
Abstract
The present study was designed to evaluate the therapeutic effectiveness and mechanism of acute intermittent hypoxia on respiratory function at distinct injury stages following mid-cervical spinal contusion. In the first experiment, adult male rats received laminectomy or unilateral contusion at 3rd-4th cervical spinal cord at 9 weeks of age. The ventilatory behavior in response to mild acute intermittent hypercapnic-hypoxia (10 episodes of 5 min of hypoxia [10% O2, 4% CO2, 86% N2] with 5 min of normoxia intervals) was measured by whole–body plethysmography at the acute (∼3 days), subchronic (∼2 weeks), and chronic (∼8 weeks) injury stages. The minute ventilation of contused animals is significantly enhanced following acute intermittent hypercapnic-hypoxia due to an augmentation of the tidal volume at all time-points post-injury. However, acute intermittent hypercapnia-hypoxia–induced ventilatory long-term facilitation was only observed in uninjured animals at the acute stage. During the second experiment, the effect of acute intermittent hypercapnic-hypoxia on respiration was examined in contused animals after a blockade of serotonin receptors, or adenosine 2A receptors. The results demonstrated that acute intermittent hypercapnic-hypoxia–induced enhancement of minute ventilation was attenuated by a serotonin receptor antagonist (methysergide) but enhanced by an adenosine 2A receptor antagonist (KW6002) at the subchronic and chronic injury stages. These results suggested that acute intermittent hypercapnic-hypoxia can induce respiratory recovery from acute to chronic injury stages. The therapeutic effectiveness of intermittent hypercapnic-hypoxia is dampened by the inhibition of serotonin receptors, but a blockade of adenosine 2A receptors enhanced respiratory recovery induced by intermittent hypercapnic-hypoxia.
Introduction
Cervical spinal cord injury is usually associated with respiratory deficiency due to damage of the spinal respiratory motoneurons and/or interruption of bulbospinal respiratory pathways. 1 –7 Currently, there are limited effective therapeutic strategies to restore the respiratory function in patients with cervical spinal cord injury. 6 Several pre-clinical and animal studies have shown that mild acute intermittent isocapnic or hypercapnic-hypoxia (mAIH) can improve the breathing capacity through the induction of respiratory neuroplasticity. For example, two independent pre-clinical studies demonstrated that minute ventilation could be enhanced for 30–40 min after exposure to mAIH in cervical spinal cord injury patients. 8,9 In addition, animal studies showed that mAIH can enhance the phrenic nerve activity ipsilateral to unilateral high-cervical hemisection at 4–8 weeks post-injury in anesthetized rats. 10,11 Moreover, mAIH can evoke augmentation in an uninjured side, but not on the injured side, of diaphragmatic activity in unanesthetized C2 hemisected animals at the chronic injury stage. 12
Our recent report further demonstrated that mAIH can significantly improve the tidal volume at 4 weeks post-injury in a clinically relevant mid-cervical contusion rodent model. 13 These results suggested that mAIH is a potential therapeutic strategy to improve respiratory function following cervical spinal cord injury. However, it is critical to evaluate a suitable time window for induction of intermittent hypoxia therapy and examine whether mAIH can still have therapeutic effectiveness on the respiratory function at a longer chronic injury stage. Accordingly, the first aim of this study was to monitor respiratory behaviors of mid-cervical contused rats in response to mAIH at the acute (i.e., 2–3 days), subchronic (i.e., ∼2 weeks), and chronic injury stages (i.e., ∼8 weeks).
It is established that intermittent hypoxia-induced respiratory neuroplasticity is primarily mediated by activation of serotonin receptors in normal rats, which causes a series of intracellular signal transduction to enhance phrenic motoneuron excitability, then to increase phrenic motor outputs. 14 –17 Dougherty and colleagues further demonstrated that daily acute intermittent hypoxia-induced recovery of ventilation in C2 hemisected rats is mediated by the serotonin-dependent and serotonin-independent mechanisms at the acute vs. chronic injury stage, respectively. 18
Moreover, Navarrete-Opazo and colleagues showed that inhibition of adenosine 2A receptors has a differential effect on the ventilatory response induced by intermittent hypoxia in awake animals at different time-points post-injury. 19,20 Specifically, the adenosine 2A receptor antagonist attenuated the enhancement of contralateral diaphragm electromyogram activity following daily acute intermittent hypoxia 2 weeks after C2 hemisection. 20 Interestingly, the same dose of the adenosine 2A receptor antagonist rather enhanced daily acute intermittent hypoxia-induced increase in the tidal volume in C2 hemisected animals at nine weeks post-injury. 19 These results implied that intermittent hypoxia-induced respiratory neuroplasticity is switched from the adenosine-dependent mechanism at the acute injury stage to the serotonin-dependent mechanism at the chronic injury stage.
However, the aforementioned studies were exclusively conducted in high-cervical hemisected animals. Since mid-cervical contusion is the most common type of spinal cord injury, and both bulbospinal respiratory pathways and cervical phrenic motoneurons were damaged by such contused injury, it is important to investigate the modulatory role of serotonin receptors and adenosine 2A receptors on intermittent hypoxia-induced respiratory plasticity from the acute to chronic injury stage. Accordingly, the second aim of our study was to evaluate the effect of mAIH on respiratory function after the blockade of serotonin receptors or adenosine 2A receptors at distinct injury stages following mid-cervical spinal cord contusion.
Methods
Animals
All experimental procedures were approved by the Institutional Animal Care and Use Committee at the National Sun Yat-sen University and performed in accordance with the guidelines for experimental animals. 21 A total of 69 adult male Sprague-Dawley rats (7–8 weeks of age) were obtained from BioLasco Taiwan Co., Ltd and housed in an animal room with access to food and water ad libitum. These animals received C3 laminectomies (n = 8) or cervical spinal cord contusion surgery (n = 61). Seven animals died after the cervical contusion surgery and were excluded from the further experimental protocol.
Two series of experiments were conducted in the study. In the first experiment, eight uninjured animals (C3 laminectomy only) and 10 mid-cervical spinal cord contused animals were used. An additional 44 mid-cervical spinal cord contused animals were used for the second experiment. The number and body weight of the animals used in each experiment are presented in Table 1.
Number and Body Weight (g) of the Animal Used in the Present Study
p < 0.05 vs. Acute.
†p < 0.05 vs. Acute and Subchronic.
#p < 0.05 significant difference between uninjured and contused animals.
mAIH, mild acute intermittent hypoxia.
Cervical spinal cord contusion
Cervical spinal cord injury surgery was similar to our previous reports. 4,13,22 Animals were anesthetized with xylazine (10 mg/kg, subcutaneous [s.c.], Rompun®, Bayer) and ketamine (140 mg/kg, intraperitoneal [i.p.], Ketalar®, Pfizer) at 9 weeks of age. After an absence of toe-pinch withdrawal reflex, the C3 vertebrate bone was clamped bilaterally by a pair of transverse clamps (#51692, Stoelting Co.) and Cunningham spinal adaptors (#51695, Stoelting Co.) mounted on a stereotaxic instrument (David Kopf Instruments). The animals in the contused group were placed on the stage of the Multicenter Animal Spinal Cord Injury Study (MASCIS) Impactor System (Model II, W.M. Keck Center for Collaborative Neuroscience) and received a unilateral spinal cord contusion by releasing the impact rod (weight: 10 g; diameter: 2 mm) at the height of 6.25 mm above the left spinal cord surface. The animal would be mechanically-ventilated (Harvard Apparatus) with 50% O2 (balance N2) until recovery of spontaneous breathing if the animal ceased breathing after the contusion injury. The dorsal skin and muscle were then sutured using 4-0 nylon (UNIK) and 4-0 chromic sutures (UNIK), respectively. Yohimbine (1.2 mg/kg, s.c., Tocris) and lactated Ringers solution (5 mL, s.c., Nang Kuang Pharmaceutical Co., Ltd.) were administered to reverse the effects of xylazine and prevent dehydration, respectively. An analgesic (buprenorphine, 0.03 mg/kg, s.c.; Shinlin Sinseng Pharmaceutical Co., Ltd.) was also provided after the animal woke. Daily oral delivery of Nutri-Cal (1-3 mL, EVSCO pharmaceuticals) and lactated Ringers solution (5 mL, s.c.), were supplied until adequate volitional drinking and eating recovered.
Drug preparation
The serotonin receptor antagonist (methysergide maleate, #1064, Tocris) was dissolved in normal saline at 2 mg/mL and stored at −20 °C. The adenosine 2A receptor antagonist (KW6002, #5147, Tocris) was dissolved in dimethyl sulfoxide (DMSO) at 5 mg/mL, stored at −20 °C, and diluted to 0.25 mg/mL in phosphate-buffered saline (UniRegion Bio-Tech) on the day of experiment. Methysergide (4 mg/kg) or KW6002 (0.5 mg/kg) was injected intraperitoneally 5 min before mAIH or normoxia (i.e., time control) treatment after a 1-h baseline recording.
Whole–body plethysmography
Ventilatory behaviors (e.g., respiratory frequency, tidal volume, and minute ventilation) of unanesthetized animals were measured using the whole–body plethysmography system (Data Sciences International), as previously described. 13,22,23 In the first experiment, animals (uninjured, n = 8; contused, n = 10) were placed in Plexiglas chambers (#PLY4213, volume: 3.9 L) balanced with air (2.5 L/min) for a 1-h baseline condition, then received a 100-min normoxia (i.e., time control) at the acute (2 days), subchronic (14.5 ± 0.1 days), and chronic (56.6 ± 0.2 days) injury stages. The same animals were exposed to intermittent hypercapnic-hypoxia (10% O2, 4% CO2, balance N2 for 5 min with a 5-min normoxia interval; 2.5 L/min) at the acute (3 days), subchronic (15.4 ± 0.1 days), and chronic (58.8 ± 0.5 days) injury stages (Fig. 1). An additional 1-h normoxic breathing was continuously monitored after a 100-min normoxia, or mAIH treatment. Because hypoxia is usually associated with hypocapnia, which may reduce the capability of inducing respiratory neuroplasticity. 24 Harris and colleagues have demonstrated that sustained elevated CO2 is essential to induce ventilatory long-term facilitation in humans. 25 Therefore, we used hypercapnic-hypoxia (10% O2, 4% CO2) as our mAIH paradigm, which has been proven to be able to induce ventilatory plasticity of mid-cervical contused animals. 13
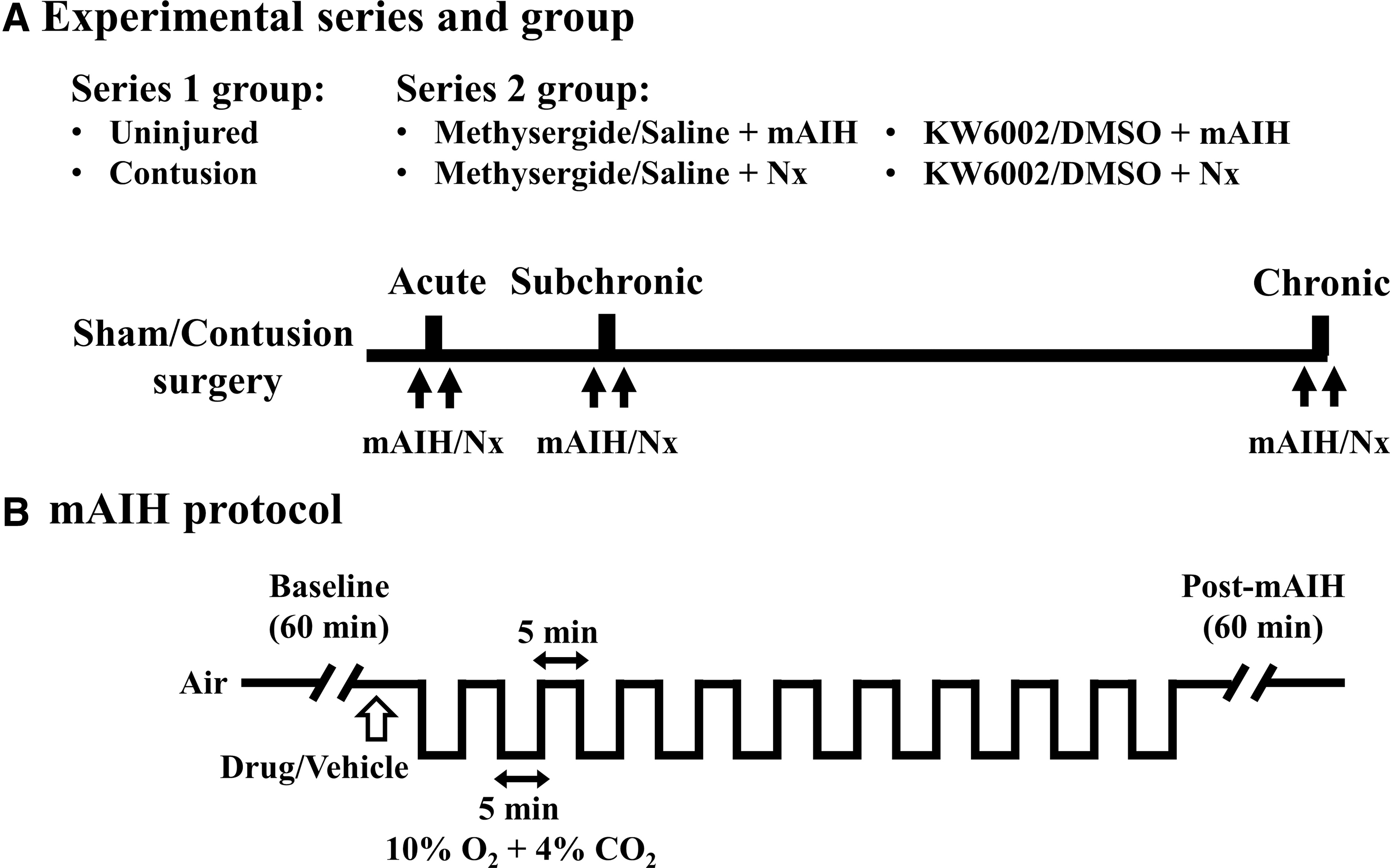
The timeline of experimental series
In the second experiment, contused animals were divided into four groups: 1) methysergide/saline + mAIH, n = 19; 2) KW6002/DMSO + mAIH, n = 17; 3) methysergide/saline + normoxia, n = 4; and 4) KW6002/DMSO + normoxia, n = 4. These animals received a vehicle (saline or DMSO) and a receptor antagonist (methysergide or KW6002) after a 1-h baseline recording at the acute (vehicle: 3.2 ± 0.1 days, antagonist: 3.8 ± 0.1 days), subchronic (vehicle: 14.0 ± 0.2 days, antagonist: 14.5 ± 0.2 days), and chronic (vehicle: 55.9 ± 0.3 days, antagonist: 56.6 ± 0.4 days) injury stages. Acute intermittent hypoxia or a 100-min normoxia followed by a 1-h normoxia baseline recording were introduced at 5 min after administration of a vehicle or a receptor antagonist.
Phrenic motoneuron labeling
The impact of mid-cervical spinal cord contusion on the phrenic nucleus was evaluated in the animals used in the first experiment. After whole–body plethysmography measurement at the chronic injury stage, the animal was anesthetized with xylazine (10 mg/kg, s.c.) and ketamine (140 mg/kg, i.p.). The diaphragm was exposed by a laparotomy and retraction of the abdominal muscle, then the animal received a 40 μL retrograde tracer (cholera toxin subunit B conjugated with Alexa Fluro 488, 0.05%; ThermoFisher Scientific) painting on the bilateral diaphragm. Three days after the tracer application, the animal was anesthetized with urethane (1.6 g/kg, i.p.; Sigma) and perfused with heparin-saline (500 mL), 4% paraformaldehyde (250 mL; Alfa Aesar) and then 10% sucrose (Sigma) in 4% paraformaldehyde (250 mL). The cervical spinal cord tissue was dissected and placed in 30% sucrose (Sigma) in a phosphate-buffered saline until the tissue sank.
The spinal cord tissue was transversely sectioned (40 μm) via a Cryostat (CM 1520; Leica), and spinal cord slices were mounted on glass slides (S7441; MATSUNAMI), and cover slips with a mounting medium containing 4',6-diamidino-2-phenylindole (Sigma). The tracer-labeled phrenic motoneuron was observed and imaged by a fluorescence microscope (BX43, Olympus) coupled with a charge coupled device (DS-5Mc; Nikon). The number of tracer-labeled phrenic motoneurons was counted manually in every slice. The phrenic motoneuron number was presented as the total number in the right and left spinal cord. In addition, the phrenic motoneuron number in every 25 slices (i.e., 1 mm) was summed and presented as the number in each 1 mm from the lesion epicenter to depict the distribution of phrenic motoneurons following mid-cervical spinal cord contusion.
Spinal cord histology
The lesion severity following mid-cervical spinal cord contusion was evaluated in the animals used in the second experiment. The anesthesia, perfusion, spinal cord tissue process, and section procedures were identical as described in the phrenic motoneuron labeling section. The spinal cord slices were mounted on glass slides and stained with cresyl violet (Acros Organics) in a standard protocol. The morphology of the spinal cord slice was examined by an upright microscope (BX43; Olympus) connected to a charge coupled device (DS-5Mc; Nikon). The area of the spinal cord and lesion cavity at the lesion epicenter in the left and right spinal cord were quantified using ImageJ software. The lesion area was presented as an absolute value (mm2) and normalized by a percentage of hemi-cord (% hemi-cord).
Statistical analysis
The ventilatory parameters (e.g., respiratory frequency, tidal volume, minute ventilation) were calculated by FinePointe software (Data Sciences International) and exported to an Excel file in a 1-min bin. A stable 10-min data during the normoxic breathing (i.e., lowest coefficient of variation of respiratory frequency) was averaged and defined as the baseline (BL) value. The first acute hypercapnic-hypoxic response was normalized by a percentage of the baseline value (% BL) and presented as the averaged data during the last 2 min of hypoxia. The ventilatory behaviors after mAIH were also normalized (% BL) and presented as the data averaged from 5 to 65 min after the last hypercapnic-hypoxic treatment. The ventilatory parameters during the baseline acute hypercapnic-hypoxic response and after mAIH were analyzed using a two-way repeated measure analysis of variance (ANOVA) followed by a Student-Newman-Keuls (SNK) post hoc test (factor one: animal group [uninjured vs. contusion] or treatment [mAIH vs. normoxia or vehicle vs. drug]; factor two: time-point [acute, subchronic and chronic injury stage]).
The impact of cervical spinal cord contusion on the phrenic motoneuron number was evaluated using a two-way mixed design ANOVA followed by a SNK post hoc test (factor one: left vs. right side; factor two: animal group or position of the slice). A paired t-test was used to compare the spinal cord area and lesion area of the left and right sides in contused animals. All data are expressed as the mean ± standard error. A p value lower than 0.05 was considered statistically significant.
Results
Animal weight
Table 1 provided the animal number and body weight in the two series of experiments. The body weight gradually increased in both uninjured and contused animals at the subchronic and chronic injury stage (p < 0.05; Table 1); however, contused animals usually had a lower body weight than uninjured animals (p < 0.05; Table 1).
Baseline breathing pattern between uninjured and contused animals
Representative examples of respiratory airflow recorded in contused animals at the acute, subchronic, and chronic injury stage are shown in Figure 2. Respiratory frequency was significantly higher in contused animals than uninjured animals at the acute (uninjured: 99 ± 5 breaths/min; contused: 132 ± 4 breaths/min) and subchronic (uninjured: 91 ± 3 breaths/min; contused: 119 ± 6 breaths/min) injury stages (p < 0.01; Fig. 3A). The higher respiratory frequency was mainly caused by a lower expiratory duration following cervical contusion (p < 0.01; Table 2). The contused animals are able to regain normal respiratory frequency at the chronic injury stage; however, they still exhibit lower tidal volume and lower mean inspiratory flow rate at all time-points post-injury (Fig. 3B; Table 2). Specifically, the tidal volume of acutely contused animal (0.96 ± 0.03 mL) was significantly lower than that of uninjured animals (1.30 ± 0.05 mL; p < 0.01; Fig. 3B). Until the chronic injury stage, the contused animal still had a significantly lower tidal volume (uninjured: 1.98 ± 0.11; contused: 1.55 ± 0.06 mL; p < 0.05; Fig. 3B). The minute ventilation was comparable between uninjured and contused animals at the acute and subchronic injury stages (p > 0.05; Fig. 3C), but chronically contused animals had a lower minute ventilation than uninjured animals (p < 0.05; Fig. 3C).
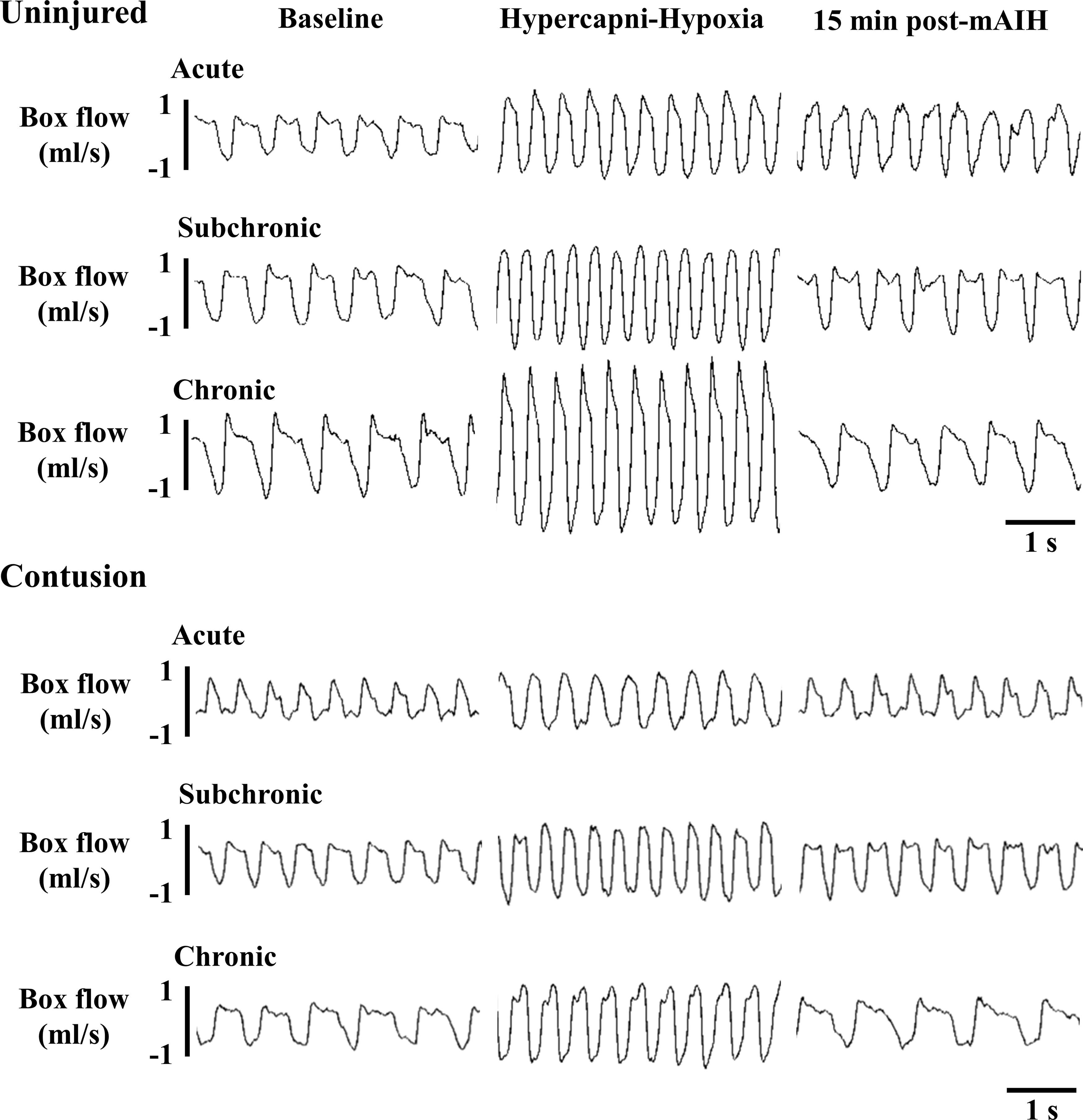
Representative examples of the respiratory airflow recorded in contused animals using the whole–body plethysmography during the baseline, first hypoxic treatment and 15 min after mild acute intermittent hypercapnic-hypoxia (mAIH) at the acute, subchronic, and chronic injury stages.
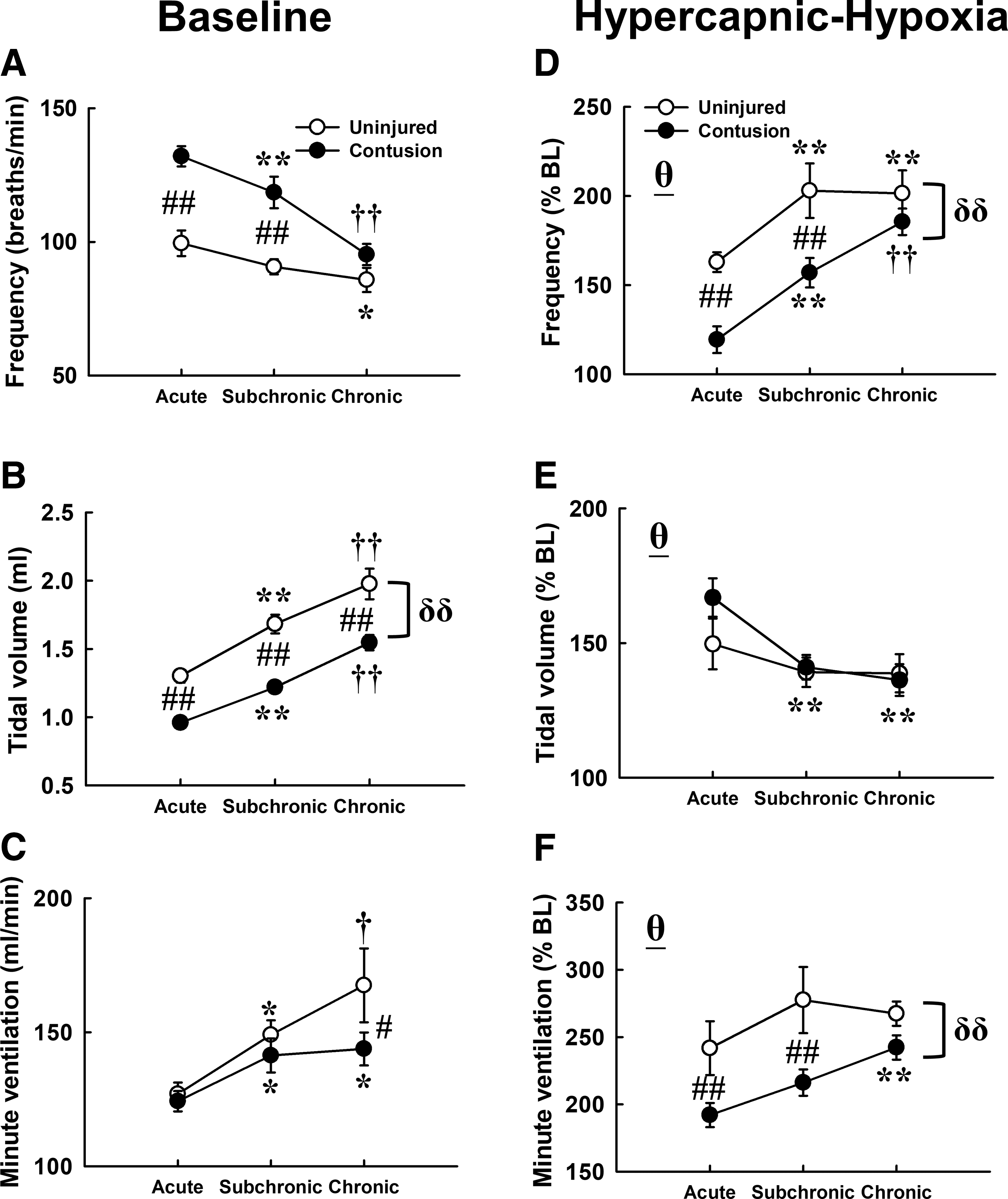
The baseline breathing pattern and first hypercapnic-hypoxic ventilatory response of uninjured and contused animals at the acute, subchronic, and chronic injury stages. The baseline breathing parameters were presented as an absolute value
Respiratory Cycle Duration and Mean Inspiratory Flow Rate of Uninjured and Contused Animals
p < 0.01 vs. Acute.
†p < 0.05.
††p < 0.01 vs. Acute and Subchronic.
p < 0.05.
p < 0.01, significant difference between uninjured and contused animals at specific injured stages.
p < 0.05.
p < 0.01, significant group effect (uninjured vs. contusion).
Acute hypercapnic-hypoxic response between uninjured and contused animals
Ventilatory parameters (respiratory frequency, tidal volume, and minute ventilation) were significantly enhanced during hypoxic treatment in both uninjured and contused animals at all time-points after spinal surgery (p < 0.01; Fig. 3D-F). However, the augmented hypercapnic-hypoxic response of the respiratory frequency and minute ventilation were relative weaker in contused animals than uninjured animals (p < 0.01; Fig. 3D and 3F). Specifically, the minute ventilation of uninjured animals can reach up to 242 ± 20% BL and 278 ± 25% BL during hypoxia at the acute and subchronic stages, but the hypercapnic hypoxia-induced increases of minute ventilation were less than 220% BL in contused animals (acute: 192 ± 9% BL; subchronic: 216 ± 10% BL; Fig. 3F). The blunted hypercapnic-hypoxic ventilator response of contused animals can gradually recover to 242 ± 9% BL at the chronic injury stage (Fig. 3F).
Acute intermittent hypercapnic hypoxia-induced ventilatory response between uninjured and contused animals
The minute ventilation of uninjured animals was significantly enhanced following mAIH in uninjured animals; however, this ventilatory long-term facilitation was only observed at the acute stage and was not induced by 100-min normoxia (i.e., time control) (Fig. 4C). Interestingly, mAIH can significantly increase the minute ventilation in contused animals from the acute to chronic injury stage (acute: 113 ± 4% BL; subchronic: 113 ± 3% BL; chronic: 110 ± 3% BL; p < 0.05; Fig. 4F). The augmented minute ventilation was mainly caused by an enhancement of the tidal volume in both groups (p < 0.01; Fig. 4B and 4E).
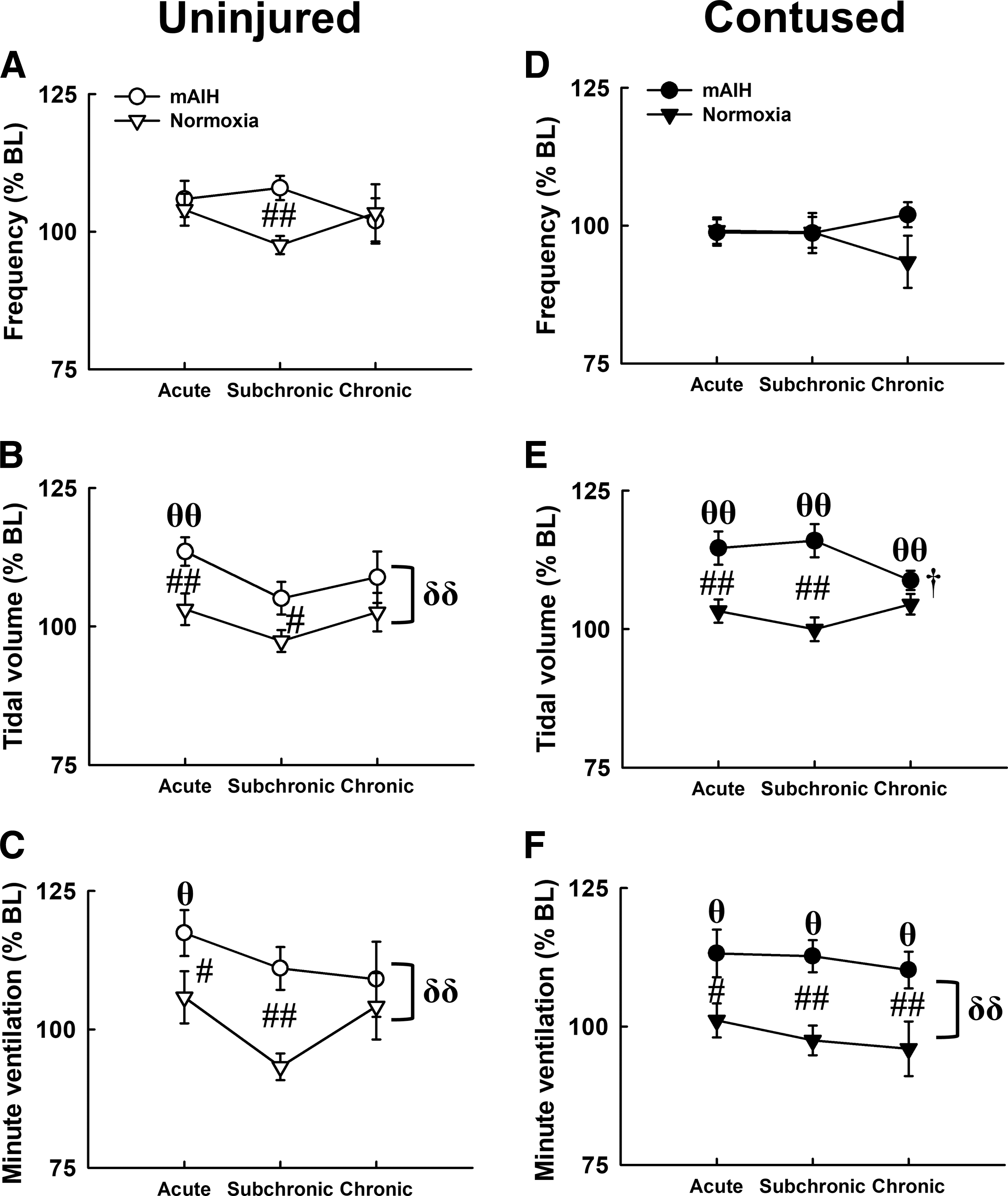
The breathing pattern of uninjured
Acute hypercapnic-hypoxic response following blockade of serotonin receptors or adenosine 2A receptors in contused animals
Acute hypercapnic-hypoxia induced a significant increase in respiratory frequency, tidal volume, and minute ventilation in contused animals that received a vehicle (saline or DMSO; p < 0.01; Fig. 5). A blockade of serotonin receptors by methysergide did not modulate the acute hypercapnic-hypoxic response; however, administration of an adenosine 2A receptor antagonist (KW6002) significantly augmented the frequency response at the subchronic and chronic injury stages (p < 0.01; Fig. 5D). Specifically, the respiratory frequency reached to ∼200% BL (subchronic: 202 ± 11% BL; chronic: 206 ± 10% BL) in contused animals that received DMSO administration, respectively (Fig. 5D). This hypercapnic hypoxia-induced increase in respiratory frequency could be significantly augmented to 232 ± 13% BL and 234 ± 11% BL following KW6002 administration at the subchronic and chronic injury stages, respectively (p < 0.01; Fig. 5D).
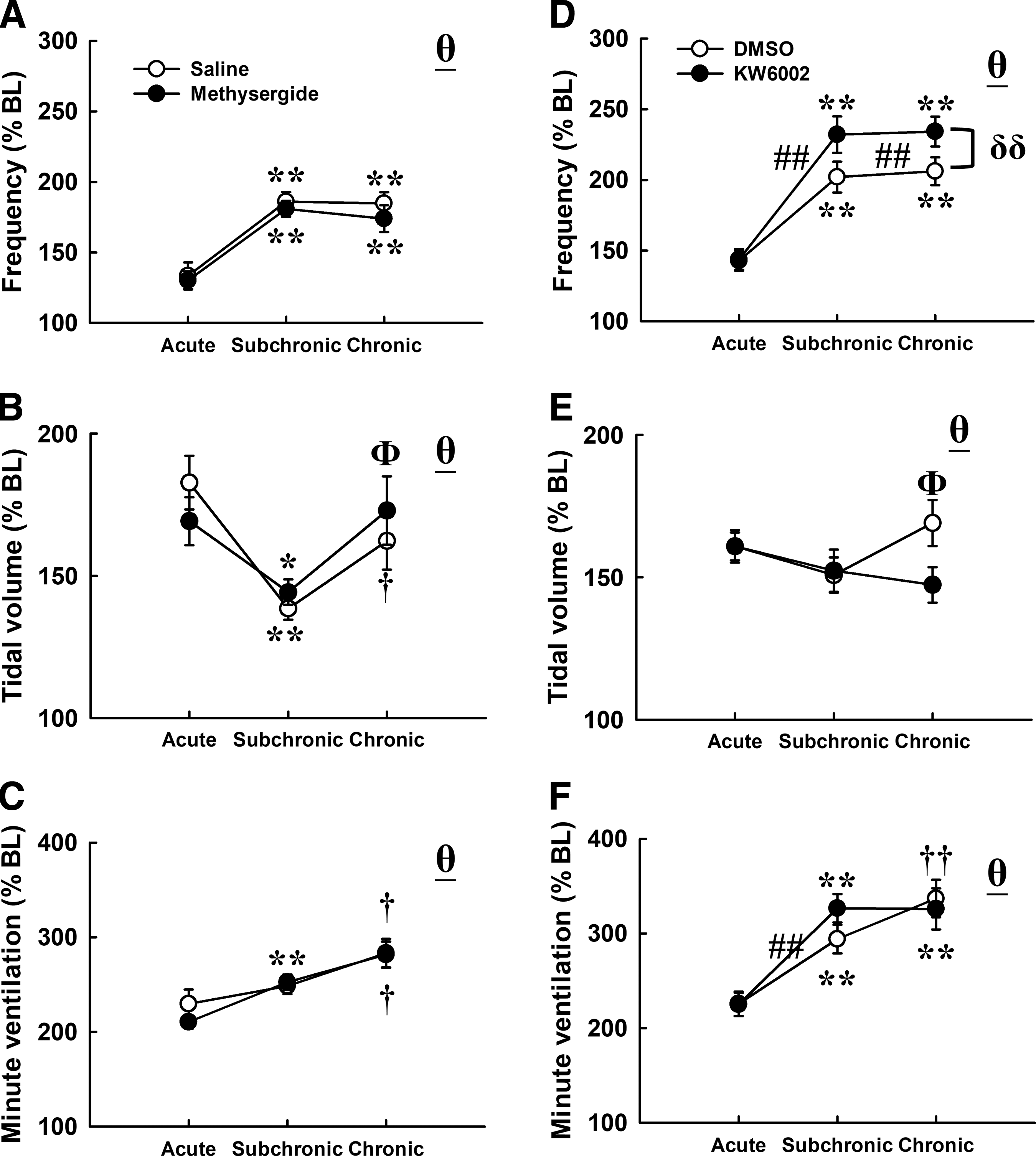
The first hypercapnic hypoxic ventilatory response of contused animals after a blockade of serotonin receptors
Acute intermittent hypercapnic hypoxia-induced ventilatory response following blockade of serotonin receptors or adenosine 2A receptors in contused animals
The minute ventilation and tidal volume were significantly elevated following mAIH in contused animals that received vehicle (saline or DMSO) treatment at all time-points post-injury (p < 0.05; Fig. 6B, 6C, 6E, 6F). This ventilatory long-term facilitation was significantly attenuated in contused animals that received methysergide administration due to a reduction of respiratory frequency at the subchronic (saline: 104 ± 3% BL; methysergide: 97 ± 2% BL) and chronic (saline: 105 ± 3% BL; methysergide: 91 ± 3% BL) injury stages (Fig. 6A). On the contrary, mAIH-induced ventilatory facilitation was augmented by KW6002, which significantly enhanced the respiratory frequency at the subchronic (DMSO: 105 ± 2% BL; KW6002: 116 ± 5% BL) and chronic (DMSO: 108 ± 4% BL; KW6002: 124 ± 6% BL) injury stages (p < 0.01; Fig. 6D). Notably, the tidal volume response following mAIH was not robustly changed by methysergide or KW6002 (Fig. 6B and 6E).
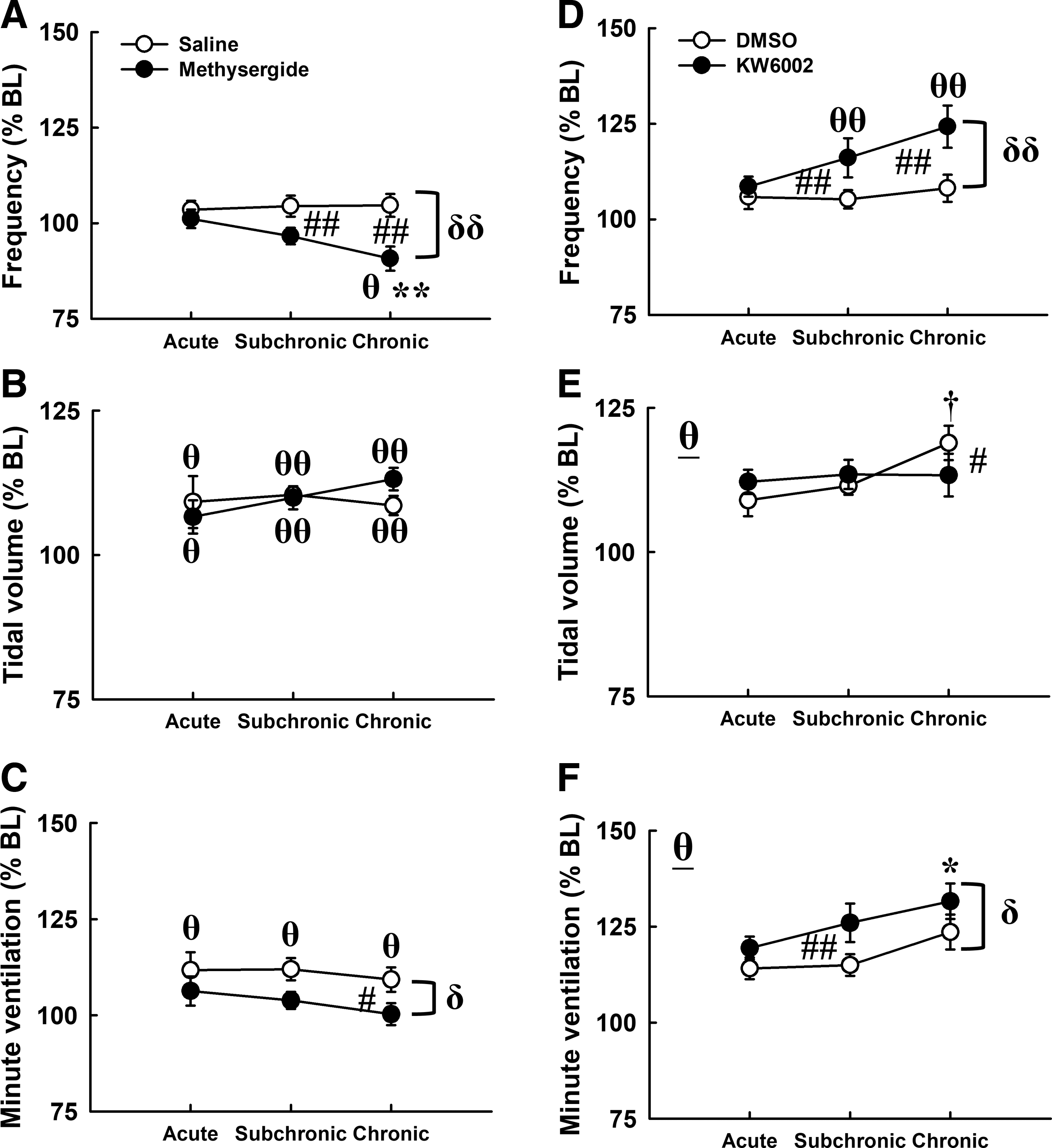
The influence of serotonin receptors
Table 3 shows the influence of methysergide or KW6002 on the ventilatory response after 100-min normoxia (i.e., time control). The result demonstrated that the minute ventilation was not significantly altered by methysergide; however, there is a trend of increasing minute ventilation after KW6002 administration.
Changes in the Minute Ventilation (% BL) of Contused Animals that Received Methysergide or KW6002 after the Normoxic Time Control Period
BL, baseline; DMSO, dimethyl sulfoxide.
Phrenic motoneuron number
The impact of mid-cervical contusion on the phrenic nucleus was evaluated in animals in the first experiment (uninjured, n = 6; contused, n = 10). Representative examples of transverse cervical spinal sections are demonstrated in Fig. 7. The tracer-labeled phrenic motoneurons were located at the ventral horn of the bilateral spinal cord grey matter in an uninjured animal (Fig. 7A). The phrenic motoneurons at the left spinal cord were lost or damaged at the lesion epicenter (Fig. 7B) in contused animals. The total number of tracer-labeled phrenic motoneurons at the left side was significantly lower in contused animals (112 ± 11) than in uninjured animals (207 ± 20; p < 0.01; Fig. 7C). Figure 7D shows the distribution of phrenic motoneurons of the bilateral spinal cord in contused animals. The result demonstrated that phrenic motoneurons were primarily lost at the middle portion of the phrenic nucleus following mid-cervical spinal cord contusion.

Representative example of transverse cervical spinal cord section in an uninjured
Spinal cord histology
The lesion severity was evaluated in animals in the second experiment (n = 44). A representative example of a transverse cervical spinal cord section of a contused animal is shown in Figure 8A. The spinal cord area of the left side (i.e., injured side, 1.07 ± 0.04 mm2) was significantly smaller than that of the right side (i.e., uninjured side, 1.83 ± 0.06 mm2; p < 0.01; Fig. 8B). Unilateral mid-cervical spinal cord contusion caused a significant cavity at the left spinal cord. The lesion cavity at the lesion epicenter is 0.41 ± 0.04 mm2 and occupied 35.3 ± 2.5% hemi-cord (Fig. 8C, 8D).
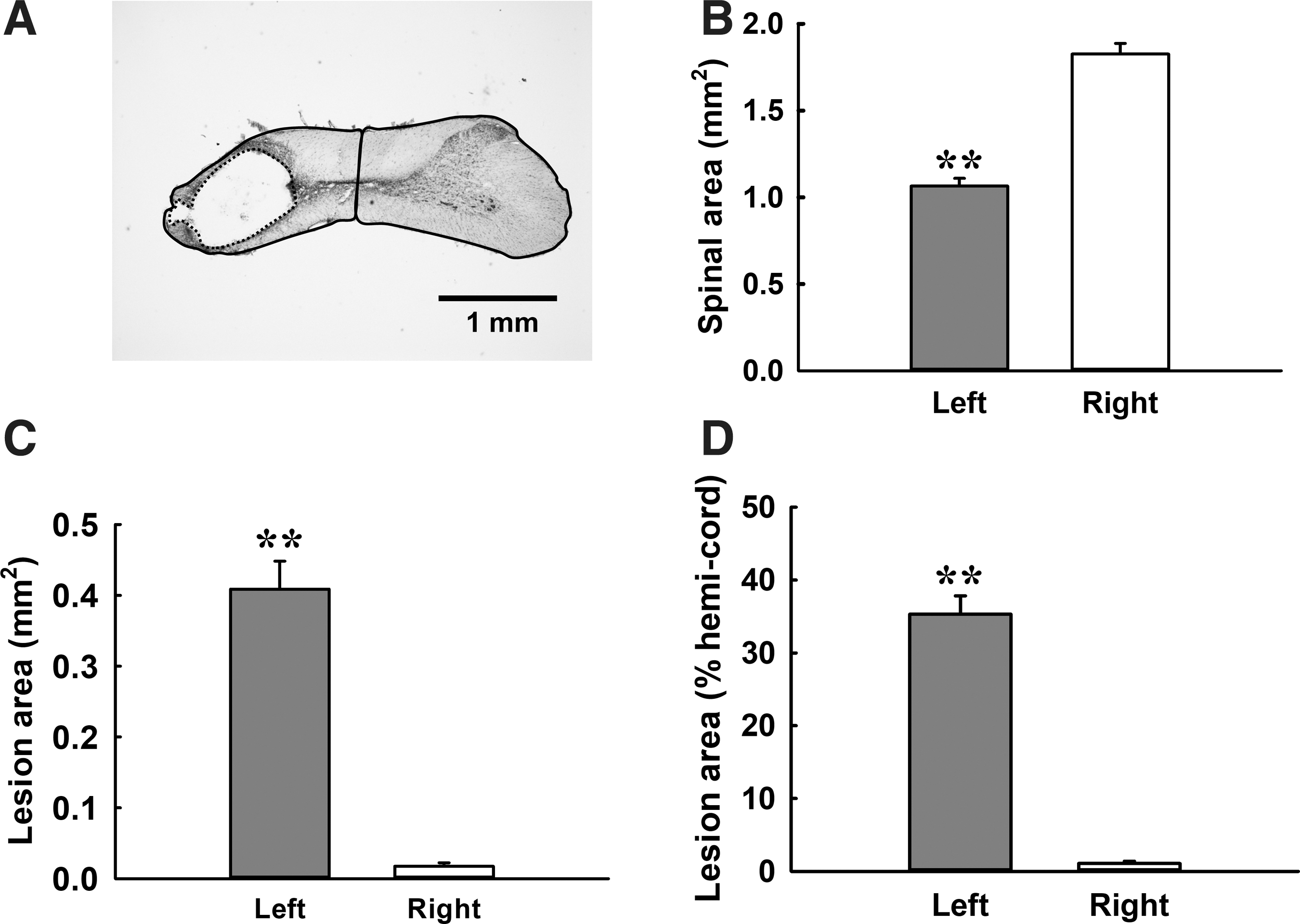
Cervical spinal cord morphology at the chronic injury stage following unilateral mid-cervical spinal cord contusion.
Discussion
There are three major findings in the present study. First, unilateral mid-cervical spinal cord contusion caused a long-term reduction in the tidal volume and attenuated the hypercapnic-hypoxic frequency response. Second, the minute ventilation could be enhanced following mAIH (i.e., ventilatory long-term facilitation) from the acute to chronic injury stage in contused animals. Third, ventilatory long-term facilitation was attenuated by a blockade of serotonin receptors but enhanced by an adenosine A2 receptor antagonist at the subchronic and chronic injury stages. Together, these findings suggested that mAIH is effective to enhance the ventilation from the acute to chronic injury stage, and the pre-treatment with an inhibitor of adenosine 2A receptor could be a potential strategy to enhance the therapeutic effectiveness of mAIH on respiratory recovery following mid-cervical spinal cord contusion.
Critique of methods
The composition of hypoxic gas used for mAIH paradigm in the present study is 10% O2 + 4% CO2; therefore, elevation of CO2 level during respiratory stimulus may have an impact on the ventilatory response in our model. The functional role of CO2 on hypoxia-induced respiratory neuroplasticity has been reported. For example, Harris and colleagues demonstrated that intermittent hypoxia-induced ventilatory long-term facilitation can be evoked only in the presence of sustained elevated CO2 in awake human. 25 Similarly, augmentation of ventilation following acute continuous hypoxia can be also observed when the CO2 is maintained above the physiological value. 26 These results suggested that increased CO2 may be an important factor for induction of respiratory neuroplasticity. 27 In addition, intermittent hypercapnia alone can also evoke another form of respiratory neuroplasticity (i.e., long-term depression), 28 which may counteract the facilitatory effect of intermittent hypoxia when both stimuli were combined. 29,30 Notably, above-mentioned studies were conducted in healthy subjects or normal animals. The future studies are necessary to elucidate the function of CO2 on hypoxia-induced respiratory neuroplasticity following cervical spinal cord injury.
The other potential concern is that the drugs (methysergide and KW6002) used in the present study were provided via systemic intraperitoneally injection. Bavis and Mitchell demonstrated that carotid-denervated animals still exhibited phrenic long-term facilitation following intermittent hypoxia. 31 Moreover, long-term facilitation of respiratory rhythm from the pre-Bötzinger complex could be evoked in in vitro brain slice preparation. 32,33 These results suggested that direct hypoxic effect on the central nervous system can contribute to induction of respiratory neuroplasticity. Accordingly, the alteration of ventilatory behaviors following drug administration could be caused by the action of these drugs on both supraspinal and spinal level. In addition, methysergide is a broad-spectrum serotonin receptor antagonist; thus, the present study can only conclude that the inhibitory effects of methysergide on mAIH-induced respiratory recovery are mediated by preventing serotonin effects. It remains to be investigated which specific type of serotonin receptor is involved in induction and/or maintenance of ventilatory long-term facilitation following mAIH in unanesthetized mid-cervical contused animals.
The baseline breathing pattern following mid-cervical spinal cord contusion
Unilateral mid-cervical spinal cord contusion caused a significant reduction in the tidal volume in contused animals at all time-points post-injury. Based on the results from the spinal cord histology and phrenic motoneuron labeling, we considered that the reduced tidal volume of contused animals was attributed by interruption of bulbospinal respiratory pathways and damage to phrenic motoneurons. Although the tidal volume was reduced following mid-cervical spinal cord contusion, the tidal volume of contused animals can gradually increase at the subchronic and chronic injury stages. This spontaneous and partial recovery of the tidal volume also was observed in other types of cervical spinal cord injury (e.g., high-cervical hemisection, mid-cervical hemisection, and midline mid-cervical contusion). 22,23
Several potential mechanisms could contribute to the recovery of the tidal volume following unilateral mid-cervical spinal cord contusion. First, the phrenic motoneurons were innervated by both ipsilateral and contralateral pathways from the brainstem and/or spinal interneurons. 34,35 Therefore, the phrenic motoneurons below the lesion could still be activated by the latent crossed phrenic pathway to promote the tidal volume recovery despite the ipsilateral bulbospinal pathway being interrupted by the unilateral cervical contusion. 36 Second, compensatory increases in supraspinal and spinal respiratory drives to the residual phrenic motoneurons located above and contralateral to the lesion could be involved in the induction of respiratory recovery following cervical spinal cord injury. 37 –39 Third, the respiratory activity of the intercostal muscles could also be evoked to maintain and improve the tidal volume in injured animals following C2 hemisection as well as C4 contusive injury. 4,20,40
The acute hypoxic response following mid-cervical spinal cord contusion
Hypoxia induced a significant increase in the respiratory frequency, tidal volume, and minute ventilation in both uninjured and contused animals at all time-points post-injury as expected. However, the hypercapnic-hypoxic frequency response was blunted in contused animals at the subchronic and chronic injury stages. The blunted frequency response was also noticed in cervical spinal cord injury animals in response to other types of respiratory stimuli (i.e., lung inflation, intra-jugular capsaicin administration). 2,22,41,42 We proposed two factors that may contribute to the blunted hypercapnic-hypoxic frequency response following mid-cervical spinal cord contusion. First, the baseline respiratory frequency was already higher in contused animals; therefore, there may a ceiling effect preventing a further increase in respiratory frequency during hypoxia. Second, cervical spinal cord injury is associated with changes of supraspinal respiratory network, 43 which may modulate hypercapnic-hypoxic frequency response in contused animals.
Ventilatory long-term facilitation following mid-cervical spinal cord contusion
The minute ventilation following mAIH was significantly enhanced in uninjured animals at the acute, but not subchronic and chronic stages. This observation is similar with a previous report demonstrating that ventilatory long-term facilitation was greater in 1- vs. 2-month old rats, 44 and suggest that aging may reduce the neuroplasticity of the respiratory motor system. Several studies indicated that mAIH was unable to induce long-term facilitation of phrenic or diaphragmatic output from the acute to subchronic injury stages in C2 hemisected animals. 11,12 However, our current study demonstrated that contused animals can exhibit ventilatory long-term facilitation following mAIH from the acute to chronic injury stages. This finding implies that the inhibitory effect of aging may be counteracted by mid-cervical spinal cord contusion. Further, different O2 and CO2 level (e.g., 10% O2 vs. 10% O2 + 4% CO) used for intermittent hypoxia protocol may cause differential long-term facilitation response. 25 Lastly, there may be a distinct mechanism underlying mAIH-induced respiratory recovery in high-cervical hemisected versus mid-cervical contusion injury models at distinct injury stages.
The magnitude of ventilatory long-term facilitation of cervical contused animal is ∼10–20% above the baseline in the current study. Other animal studies also showed that the baseline tidal volume or minute ventilation during maximal chemoreceptor activation (i.e., 7% CO2, 10.5% O2) can increase ∼10% after daily acute intermittent hypoxia. 18,19 Interestingly, the minute ventilation of subjects with spinal cord injury can be also elevated to ∼20% following acute intermittent hypoxia. 8,9 These results suggested that acute or daily acute intermittent hypoxia can significantly improve the respiratory function after cervical spinal cord injury. In our study, the expression of ventilatory long-term facilitation in contused animals was primarily due to augmentation of the tidal volume in response to mAIH. It has been shown that cervical spinal cord injury is associated with increases in ipsilateral phrenic motoneuron excitability 41,45 and compensatory function in contralateral diaphragmatic and intercostal motor outputs. 4,7,37,46,47 Therefore, we speculate that the improvement of the tidal volume could be caused by increasing the following respiratory motor outputs: 1) contralateral phrenic and intercostal motoneurons; 2) ipsilateral phrenic motoneurons above the contusion injury; and 3) ipsilateral phrenic and intercostal motoneurons below the contusion injury.
Acute hypercapnic-hypoxic response following blockade of serotonin receptor and adenosine 2A receptor in contused animals
The acute hypoxic response is primarily triggered by activation of peripheral chemoreceptor receptors, which provide excitatory drives to the respiratory circuit and then enhance respiratory motor outputs. 48 Our current study demonstrated that methysergide did not modulate the ventilatory behaviors during hypercapnic-hypoxia, but KW6002 significantly enhanced the hypercapnic-hypoxic frequency response. These results indicated that adenosine may have an inhibitory effect on respiratory frequency during hypoxia through activation of adenosine 2A receptors. Kobayashi and colleagues demonstrated that adenosine 2A receptors of the carotid body mediated adenosine-induced inhibition of voltage-dependent calcium current in type I cells. 49 Therefore, a blockade of an adenosine 2A receptor in the current study may augment hypoxia sensitivity of the carotid body and subsequently increase afferent inputs to enhance the hypercapnic-hypoxic ventilatory response.
In addition, Koos and colleagues observed that hypoxia-induced ventilatory depression was attenuated by a blockade of adenosine 2A receptors in lambs. 50 Accordingly, augmented hypercapnic-hypoxic ventilatory response following KW6002 administration could also be due to a dis-inhibitory effect induced by the blockade of adenosine 2A receptors. Notably, the effect of KW6002 was only observed at the subchronic and chronic injury stages, suggesting that a modulatory effect of adenosine 2A receptors on the hypercapnic-hypoxic ventilatory response is time-dependent in mid-cervical contused animals.
Ventilatory long-term facilitation following blockade of serotonin receptor and adenosine 2A receptor in contused animals
Ventilatory long-term facilitation was modulated in contused animals that received either methysergide or KW6002 administration, suggesting both serotonin and adenosine are involved in the regulation of respiratory neuroplasticity following mAIH. Interestingly, the influence of methysergide or KW6002 on minute ventilation was mainly due to changes of respiratory frequency but not tidal volume. These results suggest that mAIH-induced long-term facilitation of the tidal volume is independent on serotonin and adenosine 2A receptors, which is different from the mechanism underlying phrenic long-term facilitation in anesthetized uninjured and high-cervical hemisected animals or compensatory increases of the contralateral diaphragm in unanesthetized cervical hemisected animals. 11,15,20 These differences could be due to a distinct injury model (i.e., mid-cervical C4 contusion vs. high-cervical C2 hemisection) and an intermittent hypoxic condition (i.e., intermittent isocapnic, poikilocapnic or hypercapnic hypoxia).
The respiratory frequency was significantly reduced in animals that received both methysergide and mAIH; however, methysergide or mAIH alone did not influence the respiratory frequency. These findings suggest that mAIH and a blockade of serotonin receptors may be synergistic to inhibit respiratory frequency. In addition, the hypoxic condition used in the current study is isocapnic hypoxia with 10% O2 plus 4% CO2. Moreover, we cannot exclude that mild intermittent CO2 exposures may also have had an impact on the ventilatory response. It has been shown that intermittent hypercapnia caused a long-term depression in the phrenic frequency and amplitude. 28 Valic and colleagues further demonstrated that administration of methysergide can reveal intermittent hypercapnia-induced phrenic long-term depression. 51 Accordingly, a blockade of serotonin receptors by methysergide may uncover the potential respiratory depression induced by mAIH in the current model.
Gonzalez-Rothi and colleagues and Turner and colleagues proposed that there is a cross-talk inhibition between intermittent hypoxia-induced activation of Gq-coupled metabotropic receptors (e.g., serotonin 2 receptor) and Gs-coupled metabotropic receptors (e.g., adenosine 2A receptor). 16,52 Several studies have demonstrated that a blockade of an adenosine 2A receptor can enhance mAIH-induced long-term facilitation of respiratory motor outputs (e.g., phrenic, diaphragm, respiratory frequency) in uninjured animals. 12,53 Our current results also support this notion showing that ventilatory long-term facilitation of contused animals was enhanced by an adenosine 2A receptor antagonist (KW6002). However, the excitatory effect of KW6002 on the minute ventilation was mainly mediated by increasing respiratory frequency. This result suggested that adenosine 2A receptors may constrain the effect of mAIH at the supraspinal level. In addition, we noticed there is a modest increase of respiratory frequency in animals that received KW6002 without mAIH. Therefore, KW6002 may generally enhance the peripheral chemoreceptor sensitivity and, in turn, increase respiratory frequency.
Dougherty and colleagues proposed that there is a shift in mechanism underlying daily mAIH-induced respiratory recovery from a serotonin-independent mechanism at one-two weeks post-injury to a serotonin-dependent mechanism at 7–8 weeks post-injury in cervical hemisected rats. 18 The current study did not observe such transition mechanism but noticed that the effect of methysergide and KW6002 was gradually enhanced from the acute to chronic injury stages, suggesting the modulatory effect of serotonin and adenosine on mAIH-induced respiratory response may be time-dependent.
Conclusion
The present study demonstrated that mAIH can improve respiratory function in mid-cervical spinal cord contused animals from the acute to chronic injury stages. This augmentation in ventilation, named ventilatory long-term facilitation, was attenuated by a serotonin receptor antagonist, but can be enhanced after systemic adenosine 2A receptor inhibition with KW6002 injection. Indeed, pre-treatment of an adenosine 2A receptor antagonist may amplify the therapeutic effectiveness of mAIH in mid-cervical contused animals. However, the effect of KW6002 is mainly mediated by regulating central respiratory frequency. We suggest that there is a need to explore the mechanism underlying mAIH-induced tidal volume recovery in a clinically relevant mid-cervical contusion model, which can provide us with opportunities to develop potential therapies improving the breathing capacity in patients with cervical spinal cord injury.
Footnotes
Acknowledgments
Support for this work was provided by grants from the Ministry of Science and Technology (MOST 105-2628-B-110-002-MY3 & 108-2636-B-110-001), Higher Education Sprout Project (07C030111) and NSYSU-KMU Joint Research Project (107-I001).
Author Disclosure Statement
No competing financial interests exist.