
Abstract
Although the majority of traumatic spinal cord injuries (SCIs) take place at the cervical level, pre-clinical studies have been disproportionally focused on thoracic insults. With differences in anatomy, physiology, and immune response between spinal cord levels, there is evidence that injury pathophysiology may vary, requiring tailored treatment paradigms. Further, as only a few therapies have been successfully translated to the clinic, cervical models are increasingly recognized as essential for the characterization of trauma and therapy. Using a novel and clinically relevant cervical contusion-compression mouse model of bilateral incomplete injury, this study aimed to assess the role of interleukin10 (IL-10), a potent cytokine with broad anti-inflammatory effects, in SCI vascular pathology. While the effects of IL-10 loss have been previously evaluated, the vascular changes are poorly characterized. Here, using in vivo high-resolution ultrasound imaging, we demonstrate that IL-10 deficiency is associated with increased acute vascular damage. Importantly, the loss of endogenous IL-10 led to significant differences in the acute systemic response to SCI, specifically the circulating levels of IL-12 (p70), LIX (CXCL5), IL-1β, tumor necrosis factor (TNF)-α, and IL-6 relative to genotype sham controls. These effects also fostered modest impairments in long-term functional recovery, assessed by the Basso Mouse Scale, as well as histological outcomes.
Introduction
Traumatic spinal cord injury (SCI) leads to significant motor, sensory, and autonomic dysfunction. With limited treatment options, the management and care of SCI patients presents a major burden for patients and caregivers. Importantly, over 60% of injuries take place at the cervical level 1,2 and have an estimated lifetime cost of care at $1.1-$4.2 million per patient. 3 As a result, the translational importance of these injuries has been a driving force in the development and characterization of cervical SCI models in mice, 4 –7 rats, 8 –10 swine 11 as well as non-human primates. 12,13 Yet, aside from sharp transection models, which are not representative of most human SCI, few studies have applied genetic deletions to assess the mechanisms involved in cervical SCI pathophysiology. 14
SCI involves two distinct phases of tissue damage, known as primary and secondary injury. Following primary injury, resulting from mechanical trauma, a series of secondary biochemical and pathological changes exacerbate spinal cord tissue loss. Vascular pathology, which involves reduced blood flow, loss of microcirculation, blood–spinal cord barrier disruption, and hemorrhage, is a key mediator of secondary injury.
15
As one of the first steps in a complex sequela, vascular damage is directly related to subsequent pro-apoptotic signaling, release of cytotoxic factors, and inflammatory cell infiltration into the spinal cord. Importantly, various strategies shown to preserve vasculature also limit injury severity and improve recovery post-SCI. Many of these promising neuroprotective therapies, such as glibenclamide (DiaBeta,
Interleukin 10 (IL-10) has been implicated in the maintenance of vascular function and shown to counteract hypertension 16,17 as well as aging-associated endothelial dysfunction. 18 As a potent cytokine with broad anti-inflammatory effects, IL-10 has a multitude of cell sources and targets, many of which are associated with various pathologies, including rheumatoid arthritis, inflammatory bowel disease, multiple sclerosis, and systemic lupus erythematosus, 19 among others. In the context of SCI, there is considerable evidence that exogenous administration 20 and over-expression 21 of IL-10 improves outcomes, while deficiency propagates damage. 22,23 However, not all observations have demonstrated long-term functional recovery. 24 Age-dependent reductions in IL-10 expression also have been reported to exacerbate injury and impair functional recovery, specifically through changes in macrophage activation. 25 Relatedly, our previous work found a correlation between increased systemic levels of IL-10 and vascular protection following cervical SCI in the rat. 26
Expanding on this previous study, we aimed to assess the role of IL-10 in SCI vascular pathology using a novel and clinically relevant cervical contusion-compression mouse model of bilateral incomplete injury. 7 While the effects of IL-10 loss have been evaluated in SCI, 22,23 the vascular changes have been poorly characterized. Therefore, there is a need to evaluate the impact of genetic deletions on cervical level pathophysiology, which will encourage better translation of pre-clinical data to the most common type of traumatic SCI in patients. 27 Here, we demonstrate that IL-10 deficiency is associated with increased acute vascular damage, and modest impairments in long-term functional recovery and histological outcomes. Further, characterization of the acute systemic cytokine response following cervical SCI highlighted significant differences in expression of IL-12 (p70), LIX (CXCL5), IL-1β, tumor necrosis factor (TNF)-α, and IL-6 between IL-10 knockout (KO) and wild type (WT) mice (when normalized to genotype sham surgeries). These compensatory changes may account for the limited effects on animal function.
Methods
Clip-compression spinal cord injury
All animal experiments were approved by the animal care committee at the University Health Network (Toronto, Ontario) in compliance with the Canadian Council on Animal Care. Adult (10- to 11-week-old) C57BL/6J (WT) or IL-10 KO (B6.129P2- tm1Cgn /J) mice (18-21 g) were placed under anesthesia, using inhalant isoflurane (1-2%) delivered in a 1:1 mixture of O2/N2O. The incision area was shaved and cleaned. A combination of 0.05 mg/kg buprenorphine and 1 mL of saline was administered prior to beginning the surgical procedure. Following a midline skin incision at the lower cervical and upper thoracic area, a two-level laminectomy (C7-T1) was performed with micro-scissors. The C7 spinal level was subsequently compressed for 40 sec with a 5.2-g modified aneurysm clip, generating a bilateral contusion-compression injury. Post-operatively, animals were housed in standard mouse cages with absorbent bedding (at 26°C-27°C) and amoxicillin trihydrate/clavulanate potassium (Clavomox) delivered in drinking water. Their bladders were manually emptied twice daily until the return of normal bladder function.
In vivo very high-resolution ultrasound and Power Doppler imaging
In vivo very high-resolution ultrasound (VHRUS) and Power Doppler imaging were performed as previously described with minor modifications. 28 In short, at 1 day and 5 weeks post-SCI, mice were anesthetized and placed on an imaging platform (Vevo Imaging Station, VisualSonics, Toronto, Ontario). The injury was exposed through a midline incision and ultrasound gel (scanning gel, Medi-Inn, Cameron, Ontario) was placed on the dura mater. The spinal cord was scanned with the VHRUS probe (44 MHz, Vevo 770; VisualSonics) in three-dimensional (3D) Power Doppler and B-modes. The 3D B-mode and Power Doppler scans were analyzed by using ImageJ software as previously described. 26
Serum cytokine/chemokine assessment with luminex assay
Prior to transcardial perfusion, mice were anesthetized using inhalenet isoflurane and blood samples were collected via cardiac puncture. The blood was left to coagulate for 30 min, then spun down at 1500 rpm for 5 min, after which the supernatant was collected and spun again at 10,000 rpm (centrifuge 5810R, Eppendorf) for another 5 min. A mouse cytokine/chemokine 31-Plex (MD31) luminex array was used to determine the expression levels of selected cytokines from the serum supernatant (Eve Technologies, Calgary, Alberta). Blood samples were collected at 24 h post-SCI. Cytokines outside the range of detection were removed from analysis. The injured groups were normalized to their genotype controls (sham surgeries) and a heat map was generated using the BROAD Institute's R implementation of Morpheus with hierarchical clustering by average Euclidean distance.
Intraparenchymal hemorrhage quantification with Drabkin's assay
The extent of hemorrhage and thereby the degree of intraparenchymal hemoglobin post-SCI was quantified as previously described. 29 Briefly, following transcardial perfusion with 30 mL of phosphate-buffered saline (PBS), 5 mm of the spinal cord (containing the lesion epicenter) was isolated, weighed, and homogenized in 200 μL of distilled water. Following a spin at 3,000 rpm for 15 min, 50 μL of supernatant was added to 100 μL of Drabkin's reagent (Drabkin's Reagent powder [D5941]; Sigma-Aldrich] in 1,000 mL of distilled H2O and 0.5 mL of 30% Brij 35 Solution) in a clear 96-well plate. The samples were incubated for 15 min and absorbance was read at 560 nm (Wallac 1420 VICTOR2; PerkinElmer). The concentration was calculated from a linear bovine blood hemoglobin (H2500; Sigma-Aldrich) standard curve and normalized to wet tissue weight.
Basso Mouse Scale score and sub-score for neurobehavioral assessment
Locomotion was assessed by using the 11-point (0-10) Basso Mouse Scale (BMS) as previously described. 30 Mice were placed in an open field and scored daily for the first week post-injury, followed by weekly assessment for a total of 5 weeks by two observers blinded to the experimental groups. To supplement the BMS score with fine details of locomotion, a cumulative sub-score also was obtained at 5 weeks post-SCI. 30
Automatic gait analysis (CatWalk)
Mouse gait was evaluated using the automated CatWalk XT 10.6 multivariate system (Noldus Information Technology, Wageningen, the Netherlands) at 2-5 weeks post-SCI. Successful performance required three complete crossings with at least three consecutive step cycles for analysis. Paw prints were labeled by an observer blinded to the experimental groups.
Tissue processing and immunohistochemistry
Mice were perfused with 30 mL of 1% PBS, followed by 30 mL of 4% paraformaldehyde (PFA) in 1% PBS (pH 7.4). The spinal cord was dissected and incubated for 4 h within a 10% sucrose solution in 4% PFA and subsequently overnight in a 30% sucrose solution at 4°C. Next, a 1.5 cm area spanning the lesion epicenter was embedded in M-1 medium (Thermo Scientific). Cryosections were cut at 30 μm and mounted on Fisherbrand® Superfrost Plus charged glass slides (Fisher Scientific).
For immunohistochemistry, the slides were dried and rinsed with 1 × PBS and blocked (blocking solution; 2% FBS and 0.1% Triton X in 1 × PBS) for 1 h at room temperature. Fluorescein isothiocyanate (FITC)–conjugated lectin from Lycopersicon esculentum (LEA, L0401 Sigma) was diluted (1:500) in blocking solution and left overnight at 4°C. After three washes (15 min), the slides were cover-slipped and images were taken at × 20 on the Zeiss Axioplan 2 fluorescence microscope (Zeiss, Stuttgart, Germany).
Lesion morphometry
Cross-sectional (30 μm) serial sections (n = 6 per group) were stained with Luxol fast blue (LFB) and hematoxylin and eosin (H&E). Specifically, after 30 min of drying at room temperature, slides were rinsed in 1% PBS for 10 min and subsequently dipped in 50% ethanol (EtOH). Following 4 h in LFB solution at 56°C and cooling, slides were washed with 95% EtOH and distilled H2O for 5 min each. A 0.05% LiCO2 solution was used for de-staining with 70% EtOH to stop the reaction. After 20 min in hematoxylin solution, slides were washed with H2O, dipped into eosin (5 sec) and dehydrated with 95% and later 100% EtOH prior to xylene. Sections were systematically sampled at every 250 μm over 2500 μm, and unbiased measurements were made with a Cavalieri probe using Stereo Investigator (MBF Bioscience, Williston, VT) for lesional tissue, gray matter, and white matter percent area (normalized to the area of spinal cord tissue) by an individual blinded to the experimental groups.
Statistical analysis
Quantitative data are expressed as the mean ± standard error of the mean (SEM). Differences between groups were assessed by unpaired t-test as well as one- and two-way analysis of variance (ANOVA) with Tukey's post hoc tests (statistically significant at p ≤ 0.05). Eve Technologies mouse cytokine/chemokine 31-Plex (MD31) luminex array results were normalized to genotype controls (sham surgeries) and assessed by multiple t-tests with Holm-Sidak correction for multiple comparisons. All other statistical tests applied are mentioned in corresponding figure legends. Data were analyzed with GraphPad Prism (GraphPad Software Inc., La Jolla, CA).
Results
IL-10 deficiency results in greater acute vascular pathology following cervical SCI
The effect of endogenous IL-10 deficiency on acute vascular damage was assessed in WT and IL-10 KO mice (Fig.1). We used Drabkin's assay for quantification of intraparenchymal hemorrhage and VHRUS imaging for measurement of lesion volume and functional vascularity (Fig. 2). At 1-day post-SCI, there was 206.0 ± 6.4 μL of blood per mg of wet tissue in WT injured controls compared with 289.9 ± 41 μL of blood per mg of wet tissue in IL-10 KO animals (n = 5 per group, unpaired t-test, p = 0.056). The lesion volume, as measured by VHRUS, was significantly greater (unpaired t-test, p < 0.05) in IL-10 KO relative to WT mice, with volumes of 1.49 ± 0.09 mm3 and 1.16 ± 0.09 mm3, respectively. Although there was a trend, the difference in functional vascularity (active blood flow detected by Power Doppler) was non-significant (unpaired t-test, p = 0.068), with a 54.12 ± 9.2 % of sham Doppler signal in the WT group compared with a 29.35 ± 6.2% in IL-10 KO animals. The raw area of Power Doppler signal values (without normalization to genotype controls), at 1 day post-SCI, are included in Figure 8B.

Schematic of the experimental design. In short, adult C57BL/6 (wild type [WT]) or IL-10 (knockout [KO]; B6.129P2-Il10 tm1Cgn /J) mice (18-21g) received a C7 5.2g clip compression injury. Spinal cord changes in lesion volume, functional vascularity and hemorrhage were assessed at 1 day post- spinal cord injury (SCI; acute). Neurobehavioral testing (n = 9 for C57BL/6 WT sham; n = 9 for C57BL/6 WT SCI; n = 10 for IL-10 KO sham; and n = 10 for IL-10 KO SCI) was applied daily for the first 7 days post-SCI, followed by weekly assessment up to 5 weeks after SCI by examiners blinded to the experimental groups. Locomotion was examined with the 11-point (0-10) Basso Mouse Scale (BMS). Gait parameters were measured with the Noldus automated CatWalk multivariate system.

Loss of endogenous interleukin (IL)-10 increases acute lesion volume and reduces functional vascularity at 1 day following cervical spinal cord injury (SCI). Blood volume (intraparenchymal hemorrhage) was quantified via Drabkin's assay (n = 5 per group) at 1 day post-SCI and normalized to wet tissue weight
IL-10 deficiency has modest impairments in long-term functional recovery following cervical SCI
Functional recovery was assessed for 5 weeks following SCI using BMS scoring (Fig. 3) and CatWalk automated gait assessment (Fig. 4). Locomotion, assessed through BMS scoring, was significantly different between injured WT and IL-10 KO mice (n = 9-10 per group, two-way ANOVA, p < 0.05). At 5 weeks post-SCI, WT and IL-10 KO animals had BMS scores of 5.5 ± 0.4 and 4.9 ± 0.3, respectively. There was also a difference (Tukey's post hoc test, p = 0.012) in WT (4.7 ± 0.4) and KO (3.4 ± 0.3) cumulative sub-scores. Interestingly, this did not translate into quantifiable changes between the two groups in gait parameters measured by the Noldus CatWalk system.
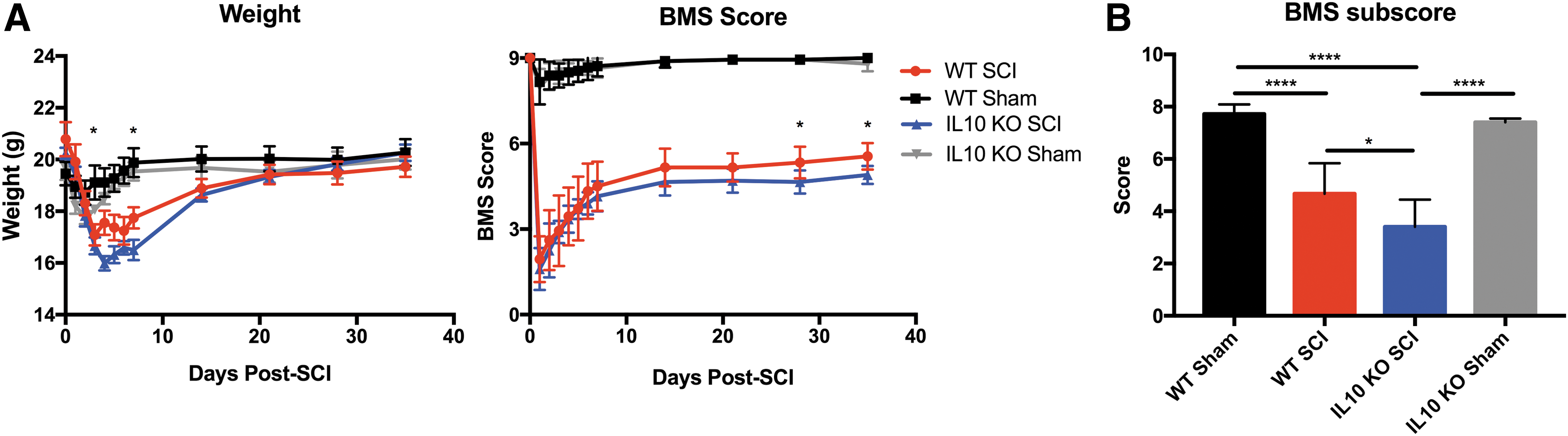
Loss of endogenous interleukin (IL)-10 impairs functional recovery following cervical spinal cord injury (SCI). Weight and locomotion
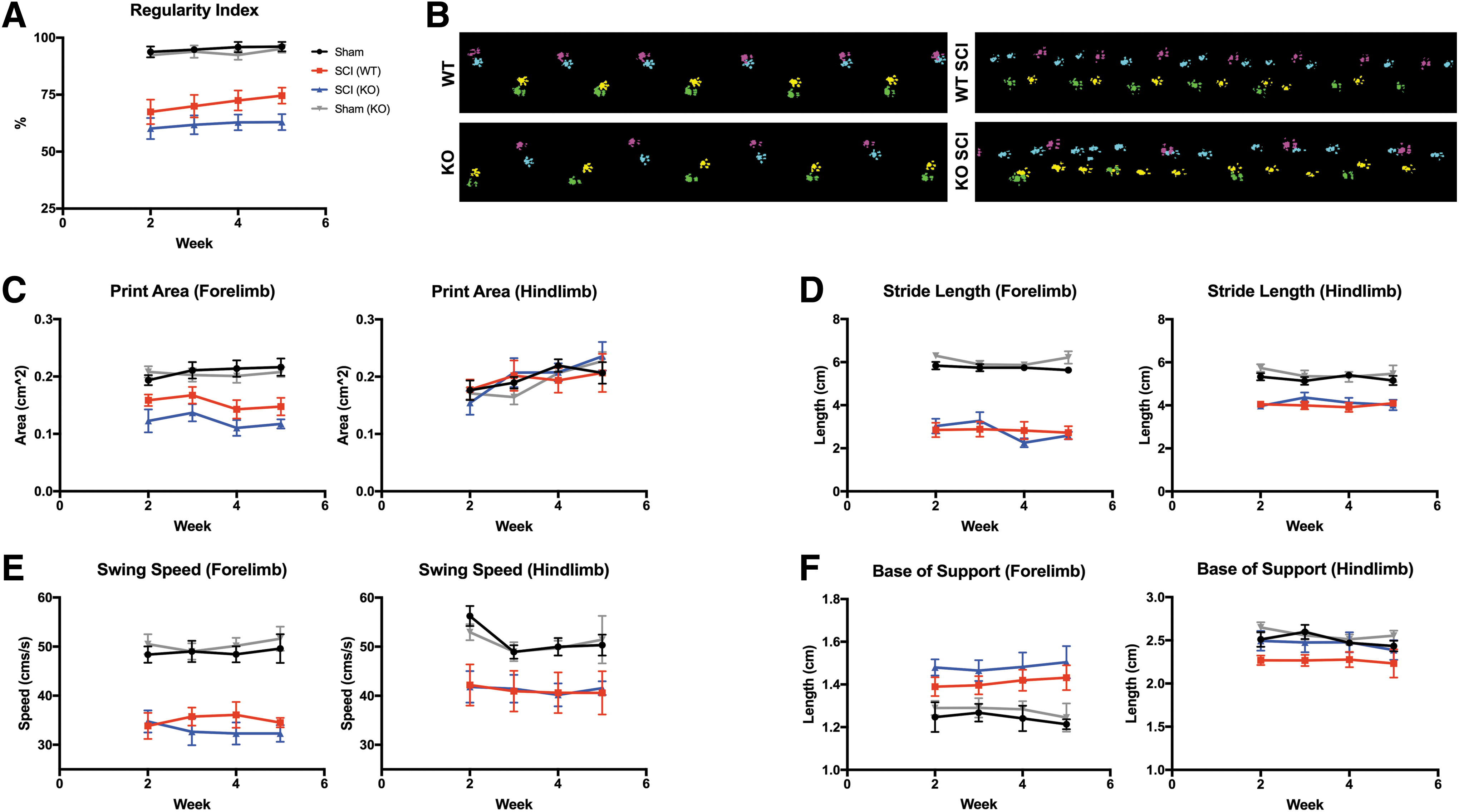
Loss of endogenous interleukin (IL)-10 did not significantly affect multivariate Catwalk gait parameters following cervical spinal cord injury (SCI). CatWalk analysis of mice (n = 9-10) at 2-5 weeks following SCI. Regularity index
IL-10 deficiency exacerbates long-term gray matter injury following cervical SCI
Histomorphometric analysis with LFB- and H&E-stained sections was used to quantify the lesion and gray and white matter tissue changes at 5 weeks post-SCI (Fig. 5A, 5B). IL-10 deficiency was associated with increased lesional tissue (two-way ANOVA, Tukey's post hoc test) at 250 μm rostral (p = 0.02) and caudal (p = 0.03) to the lesion, as well as at the lesion epicenter (p = 0.0002), compared with WT mice. IL-10 KO mice also had more gray matter loss than their WT counterparts at 750 μm (p = 0.03) rostral and 250 μm (p = 0.0001) caudal to the injury epicenter. These results were further confirmed with VHRUS imaging lesion quantification (Fig. 5C, 5D), where lesion volume was 0.31 ± 0.03 mm3 and 0.40 ± 0.02 mm3 for WT and KO mice, respectively (unpaired t-test, p = 0.04). Despite greater gray matter damage in IL-10 KO animals, which corresponds to the more vascularized region of the spinal cord, there was no significant difference (unpaired t-test, p = 0.37) in functional vascularity at 5 weeks post-SCI as detected by Power Doppler imaging (Fig. 6A, 6B). The raw area of Power Doppler signal values (without normalization to genotype controls), at 5 weeks post-SCI, are included Figure 8C.

Loss of endogenous interleukin (IL)-10 exacerbates gray matter injury and lesion volume following cervical spinal cord injury (SCI). At 5 weeks post-SCI, cross-sectional (30 μm) serial sections (n = 6 per group) were stained with Luxol fast blue and hematoxylin and eosin and systematically sampled at every 250 μm over a distance of 2500 μm, and unbiased measurements were made with a Cavalieri probe by using Stereo Investigator. Percent area of lesional tissue, gray matter, and white matter are shown
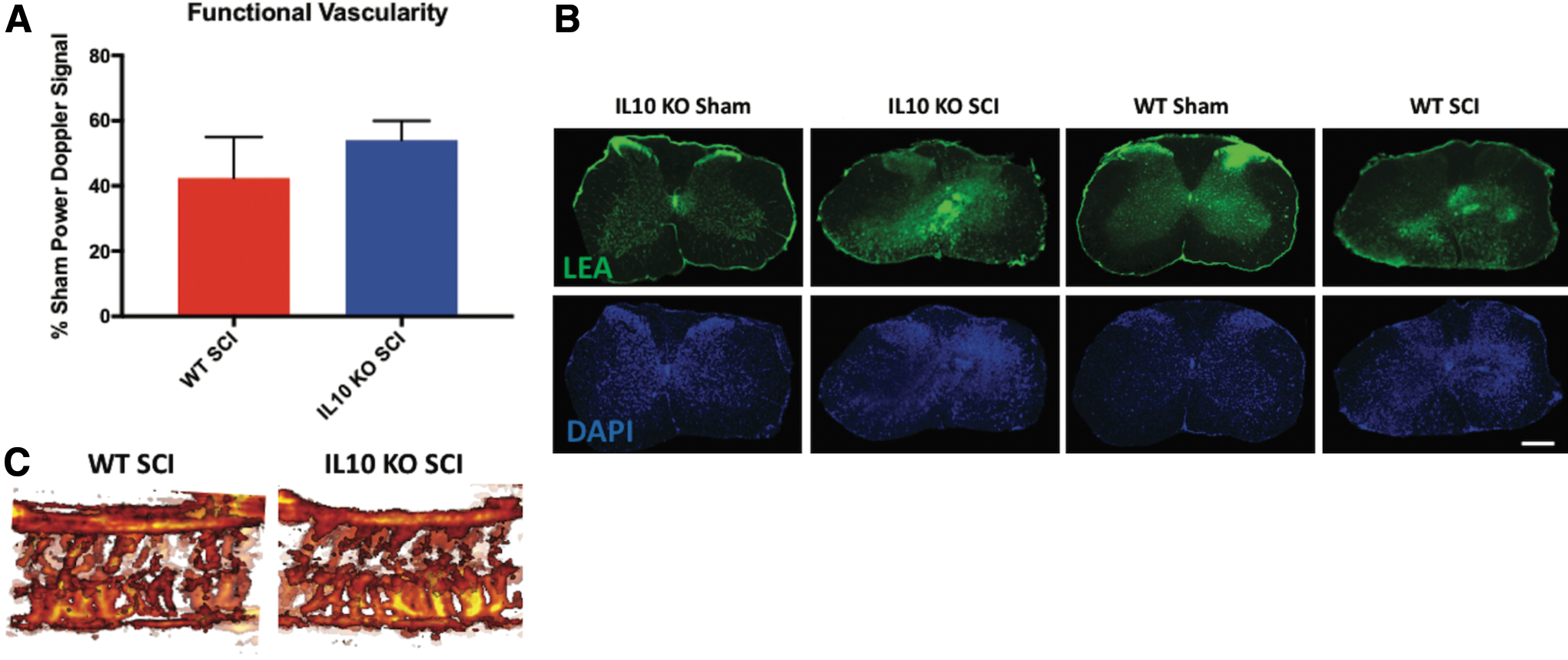
Interleukin (IL)-10 deficiency associated vascular damage dissipates by 5 weeks following cervical spinal cord injury (SCI). Power Doppler imaging was applied to quantify functional vascularity (blood flow) within the injured spinal cord normalized to sham controls. There was no difference (unpaired t-test) between wild type (WT) and IL-10 knockout (KO) functional vascularity
Altered systemic inflammatory cytokine response in SCI may account for limited long-standing effects
Serum cytokine levels were assessed by the Eve Technologies 31-Plex (MD31) Luminex assay at 24 h following cervical SCI (n = 5 for each sham group and n = 8 for each SCI group). The SCI groups were normalized to their corresponding genotype controls (sham surgery) and relative differences are shown on a heat map, with hierarchical clustering by average Euclidean distance (Fig. 7B). In the absence of endogenous IL-10, the relative levels of LIX (CXCL5) were highly elevated compared with the WT, while IL-12 (p70), IL-1β, and TNF-α were significantly reduced (multiple t-test with Holm-Sidak correction for multiple comparisons). The relative rise in IL-6 levels also was significantly lower in IL-10 KO mice compared with the WT controls (adjusted p = 0.037). The raw mean pixel density values (without normalization to genotype controls) are included in Figure 8A.

There is an altered acute systemic inflammatory cytokine/chemokine response following cervical spinal cord injury (SCI) in interleukin (IL)-10 deficient mice. Serum levels of IL-12 (p70), LIX (CXCL5), IL-1B, tumor necrosis factor (TNF)-α, and IL-6 were significantly different among IL-10 deficient and wild type (WT) mice at 24 h following cervical SCI
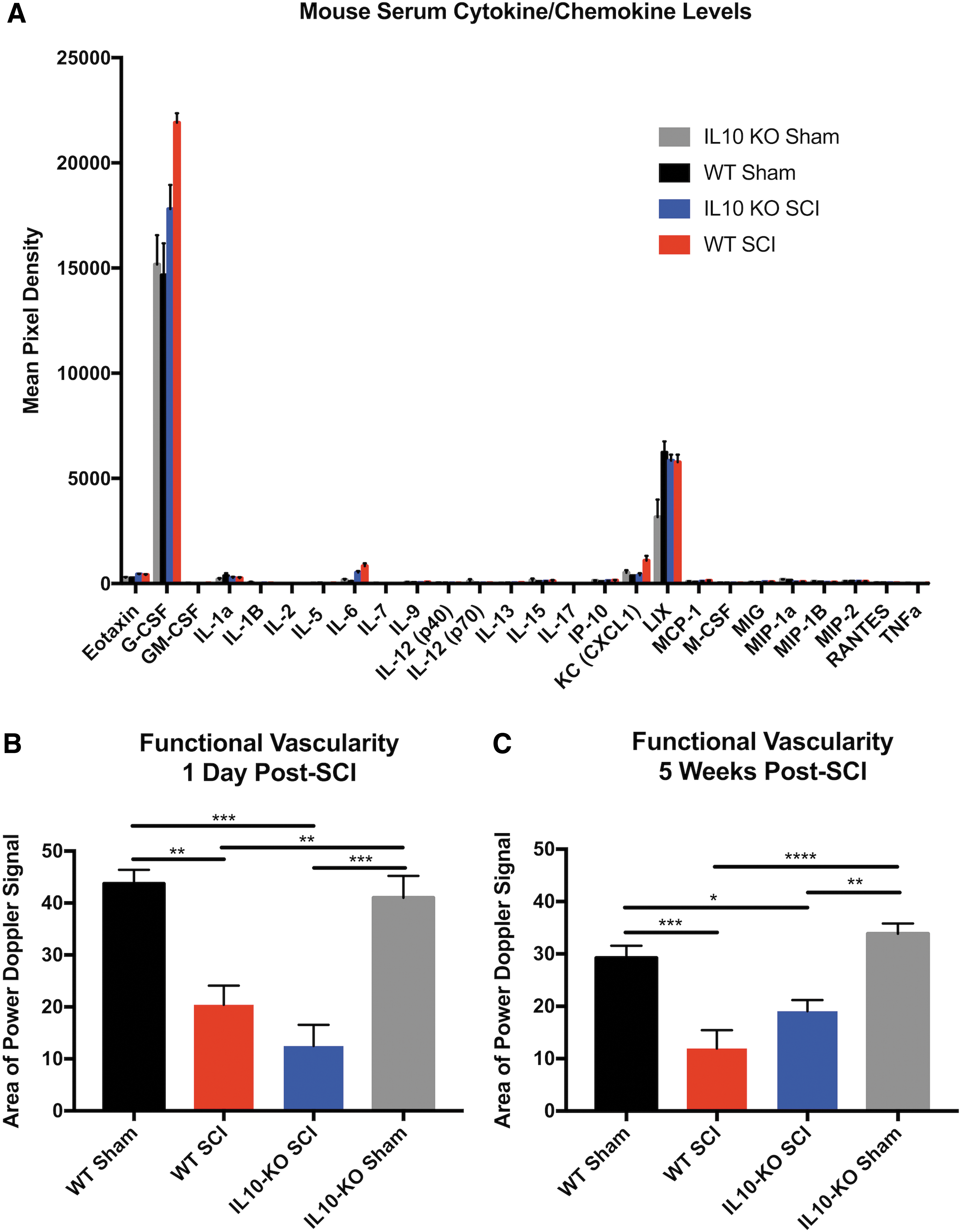
There are no significant differences in baseline cytokine levels and functional vascularity between genotypes. Mouse serum cytokine/chemokine levels 24 h following cervical spinal cord injury (SCI) or sham surgery
Discussion
This study demonstrates the successful application of a genetic deletion in a novel mouse cervical clip-compression SCI model. It is also the first characterization of cervical vascular pathology in the absence of IL-10 following SCI, further highlighting the complex interplay between neurovascular and immune function. In addition to significantly increased lesion volume, IL-10 KO mice exhibited trends towards greater intraparenchymal hemorrhage (p = 0.056) and reduced functional vascularity (p = 0.068) at 1 day post-SCI. The acute damage was further associated with less long-term tissue sparing and lower BMS scores. Interestingly, despite these effects, there was no significant difference in CatWalk gait parameters. This non-significant result may stem from the inherent variability in motivation, body weight and speed between WT and IL-10 KO mice, which has been reported to complicate direct comparisons using the CatWalk system. 31 The application of multiple experimental groups and time-points further reduced CatWalk statistical power and thereby sensitivity. 32 The differences in WT and IL-10 KO functional vascularity also were absent at 5 weeks following injury, emphasizing IL-10's dominant role in acute pathology.
Consistent with our results, loss of endogenous IL-10 has been previously reported to increase gray matter damage post-SCI, 22 as well as other secondary injury outcome measures, including edema and apoptosis. 23 These acute effects have been specifically attributed to increased levels of circulating inflammatory cells, 22 amplified pro-inflammatory cytokine signalling 23 and greater damage to vascular structures. 33 Yet, while Takami and colleagues found exogenous IL-10 administration after thoracic SCI was able to preserve gray matter integrity, this did not translate into significant effects in long-term functional recovery. 24 The modest functional recovery, shown in this study, likely stems from greater susceptibility to gray matter injury in the highly vascularized cervical spinal cord. 27 Further expanding on the vascular targets, IL-10 deficiency has been directly associated with loss of smooth muscle cells, greater collagen deposition, higher levels of matrix metalloproteinase-9 (MMP-9) and changes in the expression of adhesion molecules ICAM-1 and VCAM-1. 34 For this reason, IL-10 has been widely explored as a neuroprotective strategy for SCI. 35
However, as demonstrated by limited translational success, exogenous therapeutic application of IL-10 has been challenging. 19 In addition to a multitude of cell sources, IL-10 involves complex feed-forward and feedback pathways with temporal and spatial regulation. Beyond the intricate regulatory network, there is also the reality of short-lived pharmacological effectiveness, where the mean terminal phase half-life of recombinant human IL-10 is only about 2.7-4.5 h. 34 Therefore, it is increasingly clear that strategies aimed at modulating endogenous IL-10, such as the administration of mesenchymal stem/stromal cells (MSCs), 26 may be superior to exogenous supplementation and require greater mechanistic insight. Relatedly, MSCs that have been genetically engineered to overexpress IL-10 have also shown efficacy in various disease models. 37,38
The systemic cytokine profile in IL-10 KO animals compared with WT controls highlighted significant changes in the expression of IL-12(p70), LIX (CXCL5), IL-1β, TNF-α, and IL-6 relative to sham surgeries. Only LIX (CXCL5) expression, which stimulates the chemotaxis of neutrophils in acute inflammation, 39 was elevated with IL-10 deficiency. The others, many of which are directly regulated by IL-10, 40 were present in lower concentrations and are indicative of compensatory changes within the knockout animals. Importantly, this systemic profile contrasts with other studies involving depletion of endogenous IL-10 following SCI, where there is an upregulation in the production of inflammatory cytokines within the injured spinal cord. 23 Thus, it is possible that reduced systemic IL-1β and TNF-α levels following SCI in this study are associated with an upregulation of these two cytokines in the spinal cord, resulting in a more inflammatory environment that is less permissive for functional recovery. Future studies should explore cytokine levels in the spinal cord, exploring vasculature as the boundary between systemic and spinal cord inflammation.
Taken together, by use of a constitutive knockout this work provides a snapshot of the relationship between vascular injury and IL-10, which, to some extent, validates our previous work. 26 However, susceptible to compensatory cytokine responses stemming from overlapping function, 41 the overall changes are modest and a greater understanding of cell-specific responses is warranted. As a result, future work should assess IL-10 function through conditional knock-outs.
Conclusion
We show that IL-10 deficiency exacerbates secondary vascular pathogenesis through increased lesion volume and reduced functional vascularity. These acute effects further translated into a modest decrease in BMS score and tissue sparing, without any changes in overall animal gait. Despite trivial changes, likely due to altered cytokine response, these data support the relationship between vascular and immune function in SCI pathophysiology.
Footnotes
Acknowledgments
The authors would like to acknowledge Antigona Uldreaj for providing surgical training on the cervical mouse model and Sadiya Yousef for assistance with post-operative animal care. This work was supported by funds from the Canadian Institutes of Health Research Training Program in Regenerative Medicine (A.B.), the Ontario Graduate Scholarship (A.B.) and the James F. Crothers Family Fellowship in Peripheral Nerve Damage (A.B). M.F. was supported by the Halbert Chair in Neural Repair and Regeneration and Dezwirek Family Foundation.
Author Disclosure Statement
No competing financial interests exist.