
Abstract
Traumatic brain injury (TBI) has high morbidity and mortality rates. The mechanisms underlying TBI are unclear and may include the change in biological material in exosomes. Circular ribonucleic acids (circRNAs) are enriched and stable in exosomes, which can function as microRNA (miRNA) sponges to regulate gene expression levels. Therefore, we speculated that circRNAs in exosomes might play an important role in regulating gene expression after TBI and then regulate specific signaling pathways, which may protect the brain. We first isolated exosomes from the brain extracellular space in mice with TBI by digestion. We then investigated the alterations in circRNA expression in exosomes by high-throughput whole transcriptome sequencing, analyzed the data by gene ontology (GO) and pathway analysis, and constructed the circRNA-miRNA network. In this study, we identified 231 significantly and differentially expressed circRNAs, including 155 that were upregulated and 76 that were downregulated. GO analysis showed that these differentially expressed circRNAs might be related to the growth and repair of neurons, the development of the nervous system, and the transmission of nerve signals. The most highly correlated pathways that we identified were involved primarily with glutamatergic synapse and the cyclic guanosine monophosphate-protein kinase G signaling pathway. The circRNA-miRNA network predicted the potential roles of these differentially expressed circRNAs and the interaction of circRNAs with miRNAs. Our study broadens the horizon of research on gene regulation in exosomes from the brain extracellular space after TBI and provides novel targets for further research on both the molecular mechanisms of TBI and the potential intervention therapy targets.
Introduction
A
Although the research on TBI currently is gaining momentum, the mechanism of internal damage remains unclear. It is still important to improve the understanding of the molecular and intermolecular interactions and the key signaling pathways that are affected post-TBI.
Extracellular vesicles (EV) are spherical particles that are enclosed by a phospholipid bilayer and are released by both eukaryotic and prokaryotic cells. EV diameters typically range from 30–100 nm. Exosomes are a type of EVm first proposed by Trams and associates 5 in 1981, which were detached from the vesicles exfoliated from the extracellular enzyme activity of cells. Exosomes are secreted by active cells of various types under normal or pathological conditions and found in many types of body fluids such as saliva, serum, plasma, urine, breast milk, amniotic liquid, bronchoalveolar lavage fluid, cerebrospinal fluid, and others. 6 –9
Johnstone and colleagues 6 showed that exosomal densities can range from 1.13 g/mL for B cells to 1.19 g/mL for epithelial intestinal cells in an ultracentrifugation experiment. Exosomes contain not only cellular proteins and lipids but also messenger ribonucleic acids (mRNAs) and microRNAs (miRNAs) from the host cell, which can affect the activity of the recipient cells and become an important medium for cell-cell and cell-extracellular environment communication. 10 Because of their endosomal origin, exosomes bear specific protein markers such as membrane transport and fusion proteins (guanosine triphosphate-ases, annexins, flotillin), tetraspannins (CD9, CD63, CD81, CD82), heat shock proteins (HSP 70, HSP 90), proteins involved in the biosynthesis of multi-vesicular bodies (Alix, tumor susceptibility gene [TSG] 101), lipid-related proteins, and phospholipases. 11 –13
Studies have shown that exosomes have numerous biological functions, such as information transfer, 14 immunoregulation, 15,16 altering tumor microenvironment, and more. 17,18 With the progress of research, exosomes have attracted increasing attention and become the focus of many studies. Recently, Zhang and coworkers 19 demonstrated that mesenchymal stromal cells (MSC)-generated exosomes effectively ameliorate functional recovery by promoting endogenous angiogenesis and neurogenesis, reducing inflammation in rats after TBI. This suggests that exosomes may provide a novel therapy for TBI and possibly other neurological diseases.
Circular RNAs (circRNAs) are a group of nonprotein-coding RNA molecules. 20 Unlike linear RNAs that are terminated with 5' caps and 3' poly (A) tails, circRNAs form covalent-closed continuous loops with neither 5'-3' polarity nor a polyadenylated tail. 21 The covalent-closed loop structure enhances the stability of circRNAs, which renders them resistant to degradation by RNase R. 22 The circRNAs are produced through spliceosomal mechanisms of precursor mRNAs. The circRNAs can derive from exons, 21 introns, 23 or both. 24 Most circRNAs, however, are derived from exons of protein coding genes via “back-splicing.” 25 The circRNAs are characterized by the back-splicing junction.
Growing evidence has suggested that complementary sequences and exon skipping are responsible for the biogenesis of circRNAs. 21,26 Once generated, circRNAs appears to be nonprotein-coding because of the lack of effective start and/or stop codons. Genome analyses have indicated that the majority of circRNAs are conserved and exhibit cell-type or tissue-specific expression. 22,27
The circRNAs have gained more attention because of not only their stabilization in eukaryotic transcriptomes but also their biological functions. In 2011, Hansen and colleagues 28 recognized a highly expressed circRNA in human and mouse brains. It is well known that miRNAs, the post-transcriptional regulators of gene expression, function by direct base pairing to target sites within the noncoding region of mRNA. 29 Recent studies have shown that some circRNAs can play a role as miRNA sponges to regulate the expression levels of other related RNAs by miRNA response elements. 30,31 The circRNAs are capable of making miRNA response elements bind to miRNAs to block the inhibitory action of miRNAs on the expression of their target genes. 32
A recent study indicated that circRNAs are enriched and stable in exosomes. Alterations in circRNA expression are also involved in disease onset. 33,34 For example, a study observed a reduction in circRNAs in tumor tissues from colorectal cancer patients compared with matching normal tissues. 35 Zhong and associates 36 demonstrated that circTCF25 overexpression could downregulate miR-103a-3p and miR-107, increase CDK6 expression, and promote proliferation and migration in bladder cancer by screening differential circRNA expression profiles. Our group reported previously that miR-21 can alleviate apoptosis of cortical neurons and secondary blood-brain damage after TBI in rats. 37,38 Therefore, we speculated that circRNAs in exosomes from the brain extracellular space after TBI might play an important role in regulating gene expression and regulate specific signaling pathways, which may protect the brain.
Aiming to identify the vital circRNAs and provide novel targets for further study, we investigated the alterations in circRNA expression in exosomes from the brain extracellular space after TBI by high-throughput whole transcriptome sequencing. We used bioinformatics to predict the function of differentially expressed circRNAs by gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis. Pathway relation networks and circRNA-miRNA networks were constructed based on key differentially expressed circRNAs to explore the interaction between them. In addition, five circRNAs were selected randomly to confirm the reliability of the microarray results by quantitative reverse transcription-polymerase chain reaction (qRT-PCR). We revealed some potential roles of these circRNAs that were expressed differentially in exosomes from the brain extracellular space after TBI.
Methods
Animals
Experiments were performed using adult male C57BL/6J mice (12–16 weeks old and 22–25 g) that were housed for at least seven days before surgery. Mice were allowed free access to water and food. Water and food were withheld overnight before surgery. All animals were treated in accordance with the principles and procedures of the Laboratory Animal Care and Use Committee of Tianjin Medical University. All protocols were approved by the Tianjin University Administrative Panel on Laboratory Animal Care.
TBI model and experimental groups
In this study, we used the fluid pressure injury (FPI) method to inflict medium-sized injuries in the TBI model. 39 Mice were intraperitoneally anesthetized with 10% chloral hydrate 0.3 mL/100 g. After anesthesia, the head hair was shaved, and the skin was disinfected with iodophor. A midline scalp incision was made followed by a 3.0-mm diameter hole drilled into the skull (2.0 mm posterior from the bregma and 2.0 mm lateral to the sagittal suture) with the dura mater intact. A Plexiglas cylindrical reservoir was connected to the round skull window with glass ionomer cement. Attention was paid to expose the bone window and the Plexiglas cylindrical reservoir was filled with saline. The FPI device (Custom Design & Fabrication, Richmond, VA) was connected after the pinch tail reflex and the mouse TBI model was established according to the standard procedure. Moderate FPI was induced by a swinging pendulum, which was set to an angle of 10 degrees, and the peak pressure was 1.9 ± 0.1 atm. The sham group received an identical surgical procedure but did not receive an FPI.
To model the acute events, all mice were examined within the first 3 h after FPI. After a survival period of 3 h, the mice were anesthetized (10% chloral hydrate 0.3 mL/100g) and transcardially perfused with 100 mL of 4°C phosphate buffered saline (PBS). The traumatically injured brain tissues were obtained in the TBI group, and the corresponding brain tissues were obtained in the sham group.
Twenty-four mice were divided randomly into two groups of 12 mice each. Four random mice brains in the same group were mixed into one sample, so each group had three samples. In total, six samples (three from the TBI group and three from the sham group) were used to isolate exosomes.
Isolation of exosomes
The brain tissues were placed in a culture dish containing the cell culture medium (Hibernate-A, Invitrogen) in a sterile ventilated hood and were dissected. The shredded brain tissues were transferred to a 15 mL centrifuge tube and resuspended in 3 mL of medium. The solution was treated with 20 units/mL of papain (Worthington Biochemical Corp., Lakewood, NJ) for 15 min at 37°C. After discarding the supernatant, 6 mL of 4°C medium was added to stop the digestion. Repeated blowing with a polished fine diameter dropper was used to blow away brain tissues.
Samples were centrifuged for 30 min at 2000 × g, at 4°C to discard cells and membranes, and the supernatant was collected. The supernatant was filtered by passing through a sterile 0.2-μm filter (Thermofisher, Invitrogen). Total exosome isolation reagents (Thermofisher, Invitrogen) were added to the supernatant at the proportion of 1 mL of supernatant to 500 μL of the isolation reagents, and then samples were vortexed to mix well and incubated overnight at 4°C. The supernatant was centrifuged at 10,000 × g for 60 min at 4°C to pellet exosomes. The supernatant was removed as completely as possible, with the suspension consisting of exosomes.
Detection of exosomes
Exosomes were observed under a transmission electron microscope (TEM; Hitachi, HT7700). For an enhanced view of the plasma membrane, exosomes were loaded onto carbon-coated formvar grids for 10 min and stained with phosphotungstic acid (pH 6.8) for 5 min.
Western blot was used to detect typical exosomal markers. The hemi-brains of normal mice were processed for exosome isolation and then proteins were extracted; other hemi-brains were subjected to whole tissue protein extraction and used as controls. Equal amounts of protein were subjected to SDS-PAGE. The sources of antibodies and dilutions used were as follows: mouse anti-CD63 (Abcam, 1:400 dilution), mouse anti-GFAP (Abcam, 1:1000 dilution), rabbit anti-TSG101 (Abcam, 1:1000 dilution), mouse anti-HSP70 (Abcam, 1:1000 dilution), and mouse anti-β-actin (Abcam, 1:1000 dilution), which was used as an internal control.
RNA library construction and circRNA sequencing
Exosomes were resuspended in 500 μL of TRIzol reagent (Life Technologies, Carlsbad, CA) for subsequent detection. Total RNA was isolated from exosome samples using TRIzol reagent according to the manufacturer's instructions. RNA quantification and quality were assessed by a NanoDrop ND-1000. For spectrophotometry, the O.D. A260/A280 ratio should be near 2.0 for pure RNA (ratios between 1.8 and 2.1 are acceptable). The O.D. A260/A230 ratio should be more than 1.8. RNA integrity and DNA contamination were tested by denaturing agarose gel electrophoresis. DNA should be completely absent, and RNA should not be degraded. RNA integrity meets the requirements when RNA Integrity Number (RIN) ≥7.
The sequencing library was determined by Agilent 2100. Total RNA was used for removing the rRNAs using Ribo-Zero rRNA Removal Kits (Illumina, Sa Diego, CA). RNA libraries were constructed using rRNA-depleted RNAs with TruSeq Stranded Total RNA Library Prep Kit (Illumina) according to the manufacturer's instructions. Libraries were controlled for quality and quantity using the BioAnalyzer 2100 system (Agilent Technologies, Santa Clara CA). Libraries (10 pM) were denatured as single-stranded DNA molecules, captured on Illumina flow cells, amplified in situ as clusters, and sequenced for 150 cycles on Illumina HiSeq Sequencer according to the manufacturer's instructions.
circRNA sequencing analysis
High throughput whole transcriptome sequencing and subsequent bioinformatic analysis were both performed in this study. Briefly, paired-end reads were harvested from the Illumina HiSeq 4000 sequencer. After 3' adaptor-trimming and low-quality reads were removed by the cutadapt software (v1.9.3), the high-quality reads were aligned to the reference genome/transcriptome with STAR software. The circRNAs were detected and identified with DCC software 40 and then annotated with the circBase database (mouse MM9). We converted the coordinates of the MM9 to the MM10 using the UCSC liftover tool. Raw junction reads for all samples were normalized by the total number of mapped reads and log2 transformed. Differentially expressed circRNAs were identified by T tests and the fold change between the two groups. The p values were corrected by using multiple hypothesis testing (BH method). circRNAs exhibiting fold changes ≥2.0 with p values ≤0.05 were classified as significant.
According to the previously reported method,
41
the coordinates of the mRNA (which is defined as the circRNA-associated gene) were found through the RefSeq database based on back-splicing site coordinates of the circRNA. GO (
The top 10 significant pathways of upregulated circRNAs and downregulated circRNAs were chosen to construct the pathway relation network, which were based on the interaction data in KEGG. The pathway relation network was used to identify the regulating effect of these pathways. According to the specific complementarity of the chain, for exonic circRNAs, the genomic sequence of exons of the circRNA coordinates was spliced; for nonexonic circRNAs, the genomic sequence between the circRNA coordinates was obtained directly. After obtaining the predicted sequence of the circRNA, circRNA and miRNA interactions were predicted using customized Arraystar miRNA target prediction software based on Target Scan and miRanda. 42,43 The miRNA binding sites on differentially expressed circRNAs and the top five putative target miRNAs for differentially expressed circRNAs were identified. The circRNA-miRNA network was constructed using cytoscape software.
qRT-PCR analysis
To verify the accuracy of the circRNA-seq data, qRT-PCR was performed as a validation method. The target RNA and internal parameters of each sample respectively were subjected to RT- PCR reactions on an Applied Biosystems 7500 Fast Real-Time PCR System (Roche, Basel, Switzerland; software version 2.0.5) with SYBR Green qPCR SMix (ROX; Roche). Total RNA was reverse transcribed to synthesize cDNA using a Prime Script RT Reagent Kit (Perfect Real Time; TaKaRa, Osaka, Japan). The data were analyzed by the 2-ΔΔCT method. An outward-facing primer is designed typically for an exon sequence near the back-splice site (within 150 nt up or down) of the circRNA.
All procedures were performed as follows: 95°C, 10 min; 40 PCR cycles (95°C, 10 sec; 60°C, 60 sec [fluorescence collection]). After the amplification reaction was finished, the procedure was performed as follows: 95°C, 10 sec; 60°C, 60 sec; and 95°C, 15 sec. The temperature was slowly increased from 60°C to 99°C (Automatic instrument, Ramp Rate 0.05°C/s) to establish the melting curve of the PCR product. The primers of the five randomly selected circRNAs and internal parameters of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) are shown in Table 1.
GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Statistical analysis
All statistical analyses were performed in SPSS 21.0. Data were expressed as the mean ± standard deviation (means ± SD). Student t test was used to assess differences between two groups. A p value <0.05 was considered a significant difference.
Results
Detection of exosomes
TEM and Western blot analysis were used to identify the exosomes that were obtained. Under TEM, these exosomes were irregular spheres ranging 30–100 nm in diameter, with a clearly defined and relatively intact membrane (Fig. 1A). As Western blot analysis shows in Figure 1B, the exosomes expressed the typical exosomal markers such as CD63, TSG101, and HSP70. There was no expression of CD63 and TSG101 and low expression of HSP70 in the brain tissue, however, which may be the result of the low content in whole brain tissue. Glial fibrillary acidic protein (GFAP) is a marker of astrocytes, which these exosomes expressed, confirming they came from brain tissues. There was no expression of β-actin in these exosomes.

Detection of exosomes. (
Differential expression of circRNAs in exosomes from the brain extracellular space after TBI
The circRNAs were sequenced in three TBI and three normal exosome samples from the brain extracellular space. The circRNA-seq reads of each sample are shown in Table 2. We identified 231 significantly differentially expressed circRNAs that met the criteria of exhibiting fold change ≥2.0 and p values ≤0.05. Of these, 155 were upregulated (Supplementary Table 1; see online supplementary material at
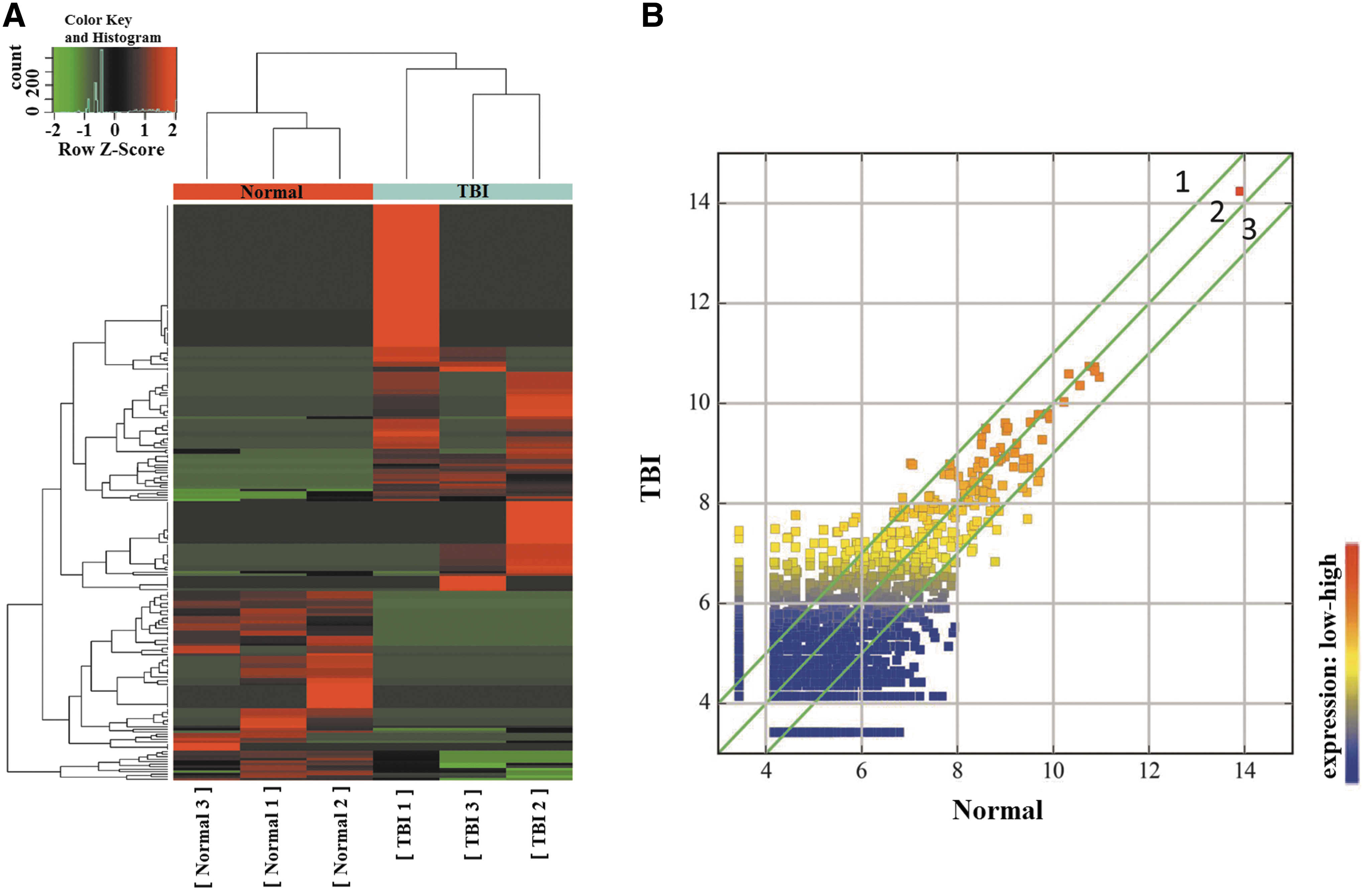
Differential expression of circular ribonucleic acids (circRNAs) in exosomes from the brain extracellular space after traumatic brain injury (TBI).
RNA, ribonucleic acid; TBI, traumatic brain injury.
GO analysis and KEGG pathway analysis of differentially expressed circRNA derived genes
The circRNA-derived genes were used for GO function analysis to annotate and speculate the function of these circRNAs (Fig. 3). GO analysis is divided into three parts: biological process (BP), cell component (CC), and molecular function (MF). GO analysis of BP showed that differentially expressed circRNAs were significantly associated with regulation of dendrite development, transport, positive regulation of CC and process, CC organization or biogenesis, positive regulation of biological process, nervous system development, and single-organism process. Significant GO CC terms of differentially expressed circRNAs showed that these circRNAs were associated with cytoplasm, organelle, cell projection, intracellular part, neuron part, synapse part, neuron projection, cell part, and intracellular organelle. For MF, these circRNAs were associated with the binding of specific substances such as protein, enzyme, kinase, protein kinase, metal ion, cation, and adenosine triphosphate (ATP).
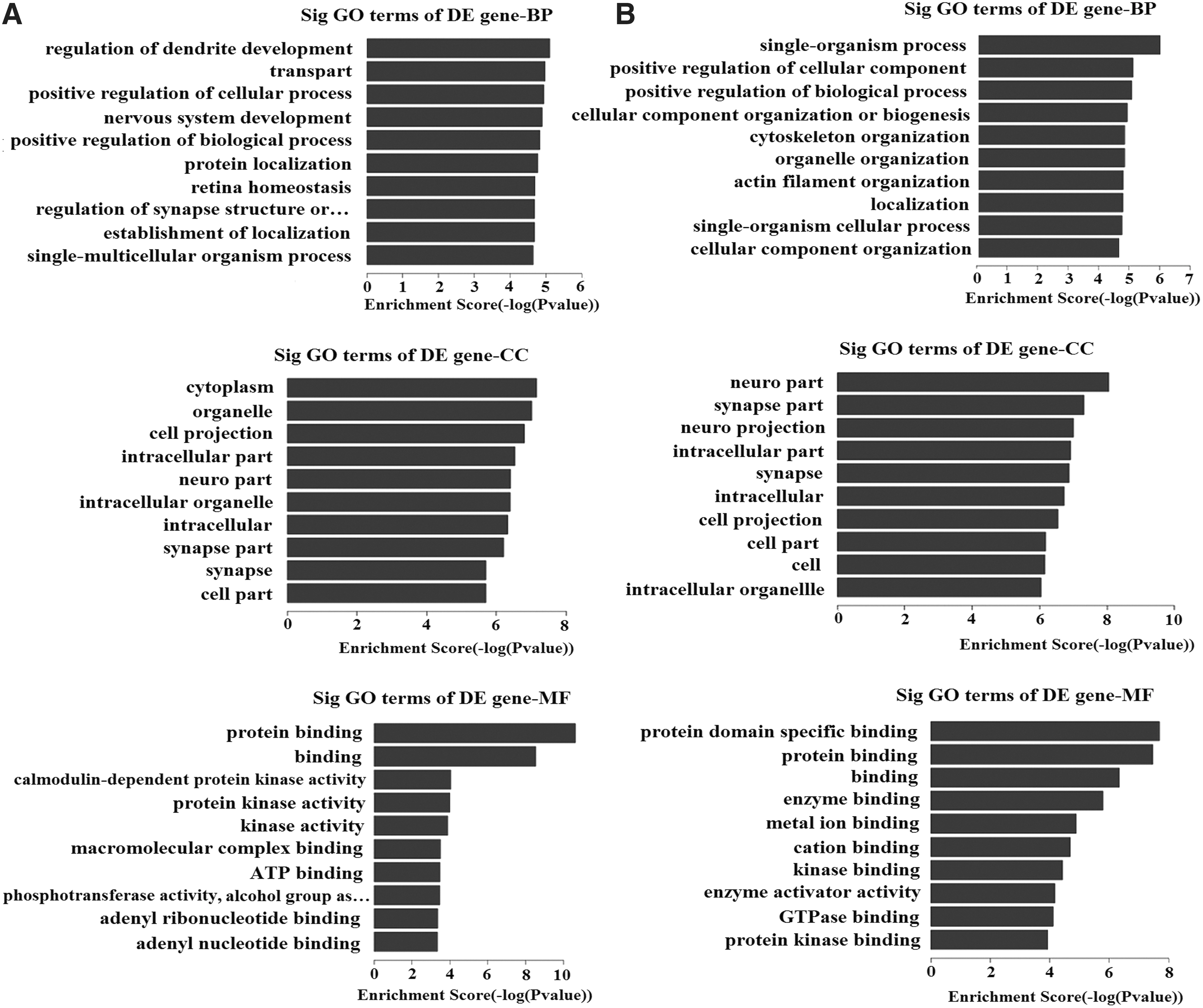
Gene ontology
The KEGG pathway dot plot shows the significant enrichment pathway with the top 10 enrichment score (-log10 (p value)) values. KEGG pathway analysis predicted the pathways affected by the variation of circRNAs in exosomes from the brain extracellular space after TBI (Fig. 4). The top 10 pathways of upregulated circRNAs were glutamatergic synapse, long-term potentiation, amphetamine addiction, oxytocin signaling pathway, amyotrophic lateral sclerosis, ubiquitin mediated proteolysis, Fc epsilon RI signaling pathway, calcium signaling pathway, B cell receptor signaling pathway, and cardiac muscle contraction. The glutamatergic synapse pathway had the highest enrichment score among these signaling pathways.

Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of differentially expressed circular ribonucleic acids (circRNAs) with top 10 enrichment score. The enrichment score was calculated as -log10 (p value). Selection counts represent the number of entities of the differentially expressed genes directly associated with the listed Pathway ID.
The top 10 pathways of downregulated circRNAs were the cyclic guanosine monophosphate-protein kinase G (cGMP-PKG) signaling pathway, citrate cycle pathway, thyroid hormone signaling pathway, N-glycan biosynthesis, lysine degradation, synaptic vesicle cycle, renal cell carcinoma, Fc epsilon RI signaling pathway, cAMP signaling pathway, and salivary secretion. The cGMP-PKG signaling pathway had the highest enrichment score among these signaling pathways. The pathway relation network of the top 10 significant pathways of upregulated circRNAs and downregulated circRNAs showed that the calcium signaling pathway had a regulating effect on upstream levels (Fig. 5).
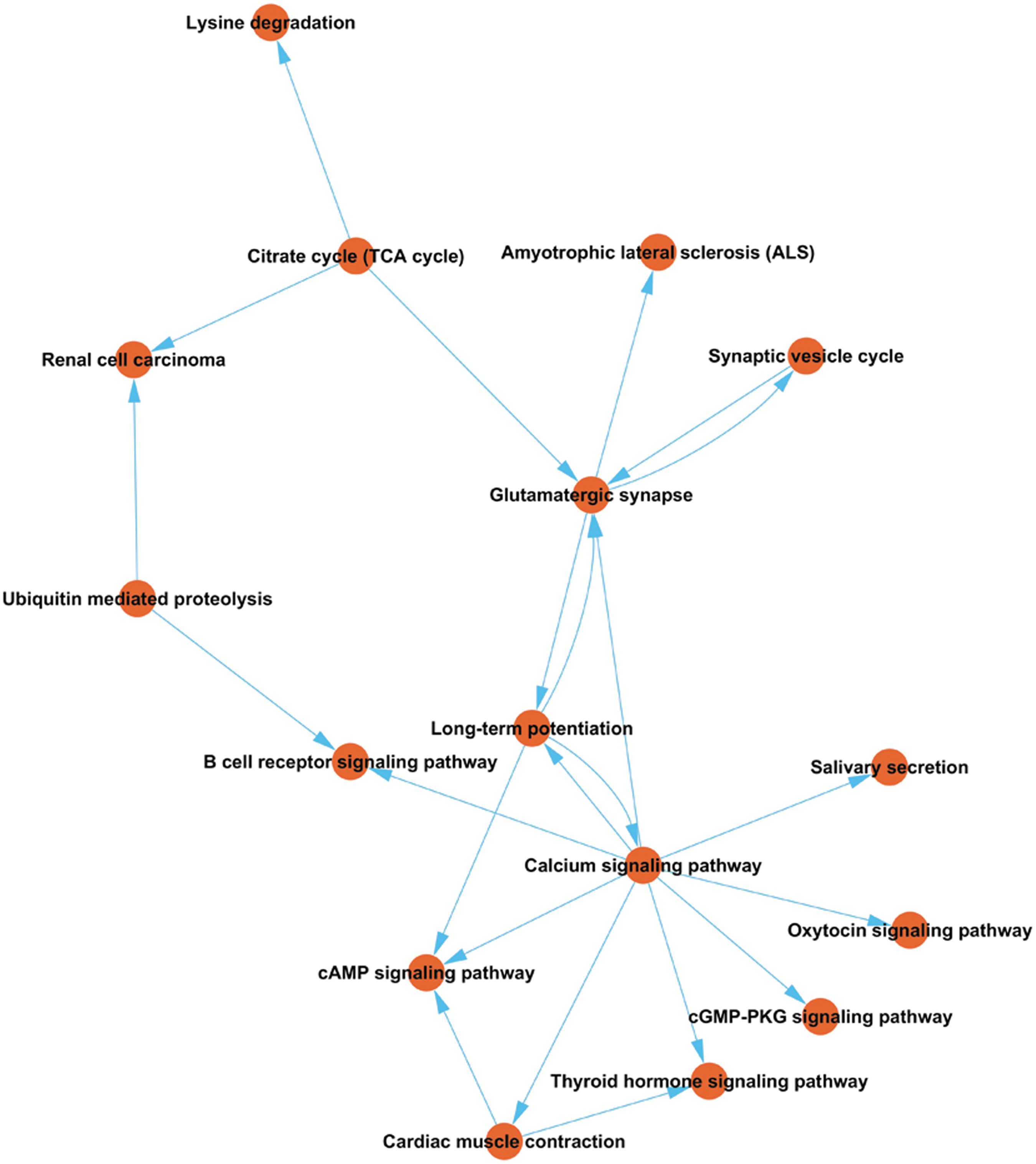
Pathway relation network analysis of the top 10 significant pathways of upregulated circular ribonucleic acids (circRNAs) and down-regulated circRNAs. The network was established based on the results of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and KEGG database search.
Identification of circRNA-miRNA network
Bioinformatic analysis has shown that some circRNAs can play a role as miRNA sponges to regulate the expression levels of other related RNAs by miRNA response elements. Therefore, it is important for us to identify the interaction of circRNAs and miRNAs. The miRNA binding sites on differentially expressed circRNAs are listed in Supplementary Table 3; see online supplementary material at

Circular ribonucleic acids (circRNA)-micro ribonucleic acids (miRNA)-network analysis. circRNAs: yellow, round; miRNA: green, triangular.
Validation of the accuracy of circRNA-seq data by qRT-PCR
Five differentially expressed circRNAs (chr10: 5403841-5426275-, chr8: 83937687-83938400+, chr12: 116871936-116901047+, chrX: 143709648-143744001+, and chr5: 115655901-115663886-) were selected randomly for validation. The target circRNA expression levels were normalized to their internal control, GAPDH. The expression levels of these circRNAs are shown in Table 3. The expression levels of these circRNAs were calculated as 2- ΔΔ CT. The data were expressed as the means ± SD. There were significant differences in all of the selected circRNAs (p < 0.05) except chr8: 83937687-83938400+. The objective circRNA validation rate was 4/5, showing that the circRNA expression profiles were reliable.
p value <0.05 vs Sham group, Student t test (two-tailed).
Discussion
The altered expression profile of circRNAs in exosomes from the brain extracellular space after TBI in mice was studied for the first time by high-throughput whole transcriptome sequencing and subsequent bioinformatic analysis. We made several novel observations that improve the understanding of post-TBI molecular and intermolecular interactions as well as the key signaling pathways they participate in.
First, we demonstrated that traumatically injured brain tissues could release exosomes to the brain extracellular space. We used the digestion method to isolate exosomes, such as the neuron culture, in which the aim was not to damage the cells. These exosomes were then confirmed by TEM and Western blot analysis (Fig. 1). The change in biological material in exosomes from the brain extracellular space can actually reflect the physiological and pathological process after TBI and may be the potential target of TBI therapy.
The circRNAs, one class of noncoding RNA molecules that can regulate the expression of miRNAs, are enriched and stable in exosomes and are involved in the onset of many diseases. 26,27 In this study, we found a total of 231 circRNAs in exosomes that are expressed differentially, including 155 upregulated and 76 downregulated circRNAs after TBI. These altered circRNAs likely participate in the progress of TBI.
Second, to gain insight into the potential function of the differentially expressed circRNAs, GO analysis and KEGG pathway analysis were performed. Gene ontology (GO) is a bioinformatic concept that unifies the genes and gene products of all species. 44 We used the circRNA-derived genes for GO function analysis to annotate and speculate the function of these circRNAs.
GO analysis of molecular function showed that the differentially expressed circRNAs were significantly associated with the binding of specific substances such as protein, enzyme, kinase, and protein kinase. Significant GO CC terms of differentially expressed circRNAs showed that these circRNAs were associated with neuron part, synapse part, cell part, intracellular part, and intracellular organelle. For biological process, these circRNAs were associated with the regulation of dendrite development, positive regulation of cellular component and process, CC organization or biogenesis, positive regulation of biological process, and nervous system development. Thus, we suspected that these differentially expressed circRNAs might be related to the growth and repair of neuron, the development of the nervous system, and the transmission of nerve signals.
KEGG pathway analysis revealed that the differentially expressed circRNAs participated in several biological pathways. Of these pathways, the citrate cycle pathway, thyroid hormone signaling pathway, and cAMP signaling pathway are related to metabolic processes. The most highly correlated pathways that we identified were involved primarily with glutamatergic synapse and the cGMP-PKG signaling pathway. Based on pathway analysis, we constructed a pathway relation network. The network indicated that the most important core upstream pathway was the calcium signaling pathway.
Many cellular functions are regulated by calcium signals, which are generated by different signaling pathways. Changes in the nature of both the primary and modulatory roles of calcium signaling are contributing factors responsible for the onset of a large number of human diseases. The calcium signaling pathway regulates both the proliferation and differentiation of brain cells.
45,46
As neurons develop, axons grow outward to interact with other neurons, and the calcium signaling pathway contributes to the mechanism that steers the growth cones toward their targets.
47
The diagram of the calcium signaling pathway suggests that calcium signaling may regulate metabolism, the mitogen-activated protein kinase (MAPK) signaling pathway, apoptosis, exocytosis secretion and other signaling pathways (Supplementary Fig. 1; see online supplementary material at
Recent studies have found that pathological axonal death after injury was associated with the activation of the MAPK signaling pathway, 48 known as MAPK signaling pathway mediated TBI injury-induced apoptosis in astrocytes. 49 Thus, we infer that these differentially expressed circRNAs may play an important role in regulating metabolism, apoptosis, and exocytosis secretion. The primary TBI-related downstream signaling pathways of the calcium signaling pathway shown on the pathway network are the thyroid hormone signaling pathway and B cell receptor signaling pathway (Fig. 5). Most of the downstream pathways are related to metabolism and immunoreaction.
The thyroid hormone signaling pathway is essential for growth, development, and energy metabolism of mammals. The primary downstream signaling pathways of the thyroid hormone signaling pathway shown on the diagram are the hypoxia-inducible factor–1 (HIF-1) signaling pathway, Wnt signaling pathway, and Notch signaling pathway (Supplementary Fig. 2; see online supplementary material at
Previous studies have shown that the Wnt and Notch signaling pathways play an important role in the proliferation and differentiation of neural stem cells. These signaling pathways can activate the proliferation and differentiation of neural stem cells through the generation of stem cell regulatory factors, hence promoting neural regeneration and repair. 53,54
The B cell receptor signaling pathway, an important immune-related pathway, impacts the development and maintenance of B cells. The primary downstream signaling pathways of the B cell receptor signaling pathway shown on the diagram are the PI3K-AKT signaling pathway, NF-
The NF-
Third, miRNAs are highly conserved during evolution and negatively regulate target mRNAs. Recent functional research found that circRNAs could harbor miRNA binding sites and function as miRNAs sponges. 31 The interaction of circRNAs with disease-associated miRNAs indicates that circRNAs play an important role in the pathology of disease. To date, few studies have focused on the interaction of circRNAs and miRNAs in exosomes from the brain extracellular space.
Here, miRNA binding sites on differentially expressed circRNAs and the top five putative target miRNAs for differentially expressed circRNAs were identified. These data will provide reference for the future study of specific circRNA-miRNA interactions. The circRNA-miRNA network is able to help us further understand the role of circRNAs in regulating the expression of specific genes. The functional effect of the interaction of specific circRNA-miRNAs will be the focus of our future research.
Conclusion
The present study identified a series of differentially expressed circRNAs in exosomes from the brain extracellular space after TBI in mice for the first time, which may be related to the physiological and pathological processes after TBI. We also predicted the potential roles of these differentially expressed circRNAs and the interaction of circRNAs with miRNAs. Our study could broadens the horizon of gene research in exosomes from the brain extracellular space after TBI and provides novel targets for further research on both the molecular mechanisms of TBI and the potential intervention therapy targets.
Footnotes
Acknowledgments
This study was supported by the National Science Foundation of China (81501057) and the Tianjin Research Program of Application Foundation and Advanced Technology (grants 16JCQNJC10600 and 15JCQNJC11300).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.