
Abstract
Traumatic brain injury (TBI) is a leading cause of death and disability in the United States, with children who sustain a TBI having a greater risk of developing long-lasting cognitive, behavioral, and motor function deficits. This has led to increased interest in utilizing large animal models to study pathophysiologic and functional changes after injury in hopes of identifying novel therapeutic targets. In the present study, a controlled cortical impact (CCI) piglet TBI model was utilized to evaluate cognitive, motor, and histopathologic outcomes. CCI injury (4 m/sec velocity, 9 mm depression, 400 msec dwell time) was induced at the parietal cortex. Compared with normal pigs (n = 5), TBI pigs (n = 5) exhibited appreciable cognitive deficiencies, including significantly impaired spatial memory in spatial T-maze testing and a significant decrease in exploratory behaviors followed by marked hyperactivity in open field testing. Additionally, gait analysis revealed significant increases in cycle time and stance percent, significant decreases in hind reach, and a shift in the total pressure index from the front to the hind limb on the affected side, suggesting TBI impairs gait and balance. Pigs were sacrificed 28 days post-TBI and histological analysis revealed that TBI lead to a significant decrease in neurons and a significant increase in microglia activation and astrogliosis/astrocytosis at the perilesional area, a significant loss in neurons at the dorsal hippocampus, and significantly increased neuroblast proliferation at the subventricular zone. These data demonstrate a strong relationship between TBI-induced cellular changes and functional outcomes in our piglet TBI model that lay the framework for future studies that assess the ability of therapeutic interventions to contribute to functional improvements.
Introduction
Traumatic brain injury (TBI) continues to be a leading cause of death and disability in children. Young children between the ages of 0 and 4 are the age group most likely to sustain a TBI as a result of a concussive injury from a fall and have the highest rate of TBI-related emergency department visits (1256 per 100,000 population). 1 Despite the perception that children's brains are potentially more “plastic” at a younger age, recent research findings suggest that the immature brain may be more vulnerable to early insult. 2,3 Injury severity plays an important role in neurocognitive outcomes. Mild TBI results in few, if any, cognitive impairments except in the case of repetitive TBI, but moderate-to-severe TBI often lead to significant impairments in cognition that persist over time. 4 –8
Disruptions in cognition can emerge in the acute and/or chronic phases of TBI. 9 Severe cognitive impairments in young children in the acute phase after TBI have been found to worsen rather than recover over time, suggesting that acute cognitive impairments may be a sensitive predictor of long-term outcomes. 10 Consequently, moderate-to-severe TBI is thought to disrupt existing neural networks and interfere with normal development, impeding the learning process and putting children developmentally and cognitively behind their peers even years after injury. 5 Similarly, TBI has been found to contribute to impairments in motor function that correlate with injury severity. 11,12 Although the majority of children who sustain a severe TBI regain independent ambulation, balance and speed of ambulation can remain impaired. 13 Like cognitive deficits, TBI at a young age is not associated with better long-term recovery in gait, which may be attributed to disruption of normal developmental processes. 13,14
Animal models of TBI have played an important role in elucidating the pathophysiology and functional outcomes associated with injury. The controlled cortical impact (CCI) model has gained popularity in recent years because it allows for precise control of injury location and severity, making it an ideal model with which to recapitulate the human condition of a focal TBI. 15 A number of studies have examined cognitive and motor function deficits and associated histological changes in rodents CCI models. 16 –19 More recently, interest in juvenile rodent CCI models have expanded in response to recent evidence that the immature brain may respond differently to injury than the mature brain. 20,21 TBI in juvenile rats has been shown to lead to cognitive deficits that persist into adulthood. 21 –23 Rodent TBI models are economically advantageous, have been found to be reproducible, and demonstrate similar pathological features to human TBI. 24 Significant credit can be attributed to pre-clinical rodent TBI studies for creating the framework of knowledge of TBI pathophysiology, functional responses, treatment strategies, and therapeutic targets.
However, rodents have a number of characteristics that may limit their translatability as a model. Compared with the lissencephalic rodent brain, the human brain is gyrencephalic, which significantly influences the movement of the brain in the skull during TBI, leading to increased brain deformation and transmission of forces at the moment of injury. 25 Differences in brain development between rodents and humans may also yield differing responses. 26 For example, the myelination time scale is much faster in developing rodent brains and the total amount of white matter is significantly reduced in rodents (12%) compared with humans (> 60%). 27,28 White matter has been found to respond differently and be more susceptible to injury, therefore key differences in white matter development may restrict the translational power of pediatric rodent models. 29 –31 These limitations likely contribute to the lack of effective therapies to treat TBI, which has led to increased interest in utilizing large animal models to study TBI.
The juvenile piglet has been implicated as a more translational model to study the effect of early life insults on neurodevelopment and to study the potential of novel therapeutic interventions. 31,32 The advantages of using pigs to study TBI processes over traditional rodent models have been recently reviewed. 33,34 For example, the pig and human brain are both gyrencephalic in structure. 35 The brain of the juvenile pig at approximately 4 weeks of age is developmentally more similar to the toddler-aged human brain in terms of both size and weight, compared with rodents. 36 –38 At this age, myelination is still incomplete. The juvenile piglet shares a more similar pattern and timing of myelination during brain development within human toddlers that continues into early adolescence, and total white matter ultimately reaches >60% in both adult pigs and humans. 27,39,40 The juvenile and human brain exhibit a similar time course of growth and development including gliosis, neurogenesis, and synaptogenesis. 41 –43 Compared with rodents, the location and orientation of the pig hippocampus is more similar to the human hippocampus which suggests pigs may be a more representative model for studying hippocampal-dependent cognitive processes and recovery mechanisms after TBI. 44,45
These characteristics suggest that the juvenile pig model may be more appropriate for studying the relationship between early life insults and consequential disruptions in normal developmental processes. As such, a number of recent studies have established the pig as an excellent model of TBI in both adults and juveniles. The piglet brain has been found to have a maturation-dependent response to injury and, similar to human infants, evidence of neuroblast proliferation and migration from the subventricular zone after injury. 36,46 –48 In addition, the pig has been well validated as an excellent model to study cognition and neurobehavioral disorders. 35 Cognitive and gross motor deficits have been observed in a non-impact axial rotation TBI piglet model, but few studies have been performed that examine the effects of CCI on cognition or motor function in a piglet model. 49,50
The purpose of this study was to assess the cognitive, motor, and corresponding histopathologic effects of CCI in a piglet TBI model. For evaluation of cognitive and neurobehavioral deficits after injury, the spatial T-maze test was utilized to measure hippocampal-dependent spatial memory, the open field test was utilized to measure exploratory behaviors, and the three-chamber social recognition test was used to assess sociability behaviors and social memory after injury. In addition, gait analysis was performed using a GAITFour® pressure mat to assess changes in pressure and spatiotemporal gait parameters after TBI. Finally, histopathologic analysis was performed and included the evaluation of neuronal cell death, microglia activation, and reactive gliosis in the perilesional cortex, neuronal cell death at the hippocampus, and neuroblast proliferation at the subventricular zone.
Methods
Animals
All work in this study was performed in accordance with the University of Georgia Institutional Animal Care and Use Committee guidelines. Two commercially bred pregnant Landrace-cross sows were housed in commercial farrowing crates and farrowed naturally. Each litter was removed from the farrowing crates two to three times per day starting immediately after birth for socialization with handlers until weaning. Socialization included holding and stroking the piglets and giving them milk replacer (Sav-a-Calf Grade A Ultra 24 Multispecies Milk Replacer, Milk Products LLC) as a treat. Preliminary studies showed that socialization was critical to reduce anxiety and increase compliance during handling and testing procedures. At weaning (3 weeks of age), five male piglets from each litter were chosen at random, group housed by litter, and fed a nutritionally complete pelleted starter diet ad libitum for the duration of the study. Room temperature was maintained at 26°C with a 12 h light/dark cycle and an overhead heat lamp provided supplemental heat.
Experimental design
Starting at 2 weeks of age, all piglets were habituated individually on the gait track twice a day to acclimate the piglets to a new environment and to being socially isolated. After weaning at 3 weeks of age, all piglets underwent gait track training for 3 to 4 days, then baseline gait analysis was collected on three separate days before the start of the study. At 4 weeks of age, each litter was randomly assigned to a treatment group: normal control (n = 5) or TBI (n = 5). Piglets at this age have been found to correspond with toddler-aged humans. 36 At the start of the study, TBI pigs underwent controlled cortical impact (CCI) surgery. Gait analysis was performed 1, 3, 7, 14, and 28 days post-TBI. The open field test was performed 1, 7, 14, and 26 days post-TBI. The three-chamber social recognition test was performed 15 days post-TBI. The spatial T-maze test was performed between 16–25 days post-TBI. Normal controls pigs underwent gait analysis collection, open field, social recognition, and spatial T-maze testing at corresponding time-points, but no TBI surgery was performed at the start of the study. All animals were sacrificed at 28 days for histological analysis of brain tissues.
Surgical preparation and controlled cortical impact
Piglets in the TBI group (n = 5) were initially anesthetized with 5% vaporized isoflurane mixed with oxygen using a nose cone until non-responsive to stimulation. For the duration of surgery, anesthesia was maintained within 2.5%-3.0% isoflurane and vitals were monitored every 5–10 min. The surgical site was prepared in a routine manner using betadine and 70% ethanol and covered with a sterile drape. A 4cm, left-sided incision was made at the top of the cranium to expose the underlying skull. A 20-mm craniectomy was performed using an air drill (Brasseler USA, Savannah, GA) at the left posterior junction of the coronal and sagittal sutures to exposure the underlying dura.
Pigs were secured in a controlled cortical impactor device (University of Georgia Instrument Design and Fabrication Shop, Athens, GA) designed specifically for pigs. The impactor tip (15 mm diameter) was centered over the exposed dura and a TBI was induced with the following parameters: 4 m/sec velocity, 9-mm depth of depression, and 400 msec dwell time. TBI was induced at the level of the parietal lobe. Although there are no clear criteria that classify mild, moderate, and severe TBI in pre-clinical models, especially in large animal models, CCI at these parameters in 4-week-old piglets is estimated to generate an injury that is moderate-to-severe in nature. 51 After TBI induction, the injury site was flushed with sterile saline, then the overlying skin was re-apposed with surgical suture. Anesthesia was discontinued and piglets were allowed to breath oxygen until recovered. Banamine (2.2 mg/kg; MERCK Animal Health) and butorphanol (0.2 mg/kg; Pfizer, New York, NY) were administered for analgesia and oxytetracycline (19.8 mg/kg; Norbrook Inc., USA, Overland Park, KS) was administered as an antibiotic during surgery and for an additional 4 days post-surgery. Once piglets were completely ambulatory, they were placed back in their home pens and monitored daily for health. Normal piglets (n = 5) did not undergo any surgical procedures.
Spatial T-maze test
The design of the spatial T-maze arena and testing procedure was adapted from Elmore and colleagues. 52 Briefly, a plus-shaped T-maze was constructed using metal grid paneling positioned over black rubber mats (Fig. 1A). North and south arms measured 2.1 m × 1.2 m and east and west arms measured 2.4 m × 1.2 m. A white shower curtain encircled the maze to reduce visual distractions and extra-maze environmental cues. Two reward bowls (University of Georgia Instrument Design and Fabrication Shop, Athens, GA) were fixed to the east and west arms. Reward bowls were filled with equal volumes of milk replacer to provide identical olfactory cues to both arms of the maze. One reward bowl was accessible and could be opened by the pigs using an inherent rooting motion and the other inaccessible reward bowl was secured shut. During initial days of testing, if pigs chose the correct reward bowl but were unable to open the lid, handlers manually opened the lid until the skill was mastered. Extra-maze visual cues (blue rectangle, gray circle, black triangle, red plus-sign) were attached to the white shower curtain. Visual cues were rotated around the arena to account for the location of the reward such that the red plus-sign was always located to the left of the reward, the blue rectangle to the right, the circle to the left of the incorrect reward, and the triangle to the right. To allow for alternating starting locations in the north and south arms, removable panels were constructed that could create a start box in the north arm and block off the south arm for “north start” configuration or a start box in the south arm and block off the north arm for a “south start” configuration (Fig.1B).
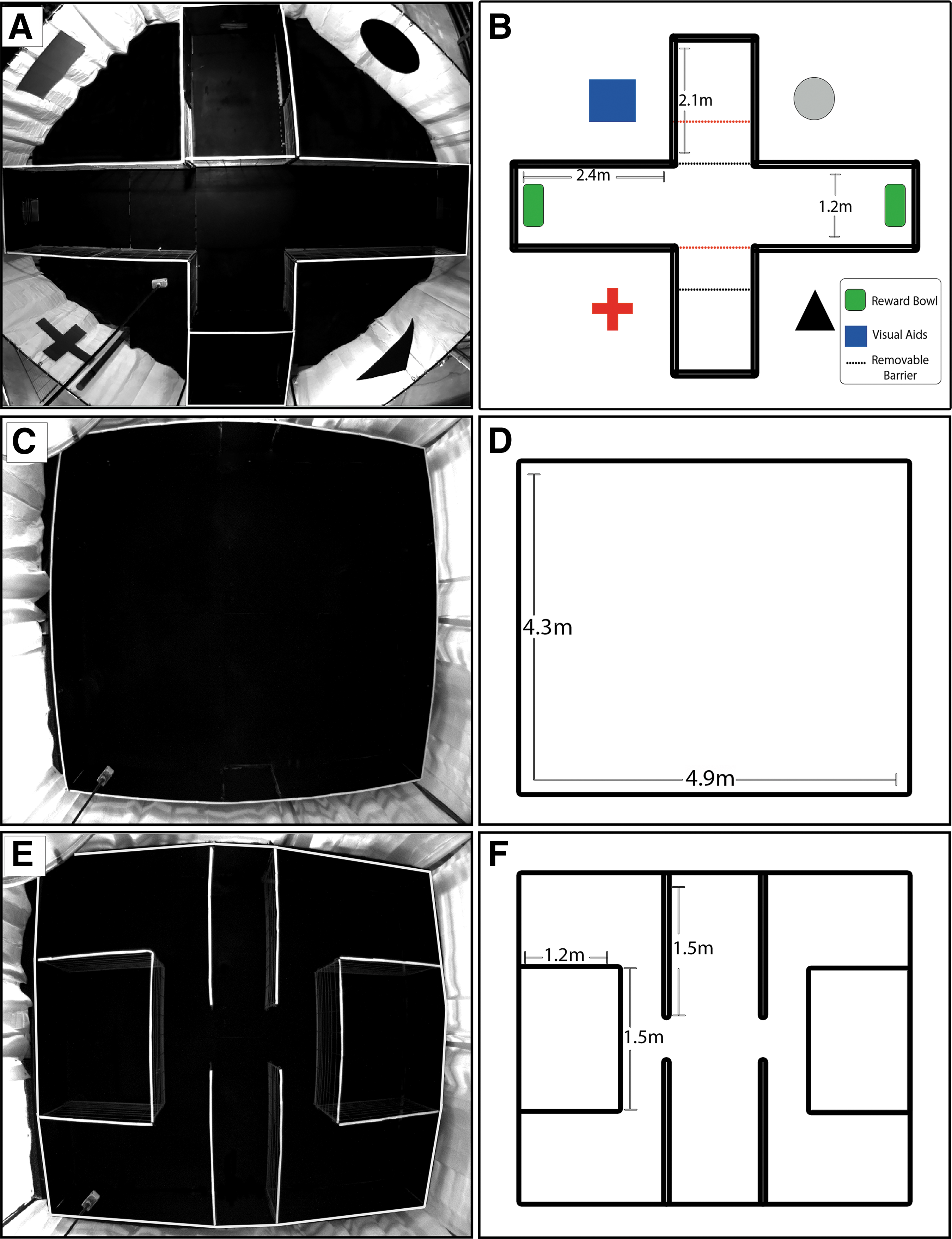
Photographic and schematic depictions of pig behavioral test arenas. Photograph of the T-maze arena depicts a start box in the south arm and restricted access to the north arm
Piglets began spatial T-maze testing after 6 h of food deprivation to increase motivation for the milk replacer food reward. The spatial T-maze test consisted of 6 days of acquisition testing in which the reward was assigned to either the east or west arms and 4 days of reversal testing where the reward arm was moved to the opposite arm. The extra-maze visual cues did not move during reversal testing. The reward location was assigned for each pig by running a spatial bias test on Day 1 of acquisition (A1). Pigs underwent five trials in which the reward was not accessible in either arm. The reward location was assigned to the arm visited least frequently by the animal to account for potential spatial bias. As such, 8/10 pigs for both TBI (n = 4) and normal (n = 4) groups exhibited a bias for the west arm and were therefore assigned to the east reward arm. Two of 10 pigs for the TBI (n = 1) and normal (n = 1) groups exhibited a bias for the east arm and were therefore assigned to the west reward arm. The starting location for each trial alternated from north to south in a pseudo-random order where no pig could start from one arm more than twice in a row. Alternating north and south arms encouraged pigs to solve the task using a hippocampal-dependent, allocentric mechanism whereby pigs use extra-maze visual cues to create spatial memories rather than a striatum-dependent, egocentric mechanism whereby muscle memory plays a role in reward selection, such as always turning the body to the left or right.
Pigs underwent 10 trials per day and had a maximum of 60 sec to reach the reward. If the pig exceeded 60 sec without choosing a reward, the pig was deemed non-compliant for that trial. For the first 2 days, if the pigs chose the incorrect arm they were corrected and taken to the correct arm. After 2 days, if the pigs chose the incorrect arm, they were returned to the start box for the next trial without any reward. Between each trial, any defecation and/or urination was removed and the entire arena was wiped down with 70% ethanol to remove any olfactory cues from trial to trial. In addition, the starting boxes were blacked out such that the pigs were unable to see the handlers refill the reward bowl between trials. An automatic tracking software (EthoVision XT 11.5; Noldus, Wageningen, the Netherlands) was used to measure latency to choice and proportion trials correct. Latency to choice was started manually once the removable panel was opened in the start box and automatically stopped once the piglet's nose-point entered a zone around a reward bowl, whether correct or incorrect. Proportion trials correct was measured as the proportion correct/total trials. The entire maze was cleaned between each pig to reduce olfactory bias.
Open field test
The open field arena measures 4.3 × 4.9 m and was constructed using metal grid paneling positioned over black rubber mats (Fig. 1C, 1D). White curtains were hung around the arena to reduce visual distractions during testing. For the open field test, pigs were placed into the arena from alternating north and south start/exit gates and allowed to freely explore the arena for 10 min. The exploratory behaviors distance traveled and velocity of the center point of the pig were measured using the automatic tracking software EthoVision for the duration of the test.
Three-chamber social recognition test
The three-chamber social recognition arena was constructed using metal grid paneling positioned over black rubber mats and is comprised of three chambers (Fig. 1E). The center chamber is the start/exit chamber made up of two panels attached to the north wall of the arena and two panels attached to the south wall of the arena, each 1.5 m in length. Both the north and south walls of the start/exit chamber contain doors that allow the pigs to enter/exit at alternating locations of arena. Two openings in the start/exit chamber lead into the two outer chambers. Each outer chamber contains a smaller social stimulus box (1.5 × 1.2 m) that can hold a single stimulus pig (Fig. 1F). The bars of the social stimulus box allowed nose contact between the test and stimulus pigs, but prevented direct social interactions that might lead to aggression. Stimulus pigs were of identical age, were from a separate litter, and had not seen or interacted with the test piglets previously. Stimulus pigs (n = 5) used for TBI pigs were different from stimulus pigs (n = 5) used for normal control pigs. All stimulus pigs were randomized such that each pig was an unfamiliar pig, familiar pig, and novel pig throughout all trials to reduce a possible novelty affect where one pig could potentially be more memorable than the others. In addition, in lieu of a social stimulus pig, a novel object could be securely attached to the front of either social stimulus box to measure object exploration. Five different objects were randomly assigned to TBI and to normal control pigs. No object bias for any objects used in this test were found (data not shown). White curtains were hung around the maze to reduce visual distractions during testing.
The three-chamber social recognition test was comprised of three separate trials: habituation, sociability trial, and social memory trial. In the habituation trial, piglets were placed into the start/exit chamber at either the north or south start locations and given 10 min to freely explore the arena. For the sociability trial, piglets were briefly removed from the arena while an unfamiliar stimulus piglet was placed into a social stimulus enclosure and a novel object was attached to the front of the opposing social stimulus box. The pigs were then placed back into the arena and allowed to explore the arena, the unfamiliar stimulus pig, and the novel object freely for 10 min.
For the social memory trial, the pigs were removed from the arena for an inter-phase interval of 10 min. The novel object was removed and a novel stimulus pig was brought into the arena and placed into a social stimulus box. The unfamiliar pig from the sociability trial remained in the arena and thus became a familiar pig for the social memory trial. The test piglets were then placed back into the arena for an additional 10 min and allowed to explore the arena, the familiar pig, and the novel pig for 10 min. The location of the unfamiliar pig, novel object, and familiar pig alternated between east and west chambers for the sociability and social memory trials between animals. The automatic tracking software EthoVision was used to measure the amount of time spent with the unfamiliar pig and novel object in the sociability trial and the amount of time spent exploring the novel pig and familiar pig in the social memory trial. Time spent exploring a stimulus pig in either the sociability or social memory trials was measured as the time at which the nose-point of the test pig was in the zone immediately surrounding the social stimulus box. The time spent exploring a novel object was measured as the time during which the nose-point of the test pig was in the zone surrounding the novel object. Any urination or defecation in the arena was wiped down with 70% ethanol between each trial as necessary to reduce olfactory bias. The arena was wiped down with 70% ethanol between each pig as well to reduce olfactory cues across pigs.
Gait analysis
The gait analysis track was constructed of metal grid paneling with an adjustable, straight chute approximately 6.7 m × 0.5 m with circular turn around pens at either end approximately 2.1 m in diameter, giving the track a “dog bone” shape. Pigs were subjected to gait assessment using a GAITFour electronic, pressure-sensitive mat (CIR Systems Inc., Franklin, NJ) that is 7.01 m in length and 0.85 m in width with an active area that is 6.10 m in length and 0.61 m in width. In this arrangement, the active area is a grid, 48 sensors wide by 480 sensors long, totaling 23,040 sensors. The GAITFour mat was placed down the length of the chute and covered with a protective no-slip, adhesive mat. After approximately 1 week of individual acclimation to the gait track, pigs were trained to travel back and forth across the track at a consistent pace using milk replacer as a reward. Two handlers, one in each of the holding pens, rewarded the pig if the animal crossed the entire length of the mat at a consistent two-beat trotting gait without stopping or breaking into a run (or three-beat gait) until five valid trials were collected.
The pressure mat recorded the footfalls of the pig as it traveled across the mat and recorded the following spatial-temporal and pressure gait biomechanical parameters: Cycle time: time elapsed between the first contacts of two consecutive hoof strikes of the same hoof (i.e., left front to left front); expressed in seconds Stance percent: percentage of time during which a hoof is in stance phase (hoof is in contact with the ground) during one stride cycle (stance time/cycle time). Hind reach: distance from the heel center of the hind hoof to the heel center of the previous front hoof on the same side (i.e., left hind to left front). The closer the hind hoof to front hoof, the more positive the value. The further the hind hoof from the front hoof, the more negative value; expressed in centimeters Total pressure index (TPI): the percent of weight distribution across all four limbs, where most quadrupeds carry 60% of their weight on their forelimbs and 40% of their weight on their hind limbs.
For each trial, a minimum of three gait cycles with less than 10% variability in velocity were selected, and all gait parameters were calculated automatically using the GAITFour software. For each pig, the trials collected at every time point were averaged and normalized to pre-TBI values to account for inter-pig variability by calculating the difference between post-TBI and pre-TBI values.
Histology
At 28 days post-TBI, pigs were sedated and euthanized via intravenous lethal pentobarbital injection. After euthanasia, a knife was used to separate the head from the body at the level of the atlanto-occipital articulation. The brain was removed from the skull using a hacksaw and stryker saw and then placed whole in 10% buffered formalin until immunohistochemical analysis. After fixation, gross brains images were acquired (Supplementary Fig. S1A, S1B), then brains were sectioned into 5 mm slices using a coronal pig-specific brain sectioner. Slices with both injured cortex and dorsal and ventral hippocampus at approximately the same level were routinely processed, embedded in formalin, and stained with antibodies specific for NeuN (Millipore; 1:500), glial fibrillary acidic protein (GFAP; Biogenex; 1:4000), Iba1 (Wako; 1:8000), and doublecortin (DCX; Abcam;1:2000). Heat induced epitope retrieval was performed for all antibodies using citrate buffer at pH 6 (DAKO). Detection was performed utilizing biotinylated antibodies (Vector Labs) and horseradish peroxidase labeled-streptavidin label (4plus Streptavidin HRP Label; Biocare) and 3,3-diaminobenzidine chromagen (DAKO). All sections were lightly counterstained with hematoxylin. Microscopic inspection and imaging were performed on a Nikon Eclipse TE300 inverted microscope utilizing SPOT Imaging v5.2 software (Sterling Heights, MI) at 5–7 consecutive fields along the perilesional border at 200 × , at the dorsal polymorphic layer of the dentate gyrus at 200 × , and at 5 consecutive fields at the subventricular zone (SVZ) at 20 × . For quantification, a region of interest (ROI) was drawn around the lesion border, the CA1/CA3 of the hippocampus, and the polymorphic layer of the dentate gyrus and NeuN+ cells were manually counted using the cell counter plugin on ImageJ 2.053 and expressed as cells/mm2. For semiquantitative analysis of GFAP and Iba1, the total area of immunoreactivity corresponding to increased optical density was measured by ImageJ 2.0 software at the perilesional area. Finally, a ROI was drawn around the SVZ and DCX+ neuroblasts were manually counted using ImageJ 2.0 and expressed as cells/mm2.
Statistical analysis
For the spatial T-maze test, open field test, and gait analysis, data analysis was conducted using the proc MIXED procedure as a two-way (treatment × day) repeated measures analysis of variance using Statistical Analysis software (SAS Version 9.1; Cary, NC). Post hoc comparisons using Pdiff were made to further examine significant effects. For the social recognition test and histological analysis, data analysis was conducted using a two-tailed t-test with SigmaPlot. Statistical significance was accepted at p < 0.05 and statistical trends at p < 0.10. Data are presented as mean ± standard error of the mean.
Results
TBI leads to spatial memory deficits that affect acquisition in spatial T-maze testing
Spatial T-maze testing was initiated 16 days post-TBI. On acquisition Day 1 (A1), both normal and TBI piglets were unfamiliar with the maze and thus exhibited a longer latency to choice indicative of a lengthier, less direct track to the reward bowl (Fig. 2A). However, latency to choice was significantly (p < 0.05) reduced by A2 for normal and TBI animals (Fig. 2A, 2B). Latency to choice continued to decrease throughout acquisition and remained unaffected by reward reversal for normal and TBI animals (Fig. 2B). For proportion trials correct, TBI piglets made significantly (p < 0.05) fewer correct reward choices than normal piglets from A1 through A4 (Fig. 2C). In addition, compared with the first day of testing on A1, normal piglets made significantly (p < 0.05) more correct choices by A2, while TBI piglets took 1 day longer and made significantly more correct choices on A3 (Fig. 2C). Both normal and TBI pigs showed a temporary significant (p < 0.05) decrease in correct choices on reversal day 1 (R1). Together, these data suggest that TBI leads to spatial memory deficits that affect acquisition of the reward location in a spatial T-maze.
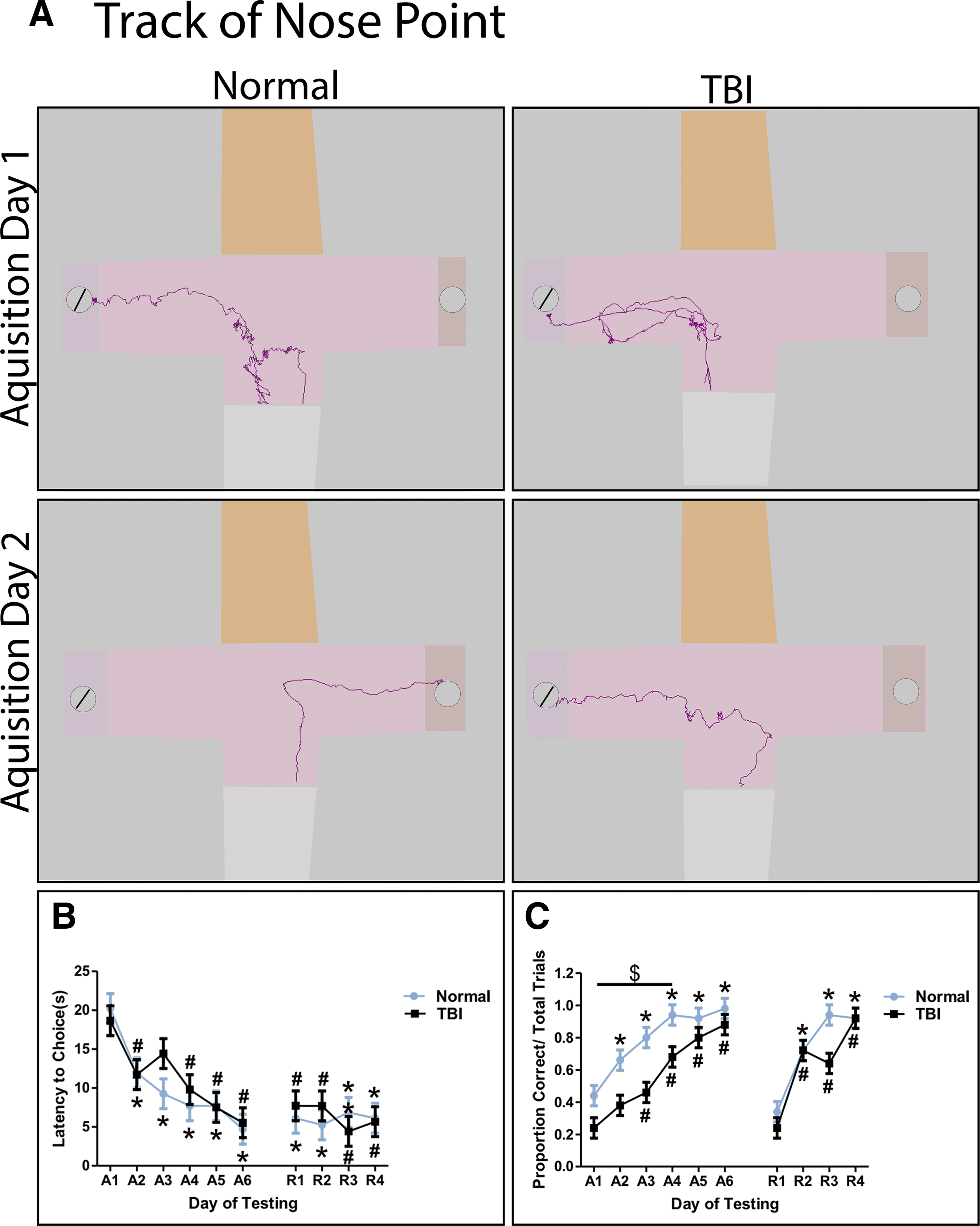
Traumatic brain injury (TBI) results in impairments in spatial memory formation assessed in a spatial T-maze test. Latency to choice and proportion trials correct was assessed in a spatial T-maze test during an acquisition phase (A1-A6) and a reversal phase (R1-R4). Representative nose point tracks of normal and TBI piglets on A2 show a more direct track to reward compared with A1 during T-maze testing. Open gray circles represent the correct reward, crossed circles represent the incorrect reward
TBI leads to a decrease in exploratory behaviors followed by an increase in hyperactivity in open field testing
Piglets were tested in an open field test 1 day, 7 days, 14 days, and 26 days post-TBI. TBI piglets traveled significantly (p < 0.05) less distance 1 day post-TBI compared with normal controls (Fig. 3A, 3B). Normal pigs showed the highest exploration and traveled the greatest distance on Day 1, and, as expected, became habituated to the testing arena (Fig. 3A). As a result, distance traveled generally decreased over time and was significantly (p < 0.05) reduced by Day 14 (Fig. 3B). However, for TBI pigs, distance traveled generally increased over time and was significantly (p < 0.05) increased by 26 days post-TBI (Fig. 4A, 4B). Similarly, TBI piglets moved at a significantly (p < 0.05) slower velocity 1 day post-TBI compared with normal controls (Fig. 3C). Normal pigs showed the highest interest in their environment and greatest velocity on Day 1 that generally decreased over time and was significantly (p < 0.05) reduced by Day 14 (Fig. 3C). However, for TBI pigs, velocity generally increased over time and was significantly (p < 0.05) greater on Days 14 and 26 post-TBI compared with 1 day post-TBI (Fig. 3C). Additionally, the velocity of TBI piglets was significantly (p < 0.05) higher than normal pigs 14 days post-TBI. These data suggest that TBI leads to an immediate decrease in exploratory behaviors that is followed by a period of hyperactivity in the weeks following TBI.
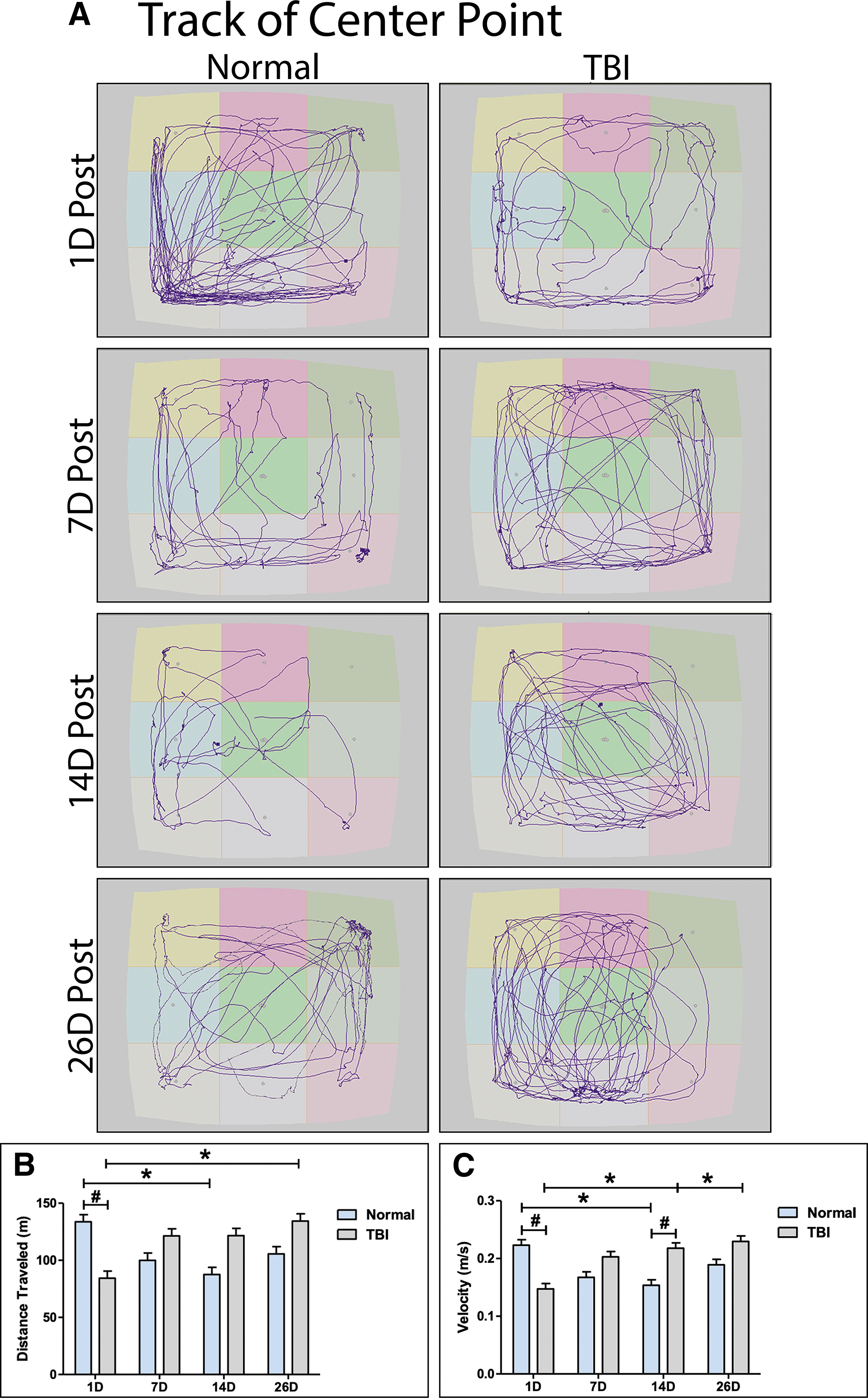
Traumatic brain injury (TBI) impairs normal exploratory behaviors assessed in an open field test. Representative center point tracks of normal and TBI pigs 1, 7, 14, and 26 days post-TBI
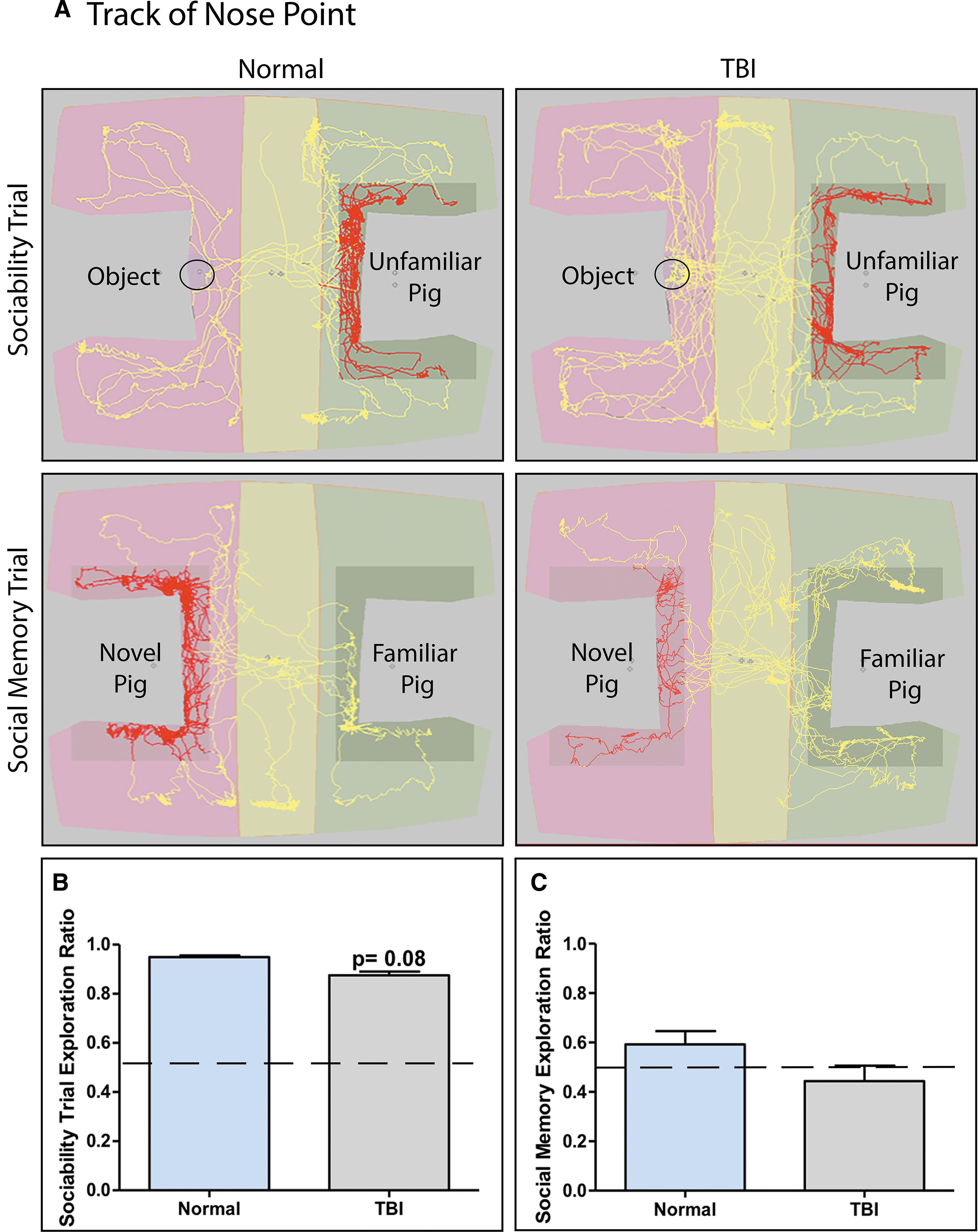
Traumatic brain injury (TBI) alters sociability and social memory behaviors assessed in a three-chamber social recognition test. Representative nose point tracks of sociability and social memory trials for normal and TBI pigs
TBI may disrupt normal sociability and social memory behaviors in a three-chamber social recognition test
Pigs were tested in a three-chamber social recognition test on Day 15 post-TBI. In sociability trials, both normal and TBI pigs exhibited a strong preference for an unfamiliar pig over a novel object. However, TBI piglets overall spent less time with the unfamiliar pig than the novel object compared with normal controls (0.875 ± 0.034 vs. 0.950 ± 0.014, respectively), but significance was only trending (p = 0.079; Fig. 4A, 4B). In social memory trials, normal pigs spent proportionally more time with the novel pig compared with the familiar pig, while TBI pigs spent proportionally less time with the novel pig compared with the familiar pig (0.592 ± 0.121 vs. 0.444 ± 0.140, respectively); however, there was no significant difference between treatment groups (Fig. 4A, 4C). These data suggest that TBI may potentially disrupt sociability and social memory behaviors, but no significant effects on these behaviors were detected.
TBI contributes to transient increases in cycle time and stance percent that alters normal gait function
Changes in the gait parameter cycle time was assessed by comparing each time-point to pre-TBI for the left front, right front, left hind, and right hind limbs. TBI pigs showed a significant (p < 0.05) increase in cycle time in all limbs 1 and 3 days post-TBI compared with pre-TBI and a significant increase in cycle time 1 day post-TBI compared with normal controls for all limbs (Fig. 5A-D). No change in cycle time was observed on Days 1 and 3 in normal control pigs compared with pre-TBI; however, from Days 7 through 28, a significant increase in cycle time was observed for all limbs that is indicative of normal changes associated with growth of the pig (Fig. 5A-D). TBI animals showed normal changes in cycle time at 7 and 14 days post-TBI for all limbs, but no significant change in cycle time was observed 28 days post-TBI, likely as a result of an overall preference for TBI pigs to move at a faster velocity than normal animals at this time-point (Fig. 5A-D).
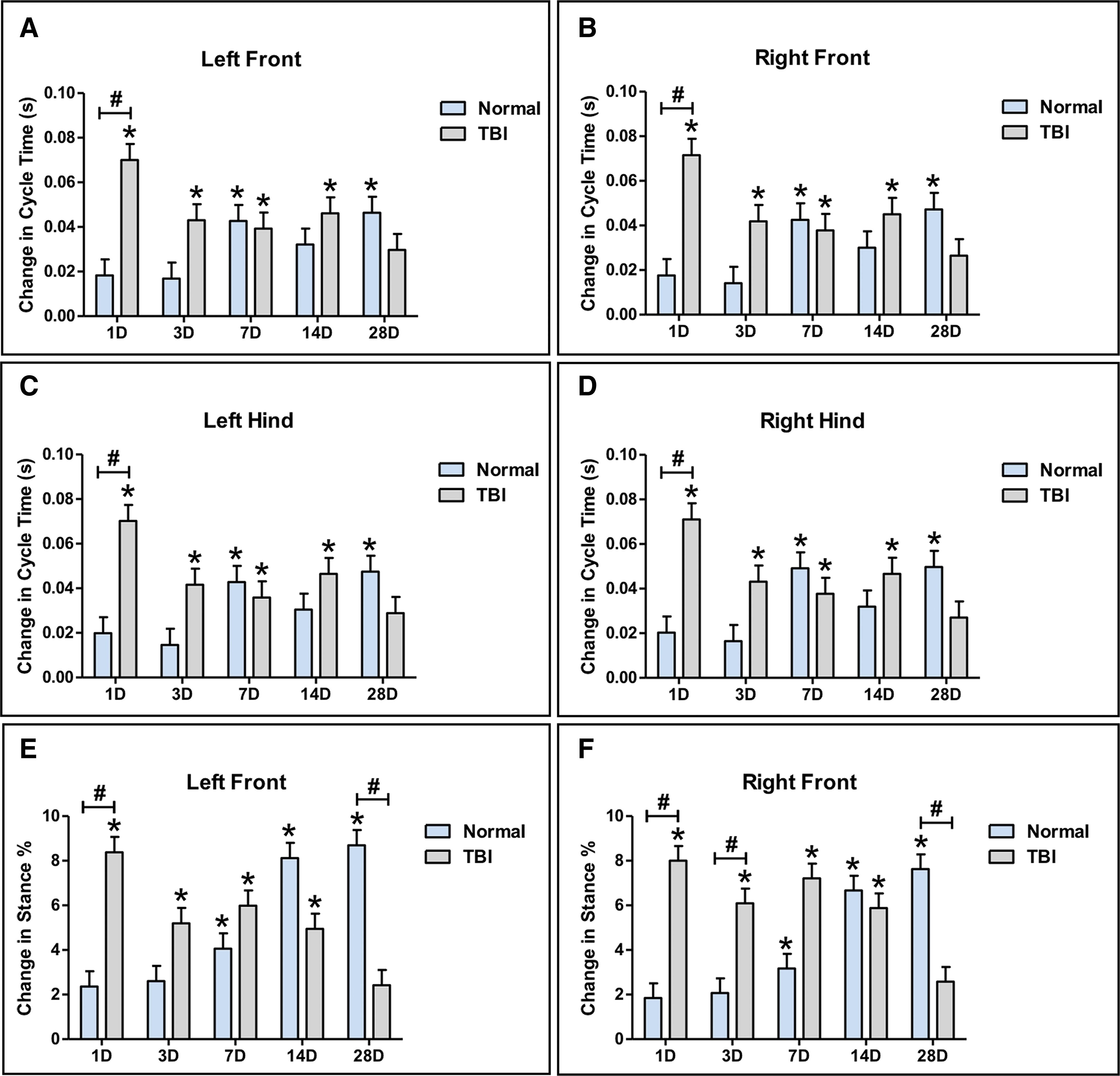
TBI leads to transient increases in the spatiotemporal gait parameters cycle time and stance percent. Significant increases in cycle time were observed 1 and 3 days post-TBI in the left front
Changes in the gait parameter stance percent was assessed by comparing each time-point to pre-TBI for the left and right front limbs. TBI pigs showed a significant (p < 0.05) increase in stance percent 1 and 3 days post-TBI compared with pre-TBI for the left and right front limbs (Fig. 5E, 5F). In addition, a significant (p < 0.05) increase in stance percent was observed 1 day post-TBI for the left front limb and 1 and 3 days post-TBI compared with normal controls for the right front limb (Fig. 5E, 5F). Similar to cycle time, no change in stance percent was observed on Days 1 and 3 in normal control pigs compared with pre-TBI. However, from Days 7 through 28, a significant (p < 0.05) increase in stance percent was observed for the left and right front limbs that is indicative of normal changes associated with growth of the pig (Fig. 5E, 5F). TBI animals showed normal changes in stance percent on 7 and 14 days post-TBI for all limbs, but no significant change in stance percent was observed 28 days post-TBI, likely as a result of an overall faster velocity than normal animals at this time-point (Fig. 5E, 5F). These data suggest that TBI contributes to transient changes in cycle time and stance percent that alter normal gait.
TBI leads to changes in hind reach and total pressure index that disrupt normal gait function
Changes in the gait parameter hind reach, which provides a measure of pig stability and balance, was assessed by comparing each time-point to pre-TBI for the left and right hind limbs. TBI animals showed a significant (p < 0.05) increase in hind reach 1 and 3 days post-TBI compared with pre-TBI for the left and right hind limbs (Fig. 6A, 6B). In addition, a significant (p < 0.05) increase in hind reach was observed 1 day post-TBI for the left hind limb and 1 and 3 days post-TBI compared with normal controls for the right hind limb (Fig. 6A, 6B). No change in hind reach was observed on Days 1, 3, and 7 in normal control pigs compared with pre-TBI. However, from Days 14 through 28, a significant decrease in hind reach was observed for the left and right hind limbs that can be attributed to growth of the pig (Fig. 6A, 6B). TBI animals show normal changes in hind reach 7, 14, and 28 days post-TBI for the left hind limb, but at 14 days post-TBI, hind reach was significantly reduced for the right hind limb compared with normal control pigs. Changes in right hind reach returned to normal by 28 days post-TBI (Fig. 6A, 6B).
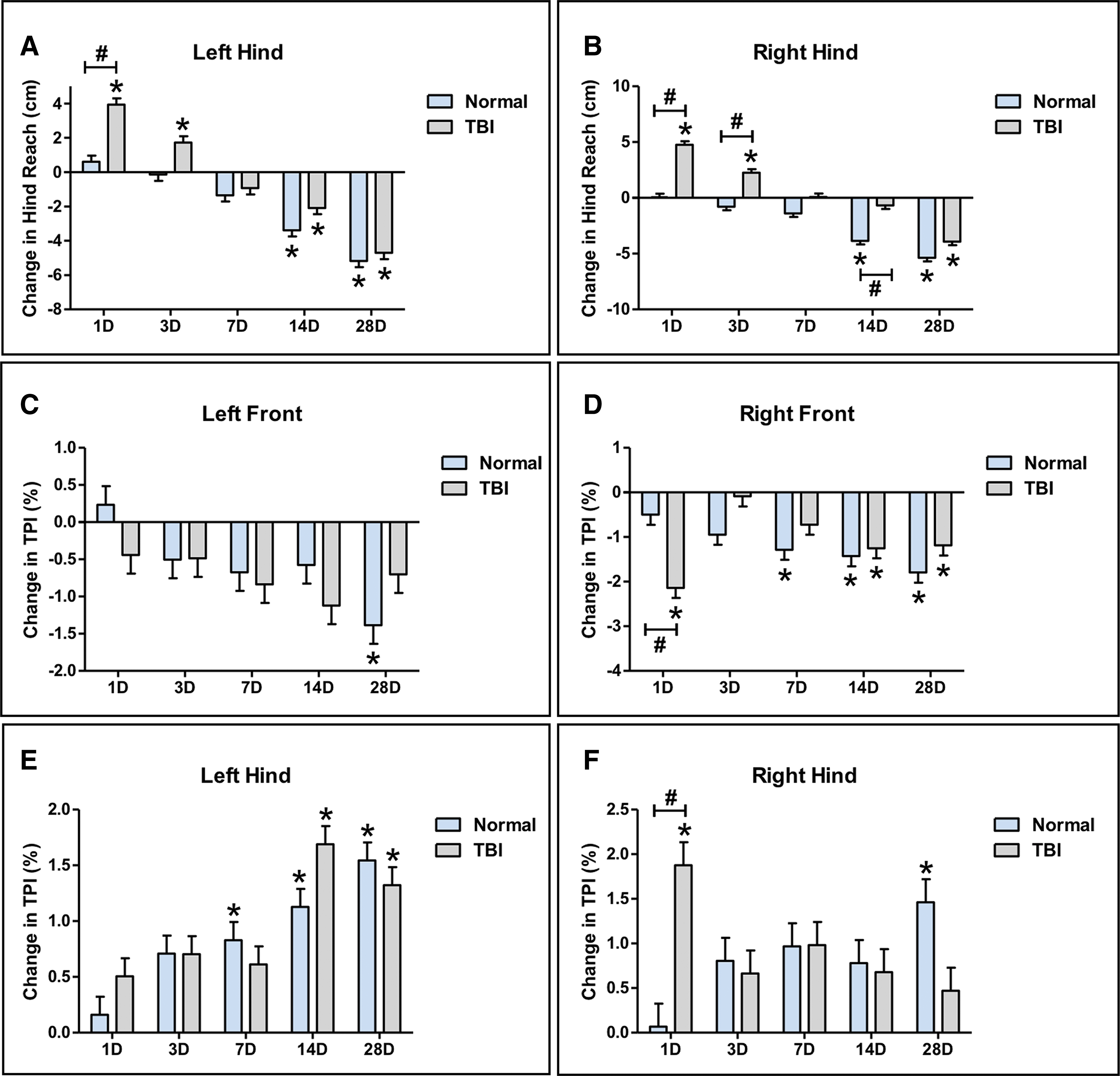
Traumatic brain injury (TBI) leads to transient changes in the gait parameters hind reach and TPI. Significant increases in hind reach were observed at 1 and 3 days post-TBI in the left hind
Changes in the gait parameter TPI was assessed by comparing each time-point to pre-TBI for the left front, right front, left hind, and right hind limbs. A significant (p < 0.05) decrease in TPI in the right (contralateral) front and a corresponding significant (p < 0.05) increase in TPI in the right hind was observed 1 day post-TBI compared with pre-TBI, and no change in TPI was observed in the left front or hind limbs (Fig. 6C-F). Although significant (p < 0.05) changes in TPI are observed in the right front limb starting at 7 days but not in the left limb until 28 days in normal pigs, overall, TPI appears to decrease in both front limbs over time as the animal ages (Fig. 6C-F). Further, significant (p < 0.05) changes in TPI are observed in the left hind limb starting at 7 days but not in the right hind limb until 28 days in normal pigs, thus overall, TPI appears to increase in both hind limbs over time (Fig. 6C-F). These data suggest that hind reach on the contralateral right side may be affected for up for 14 days post-TBI but that changes in TPI are transient and affect weight carriage only immediately after TBI.
TBI leads to changes in neurons, glia, and neuroblast distribution
Parenchymal tissues were assessed for cellular changes at the perilesional site, hippocampus, and SVZ 28 days after TBI. A significant (p < 0.05) loss of NeuN+ neurons was detected at the perilesional site for TBI pigs compared with normal control pigs (51.055 ± 13.129 cells/mm2 vs. 94.907 ± 4.272 cells/mm2, respectively; Fig. 7A-C). In addition, a significant (p < 0.05) loss of NeuN+ neurons were observed at the polymorphic layer of the dorsal dentate gyrus in TBI pigs compared with normal control pigs (44.369 ± 2.154 cells/mm2 vs. 52.901 ± 1.943 cells/mm2, respectively; Fig. 7D-F). No change in NeuN+ neurons was observed at the CA1/CA3 of the hippocampus (data not shown). TBI pigs also showed a significant (p < 0.05) increase in Iba1+ microglia reactivity at the perilesional site relative to normal control pigs (25.211 ± 3.218% positive area vs. 16.353 ± 1.306% positive area, respectively; Fig. 7G-I). In an assessment of reactive astrocyte activity, a significant (p < 0.05) increase in the area of GFAP+ astrocyte at the perilesional site in TBI pigs was observed relative normal control pigs (36.217 ± 5.736% positive area vs. 17.880 ± 2.279% positive area, respectively; Fig. 7J-L). Representative images of Iba1 and GFAP immunoreactivity were acquired at higher magnification that showed characteristic changes associated with microglia activation (Supplementary Fig. S2A, S2B) and astrogliosis/astrocytosis (Supplementary Fig. S2C, S2D). Finally, significant (p < 0.05) recruitment in DCX+ neuroblasts was observed at the SVZ in TBI pigs compared with normal control pigs (2.136 ± 0.306 cells/mm2 vs. 0.937 ± 0.239 cells/mm2, respectively; Fig. 7M-O). Taken together, these data suggest that TBI contributes a number of cellular changes at the perilesional site as well as at more remote zones such as the hippocampus and SVZ.
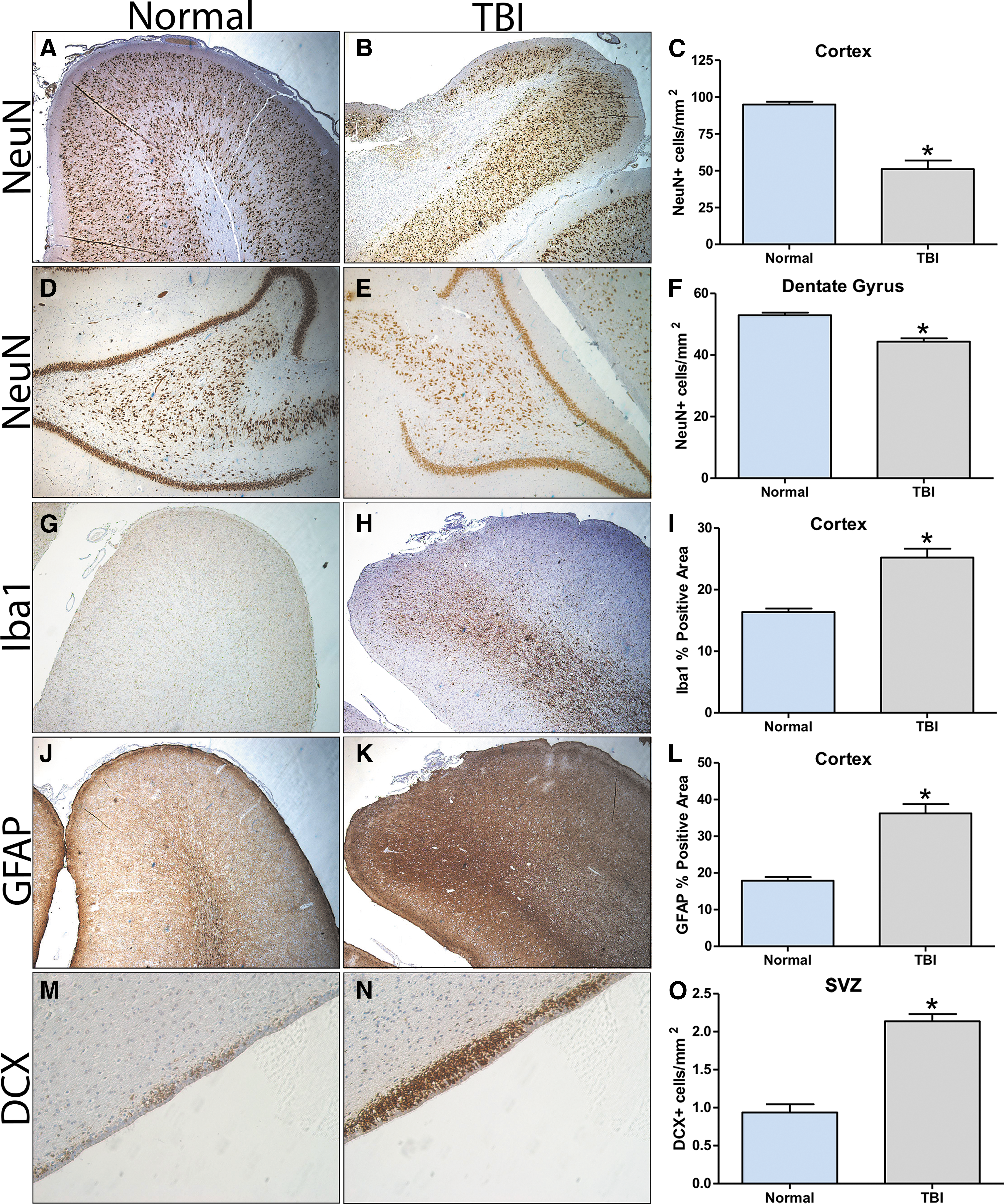
Traumatic brain injury (TBI) results in cellular changes at the perilesional site, dorsal hippocampus, and subventricular zone (SVZ). NeuN staining at the cortex in normal
Discussion
In the present study, we have shown that experimental TBI using CCI in a piglet model induces cognitive, motor, and histopathological deficits pathologically consistent with features found in human patients. TBI piglets exhibited significant spatial memory deficits and showed reduced exploratory behaviors immediately after TBI that was followed by a period of hyperactivity that persisted over time in spatial T-maze and open field testing. Changes in normal sociability and social memory behaviors was observed in TBI pigs in a three-chamber social recognition test. In addition, transient gait impairments in cycle time, stance percent, hind reach, and TPI were observed between 1–3 days post-TBI. Histological analysis revealed loss of neurons in the cortex and hippocampus, an increase in microglia activation and astrogliosis/astrocytosis, and stimulation of endogenous neuroblast populations at the SVZ. To our knowledge, this is the first report that uses a piglet CCI model to quantitatively measure histopathological changes and functional deficits using a comprehensive battery of behavior tests and gait analysis.
Children who sustain a severe TBI often show evidence of spatial learning and cognitive mapping deficits initially after TBI and can remain impaired for many years. 54,55 The spatial T-maze test has been well validated as a means to measure early life insults on cognition, specifically hippocampal-based spatial memory formation in pigs. 52,56 –59 This is the first time this test has been performed in a piglet CCI model. Previous studies utilized a traditional T-maze test in a diffuse axonal injury (DAI) piglet model; however, differences in correct reward choice and latency to choice between control and TBI animals did not reach significance. 49,60,61 In this study, the spatial T-maze test was performed 16 days post-TBI to avoid potential confounds with motor function deficits in the first days after injury. 18 All pigs acquired the “rules” of this task exceedingly fast with both normal and TBI pigs showing similar significant improvements in latency to choice as soon as A2. Motivation for seeking the reward was high given that milk replacer had been used previously as a reward for gait analysis training and the animals were feed restricted the night prior to testing.
However, TBI pigs showed significant impairments in choosing the correct reward location during the initial days of acquisition. The underlying principle of this test assesses the pigs' ability to use a spatial memory-based, allocentric mechanism to learn the location of a fixed reward, despite alternating the starting position between trials, by using extra-maze cues. 52,62,63 Allocentric navigation is hippocampal-based and is characterized by the ability to navigate using distal cues. This is in contrast to egocentric navigation which is striatal-based and is characterized by the ability to navigate using internal cues. 64 This type of navigation is employed in standard T-maze tests without alternating starting positions. Taken together, we show that, compared with normal controls, TBI impedes allocentric navigation and spatial memory formation in a spatial T-maze test.
Evaluation of the effect of TBI on open field testing revealed an initial reduction in exploratory behaviors 1 day post-TBI, followed by marked hyperactive exploration in the weeks after TBI. A decrease in exploratory behaviors immediately after TBI has been reported in other experimental animal models. 65 –67 Friess and colleagues showed that piglets with DAI spent significantly less time sniffing the walls of the arena during open field testing compared with sham pigs at 1 day post-TBI. 68 Sullivan and colleagues found that piglets with DAI were significantly more stationary at 1 day post-TBI compared with pre-TBI, which supports the results of this study that TBI leads to reduced exploratory behaviors immediately following TBI. 49,69 These changes may be attributed to anxiety or depressive symptoms, or a loss in exploratory interest. 21,70
In this study, open field behaviors were noted to significantly change over time in both TBI and control animals. Characteristic hyperactivity behaviors, increased velocity and distance traveled, were elevated starting at 7 days post-TBI and reached significance by 14 and 26 days post-TBI, respectively. This is in contrast to normal animals who showed a marked decrease in distance traveled and velocity over time, indicative of a habituation effect due to repeated testing. 71 However, Friess and colleagues observed no changes in open field behaviors across multiple timepoints at 1, 4, 6, 8, and 11 days post-injury for injured or sham animals. 68 These differences may be attributed to the use of a novel object during open field testing, or differences in the type and severity of TBI and testing time-points.
Increased hyperactive exploration of the open field arena is frequently reported in rodent CCI studies at the parietal cortex. 19,72 –75 Although the precise mechanism of TBI contribution to a hyperactive phenotype has not yet been fully elucidated, a study by Li and colleagues suggested that deficits in spatial and working memory reflect impairments in habituation and the ability to form and retrieve spatial maps, leading to hyperactivity. 76 Further, the parietal cortex and dorsal hippocampus, both regions affected in this study, have been shown to be part of a complex control system that modulates and suppresses a basally hyperactive locomotor tone. 77 Disruption to this system by TBI would therefore lead to increased locomotor activity. Clinically, human patients often display depressive symptoms shortly after sustaining a TBI that may coexist with anxiety disorders. 78,79 Children who sustain a TBI at a young age are three times more likely to develop attention-deficit hyperactivity disorder than their peers. 80 Behavioral problems and disorders that often accompany pediatric TBI continue to be both prevalent and long-term problems in affected individuals, highlighting the need for additional studies that further define TBI-induced behavioral changes. 81
Children affected by moderate-to-severe TBI are also at risk for social impairment. 82 Social participation, or the intensity of participation in social activities, has been found to be vulnerable to TBI. 83 In addition, TBI at a young age may lead to impairments in recognition memory such as facial recognition memory. 84 Evidence of long-term social dysfunction after pediatric TBI has garnered recent interest in incorporating social behavior tests in experimental animal models of brain injury. 85 –87 Although the three-chamber social recognition test has been performed previously in pigs, to our knowledge this is the first time this test has been performed in a piglet CCI model. 88 In this study, we observed changes in normal sociability behaviors. TBI pigs showed a preference for an unfamiliar pig over a novel object, but compared with normal control pigs, social preference for the unfamiliar pig was diminished, although significance was not reached. TBI pigs on average spent more time with the familiar pig than the novel pig, suggesting social memory impairments; however, results were not significant likely as a result of high variability between pigs. Social processing is highly complex, involving numerous brain structures that together help shape social behavior. Therefore, additional studies are needed to study TBI effects on sociability and social memory behaviors. 89
Limited studies have investigated the relationship between cognitive deficits and hippocampal histopathology in pigs after CCI. 50 Here, we report a loss of NeuN+ neurons in the dorsal polymorphic layer of the dentate gyrus. The polymorphic layer of the dentate gyrus is comprised of a number of different neuronal cell types that give rise to ipsilateral associational projections that innervate the molecular layer, as well as commissural projections that innervate the contralateral polymorphic layer. 90 Disruption of neurons within the polymorphic layer can result in aberrant hippocampal function. 91 Recent evidence suggests that there may be a gradient of function in the hippocampal long axis. 44 The ventral (or anterior hippocampus in humans) has been implicated in episodic-memory formation and emotional processes and the dorsal (or posterior hippocampus in humans) has been implicated in spatial memory formation. 59,92 –94 Insults to the dorsal hippocampus as a result of TBI, as we have demonstrated in this study, have been found to result in spatial memory deficits that are dependent on injury severity. 95
Children who sustain a TBI at a young age are also at risk for acquiring motor function deficits, particularly those relating to balance and gait. 96 Analysis of spatiotemporal gait parameters in children after TBI have shown an increase in step cycle time and a corresponding decrease in gait velocity. 12,14 Previous studies in pig TBI models provided gross measurements of motor function after injury using either a balance beam test, inclined balance beam test, or a hurdle cross test. 49,50,68 Baker and colleagues performed quantitative gait analysis using high speed cameras and manual calculation of spatiotemporal gait parameters. 48 In this study, a GAITFour pressure mat automatically tracked and measured both spatiotemporal parameters and changes in pressure and provided a more sensitive and accurate measurement of changes in motor function after injury. Pigs displayed a similar increase in cycle time in all limbs 1 and 3 days post-TBI compared with normal control pigs. A corresponding increase in stance percent was observed.
Reports of changes in stance percent are varied in children, with some studies reporting no change in overall stance percent and others that show an increase in stance percent in the unaffected limb (ipsilateral to TBI) but no change in stance percent in the affected limb. 12,97 Here, stance percent is increased overall in both front limbs, but compared with normal controls, stance percent in the right (contralateral) limb was more prominently altered than the left (ipsilateral) limb. Pigs are quadrupeds as opposed to bipeds, therefore variations in gait may account for these differences. Duberstein and colleagues showed that, contrary to stroke patients, stroked pigs exhibited increased stance percent of the affected limb and suggested this may be due to either lack of propulsive forces of the affected limb or an effort to maintain ground contact of the affected limb in order to better stabilize weight distribution on the affected side. 98
TBI pigs also showed an increase in hind reach for the left and right hind limbs 1 and 3 days post-TBI coupled with a decreased in TPI in the right front limb and a corresponding increase in TPI in the right hind limb 1 day post-TBI. These results suggest that TBI pigs brought their hind limbs more underneath of their body, tracking up closer to their front limbs, and in conjunction, shifted their weight from the front to the hind limbs on the affected side, likely in an effort to better stabilize their gait and improve balance. Recently, rodent TBI studies utilizing a Catwalk imaging method have found similar responses in stance percent, relative paw placement (hind reach), and intensity (TPI) acutely but not long-term. 99 –101 Here, we report only transient gait deficits that persist for only a few days after TBI, likely a result of TBI location and severity. TBI was induced at the level of the parietal lobe, just caudal to the motor cortex. Due to the close proximity of the impact site, the motor cortex is likely to be affected by secondary injury mechanisms and would explain the transient motor function impairments.
A small number of pig CCI studies have both qualitatively and quantitatively analyzed immunohistological changes after TBI. Duhaime and colleagues described cell loss at the lesion site and gliosis adjacent to the lesion. 36 Manley and colleagues showed loss of MAP-2+ and NeuN+ neurons and an increase in Fluoro-Jade B+ degenerating neurons near the injury site. 102 The Margulies group has performed several studies utilizing behavior and motor function tests in a piglet TBI model of closed head rotational injury, and histological analysis revealed increased β-amyloid precursor protein staining. 49,61,69 Here, we show a loss of NeuN+ neurons and an increase in Iba1+ and GFAP+ area at the perilesional site. These results are consistent with reports from other experimental CCI studies. 17,67,103,104 Cell loss following both experimental and clinical TBI has been correlated with poor prognosis and cognitive deficits. 105 In response to injury, characteristic features of astrogliosis/astrocytosis were observed including the upregulation of GFAP, cellular hypertrophy, increased proliferation, and overlapping of neighboring astrocytes processes. Significant astrogliosis/astrocytosis can lead to the formation of dense, compact glial barriers, called glial scars, that can act as a neuroprotectant against inflammatory cells but can also inhibit axonal regeneration, contributing to functional impairments. 106 Similarly, in response to injury, characteristic features of activated microglia were observed such as increased expression of Iba1, change from a ramified to an ameboid morphology, and increased proliferation and migration to the site of injury. 107 In addition, activated astrocytes and microglia following TBI can secrete pro-inflammatory cytokines that can alter synaptic plasticity mechanisms, contributing to functional deficits like memory loss or motor function deficits. 108,109 Lastly, we observed a significant increase in DCX+ neuroblasts at the SVZ following TBI. Costine and colleagues was the first to identify the neurogenesis of DCX+ neuroblasts at the SVZ in a TBI piglet model and note that the long-term outcomes of alterations in normal neuroblast migration are unknown, but that they could potentially play role in repair or remodeling mechanisms in the developing brain. 46
Conclusions
The piglet brain is more similar to the human brain in terms of size, structure, white matter composition, and injury responses, compared to traditional rodent models which suggests it may be a more advantageous pre-clinical animal model. We have developed a clinically relevant piglet CCI TBI model that demonstrates functional deficits with corresponding histological changes. Neuronal loss at the hippocampus and cortex and presence of astrogliosis/astrocytosis and microglia activation at the perilesional site suggest a possible mechanism for the progression of cognitive, behavioral, and motor function impairments after injury. This model can be utilized in future studies to help elucidate the effects of early life insult on development in the young gyrencephalic brain and to target and test novel therapeutics that lead to functional improvements.
Footnotes
Acknowledgments
We would like to thank Madelaine Wendzik, Shelley Tau, Kayla Hargrove, Jaime Certusi, Sarah Lewis, Olivia Fuller, Caroline Coleman, Kathryn Sellman, Alex Ross, Natalie Bishop, Julianne Gillis, and Andrea Kuehndorf for their significant and integral involvement in all pig work. We would also like to thank Rick Utley and Kelly Parham at the UGA Swine Unit for their pig expertise and for providing us with piglets. Lastly, we would like to thank Randy Nation for his help in designing and building our behavior testing arenas. This work was supported by The University of Georgia Research Foundation.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.