
Abstract
Targeting mitochondrial ion homeostasis using Kaempferol, a mitochondrial Ca2+ uniporter channel activator, improves energy metabolism and behavior soon after a traumatic brain injury (TBI) in developing rats. Because of broad TBI pathophysiology and brain mitochondrial heterogeneity, Kaempferol-mediated early-stage behavioral and brain metabolic benefits may accrue from diverse sources within the brain. We hypothesized that Kaempferol influences TBI outcome by differentially impacting the neural, vascular, and synaptic/axonal compartments. After TBI at early development (P31), functional magnetic resonance imaging and diffusion tensor imaging (DTI) were applied to determine imaging outcomes at adolescence (2 months post-injury). Vehicle and Kaempferol treatments were made at 1, 24, and 48 h post-TBI, and their effects were assessed at adolescence. A significant increase in neural connectivity was observed after Kaempferol treatment as assessed by the spatial extent and strength of the somatosensory cortical and hippocampal resting-state functional connectivity (RSFC) networks. However, no significant RSFC changes were observed in the thalamus. DTI measures of fractional anisotropy (FA) and apparent diffusion coefficient, representing synaptic/axonal and microstructural integrity, showed significant improvements after Kaempferol treatment, with highest changes in the frontal and parietal cortices and hippocampus. Kaempferol treatment also increased corpus callosal FA, indicating measurable improvement in the interhemispheric structural connectivity. TBI prognosis was significantly altered at adolescence by early Kaempferol treatment, with improved neural connectivity, neurovascular coupling, and parenchymal microstructure in select brain regions. However, Kaempferol failed to improve vasomotive function across the whole brain, as measured by cerebrovascular reactivity. The differential effects of Kaempferol treatment on various brain functional compartments support diverse cellular-level mitochondrial functional outcomes in vivo.
Introduction
A large interdependence exists between neural circuit activity, the cerebrovascular supply supporting circuit function, and synaptic/axonal microstructure, which act in concert to determine systemic sensorimotor and cognitive outcomes. Traumatic brain injury (TBI) pathology is broad, affecting multiple cellular types and hence different neurophysiological variables. 1 TBI leads to an energy metabolic decline in pediatric patients and developmental pre-clinical models of injury, 2 –7 consequently affecting downstream function in various cellular compartments. 1 Hence, mitochondria, the subcellular organelles responsible for oxidative energy metabolism and enriched in diverse cell types of the brain, are effective targets for TBI treatments. It can be hypothesized that treatments targeting mitochondria can promote diverse cell-type–specific downstream repair mechanisms in neurons, glia, vascular smooth muscle, and endothelium, which can synergistically improve neurophysiological outcomes. Given that energy metabolism is upstream to various cellular processes during recovery from TBI, treatments maintaining mitochondrial ion homeostasis, and hence their energy metabolic capacity, may differentially impact injury prognosis depending on the mitochondrial properties of each brain compartment.
Preservation of energy metabolism in the developing brain is not only crucial for efficient post-TBI repair by downstream energy-dependent mechanisms, but also to sustain normal ongoing developmental changes. Given that cerebral vasculature, neural, and axonal/synaptic microstructures dynamically change during development, 8 –12 TBI in children are more devastating because of impediments of ongoing normal development. Given the temporally variable characteristics of TBI pathology, 1 pre-clinical experiments reporting on multiple biomarkers of brain function during development, preferably in a simultaneous manner and translatable to humans, are necessary. Addressing this gap, our previous study utilized magnetic resonance imaging (MRI)-based functional MRI (fMRI) and diffusion tensor imaging (DTI) modalities to simultaneously determine brain structural and functional changes at adolescence after TBI using the development rat model. 13 The multi-modal MRI biomarkers obtained at the adolescent stage strongly correlated with metabolic decline, reduced neural viability, and behavioral debilitation observed during the early acute/subacute stage (0–72 h) after TBI. 2
Acutely targeting mitochondrial ion homeostasis using Kaempferol, a pharmacological enhancer of mitochondrial Ca2+ uniporter channel (mCU) activity and hence mitochondrial Ca2+ cycling, 14,15 mitigated TBI-induced early energy metabolic decline and improved behavioral performance. 16 The early-window Kaempferol treatment regimen (1 mg/kg, intraperitoneal [i.p.]) at 1, 24, and 48 h post-TBI also improved prognosis at adolescence revealed by better stimulus-induced neurovascular coupling responses and sensorimotor behavior. 17
Using a translatable experimental design and clinically relevant imaging markers, the present study tested the hypothesis that Kaempferol-mediated effects on the brain are diverse, through its variable effects on mitochondria in different cellular types. Rats (age, P31) underwent a fluid percussion TBI in mild-to-moderate intensity and were randomly assigned to vehicle or Kaempferol (1 mg/kg) groups, where treatments were made at 1, 24, and 48 h after injury. Using fMRI and DTI measures at adolescence (2 months after injury), various system-level biomarkers informing on the neural, vascular, and synaptic/axonal compartments were determined to assess TBI prognosis and the effects of Kaempferol treatment.
Methods
Animals and treatment
Male Sprague–Dawley rats (n = 16; 23–24 days of age; weighing 60–80 g) were procured from Charles River Laboratories (Wilmington, MA) and housed in pairs under controlled conditions. All experimental on animals were carried out in accordance with the protocol approved by a local Institutional Animal Care and Use Committee of Rutgers Biomedical and Health Sciences–New Jersey Medical School (Newark, NJ) and Yale University School of Medicine (New Haven, CT). Kaempferol used in the current study was of commercial grade (Sigma-Aldrich, St. Louis, MO) and freshly prepared into working solution using sterile saline. Routes of administration for vehicle (10% dimethyl sulfoxide in saline) and Kaempferol were through i.p. injection.
Lateral fluid percussion injury
Rats (age, P31) were subjected to lateral fluid percussion injury (FPI) as previously described. 18,19 The developmental rat age at P31 approximately corresponds to between 2 and 4 years of human age, 20 The FPI method produced diffuse moderate TBI, very similar to that observed in humans. Briefly, animals were anesthetized with ketamine (80 mg/kg, i.p.)/xylazine (10 mg/kg, i.p.) and positioned on a stereotaxic frame after confirming surgical plane anesthesia. A 3-mm craniotomy was performed on the left side of the skull −5 mm posterior to the bregma and 3 mm lateral to the sagittal suture while retaining the dura. A Luer–Lock syringe hub was attached surrounding the exposed dura using cyanoacrylate adhesive. Twenty-four hours after the Luer–Lock placement, TBI was induced by attaching the Luer–Lock hub of each isoflurane-anesthetized animal to the FPI device (Virginia Commonwealth University, Richmond, VA). A pendulum drop delivered a brief 20-ms impact on the intact dura. Impact pressure was measured by an extracranial transducer and controlled between 1.8 and 2.0 atmospheres.
All surgical and experimental conditions in the current treatment groups were kept similar to our previous sham and untreated TBI study for consistency and which has been published recently. 13 Immediate neurological parameter observation after injury did not lead to any seizure-like behavior in all injured animals. Vehicle- and Kaempferol-treated TBI animals were monitored within their cage environment on a daily basis. Mortality was observed in 2 vehicle-treated animals and 1 Kaempferol-treated animal within 3 days post-injury. Surviving animal groups, that is, TBI + vehicle (n = 5) and TBI + Kaempferol treatment (n = 8) underwent fMRI and DTI measurements for the present study.
Magnetic resonance imaging
MRI experiments were performed at adolescence (2 months after TBI), translating to approximately 12–14 years of human age. 20,21 Animals were anesthetized using i.p. injection of urethane (1.3 mg/kg body weight). A tracheostomy was performed on animals to administer a mixture of O2 and N2O (30%/70%) through a breathing tube. Animals were then placed into a custom-built frame and fixed to ear bars to minimize motion and produce consistent spatial positioning within the radiofrequency coil. Body temperature was monitored throughout the procedure using a rectal probe and maintained within 35–37°C using a warm-water–circulated pad. Chronology of anatomical MRI, fMRI, and diffusion MRI measures were randomly varied across animal subjects, except the hypercapnic (CO2) stimulation experiments, which were always performed last.
Anatomical magnetic resonance imaging
MRI scanning was performed on a modified 9.4 Tesla system with a Bruker spectrometer and custom-built 1 H ellipsoidal surface coil (5 × 3 cm; Bruker, Billerica, MA). MR images were acquired over 12 contiguous coronal slices (thickness = 1 mm), covering the parenchyma between the olfactory bulb and cerebellum, with an in-plane field of view of 3.2 × 2.4 cm. Anatomical reference images (TR/TE = 4000/30 ms, 2 averages) were acquired in a 128 × 96 matrix, using a rapid imaging with refocused echoes sequence providing an in-plane resolution of 250 × 250 μm. Additionally, a fast three-dimensional (3D) anatomical scan (TR/TE = 50/5.6 ms, flip angle = 20 degrees, 2 averages) was acquired with an isotropic resolution of 250 μm, for image registration purposes.
Diffusion magnetic resonance imaging
Diffusion-sensitive images for DTI (TR/TE = 4000/20 ms, 4 averages) was acquired as a four-segment echo planar imaging (EPI) in a 64 × 48 matrix (in-plane resolution = 500 × 500 μm), with 5 A0 images, 15 diffusion directions, and a b-value of 1000 s/mm2.
Resting state and cerebrovascular reactivity functional magnetic resonance imaging
Resting-state and cerebrovascular reactivity fMRI were performed using the gradient echo (GE) EPI sequence (TR/TE = 1000/15 ms, preceded by eight dummy scans) using the same geometry as diffusion scans (64 × 48 matrix, 500 × 500 μm in-plane resolution), with the GE-EPI sequence producing image contrast sensitive to blood oxygenation changes (blood oxygen level dependent; BOLD). 22 The resting-state fMRI paradigm consisted of a 5-min scan (300 repetitions), acquired in four experimental trials per animal. Cerebrovascular reactivity fMRI scans were obtained using similar GE-EPI parameters as the resting-state fMRI, which lasted for 12 min (720 repetitions). A 10% carbon dioxide gas (CO2) was added to the breathing gas mixture of O2 and N2O (30%/70%) between minutes 3 and 6 of the acquisition and repeated twice per animal.
Statistical analysis
All EPI images were linearly registered to the subject's native anatomical space before further processing. BioImage Suite software (
For resting-state functional connectivity (RSFC) analysis, preprocessed EPIs were band-pass filtered (0.01–0.15 Hz). Six seed voxels were chosen from each region of interest (ROI) from the right hemisphere (contralateral to the injury). ROIs were somatosensory cortex, hippocampus, and thalamic regions, ensuring that the seed voxels randomly identified always lie within these respective ROIs identified using the rat brain atlas. 25 Signal time series from each selected seed voxel was cross-correlated with the entire brain, producing six cross-correlation maps for each experimental trial. As demonstrated in our earlier studies in normal 26 and TBI animals, 13 the average RSFC map obtained from the six different cross-correlation maps represented a more accurate topology of the RSFC network. In a similar manner, the six correlation maps obtained using six random seed voxels within each respective ROI (somatosensory cortical, hippocampus, and thalamus) were obtained for each resting-state scan leading to 18 correlation maps for three experimental trials performed on each animal. Subsequently, all correlation maps were averaged across trials and animals in each group. Fisher's z-transformation was used to convert the correlation coefficients to z-scores before averaging. After averaging, an inverse transform was subsequently used to obtain voxel-wise average correlation coefficient values in the group RSFC maps.
For group statistical comparisons, 3D anatomical images were used to generate a non-linear transform from each animal subject's native space to a common space, consisting of an averaged brain from 8 sham animals obtained from our earlier analysis of sham animals. 13 Given that a small ipsilateral distortion occurred along the skull and parenchymal area (injury epicenter) where a fluid percussion Luer–Lock existed to enable the fluid percussion TBI, a non-linear registration was used for the final statistical parameters. Registered anatomy from each animal subject was visually inspected for alignment by overlaying the registered images on the average-sham anatomical MRI template. Subsequently, the respective transforms were applied to all parametric maps of FA, apparent diffusion coefficient (ADC), cerebrovascular reactivity (CVR), and RSFC networks obtained from all TBI animals (both vehicle- and Kaempferol-treated).
Regions of interest (ROIs) included the cortex, hippocampus, thalamus, hypothalamus, amygdala, and white matter areas such as the corpus callosum, internal capsule, and cingulum. ROIs were drawn on the coregistered average sham brain using the rat brain atlas 25 and used across each coregistered individual brain (Supplementary Fig. S1). Similar to our earlier analyses of the sham and untreated-TBI groups, 13 cortical ROIs encompassed motor, somatosensory, parietal, auditory, visual, perirhinal, and entorhinal areas for consistent comparisons (Supplementary Fig. S1). For each modality, group differences were calculated using a voxel-level general linear model (t distribution), with treatment as the contrast and standard deviation calculated at each voxel. Significance was defined as a threshold of p < 0.05 after correcting for multiple comparisons using a random field theory approach, accounting for the cluster size of contiguous significant voxels. 27
Results
Multimodality imaging with DTI-based FA and ADC variables were used as markers of brain parenchymal changes, which is known to correlate with synaptic/axonal microstructure. fMRI-BOLD–based RSFC was used as a marker of neural circuit activity and transient hypercapnia-induced fMRI-BOLD as a surrogate for CVR. Together, these parameters provide a multi-dimensional assessment of the effects of Kaempferol treatment in improving outcomes of developmental TBI.
In our earlier studies, comparison of DTI responses between sham and TBI groups revealed a significant reduction in FA and increased ADC, spreading bilaterally and across the interhemispheric white matter regions such as the corpus callosum. 13 Specifically, bilateral FA decreases were observed in the somatosensory and entorhinal cortices, whereas unilateral FA decreases were prominent in the ipsilateral cingulum and internal capsule ROIs. 13 ADC values between sham and TBI in our previous studies increased significantly in the same ROIs, indicating increased isotropic diffusion after TBI and consistent with the decreased FA. 13 In the current study comparing Kaempferol and vehicle treatments in TBI animals, strong uni- and bilateral increases in FA occurred after Kaempferol treatment when compared to vehicle (Fig. 1A). As observed by the group statistical maps of vehicle versus Kaempferol, FA increases were mostly across the perilesional regions away from the injury epicenter (Fig. 1A). Unilateral FA increases were observed within the ipsilateral perirhinal cortex, hippocampus, hypothalamus, thalamus, and amygdala, whereas bilateral FA increases were observed across motor, somatosensory cortices, insula, and caudate putamen (Fig. 1A). FA increases with Kaempferol treatment across various anatomical regions, statistically determined in Figure 1A, are depicted in Supplementary Figure S2B.
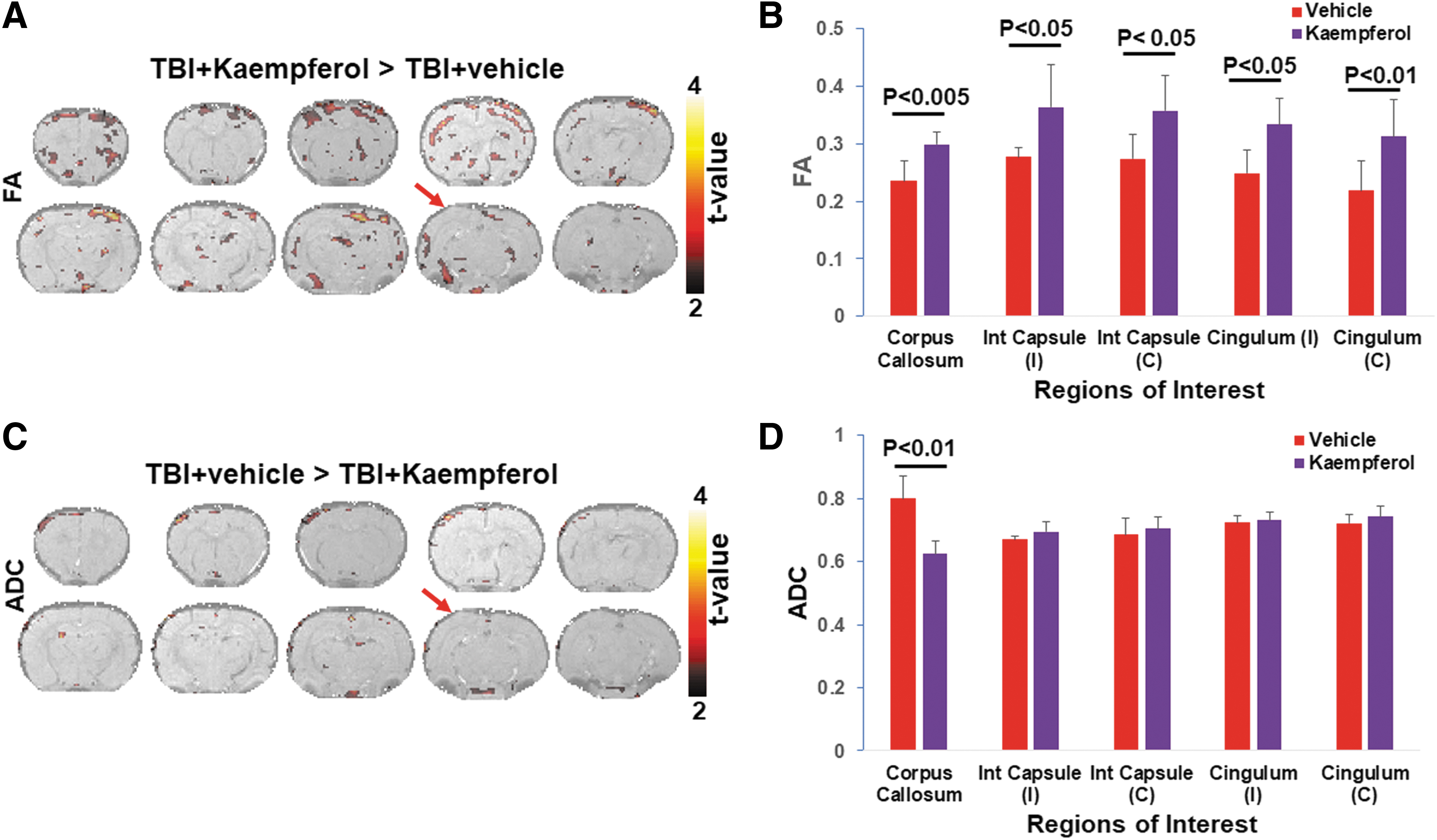
Diffusion tensor imaging group average of fractional anisotropy (FA) and apparent diffusion coefficient (ADC) in TBI animals treated with Kaempferol and vehicle. (
Significant Kaempferol-induced microstructural integrity improvement, as determined by increased FA, was observed in various white matter ROIs, including the corpus callosum and across both hemispheres in the cingulum and internal capsule (Fig. 1B). ADC values decreased after Kaempferol treatment compared to vehicle. Although ADC decreases were relatively sparse compared to FA, the group statistical maps indicated highly specific ADC decreases across ipsilateral cortical regions, hippocampus, and internal capsule and bilaterally across the hypothalamus (Fig. 1C). ADC changes determined across different white matter ROIs showed a significant ADC decrease only across the corpus callosum after Kaempferol treatment when compared to vehicle (Fig. 1D). ADC decreases after Kaempferol treatment across various anatomical regions, statistically determined in Figure 1B, are depicted in Supplementary Figure S2D. Taken together with our earlier results of untreated TBI versus sham, 13 the current DTI results after treatment highlights the protective effect of Kaempferol not only in the most vulnerable regions after TBI, such as the perirhinal cortex, hippocampus, hypothalamus, thalamus, amygdala, and corpus callosum, but also across additional structures such as the insula and caudate putamen.
From our previous studies, all RSFC networks (somatosensory cortical, hippocampal, and thalamic seed voxels) were observed to have higher bilateral symmetry in sham compared to TBI animals. A significant ipsilateral decrease in RSFC network spatial extent was observed in TBI when compared to sham. 13 In the current treatment study, all RSFC networks (somatosensory cortical, hippocampal, and thalamic) showed similar ipsilateral RSFC spatial extent decrease in the vehicle-treated TBI animal group (Figs. 2A, 3A, and 4A) and comparable to the untreated TBI group observed earlier. 13 Kaempferol's effect on RSFC was investigated by comparing the vehicle- and Kaempferol-treated TBI groups across the somatosensory cortical, hippocampal, and thalamic RSFC networks. A significant bilateral increase in RSFC network spatial extent occurred across the cortical and hippocampal networks (Figs. 2A and 3A), whereas no significant changes were apparent in the thalamic RSFC network (Fig. 4A). RSFC strength in various anatomical ROIs within these RSFC networks has been shown previously to decrease after TBI when compared to sham. 13 RSFC strength was estimated from the frontal and parietal cortical, hippocampal, and thalamic ROIs from the vehicle- and Kaempferol-treated TBI groups.
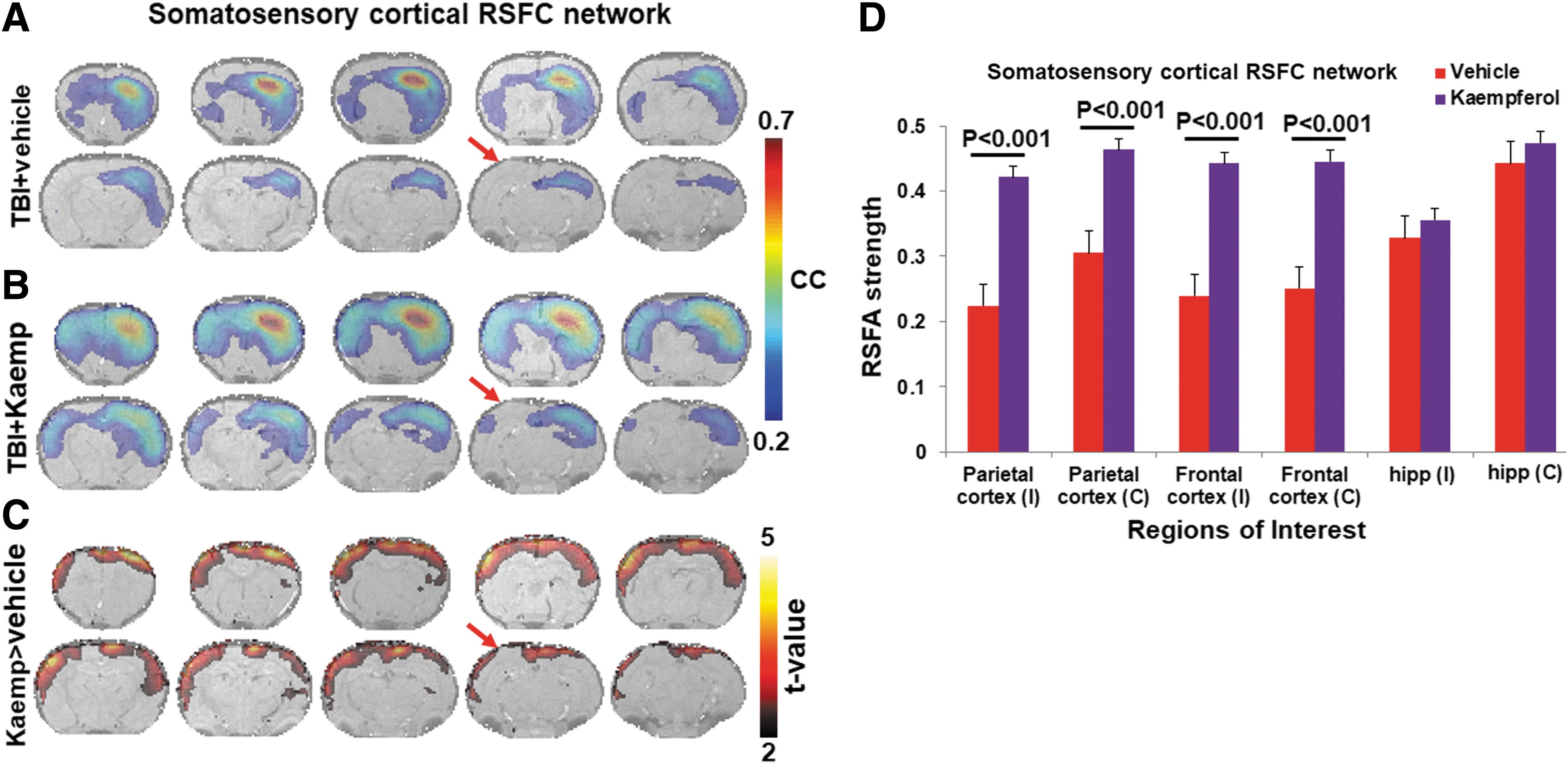
Resting-state functional connectivity (RSFC) maps obtained from the vehicle- and Kaempferol-treated TBI animal groups. Six random seeds from the right somatosensory cortical ROI were used from each subject. Cortical RSFC networks were obtained after cross-correlating the seed voxel fMRI-BOLD time series with the signal time series across all voxels in the brain and subsequently averaged across trials and animals within each group after Fisher's Z-transform. Voxels with average correlation coefficients (cc) >0.2; corresponding to a corrected p < 0.05 represented active voxels of the cortical RSFC network. (

Resting-state functional connectivity (RSFC) maps obtained from the vehicle- and Kaempferol-treated TBI animal groups. Six random seeds from the right hippocampal ROI were used from each subject. Hippocampal RSFC networks were obtained after cross-correlating the seed voxel fMRI-BOLD time series with the signal time series across all voxels in the brain and subsequently averaged across trials and animals within each group after Fisher's Z-transform. Voxels with average correlation coefficients (cc) >0.2; corresponding to a corrected p < 0.05 represented active voxels of the hippocampal RSFC network. (

Resting-state functional connectivity (RSFC) maps obtained from the vehicle- and Kaempferol-treated TBI animal groups. Six random seeds from the right thalamic ROI were used from each subject. Thalamic RSFC networks were obtained after cross-correlating the seed voxel fMRI-BOLD time series with the signal time series across all voxels in the brain and subsequently averaged across trials and animals within each group after Fisher's Z-transform. Voxels with average correlation coefficients (cc) >0.2; corresponding to a corrected p < 0.05 represented active voxels of the cortical RSFC network. (
Kaempferol treatment led to significant increases in RSFC strength when compared to vehicle over multiple anatomical ROIs spanned by the somatosensory cortical RSFC network (Fig. 2B). A similar Kaempferol-induced increase in RSFC strength was observed across multiple ROIs spanned by the hippocampal RSFC network (Fig. 3B). As observed by the relatively larger statistical clusters in the ipsilateral hemisphere adjoining the injury site between vehicle- and Kaemferol-treated TBI animal groups (arrows; Figs. 2C and 3C), Kaempferol was found to reduce interhemispheric asymmetry in the somatosensory cortical and hippocampal RSFC networks.
fMRI measures of CVR were obtained using a transient (3-min) inhalation of 10% CO2 gas mixture to induce hypercapnia as a vasodilatory stimulus. CVR maps, reporting on regional vasomotive functions, were obtained on a voxel-wise-manner basis, based on fMRI-BOLD response amplitude to the CO2 stimulus, as assessed by our previous studies. 13 Whereas sham animals had a significantly higher global CVR compared to untreated TBI as assessed by our previous studies, 13 the current results assessing Kaempferol effects against TBI showed no significant effect on the CVR when compared to vehicle-treated TBI animals (Fig. 5). Quantitative statistical comparisons using a t-test between vehicle and Kaempferol groups, performed voxelwise throughout the brain, did not reveal any significant clusters of altered CVR (data not shown).

Cerebrovascular reactivity response to transient (10%) carbon dioxide inhalation in (
Discussion
Kaempferol dose used in the present study was in the range of human dietary intake of flavonol-rich foods 28 and 1000 times lower than the human safety limits of 1000 mg/kg. 29 No significant differences were observed in mortality between vehicle- and Kaempferol-treated TBI animals. As a natural flavonoid compound, Kaempferol has been shown to specifically act on the mitochondrial Ca2+ uniporter (mCU) channel by enhancing it activity in vitro 14 and be capable of crossing the blood–brain barrier. 30 Low-dose acute Kaempferol treatments in vivo (i.e., 1 mg/kg intravenous) have demonstrated its neuroactive effects of boosting stimulus-induced neurovascular coupling, 31,32 neuronal electrical activity and baseline oxidative metabolism in the normal brain. 32 Studying Kaempferol's mechanism of action in vivo, comparable doses of 1-mg/kg i.p. Kaempferol treatments were made during the acute/subacute window (1, 24, and 48 h post-TBI) and assessed in vivo using the rat model of developmental TBI. 16 This early Kaempferol treatment mitigated TBI-induced brain energy metabolic decline and improved neural viability and behavior at 72 h post-injury. 16
A metabolome-wide analysis of the brain indicated that Kaempferol's effects in the post-TBI brain were predominantly by its action on mitochondria-related biochemical pathways. 16 Kaempferol's mitochondrially mediated effects, including early bioenergetic stress mitigation after TBI, significantly altered the trajectory of developmental TBI at adolescence. Such a long-term beneficial effect on TBI prognosis was supported by our Kaempferol treatment studies, which improved behavioral and stimulation-induced cerebral blood flow (CBF) responses at adolescence (2 months after TBI). 17
Mitochondrial heterogeneity and the hypothesis of differential treatment outcomes across brain compartments and regions
Although in vivo treatment studies have so far established Kaempferol's mitochondrial mechanism of action, neuroactive impact in the working brain, and beneficial effects in developmental TBI outcomes, 16,31 –33 its overall systemic impact within the brain can be hypothesized to vary depending on mitochondrial structural and functional heterogeneity across various cellular populations. Brain regional mitochondrial heterogeneity and its placement within the context of cellular, functional, developmental, and neuroanatomical environment are crucial to interpret pediatric brain pathophysiological changes. Additionally, active consideration of mitochondrial heterogeneity, which is the true in vivo scenario, may help reconcile the differential impact of mitochondrially targeted treatments across various brain compartments. 34 Hence, Kaempferol-induced mitochondrial functional changes and its consequent downstream impact on cellular pathways were hypothesized to manifest in a heterogeneous manner in a variety of cellular populations. The current multi-modal imaging design with clinically relevant markers enabled testing across cellular compartments and brain regions. Different multi-modal imaging outcome results demonstrate the heterogeneous impact of mitochondrially targeted Kaempferol treatment across different cell types of the brain in vivo.
Neuronal (synaptic/non-synaptic) and astroglial mitochondrial impact
RSFC topology was significantly improved across the somatosensory cortical and hippocampal, but not thalamic, networks after Kaempferol treatments, signifying a favorable outcome on neuronal mitochondrial compartments (Figs. 2, 3, and 4). 35 Both synaptic and non-synaptic mitochondria seem to be affected, as observed from the increases in FA (Fig. 1A,B) and somatosensory cortical RSFC network topology and strength (Fig. 2). Earlier studies with the same dose of Kaempferol showed improved stimulation-induced CBF responses, suggesting improved neural circuit activity and neurovascular coupling. 17 Given that astrocytes significantly enable the neurovascular coupling process, their mitochondria are likely to have been impacted, leading to better astrocytic Ca2+ signaling and neurovascular activity. Given that Ca2+ overload occurs in the temporal window of approximately 6 h after TBI through mitochondrial dysfunction and arrest of Ca2+ transport, 36 the ion homeostatic benefits derived by both neuronal and astroglial mitochondria from Kaempferol treatment improve their viability and function.
However, intensity of Kaempferol action is likely to vary depending on functional differences of neuronal and astroglial mitochondria. Even across neurons, both synaptic and non-synaptic mitochondria may derive different Kaempferol treatment benefits with synaptic mitochondria receiving a higher impact. This possibility is attributable to a relatively greater TBI-induced oxidative damage of synaptic mitochondria when compared to non-synaptic. 35 Further heterogeneous effects could be observed in the thalamic RSFC network, which showed no significant impact of Kaempferol (Fig. 4), contrasting the robust neural connectivity improvements within the somatosensory cortical (Fig. 2) and hippocampal RSFC networks (Fig. 3).
Although the thalamic region is very functionally diverse, containing several nuclei with specific functions, the individual thalamic RSFC networks obtained from the six seed voxels placed randomly in different locations within the thalamic ROI have been shown to be topologically stable, as demonstrated our earlier study. 26 Hence, spatially separated selection of the six random seed voxels within all ROIs, including the thalamic ROI, was unlikely to cause any significant RSFC variability, resulting in no improvement of thalamic RSFC after Kaempferol treatment. Thalamic regions also do not display neurodegeneration, but show gradual neuronal atrophy from early perisomatic axotomy after a diffuse brain injury, very similar to our current model. 37 The subsequent post-TBI reorganization of the thalamus is also complex, leading to hypersensitivity in somatosensory responses and hypothesized as a maladaptive reorganization extending into the chronic time periods after TBI. 38 A diminishing RSFC spatial extent and strength in the ipsilateral thalamic regions at adolescence corroborate earlier neuronal atrophic evidence. 37 However, Kaempferol's inability to improve thalamic RSFC strength, in the current study, may point to a differently vulnerable mitochondrial population supporting a prolonged neural atrophic mechanism.
Endothelial and smooth muscle mitochondria impact
Extracellular mitochondria after a central nervous system (CNS) injury 39,40 have been proposed as a cellular rescue signal 41 and been demonstrated to improve endothelial function in vivo after a stroke. 42 Although endothelial mitochondrial properties can be expected to differ from neural mitochondria in certain specific functions, they would generally benefit from Kaempferol treatment given that the mitochondrial Ca2+ uniporter channel (mCU), which is the target for Kaempferol, is expressed in all mitochondrial types. Although it is not known whether mCU densities varied across various distinct cellular mitochondria within the CNS, enhanced survival capability of extracellular endothelial mitochondria from damaged vasculature can potentially alter the overall endothelial population responses post-TBI in a beneficial direction. Supporting this view, improved stimulation-induced neurovascular activity after TBI suggest that the capillaries of the neurovascular units functioned better after Kaempferol treatment. 17 However, no significant improvement was observed in CVR in response to a transient hypercapnic challenge, which predominantly tests the functionality of large and intermediate cerebral arteries. 43,44
The current result infers that smooth muscle mitochondria, and hence their function, were the least affected by Kaempferol treatment. Supporting this result, regional baseline CBF perfusion deficits after TBI, stemming from mostly large and intermediate vessel disruptions after injury, were observed to persist through development in the current model of TBI. 17 Kaemferol treatment, although helpful in improving behavioral and brain responses, had no effect on the decreased baseline CBF perfusion observed ipsilateral to the TBI. 17 Decreased CBF is a hallmark of concussive TBI in humans, 45 and its acute vascular origins and cellular mechanisms have been well studied in pre-clinical animal models of TBI of various severity. 46 Although chronic vascular rearrangement continues beyond 6 months after TBI with significant vessel proliferation in both the lesional and perilesional regions, there are no direct correlations between CBF and regenerated vessel density. 47 Further, prolonged inflammation, blood–brain barrier disruption, and white matter damage may sustain chronic microvascular dysfunctions after TBI. 48
Recent studies indicate impaired myogenic constriction of cerebral arteries through mitochondrial oxidative stress after a TBI, which was restored by mitoTEMPO, a mitochondria-targeted antioxidant. 49 Hence, TBI-induced mitochondrial responses may be very different in vascular cells, necessitating entirely different doses of Kaempferol for optimal effects. Higher doses of Kaempferol are known to induce significant antioxidative and anti-inflammatory responses. Hence, future dose escalation studies may help determine possibilities of cerebrovascular protection through mitochondrially triggered redox- and immunomodulations.
To determine vehicle effects within the brain, untreated and vehicle-treated TBI animal comparisons were made in our earlier study, which showed no significant vehicle-induced effects on the brain metabolome. 2 Hence, the vehicle-treated sham group was not considered in the current study. Further, given Kaempferol's properties as a dietary flavonol compound with proven safety in humans, 28 it is highly unlikely that Kaempferol may have significant effects on the structural or functional components within the normal working brain. Hence, the absence of a Kaempferol-treated sham group in the current study, although a moderate weakness, is very unlikely to alter the overall conclusion of the study.
Conclusion
Given that TBI pathology is broad, its prognosis and treatment outcomes can be best understood multi-dimensionally using a systems approach spanning the genomic, proteomic, metabolomic, tissue, organ, and organism levels. 50 The demonstrated multi-modal translational imaging of TBI treatment and its ability to distinguish mitochondrial treatment responses across various tissue compartments and regions indicate its critical value within this futuristic systems framework. Although the Kaempferol dose used in the current pre-clinical trial favorably impacted neural populations, a lack of cerebrovascular improvement and minimal neural impact in the thalamic region indicated mitochondrial functional diversity. The current results justify future Kaempferol dose/treatment window optimizations extending to cerebrovascular protection, with potential to further enhance the improved behavioral and brain responses after TBI.
Footnotes
Funding Information
Funding from the New Jersey Commission for Brain Injury research (CBIR15IRG010; to S.K.), NIH R01 NS097750 (to V.S.), NIH R01 MH067528 (to F.H.), and NIH P30 NS052519 (to F.H.) are acknowledged.
Author Disclosure Statement
No financial conflicts exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.