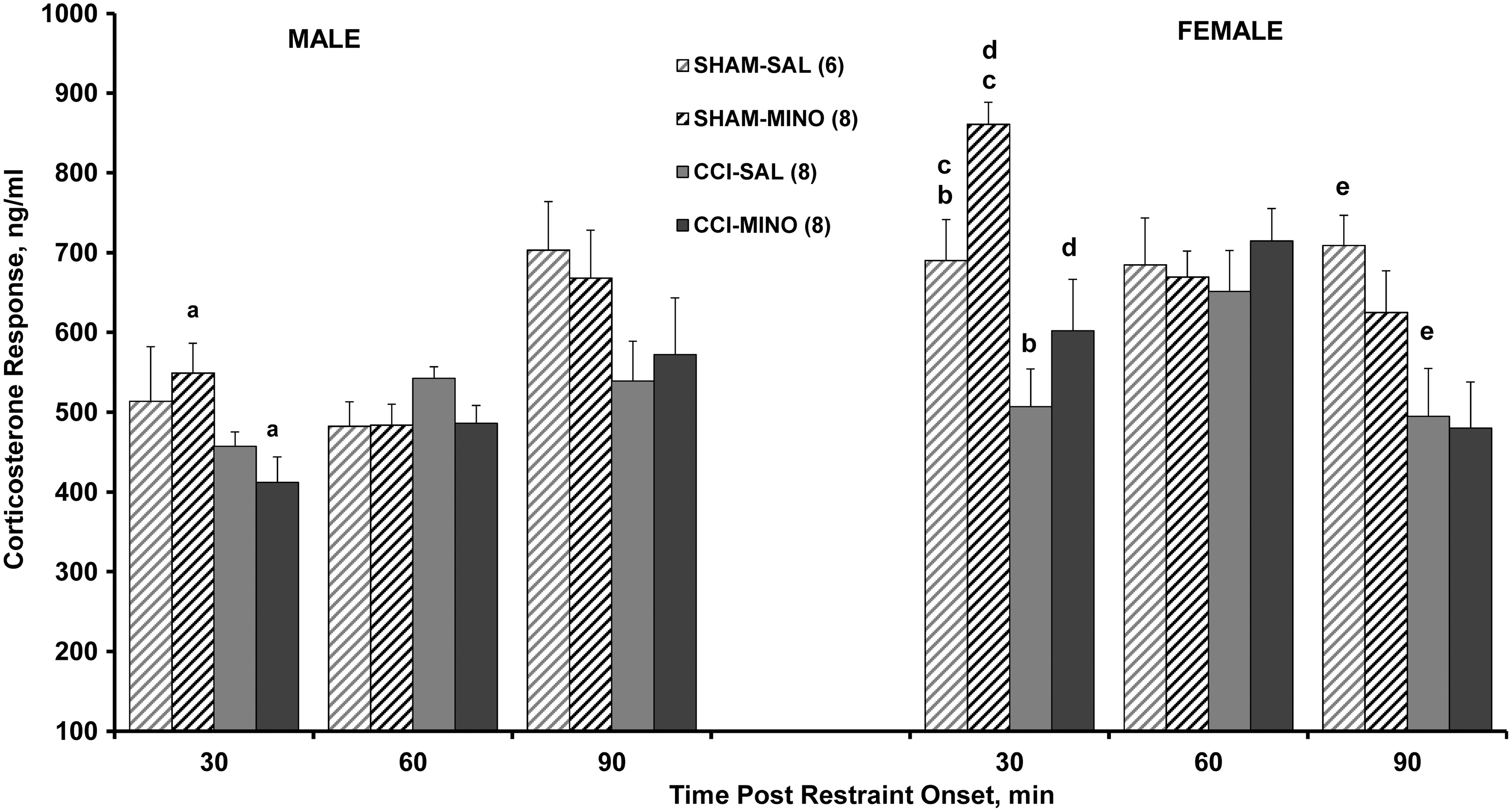
Effect of restraint stress on the corticosterone (CORT) response (mean ± SEM, ng/mL) at 28 days after moderate cortical contusion injury (CCI) or sham surgery in male and ovariectomized female rats treated with saline (SAL) or minocycline (MINO) on Days 1–4 after injury. The numbers in the parentheses indicate the sample size of each group, and are valid for both sexes. At 30 min post-restraint onset, analyses of variance (ANOVA) indicated significant effects of group (F
3,52 = 11.00, p < 0.0001), sex (F
1,52 = 32.86, p < 0.0001), and their interaction (F
3,52 = 3.08, p < 0.05); at 60 min, ANOVA indicated significant effects only of sex (F
1,52 = 49.83, p < 0.0001); and at 90 min, ANOVA indicated significant effects only of group (F
3,52 = 4.78, p < 0.01). The CORT response in CCI-MINO males (
Minocycline and cytokines
On Day 35 post-injury, although there was no effect of minocycline on the terminal plasma CORT levels in either CCI or sham males and females, CORT levels were significantly (p ≤ 0.01) higher in the sham than in the CCI males and females irrespective of post-CCI treatment with minocycline or saline (Table 1). These differences were the result of decapitation and blood collection in the afternoon rather than in the morning as in the CCIs.
On Day 35 after CCI, IL-1β, and TNF-α were extracted from plasma, and IL-6 was extracted from serum, and all three cytokines were extracted from ipsilateral cortical and hippocampal tissues. Levels of cytokines were undetectable in the plasma or serum. Although there were no effects of either injury or minocycline/saline treatment on ipsilateral cortical cytokines, there were marked sex differences in the cytokine levels (Table 2), with cortical IL-1β and TNF-α being significantly higher in males than in ovariectomized females (p < 0.01 and p < 0.0001, respectively), and cortical IL-6 being significantly higher (p < 0.0001) in females than in males (Table 2). In contrast, hippocampal IL-1β and IL-6 were significantly higher (p < 0.01 and p < 0.0001, respectively) in the ovariectomized females than in the males (Fig. 4A and B), whereas TNF-α was significantly lower (p < 0.0001, Fig. 4C).

Interleukin (IL)-1β
In both sexes, there were eight rats per group, except for the Sham-Saline group in which there were six rats per group.
Interleukin (IL)-1β: Analyses of variance (ANOVA) indicated significance in sex (F 1,52 = 9.76, p = 0.003), but not in group or their interaction. Whereas all female levels were lower than male levels, post-hoc t tests revealed a significant (a p = 0.05) sex difference for the sham-salines.
IL-6: ANOVA indicated significance in sex (F 1,52 = 100.08, p < 0.0001), and a trend for its interaction with group (F 3,52 = 2.77, p = 0.051) but not in group. Whereas all female levels were higher than male levels, post hoc t tests revealed significant (b–e p ≤ 0.01) sex differences for each group.
Tumor necrosis factor (TNF)-α: ANOVA indicated significance in sex (F 1,52 = 697.36, p < 0.0001), but not in group or their interaction. Whereas all female levels were lower than male levels, post hoc t tests revealed significant (f–i p < 0.001) sex differences for each group.
In the males, there was no effect of injury or treatment on the levels of these cytokines in hippocampal supernatants. In contrast, in the females, IL-1β and IL-6 extracted from the hippocampus ipsilateral to the injury on Day 35 after CCI showed significant effects of CCI with or without minocycline (Fig. 4 A and B). Hippocampal IL-1β was significantly (p < 0.05) higher in the ovariectomized CCI-minocycline females than in their sham-minocycline controls (Fig. 4A). The same differences (p < 0.001) were found in hippocampal IL-6 (Fig. 4B). Additionally, hippocampal IL-6 was significantly (p < 0.01) higher in the CCI-SAL females than in their SHAM-SAL controls (Fig. 3B). Hippocampal TNF-α levels, as in the males, were unaffected by injury or treatment (Fig. 4C).
Discussion
We herein report several novel findings with respect to moderate CCI with or without minocycline treatment during the first 4 days post-CCI in male and ovariectomized female rats. There is a sex difference in the induction of hyperthermia by CCI, as we reported previously for male rats 8 ; that is, the first 5–6 days of post-CCI hyperthermia in male rats occurred only on the day of CCI in the ovariectomized females (Fig. 1). Further, minocycline prevented the occurrence of post-CCI hyperthermia in both sexes, particularly during the dark phase (Fig. 2). Unlike minocycline's protective action against TBI-induced neurobehavioral effects, 7 in both sexes minocycline facilitated suppression of the CORT response to restraint stress (Fig. 3), which we have identified as a hallmark of moderate CCI at 28 days post-injury in male rats. 9,10 Finally, whereas at 35 days post-injury cortical IL-6 was higher in the ovariectomized females than in the males and cortical IL-1β and TNF-α were higher in the males than in the females, hippocampal IL-1β and IL-6 were higher and TNF-α was lower in the females than in the males (Table 2 and Fig. 4). In the females, but not the males, 4 days of post-injury minocycline or saline treatment augmented hippocampal IL-6 content on Day 35 post-CCI; hippocampal IL-1β content was also augmented in the CCI minocycline-treated females compared with their sham-injured controls (Fig. 4).
One of the first studies that examined the effect of minocycline treatment following experimental TBI showed that this anti-inflammatory tetracycline derivative improved neurological function, decreased lesion volume, and attenuated production of cerebral IL-1β in CCI mice compared with saline control mice. 15 In a closed head injury model of TBI, neuroprotection by minocycline was shown to be associated with microglial activation and reduced IL-1β expression in the injured cortex. 16 In clinical and experimental studies, TBI has been reported to cause prolonged microglial activation leading to a pro-inflammatory state and worse functional outcomes. 17 –19 Further, as recently reviewed, microglia may have sex-specific responses to TBI in experimental models. 20 Although several studies have shown that sex hormones are important mediators of the microglial response after TBI, 21,22 other data from a CCI model indicate that microglial activation in the hippocampus did not differ between males and females and was unaffected by basal levels of estrogen or estrogen supplementation. 23
We have shown previously that moderate CCI in male rats induces hyperthermia that persists for at least 6 days post-injury; 8 female rats were not studied previously. In this study, we monitored body temperature for the first 6 days after moderate CCI in both sexes, and observed hyperthermia throughout most of this period following saline injection in CCI males, but only on the day of CCI in ovariectomized females (Figs. 1 and 2). The CCI-induced fever is one component of the constellation of acute-phase responses to injury that are mediated by the release of pro-inflammatory cytokines immediately after injury. 11,12 As would be expected after treatment with the anti-inflammatory drug minocycline, body temperature after CCI in males and females injected with minocycline did not differ from that in their respective sham controls (Figs. 1 and 2). The sex differences that we observed in body temperature during the first 6 days post-CCI may reflect the differences in male and female IL-1β and TNF-α cytokine mRNA expression levels in CCI mice recently reported by Villapol and colleagues. 24 Their report indicated that male mice presented with a sustained cortical cytokine peak between 1 and 7 days post-injury, whereas in female mice, there was a brief peak in pro-inflammatory cytokines at 4 h post-CCI, and a second peak at 7 days post-injury. 24 On Days 3 and 4 of minocycline treatment in male rats post-CCI, body temperatures were significantly lower than in the CCI-SAL group (Fig. 1A). Therefore, the prevention of post CCI hyperthermia by minocycline in rats of either sex can be added to the documented neuroprotective actions of this drug in TBI. 7 Whereas intact females have been found to display greater endogenous neuroprotection from TBI than ovariectomized females or males, 25 systemic hyperthermia post-CCI has seldom been reported in females. The finding that a brief period of induced hyperthermia after TBI by fluid percussion injury in ovariectomized females worsened histopathological outcome compared with that of intact females suggests that gonadal hormones may protect against secondary hyperthermic insults in TBI. 26 However, factors other than gonadal steroids; for example, microglial activation 20 or hippocampal organization, 27 have yet to have the observed sex differences confirmed.
In addition to attenuation of post-CCI hyperthermia by minocycline, as described, minocycline did not prevent our typically observed CCI-induced suppression of the CORT response to restraint stress at 28 days after injury in males, 9,10 nor did minocycline prevent suppression of the CORT response to restraint stress in the ovariectomized females (Fig. 3). CCI suppression of the stress-induced CORT response at 30 min was enhanced by minocycline in comparison with saline in females (Fig. 3). Minocycline's enhancement of this neurobehavioral parameter may reflect the time that elapsed since the rats had been exposed to minocycline; that is, 24 days. When male rats were exposed to minocycline 1 week after a psychogenic stress, attenuation of anxious-like behaviors and brain cytokine levels has been reported. 28 Moreover, the more robust CORT response in females is not surprising given the well-known differences in HPA responsiveness between males and females. 29,30
We also observed significant sex differences in cortical and hippocampal levels of IL-1β, IL-6, and TNF-α at 35 days post-CCI or sham surgery with or without minocycline treatment in the first 4 days post-injury. Irrespective of injury or treatment, our results indicated higher levels in ipsilateral cortical IL-1β and TNF-α in males than in ovariectomized females, whereas cortical IL-6 showed the opposite pattern (Table 2). In contrast, hippocampal levels of IL-1β and IL-6 were significantly higher in females than in males, and in the females, unlike in the males, were significantly affected by CCI with or without minocycline at this delayed time point (Fig. 4). In an earlier report, Taupin and colleagues showed marked effects of lateral fluid percussion injury in male rats on these cytokines in ipsilateral cortex and ipsilateral hippocampal IL-6 at 1–48 h after the injury. 4 Villapol and colleagues recently reported a delayed cortical anti-inflammatory mRNA peak in females, but not in males, at 30 days post-injury. 24 Here we show long-term effects of CCI that persist to 35 days after the injury on hippocampal IL-6 and IL-1β in ovariectomized female but not in male rats. Glucocorticoids, such as CORT, have been shown to possess potent anti-inflammatory properties, 31 although recent evidence indicates that they can also be pro-inflammatory. 32 Whether the CCI-related sex differences in these cytokines reflect the higher basal levels of corticoids in CCI-females remains to be determined.
Conclusion
In conclusion, protection by minocycline against CCI-induced hyperthermia during the 1st week post-injury in both sexes may be added to its reported protective action against TBI-induced neurobehavioral effects. 7 In contrast, at 24 days after treatment of both sexes, minocycline facilitated CCI-induced suppression of the CORT response to restraint stress, and 1 week later, in ovariectomized CCI females, it augmented hippocampal content of IL-6 and IL-1β. Thus, the short-term protective actions of minocycline on CCI-induced changes in body temperature are not sustained in long-term effects of CCI on neuroendocrine and inflammatory parameters.
Footnotes
Author Disclosure Statement
No competing financial interests exist.