
Abstract
The alpha 7 nicotinic acetylcholine receptor, α7 nAChR, plays a central role in regulating inflammatory responses. Previous studies showed that pharmacological inhibitors of α7nAChR have a pro-inflammatory effect, increasing the circulating levels of cytokines such as tumor necrosis factor alpha (TNFα). This study focused on how genetic polymorphisms of the partially duplicated α7nAChR gene (CHRFAM7A), which is highly expressed in peripheral blood cells, contribute to functional outcome after spinal cord injury (SCI). In a cohort of 27 SCI patients and 25 emergency room consented controls (% F/M: 15/85, 24/76; mean ± SE age: 35 ± 1.38 and 35 ± 2.0 respectively), a panel of circulating cytokines, noradrenergic metabolite (normetanephrine [NMN]) levels, and clinical data were available within the first 7 days post-injury (DPI) up to 90 DPI, and were investigated in the acute/subacute (DPI 1–21) and intermediate (DPI 22–90) temporal periods. Cytokine and NMN plasma levels on different DPI were analyzed as a function of CHRFAM7A genotype. TNFα levels, as a representative of some elevated inflammatory mediators, were nearly threefold higher in individuals carrying the del-2bp variant of the CHRFAM7A gene compared with that in the no-deletion genotype (p = 0.001 analysis of variance [ANOVA]) 3 weeks DPI, and twofold higher than genotype-matched acute/subacute non-SCI injury controls within 7 days DPI. In contrast, NMN levels were initially unchanged, although after 3 weeks, NMN levels were significantly decreased in SCI individuals carrying the del-2bp variant compared with non-carriers (p = 0.011 ANOVA). Numerical pain scores over this same period post-injury were significantly elevated in SCI patients carrying the del-2bp variant relative to non-carriers (p = 0.001 ANOVA). Taken together, these data reveal that pro-inflammatory responses associated with CHRFAM7A gene variation may also be associated with differences in pain experience in patients following SCI, at least during the intermediate phase.
Introduction
Spinal cord injury (SCI) exacts a tremendous financial, physical, and psychological toll on those affected. In the United States, more than $2 billion per year are directed to the care and support of those who sustain an SCI. 1 There is a great clinical need for new approaches to assessing/diagnosing and treating SCI. Currently, most available treatments for SCI aim only to minimize further damage caused by the injury by means of resuscitation, restoration of homeostasis, spinal stabilization, and spinal canal decompression.
The neurological deficit resulting from acute SCI is not simply a consequence of the initial tissue impact, but rather a combination of the resultant injury and the secondary inflammatory response, which in turn leads to further neural destruction in the first several weeks. In particular, cell signaling mechanisms that increase the levels of pro-inflammatory cytokines can have a deleterious effect on both cellular and functional recovery. 2 –4 In contrast, adoptive transfer of conditioned lymphocytes was shown to promote axonal and functional recovery after SCI. 5 Therefore, a better understanding of cellular and molecular mechanisms of the inflammatory response to injury is needed for advancing and personalizing therapy options.
The alpha 7 nicotinic acetylcholine receptor (α7 nAChR) plays a central role in regulating the body's inflammatory response. It is known that the parasympathetic nervous system, via the vagus nerve, can inhibit macrophage-dependent release of necrosis factor alpha (TNFα) and other cytokines, such as interleukin 1 beta (IL-1β) through cholinergic stimulation of the α7 nAChR. 6 This is supported by findings that genetic-based deficiency of the α7 nAChR promotes a pro-inflammatory phenotype in animal models. For example, in α7 nAChR-deficient mice, electrical stimulation of the vagus nerve no longer inhibits TNFα synthesis. 7
In humans, the α7 nAChR is encoded by the CHRNA7 gene, which is located on chromosome position 15q13.3. Genetic analysis of CHRNA7 has supported a role for this gene in neuropsychiatric disorders and neurodegenerative disorders. Reduced CHRNA7 gene function has been associated with schizophrenia, autism, epilepsy, and neurodegenerative diseases. 8 –12 However, the interpretation of these findings has been complicated by the discovery of a recently evolved, human-specific gene that resulted from a partial duplication of the CHRNA7 gene which is located ∼1.6 Mb centromeric of CHRNA7. 12 This gene, called CHRFAM7A, is a fusion of CHRNA7 and a partial duplication of another gene, called FAM7A, inserted upstream of the CHRNA7 duplication. 13 The CHRFAM7 gene maps to chromosomal location 15q14 and is transcribed in the opposite orientation of CHRNA7 (Fig. 1).
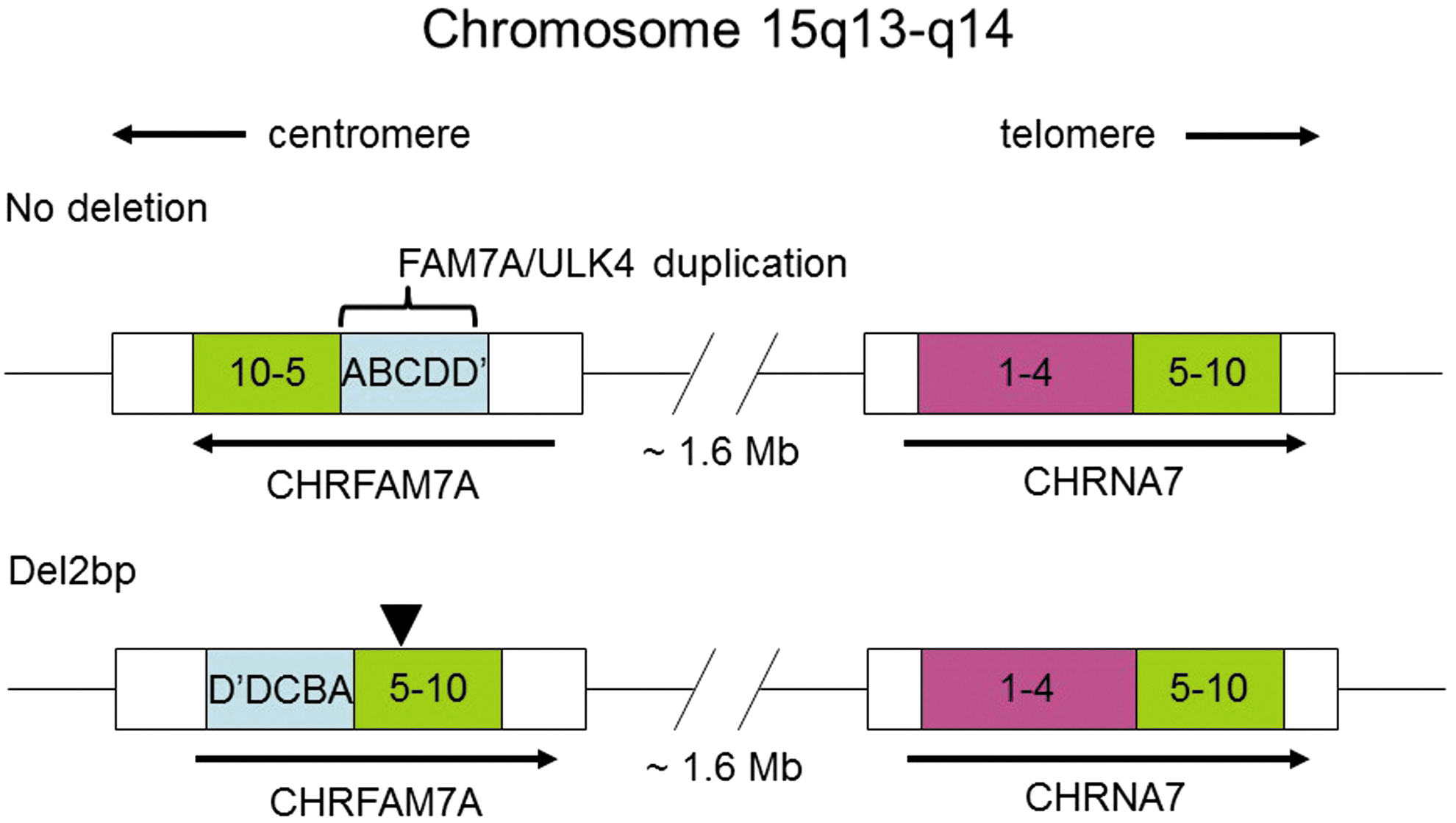
Genomic map of the CHRNA7 gene and its partial duplication CHRFAM7A. Orientation of the two genes on chromosome 15 are indicated with arrows based on the direction of transcription. Coding exons from CHRNA7 are labeled 1–10. Exons D′DCBA represent the duplication, FAM7A, from the ULK4 gene on chromosome 3. The position of the del-2bp variant is shown by an arrowhead.
Genetic polymorphisms of the CHRFAM7A gene are known. In particular, a functional polymorphism represented by a 2 base pair deletion (del-2bp) that occurs in exon 6 and is present at high frequency in different human populations (42% in individuals of European origin, and 14% of those of African origin). 10 The CHRFAM7A del-2bp gene variant encodes a protein called “dupΔα7.” 14 When dupΔα7 is co-expressed in cells with the α7 nAChR, α7 nAChR function is suppressed, in particular α7-mediated anti-inflammatory responses. 14 Therefore, within a cellular context, it is predicted that carriers of the del-2bp polymorphism would have a “pro-inflammatory” phenotype.
The goal of the present study was to determine if the del-2bp polymorphism of the CHRFAM7A gene is associated with increased TNFα production in individuals after SCI, compared with non-carriers. For the purposes of this study, we divided the stages after SCI into (1) an acute/subacute phase (days post injury [DPI] 1–21), a time when secondary damage is thought to occur 15 and (2) an intermediate phase (DPI 22–90). In addition, because catecholamine regulation and dysregulation are important in inflammation, we incorporated norepinephrine (NE) metabolite measures into the genotype stratified analysis. 16,17 Results from animal models of pain support roles for catecholamines and their interaction with pro-inflammatory molecules. 18,19 Therefore, we also determined if there was an association of the CHRFAM7A del-2bp polymorphism with development of pain in the same cohort of SCI subjects.
With no current effective therapies, there is a critical need to better understand the pathophysiology of SCI in the hope of facilitating more targeted treatments. The acute/subacute phase represents an earlier stage after SCI, which could be the target for neuroprotective therapies. Therefore, a study within this time frame may provide useful information toward this goal.
Methods
Participants
The study was approved by the Institutional Review Boards of the University of Pittsburgh and Inova Fairfax Hospital. A cohort of 27 acute traumatic SCI subjects with ratings of American Spinal Injury Association (ASIA) Impairment Scale (AIS) A/B was recruited at the University of Pittsburgh Medical Center (UPMC) from November 2009 to July 2012. The extent of initial SCI was classified using the AIS. Individuals with grades A and B (including 10 cervical and 17 thoracic spinal cord injuries) were eligible to participate in this study.
The inclusion criteria was a magnetic resonance imaging (MRI) confirmed diagnosis of acute traumatic spinal cord injury. Only individuals with AIS A or B were included in this study.
The exclusion criteria were pre-existing immune system related diseases and medications (including anti-inflammation or immune suppression treatments) as well as any previous SCI or other neurological diseases that affect motor and sensory function, or non-traumatic etiology. All medications taken during enrolment were recorded.
Twenty-five injury controls were recruited (orthopedic injuries ranging from contusions to single bone fractures) from Inova Emergency Department in Fairfax Hospital, from January 2014 to June 2015.
Control and SCI subjects were all ≥18 years of age. Demographic characteristics of the subjects are described in the Table 1.
Demographic Characteristics of the Two Subject Groups: Injury Control (Control) and Spinal Cord Injury (SCI)
A prospectively collected cohort of 27 patients with acute traumatic SCI recruited at the University of Pittsburgh Medical Center from November 2009 to July 2012. Recruitment of 25 trauma controls (orthopedic injuries ranging from contusions to single bone fractures) was from Inova Fairfax Hospital Emergency Department, January 2014 to June 2015.
Blood sample collections, biochemical analyses, and pain score rating
For the SCI patients, an initial blood sample from each patient was obtained within the 1st week post-SCI (mean 4.36 ± 1.87 days). This was used for a comparison to the trauma control samples taken in the acute phase post-injury (mean 1.87 ± 0.97 days). Subsequent blood samples were drawn one to three times per week post-SCI. The resulting plasma samples were aliquoted and stored at −80°C for later analysis. Aliquots (25 μL) in all serial blood samples collected were analyzed on a Luminex™ LX100 apparatus (MiraiBio, Austin, TX) using a human-specific 25-analyte cytokine/chemokine assay kit that included: EOTAXIN, granulocyte-macrophage colony-stimulating factor (GM-CSF), interferon (IFN)-α2, IFN-γ, IL-1Ra, IL-1β, IL-2, SIL-2RA, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12p40, IL-12p70, IL-13, IL-15, IL-17A, IP-10 (CXCL10), monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein (MIP)-1α, MIP-1β, TNF-α, and MIG (CXCL9) (Millipore, Billerica, MA). The median fluorescent value of each analyte was reported and compared with the fluorescent value of a standard curve with a range of 3.2 pg/mL to10,000 pg/mL for all analytes.
Norepinephrine metabolites (normetanephrine [NMN]) were determined using an enzyme-linked immunosorbent assay (ELISA) (Eagle Biosciences Inc, Nashua, NH).
Patient self-reported pain scores were captured using a numerical rating scale (NRS) from 0 to 10 (0 being no pain and 10 being the most severe pain). Pain scores were collected at least once a week by a researcher during a brief interview with the SCI patient. 20
Genotyping
CHRFAM7A 2bp deletion polymorphism was assessed as follows: the 2bp polymorphism (rs67158670) was genotyped using a custom Taqman allele discrimination assay and an ABI7000 Sequence Detection System (SDS) and SDS software version 1.2.3 (Thermo Fisher Scientific, Waltham, MA). The forward polymerase chain reaction (PCR) primer was 5′-CGCTGGTTTCCCTTTGATGTG-3′; the reverse PCR primer was 5′-GGACCAGCCTCCGTAAGAC-3′; the reporter probe for the non-deletion was 5′-VIC-CTGCAAACTGAAGTTT-3′; and the reporter probe for the deletion allele was 5′-FAM-CACTGCAAACAAGTTT-3′. Cycling conditions were 95°C for 10 min followed by 50 cycles of 95°C for 15 sec and 58°C for 1.5 min. Genotypes were called by two different individuals blinded to phenotypes, with discrepancies decided by review of raw data or re-genotyping.
Statistical analysis
Two temporal groups of data were analyzed: DPI 1–21 and DPI 22–90, the former representing the acute/subacute phase and the latter representing intermediate conditions after SCI. For genetic association analyses, subjects were stratified based on CHRFAM7A del-2bp carrier status.
All the inflammatory mediators measured were first examined using a non-parametric (Mann–Whitney) test between the two genotypes in the acute/subacute and intermediate phases. Then, for further multiple assessments of the associations between the inflammatory mediators/pain and the effects of the genotype and DPI, the analysis of variance (ANOVA) with repeated measurements followed by Bonferroni post–hoc test were employed. This analysis was also used when performing multiple comparisons of TNFα levels in the orthopedic injury control group versus that in the SCI group between different genotypes in the acute phase post-injury. When it was necessary, the Pearson correlation analysis was also applied. All analyses were performed using SPSS and SYSTAT statistical programs (Evanston, IL). Statistics significance was assumed at p < 0.05.
Results
The demographic characteristics of the SCI and the control subject groups were similar in age and sex. Although the majority ethnic group represented in this study was white, the number of individuals from minority ethnic populations differed in the SCI and control groups (Table 1).
Multiple inflammatory mediators are increased during the acute/subacute phase (DPI 1–21) in SCI subjects carrying the CHRFAM7A del-2bp allele. The data in Table 2 summarize statistically significant differences of the inflammatory mediators based on CHRFAM7A 2bp carrier status in the SCI patients using a non-parametric ranking test (Mann–Whitney test) during the acute/subacute phase (DPI 1–21). As shown in Table 2, the following inflammatory mediators were statistically significantly increased between the genotype groups: TNFα, GM-CSF, IFNγ, IL-1β, IL-7, IL-8, IL-12p40, IL-17A, MCP-1, MIP-1α, and MIP-1β in the del-2bp carriers during the acute/subacute phase post-injury. No such significant differences in the level of inflammatory mediators were detected in the two genotype groups during the intermediate phase (DPI 22–90) (data not shown). Limited by resources in the current study, we focused on TNFα as a representative inflammatory mediator. During the acute/subacute phase, in particular, in the third week DPI, del-2bp carriers had ∼2.8-fold higher levels of TNFα than non-carriers (8.3 ± 1.9 pg/mL for del-2bp carriers versus 3.0 ± 0.8 pg/mL for non-carriers, p = 0.048 Student's t test) (Fig. 2). To determine if the difference in TNFα levels between the genotype groups among SCI subjects was caused by the differences in DPI, we analyzed TNFα levels testing interactions between genotypes and DPI. Based on ANOVA, differences in TNFα levels between the two genotypes over DPI 1–21 were mainly associated with the different genotypes rather than being associated with differences in DPI (Fig. 2).
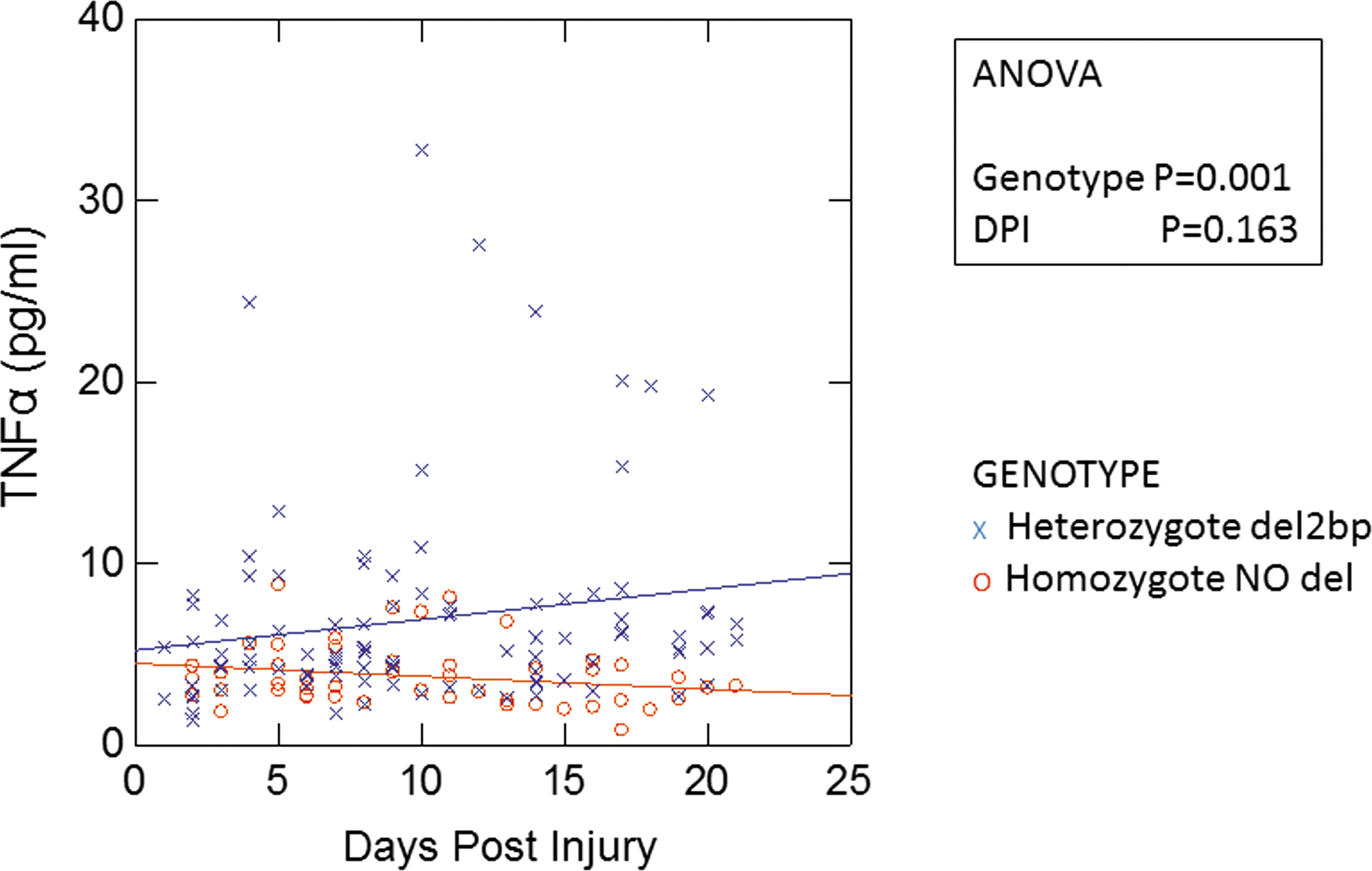
Plots of plasma tumor necrosis factor (TNF)α levels (pg/mL) in spinal cord injury (SCI) American Spinal Injury Association AIS A/B subjects during 1–21 days post-injury (DPI). The interaction between the DPI and CHRFAM7A del-2bp genotype over the effects on TNFα levels was examined by analysis of variance (ANOVA) followed by Bonferroni post-hoc test. Statistically significant differences are indicated in the figure. Fitting regression lines by genotype were added to visualize the main genotypic effect on the TNFα levels.
Genotype Effects on Inflammatory Cytokines in the Spinal Cord Injury (SCI) Patients during the Acute/Subacute Phase (1–21 Days Post-Injury [DPI])
GM-CSF, Granulocyte-macrophage colony-stimulating factor; IFN, interferon; IL, interleukin; MCP-1 (CCL2), monocyte chemoattractant protein 1; MIP-1α (CCL3), macrophage inflammatory protein 1 alpha; MIP-1β (CCL4), macrophage inflammatory protein 1 beta; TNF, tumor necrosis factor; MWRT Mann–Whitney ranking test.
Plasma TNFα levels are increased in SCI subjects compared with trauma injury controls during the initial time point (1–7 DPI)
To determine if the TNFα levels were altered by SCI, we compared the initial samples taken on DPI 1–7 from SCI patients with that from the acute injury control patients. There was a significant difference between the two injury groups for TNFα (increase of twofold SCI vs. control, ANOVA injury group effects p = 0.004) (Table 3). To determine if there was a difference in TNFα levels between the genotype groups soon after their injury, we also compared TNFα levels in the initial samples, stratified by CHRFAM7A del-2bp genotype. Based on ANOVA of the effects of injury group and genotype for this initial time point, although TNFα levels were increased in the SCI group, there was no statistical difference in TNFα levels between the genotype groups for either SCI or control subjects (Table 3).
Initial Plasma TNFα levels (Mean ± SE pg/mL) for Two CHRFAM7A Genotypes (no del or del-2bp) in the Two Subject Groups: Orthopedic Injury Control (Control) and Spinal Cord Injury (SCI) 1–7 Days Post-Injury
p value generated by analysis of variance (ANOVA).
TNF, tumor necrosis factor.
Plasma levels of the NE metabolite, NMN, are decreased during the intermediate phase of injury in subjects carrying the CHRFAM7A del-2bp allele
In peripheral monocytes and macrophages, cytokine genes are targets for NE action. In a beta2-adrenergic receptors (β2AR)-dependent mechanism, increased levels of catecholamines are brought about by decreasing their metabolism, which occurs by inhibiting the enzyme catechol-O-methyltransferase (COMT), thereby increasing circulating TNFα. 18 This increase in TNFα is expected to tip the inflammatory balance to a pro-inflammatory state.
We next wanted to determine if CHRFAM7A genotypes that were associated with increased TNFα levels following SCI were correlated with enhanced NE levels. To do this, we used an indirect approach, determining plasma levels of the NE metabolite, NMN, in the same group of SCI subjects. Although there were no significant differences between the genotypes in the acute/subacute phase, in the intermediate phase following injury (DPI 22–90), plasma NMN levels decreased more rapidly in del-2bp carriers than in non-carriers (ANOVA p = 0.011).The decrease in NMN levels in del-2bp heterozygotes was not dependent on DPI (ANOVA p = 0.26) (Fig. 3). Taken together, these data support the idea that acute/subacute TNFα responses and intermediate NE responses are both associated with the CHRFAM7A del2bp genotype.

Plots of normetanephrine (NMN) plasma levels (ng/mL) in spinal cord injury (SCI) American Spinal Injury Association AIS A/B subjects during 22–90 days post-injury (DPI). The interaction between the DPI and CHRFAM7A genotype over the effects on tumor necrosis factor (TNF)α levels was examined by analysis of variance (ANOVA) followed by Bonferroni post-hoc test. Statistically significant differences are indicated in the figure. Fitting regression lines by genotype were added to visualize the main genotypic effect on the NMN levels.
Numerical pain scores from intermediate SCI subjects are increased in carriers of the del2bp CHRFAM7A gene polymorphism
The α7 nAChR is expressed in various cell types in the central nervous system (CNS) and the periphery, contributing to acute and chronic pain in humans, and has been shown to be a therapeutic target in animal models of inflammatory pain. 21 –24 Given the fact that α7 nAChR activation also suppresses TNFα production by central microglia 25 and monocyte/macrophages in the periphery, 26,27 we asked if SCI subjects who carried the del-2bp had a higher level of neuropathic pain than del-2bp non-carriers. Numerical pain scores from SCI subjects were analyzed based on carrier status of the del-2bp CHRFAM7A polymorphism. The data in Figure 4 show that pain scores were higher in del-2bp carriers than in non-carriers in the intermediate phase (22–90 DPI) (ANOVA p = 0.001). This increase in pain score among del-2bp carriers compared with the non-carriers was not dependent on the DPI (ANOVA p = 0.13) (Fig. 4). Moreover, the NMN levels showed a trend correlation with the pain score: Pearson correlation p = 0.069 independent of CHRFAM7A del-2bp genotype. However, the correlation between NMN and genotype was statistically significant: Pearson correlation p = 0.017. TNFα levels also showed a significant correlation with the pain score: Pearson correlation p = 0.025 in the SCI patients, but no correlation with the genotype. Finally, the increase in pain score among del-2bp carriers was evident during the intermediate phase. No significant genotype-based differences in pain scores were observed in the SCI subjects within 21 DPI (data not shown).

Plot of numerical pain score rating (from 0 to 10) in spinal cord injury (SCI) American Spinal Injury Association AIS A/B subjects during 22–90 days post-injury (DPI). The interaction between the DPI and CHRFAM7A genotype over the effects on tumor necrosis factor (TNF)α levels was examined by analysis of variance (ANOVA) followed by Bonferroni post-hoc test. Statistically significant differences are indicated in the figure. Fitting regression lines by genotype were added to visualize the main genotypic effect on the pain score.
Discussion
This study showed that individuals with AIS A–B, up to 21 DPI had circulating TNFα levels that were significantly higher in individuals carrying the del-2bp allele of the CHRFAM7A gene than in non-carriers. Temporal increases in TNFα levels above initial values have been seen in the blood of SCI patients up to 56 days following injury. 28 Further, longitudinal studies have been conducted that determined serum chemokines levels up to 3 months after SCI. 29 However, this is the first report involving subjects with SCI in whom levels of a biomarker associated with inflammation were differentiated by CHRFAM7A genotype. In addition, our findings suggest a role for the α7 nAChR in the regulation of inflammation in the context of injury. Although it is appreciated that post-injury inflammation is a dynamic response associated with time-dependent changes in TNFα, when DPI was used as a covariant in ANOVA in this cohort of subjects, time post-injury did not account for differences between the genotypes in plasma TNFα levels during the acute/subacute phase after injury.
The α7nAChR is known to regulate the inflammatory immune response within the central and peripheral nervous system. Pharmacological inhibitors of α7 nAChR are pro-inflammatory, 30 whereas activators of the α7 nAChR maintain robust anti-inflammatory properties. 7,31 The CHRFAM7A del-2bp gene variant encodes a truncated α7 subunit that reduces sensitivity of the receptor to its agonist and causes α7 receptors to desensitize more rapidly following exposure to physiological concentrations of agonists such as choline. 14 The reduction in α7 nAChR activity is predicted to produce a pro-inflammatory phenotype. Consistent with this notion, we found that TNFα levels were increased along with other inflammatory mediators, namely GM-CSF, IFNγ, IL-1b, IL-7, IL-8, IL-12p40, IL-17, MCP-1, MIP-1a, and MIP-1b in the CHRFAM7A del-2bp carriers 3 weeks post-SCI, a time when the secondary damage from inflammation has been suggested to occur after SCI.
One modulator of TNFα production is the hormone and neurotransmitter, NE. Because urine samples were not available and because NE is not stable in blood, we determined plasma levels of the NE metabolite, NMN, in the SCI cohort and found that in the intermediate phase following injury, namely 22–90 DPI, plasma NMN levels decreased more rapidly in del-2bp carriers than in non-carriers, supporting the idea that an anti-inflammatory component may be diminished in carriers of the del-2bp polymorphism. This association corresponded with increased pain scores observed in the SCI patients carrying the del-2bp variant relative to non-carriers during the same post-injury period, which was not dependent on DPI. These systemic changes may be reflections of cellular responses within the injured spinal cord, where infiltrating monocytes/macrophages and microglia promote a chronic inflammatory state. 32 Taken together, our findings are consistent with results from experimental pain models, where blockade of α7 nAChRs by specific antagonists, such as methyllycaconitine, systemically increases hyperalgesia. 23,33
The influence of systemic changes in immune function following SCI is important in the intermediate and chronic phases of SCI, and to the intraspinal inflammation seen following SCI. For example, elevated levels of serum cytokines, including TNFα, have been observed in chronic SCI patients independent of secondary medical complications such as bacterial infection or pressure ulcers. 34,35 This coincides with aberrant accumulation of NE and glucocorticoids, leading to enhanced activation of beta2-adrenergic receptors (β2 ARs) and glucocorticoid receptors. 32 The relationship of these biochemical changes to the causative mechanism of neuropathic pain is not well understood. Enhanced activation of β2 ARs after injury may be a compensatory mechanism. It is thought that α2AR activation suppresses neuropathic pain, but the effect is lost over time following nerve injury. 36 Evidence for an AR-dependent mechanism is supported by the effect of serotonin-NE reuptake inhibitors (SNRI) on pain relief. In a randomized, double-blind, placebo-controlled trial of 48 subjects, which included SCI patients with central pain, duloxetine (a mixed SNRI), showed a trend toward decreased mean pain scores after 8 weeks of treatment and a significant improvement in bodily pain domain measures taken from the patient-reported 36 Item Short Form (SF-36) Health Survey. 37 The results of the duloxetine clinical trial suggested that increasing NE α2AR-dependent signaling may be beneficial to recovery over time, allowing for reduced pain scores. In the current study, we observed a stable temporal profile for the NE metabolite NMN in SCI patients for non-carriers of the del-2bp polymorphism, suggesting an association with reduced pain. This association occurred in the absence of other genotype-related changes in clinical complications, which included the rate of developing pressure ulcers (W.H., unpublished observation). In contrast, del-2bp carriers had reduced plasma NMN levels past DPI 21. One interpretation of this observation is that reduced NMN levels may indicate reduced catabolism of NE by COMT, thereby increasing NE levels. Although not understood in clinical populations, it is known from animal studies that β2AR or β3AR activation provides a mechanism whereby COMT increases TNFα and other pro-inflammatory cytokines. 18 Thus, in this context, increased plasma TNFα and NE levels are associated with increased algesia, which is opposite to the proposed effects of SNRIs on pain relief via α2AR activation, but may underlie the development of chronic pain in patients. 38 We noted that the TNFα levels remained elevated over time and correlated with pain score. However, there was no statistically significant difference in plasma TNFα levels between the genotype groups based on del-2bp carrier status during the intermediate phase. It is plausible that the CHRFAM7A gene modifies acute/subacute TNFα levels, and that this effect is part of a sequentially regulated phenomenon with NE that increased pain scores in SCI subjects after DPI 21, but only in del-2bp carriers.
Using our findings that decreased circulating NMN levels are associated with increased circulating TNFα levels and increased pain scores, we created a concept defining a role for the CHRFAM7A del-2bp polymorphism in the intermediate phase of SCI in placing α7 nAChR as a central regulator of anti-inflammatory responses (Fig. 5). Expression of functionally altered subunits, including those encoded by CHRFAM7A del-2bp, is expected to impact cholinergic anti-inflammatory responses. We propose that following SCI, the del-2bp variant α7 nAChR will tip the balance of cellular responses to a more inflammatory state.

Proposed concept for alpha 7 nicotinic acetylcholine receptor (α7 nAChR)-mediated regulation of tumor necrosis factor (TNF)α responses in the periphery: interaction with norepinephrine (NE)-mediated regulation of TNFα. Presence of the CHRFAM7A del-2bp variant is predicted to tip the balance toward a more inflammatory state following spinal cord injury (SCI), increasing peripheral and central TNFα levels (shown by the plus symbols). NE signaling through beta2-adrenergic receptors (β2AR) potentiates TNFα production, which can be further enhanced by reduced catecholamine-O-methyltransferase (COMT) enzyme activity. In our study, reductions in the level of NE metabolite, normetanephrine (NMN), suggested an alteration in NE levels after SCI, leading to increased TNFα levels. Conversely, in the absence of the CHRFAM7A del-2bp variant (NO del), activity at α7 nAChR is normal, producing a reduction TNFα levels (shown by the minus symbol).
Future investigations are envisioned using patient-derived monocytes and inducible pluripotent stem-cell-derived microglia on different CHRFAM7A genetic backgrounds to describe the cellular responses to α7 nAChR activators such as choline, which exert anti-inflammatory and antinociception activity. 21,22,24 Because of the contribution of β2AR and β3AR to this COMT-dependent mechanism, it likely originates from the periphery. 38,39 Therefore, there may be therapeutic targets on peripheral mononuclear leukocytes in addition to sites on nociceptors that could be used to treat SCI patients with chronic pain.
Strengths and limitations of the study
A number of limitations are noted. The strongest findings of this study are the relationship between CHRFAM7A del-2bp carrier status and plasma TNFα levels and between CHRFAM7A del-2bp carrier status and pain score. These novel findings were based on blood samples obtained 21 DPI (TNFα) and >21 DPI (Pain Score). The TNFα finding was limited by the fact that we could not link the finding causatively to SCI, as there was only one blood sample taken for the cytokine determination in the injury control group; namely, at the initial time point following recruitment. Future studies should incorporate a control group series with samples taken during the acute/subacute and intermediate phases following injury. In addition, although the SCI and control groups used in this study were demographically identical in age and sex, they differed in the proportion of individuals by ethnic group. Because participants were recruited from two geographically different study sites, there could have been recruitment bias. Another limitation of the study was the number of participants. SCI is a complex traumatic injury with a number of confounding factors. Although we attempted to limit those variables through the inclusion/exclusion criteria, recruitment of a larger study population is needed. 15 This is true for the interventional clinical trials as well as for observational studies in SCI. Therefore, our results should be interpreted with caution until a replication study is performed.
With these limitations noted, knowledge of CHRFAM7A genotype may guide future anti-inflammatory therapies, allowing stratification of SCI patients at increased risk of inflammation-associated complications, including development of neuropathic pain. Because inflammatory processes are complex, more than a single gene is predicted to modify risk of functional decline in SCI patients. Additional studies will be necessary in different clinical populations to determine if variation at CHRFAM7A is associated with SCI or may modify phenotypes that are associated with injury in general.
Footnotes
Acknowledgments
The authors thank Sandra Deslouches and Derek Barclay for excellent technical assistance. This project was funded in part by a grant from the Wings for Life Spinal Cord Research Foundation (N.K. and R.H.L.).
Author Disclosure Statement
No competing financial interests exist.