
Abstract
Traumatic brain injury (TBI) can lead to chronic diseases, including neurodegenerative disorders and epilepsy. The hippocampus, one of the most affected brain region after TBI, plays a critical role in learning and memory and is one of the only two regions in the brain in which new neurons are generated throughout life from neural stem cells (NSC) in the dentate gyrus (DG). These cells migrate into the granular layer where they integrate into the hippocampus circuitry. While increased proliferation of NSC in the hippocampus is known to occur shortly after injury, reduced neuronal maturation and aberrant migration of progenitor cells in the hilus contribute to cognitive and neurological dysfunctions, including epilepsy. Here, we tested the ability of a novel, proprietary non-invasive nano-pulsed laser therapy (NPLT), that combines near-infrared laser light (808 nm) and laser-generated, low-energy optoacoustic waves, to mitigate TBI-driven impairments in neurogenesis and cognitive function in the rat fluid percussion injury model. We show that injured rats treated with NPLT performed significantly better in a hippocampus-dependent cognitive test than did sham rats. In the DG, NPLT significantly decreased TBI-dependent impaired maturation and aberrant migration of neural progenitors, while preventing TBI-induced upregulation of specific microRNAs (miRNAs) in NSC. NPLT did not significantly reduce TBI-induced microglia activation in the hippocampus. Our data strongly suggest that NPLT has the potential to be an effective therapeutic tool for the treatment of TBI-induced cognitive dysfunction and dysregulation of neurogenesis, and point to modulation of miRNAs as a possible mechanism mediating its neuroprotective effects.
Introduction
Traumatic brain injury (TBI) often leads to permanent disability and chronic disease. 1 More than 10 million TBI cases are reported each year worldwide, with an annual health care cost exceeding 50 billion dollars in the United States. 2 Contact sport players and soldiers are at increased risk of TBI and of developing TBI-associated long-term neuropathology and cognitive dysfunctions. 3 –5 TBI is often related to the onset of neurodegenerative disorders, such as Alzheimer's disease, and epilepsy, 6 –8 with the incidence of seizures increased twofold after mild TBI and up to sevenfold after severe TBI. 9
The hippocampus, one of the most affected areas after TBI, plays a key role in memory and cognitive function and is one of the two brain areas in which new neurons are generated throughout life from neural stem cells (NSC) in the subgranular zone (SGZ) of the DG, which migrate in the granular layer and fully integrate into a pre-existing neuronal network. 10 –13 Neurogenesis is a highly regulated process that is under tight epigenetic control (i.e., DNA methylation, histone modification, and microRNAs [miRNAs]). 14,15 Although physical exercise and exposure to an enriched environment enhance the generation and integration of new neurons, 16 this process declines with aging, in neurodegenerative disorders (e.g., Alzheimer's disease), and after TBI, resulting in impaired memory and cognitive function. 17 –19 Whereas in animal models of TBI, increased proliferation of NSC in the hippocampus is known to occur shortly after injury, 20 recent reports have demonstrated reduced neuronal maturation and aberrant migration of progenitor cells in the hilus, a phenomenon linked with the onset of cognitive dysfunction and epilepsy. 21
Recently, non-invasive therapies (delivered transcranially) such as near infrared laser (NIL) therapy (600–1000 nm) and ultrasound waves (20 kHz–2MHZ) have received increased interest for their neuroprotective and anti-inflammatory properties. 22 –27 NIL has been reported to exert neuroprotective effects and stimulate neurogenesis in animal models of TBI, and to produce beneficial cognitive and emotional effects in individuals affected by brain injury. 28,29 Moreover, the application of low-intensity NIL has been shown to decrease headache, sleep disturbance, and irritability in subjects affected by TBI. 30,31 Ultrasound waves (US) at low-frequencies (20 kHz) have been shown to stimulate brain circuits and be neuroprotective. 23,27,32
We have developed a highly innovative non-invasive device for the transcranial delivery of NPLT. 33 This therapy is unique because it combines the benefits of pulsed (20 Hz) NIL light (808 nm) and low energy ultrasound waves, optoacoustically generated with each short (10 ns) high-energy (15 mJ) laser pulse that can travel to deep structures within the brain (including the hippocampus). 33,34 Our proprietary NPLT device is safe and has been already tested in a clinical setting to monitor adult superior sagittal sinus blood oxygenation. 35
We have recently shown that NPLT is neuroprotective in a rat model of blast-induced neurotrauma. 36 In this work we tested the ability of NPLT, applied transcranially to rats 1 h after fluid percussion injury (FPI), to reduce vestibulomotor and working memory dysfunction, regulate neurogenesis, and prevent aberrant migration of neuronal progenitors in the hippocampus hilus, a phenomenon linked with the onset of epilepsy. 21,37 Moreover, we tested the ability of NPLT to modulate microglia activation and the expression of specific regulatory miRNAs known to play a key role in the epigenetic regulation of NSC proliferation and differentiation. 38
Significance
TBI leads to chronic neurological problems including epilepsy, neuropsychiatric disorders, and dementia. Although TBI stimulates proliferation of neuronal progenitor cells in the hippocampus dentate gyrus (DG), it decreases neuronal maturation and induces aberrant migration of newborn neurons. Our study is the first to demonstrate that, after fluid percussion injury (FPI), treatment of rats with a novel, proprietary non-invasive optoacoustic laser therapy (NPLT) improves cognitive function and prevents TBI-induced impaired maturation and aberrant migration of neural progenitor cells in the hippocampus DG. Moreover, our data suggest that NPLT exerts its protective effect at least in part by mitigating TBI-driven dysregulation of specific microRNAs known to modulate neurogenesis in the hippocampus. Altogether, our study identifies NPLT as a potential therapeutic approach for the treatment of post-TBI cognitive dysfunctions and dysregulation of hippocampal neurogenesis leading to the onset of epileptic seizures.
Methods
Animals
All animal experiments were approved by the Institutional Animal Care and Use Committee of the University of Texas Medical Branch, Galveston, Texas and conducted according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Experimental design
The experimental timeline is shown in Figure 1. Rats were randomized to receive FPI or sham injury and further randomized to receive NPLT or no treatment 1 h later. A total of 18 rats per group were used in the study. Five rats per group were euthanized 24 h after surgery and the brains were processed for immunofluorescence analysis of 5-bromo-2'-deoxyuridine (BrdU) incorporation. Fourteen rats per group were used for behavioral studies and then euthanized 2 weeks after surgery; of these, eight rats/group were used for immunohistological analysis and six rats/group were used for laser-capture microdissection and quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis of microRNA expression. Of these, one brain/group was lost during processing and, therefore, the number of samples for the RT-PCR results is five rats/group.
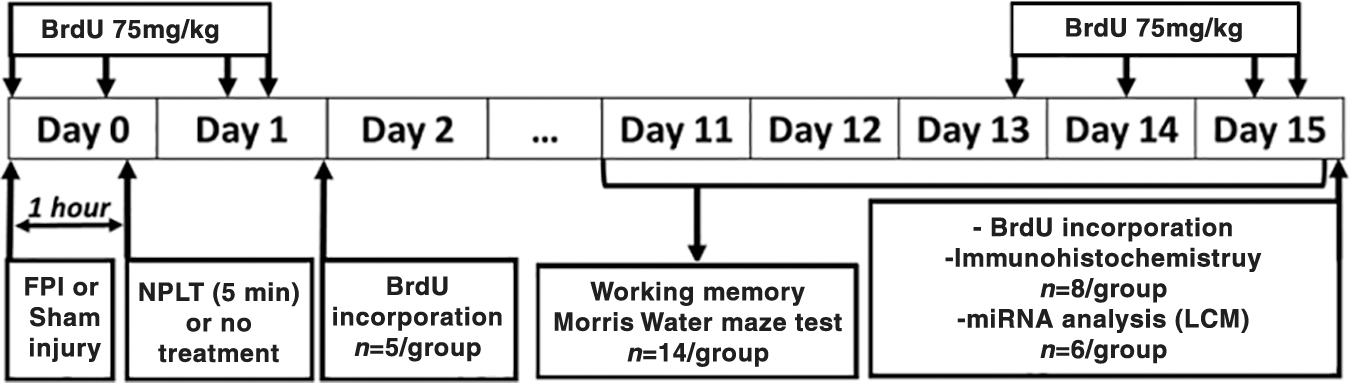
Experimental design. Rats were randomized to receive fluid percussion injury (FPI) or sham injury and further randomized to receive nano-pulsed laser therapy (NPLT) or no treatment. One cohort of rats (n = 5/group) were treated with bromodeoxyuridine (BrdU) (75mg/kg i.p.) at 2, 4, 20, and 22 h after surgery and euthanized 24 h after surgery. A second cohort of rats (n = 14/group) were tested on the neuroscore, beam-balance and beam-walk tasks on post-injury days (PIDs) 1–5 and on the working memory paradigm of the Morris water maze test (MWM) on PIDs 11–15. At the end of the last day of MWM testing, a subset of rats (8/group) were treated with BrdU (75mg/kg i.p.) at 48, 24, 4, and 2 h before euthanasia, and the brains were processed for BrdU and immunohistochemistry analysis, and another subset of rats (6/group) were euthanized, and the brains immediately dissected out and frozen on dry ice for laser capture microdissection (LCM) of the subgranular zone of the dentate gyrus for quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis of microRNAs expression.
For the isolation of NSC, three adult male Sprague–Dawley rats were subjected to FPI and euthanized 1 month after injury.
FPI
Male Sprague–Dawley rats (350–400 g) were housed with food and water ad libitum in a vivarium with constant light cycle, temperature, and humidity. The rats were anesthetized with isoflurane in an anesthetic chamber for 5 min, intubated, and mechanically ventilated with 1.5–2.0% isoflurane in O2: room air (70:30) using a volume ventilator (EDCO Scientific, Chapel Hill, NC). Rats were prepared for moderate (2 atm) parasagittal FPI as previously described. 39 Sham rats underwent craniotomy and were attached to the FPI device but were not subjected to injury. After FPI or sham injury, rats were disconnected from the device, placed in a supine position, and monitored until a righting reflex was observed. Rats were then replaced under isoflurane anesthesia until ready for treatment.
NPLT
Rats were maintained under isoflurane anesthesia for 1 h after FPI or sham procedure and treated for 5 min with NPLT. Our system consists of an optical parametric oscillator that generates short (10 ns) pulses of NIL (808 nm) at energy of up to 15 mJ and pulse repetition rate of 20 Hz. These pulses generate low-level optoacoustic waves that travel deep into the brain. Rats were treated 1 h after FPI for a duration of 5 min to provide a dose of 300 J/cm2. NPLT was delivered through a 3 mm diameter optical fiber on the site of the craniotomy. For untreated animals, the optic fiber was positioned on the craniotomy site for 5 min. The animals were allowed to fully recover from the surgery/anesthesia before being returned to their cage with ad libitum food and water.
Working memory test
To test the effect of NPLT on FPI-induced cognitive deficits, rats were subjected to a working memory paradigm of the Morris water maze (MWM) on post-injury days (PIDs) 11–15 as previously described by Sell and colleagues. 39 Briefly, the experimental animals were placed in a tank filled with water to a level that was 2 cm higher than the hidden platform. Rats were assigned four starting points and four platform locations in a balanced order to avoid starting points too close to the platform. For Trial 1, rats were placed in the tank and allowed 120 sec to find the platform. Once on the platform, the rats were allowed 15 sec to rest and then were placed in the tank again from the same starting point to begin Trial 2. They were again allowed 120 sec to find the platform. Rats were rested 4 min in a heated enclosure before starting a second pair of trials, which used different platform and starting locations. Rats received four pairs of trials daily for 5 consecutive days. All rats received the same sequence of starting points and platform locations.
Vestibulomotor function tests
Neurological function, gross vestibulomotor function, and fine motor coordination were assessed on days 1–5 post-surgery. Rats were pre-trained to each of these tasks 2 days (day -3 to day -2) prior to surgery and assessed for baseline performance 1 day prior to surgery (day -1). 40
Neuroscore
The following reflex tests were administered in order and repeated three times. A normal response received a score of 0 whereas an abnormal response received a score of 1 for each trial of each test for a total possible score of (7 × 3 = 21), the higher the score, the greater the deficit.
Forelimb Flexion Test (0-1)
Hind Limb Flexion Test (0-1)
Visually Triggered Placing Test (0-1)
Contact Triggered Placing Test, right (0 -1)
Contact Triggered Placing Test, left (0 -1)
Hind Paw Grasping Reflex Test, right (0-1)
Hind Paw Grasping Reflex Test, left (0-1)
Beam balance
The balance beam apparatus consisted of a beam 60 cm in length, 1.75 cm in width, 4.0 cm in height, elevated 90 cm off the floor, with a barrier 30 cm in height and 30 cm in width. For training, each rat was placed on the balance beam for a 60 sec trial. The rat was removed from the beam for a brief moment between trials. If the rat failed to balance on its own, it was allowed to fall into a padded safety box. Rats were rated using an ordinal scale with (1) being normal balance and (6) being unable to remain on the beam.
Beam walk
The beam walk apparatus consisted of a beam 100 cm in length, 2.5 cm in width, and 4.0 cm in height with four equally spaced pegs (2 cm in height, 25 cm apart) and a black goal box (28 cm in length, 18 cm in height, 18 cm in width) at one end. The beam was elevated 1 m off the floor, and a bright light and white noise source were positioned near the starting end. 39,40 On day -1, rats underwent a pre-assessment consisting of three timed trials. Three trial assessments were performed on days 1–5 after surgery.
BrdU treatment
Experimental rats received four intraperitoneal (i.p.) injections of BrdU (Sigma Aldrich; 75 mg/kg). The rats in the 24 h experimental group were treated on the day of surgery (immediately after surgery and 2 h later) and the following day at 4 and 2 h before euthanasia. The rats in the 2 weeks experimental group were treated with BrdU at 48 h, 24 h, 4 h, and 2 h before euthanasia.
Tissue processing and immunostaining
One cohort of rats was euthanized 24 h after surgery using 2–3% isoflurane followed by decapitation. The brains were immediately dissected out, frozen on dry ice, and stored at -80°C until further processing. For immunofluorescence analysis of BrdU incorporation, the brains were embedded in optimal cutting temperature compound (OCT), and 10 μm thick sections were cut on a cryostat and mounted on superfrost clean slides. Frozen sections were fixed in ice-cold 10% buffered formalin at room temperature for 30 min. After two washes in phosphate-buffered saline (PBS) (10 min each), sections were incubated with 2N HCl al 37°C for 30 min to denature DNA and unmask the BrdU. The sections were neutralized by washing in 0.1M borate buffer (pH 8.5) twice for 5 min each and then blocked and permeabilized with a solution of 10% normal goat serum and 0.3% Triton X-100 in PBS for 30 min. Sections were incubated with a mouse primary anti-BrdU antibody (1:100 DAKO) and a rabbit anti-NeuN antibody (1:100 Millipore; to label neurons), overnight at 4°C. The day after, the sections were washed three times in PBS for 10 min each and then incubated 1 h at room temperature with Alexa-conjugated antibodies diluted in 1.5% normal serum. Finally, the sections were mounted with FluorSave (Millipore) and viewed with a fluorescence microscope BZ-X710 (Keyence) supported by BZ-X analyzer software (Keyence). The slides were stored in the dark at 4°C.
A second cohort of rats was euthanized 2 weeks after surgery. For immunohistological and immunofluorescence analysis, rats were anesthetized and perfused with saline followed by freshly prepared phosphate-buffered formaldehyde solution (pH 7.4). The brains were dissected and post-fixed in formaldehyde for 12–16 hours at room temperature, transferred to a phosphate buffered solution (PBS) and shipped to NeuroScience Associates (Knowxville, TN) for tissue processing and immunohistological staining using a patented Multibrain® technology. Briefly, 16 rat brains were embedded in one single block (each block containing four brains/group for a total of two blocks), sectioned on a microtome at 40 μm thickness in the coronal plane and collected every 480 μm throughout the hippocampus. The sections were adhered to glass slides (5 × 4 cm) and processed for immunohistochemistry staining of BrdU (to evaluate the number of proliferating cells), doublecortin (DCX) (to evaluate the number of neural progenitor cells) and microglia markers (ionized calcium binding adaptor molecule 1 [Iba1] and cluster of differentiation [CD]68), and for double immunofluorescence staining of DCX and the neuronal marker NeuN (to evaluate the number of neural progenitor cells differentiating into mature neurons). Immunohistochemically stained slides were imaged with a BZ-X710 microscope (Keyence America, Itasca, IL) supported by the BZ-X analyzer software (Keyence America, Itasca, IL). Immunofluorescence-stained slides were viewed with an Olympus BX51 epifluorescent microscope equipped with a cooled charge-coupled device (CCD) camera. Stereological analyses were performed with the aid of a semiautomated, computer-based microscopy systems (Stereo Investigator, MBF Bioscience). Slides were stored in the dark at 4°C (immunofluorescence) or at room temperature (immunoperoxidase).
Cell counts and immunofluorescence quantitative analyses
BrdU incorporation
BrdU+ cells were counted in the SGZ of the DG (defined as a two cell-body thickness region below the granular layer) throughout the hippocampus (bregma -2.16 mm to -6.12mm; 10 sections per brain) by two independent investigators who were blinded to the experimental groups. The length of the SGZ in each section was measured using ImageJ and used to normalize the mean counts of the two investigators. Total mean count for each brain was calculated by summing the mean of the normalized values of each section.
Aberrant migration of DCX+ cells
DCX immunoreactivity in the hilus was quantified using ImageJ and normalized to total hilus area. Specifically, Paxino and Watson's rat brain atlas was used to determine the boundaries and borders of the components of the DG. Then, the area was traced and calculated. DCX+ cells in the granular and subgranular layers of the DG were counted by two investigators who were blinded to the experimental groups, in coronal sections (bregma -2.40 mm to -6.12 mm, for a total of eight sections per brain). The mean of the values obtained by the two investigators for each section was normalized for the area of the granular layer. Total mean count for each brain was calculated by summing the mean of the normalized values of each section.
In order to quantify ectopic migration of the DCX+ cells, the granular layer of the DG was divided in two sublayers (layer I and layer II) arbitrarily selected as two cell-body thicknesses starting from the SGZ. DCX+ cells in each layer were counted by two independent investigators and normalized for the length of the SGZ for each section. The mean of the values of each section were used as total mean count for each brain (eight per experimental group).
CD68+ microglia
CD68+ cells in the DG were counted by two investigators who were blinded to the experimental groups, in coronal sections (bregma -2.40 mm to -6.12 mm, for a total of eight sections per brain). The mean of the values obtained by the two investigators for each section was normalized for the area of the hilus and granular layer, respectively. Total mean count for each brain was calculated by summing the mean of the normalized values of each section.
Iba1+ microglia
Stereological analysis of Iba1+ microglia in the hilus was performed using StereoInvestigator software (MBF) in coronal sections (bregma -2.40 mm to -4.36 mm; total of four sections per brain). Specifically, the contour of the hilus and a grid size with 10–12 random squares (containing an average of five to seven cells) was selected in one coronal section at bregma level -4.16 mm. Mounted section thickness (40 μm) and distance from one section to the other (480 μm) was inserted in the parameters to calculate the total number of cells by the software.
Neuronal differentiation of DCX+ progenitor cells
Stereological analysis of DCX+ and DCX+/NeuN+ cells in the DG was performed using StereoInvestigator software (MBF) in coronal sections (bregma -2.40 mm to -4.36 mm; total of four sections per brain). Specifically, the contour of the SGZ and a grid size with 10–12 random squares (containing an average of five to seven cells) was selected in one coronal section at bregma level -4.16 mm. Mounted section thickness (40 μm) and distance from one section to the other (480 μm) was inserted in the parameters to calculate the total number of cells by the software.
Isolation and in vitro expansion of rat hippocampal stem cells
Reagents for the isolation procedure were obtained from Invitrogen Co. (Carlsbad, CA), except where noted. Three adult male Sprague–Dawley rats (200–250g) were subjected to FPI as described. At 1 month after injury, the rats were anesthetized with isoflurane (4% by inhalation) and euthanized by decapitation. The brain was rapidly removed and the hippocampi dissected out and collected into a 50 mL Falcon tube containing sterile Dulbecco's Modified Eagle Medium: Nutrient Mixture F-12 (DMEM/F12) media with antibiotics (penicillin and streptomycin) and kept on ice. The tissue was minced into small pieces in cold Hanks' Balanced Salt Solution (HBSS) media containing 1 mM egtazic acid (EGTA) in a sterile petri dish. The tissue pieces were transferred into a sterile 50 mL tube containing HBSS (without Ca2+ and Mg2+ with 0.1% collagenase/dispase, 0.01% DNase I, 1 mL for 100 mg of tissue) and incubated for 30 min at 37°C. The tissue was triturated every 10 min at 37°C using a sterile disposable 5 mL pipette. At the end of the incubation, the cell suspension was centrifuged at 200g for 5 min at room temperature. The pellet was resuspended in HBSS containing 0.025% trypsin/ethylenediaminetetraacetic acid (EDTA) and incubated for 10 min at 37°C. An equal volume of DMEM/F12, 2mM L-glutamine, 10% fetal bovine serum (FBS), and antibiotics was added to stop the cell digestion, and the cell suspension was centrifuged at 200g for 5 min. The pellet was resuspended in sterile DMEM/F12 with L-glutamine, 10% FBS, and antibiotics, triturated using a 1 mL tip and centrifuged. This step was repeated two to four times until single cell suspension was obtained. The cell suspension was filtered through a 70 μm Falcon filter, centrifuged at 200g for 5 min and resuspended in complete growth media: neurobasal A media containing the serum-free supplement B27 (without retinoic acid), 2 mM L-glutamine, 20 ng/mL of epidermal growth factor, 20 ng/mL of basic fibroblast growth factor 2, and penicillin/streptomycin. The cells were plated in an uncoated T25 flask at 1 × 106 cells/10 mL of complete growth media. The media was changed after 24 h and every other day after that. Neural stem cells were isolated from the hippocampus of TBI rats and cultured as neuropsheres for 30 days before being used for the NPLT treatment. Adult naïve rat hippocampal primary NSC (Millipore) were cultured following the manufacturer's protocol and used as controls. These cells were plated and cultured as described.
Characterization of rat hippocampal stem cells
To evaluate the stemness of the NSC isolated from TBI rats, we performed Western blotting analysis. For total proteins extraction, NSC isolated from TBI rats (1 month after injury) or control rats growing in suspension, were collected at passage 5 by centrifugation and washed in PBS once before being lysed using RIPA lysis buffer (Thermo Fisher) (500 μL of buffer per 2 × 106 cells) containing 1 μL of protease inhibitor and 1 μL phosphatase inhibitor per 100 μL of buffer to prevent degradation and modification of the phosphorylated proteins and incubate for 10 min on ice. The samples were pelleted by centrifugation (15 min at 14,000g) and the supernatant was aliquoted into clean tubes and stored at -20°C until use. Total protein concentration was determined using the BCA assay kit according the manufacturer's protocol (Thermo Fisher).
Proteins (25 μg per sample) were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) using pre-casted 4–20% gradient gels (BioRad) and transferred to polyvinylidene fluoride (PVDF) membranes. The membranes were blocked with a 5% non-fat dry milk solution in tris-buffered saline (TBS) containing 0.05% Tween-20 (T-TBS) for 1 h at room temperature and incubated with primary antibodies diluted in 0.5% non-fat dry milk in T-TBS overnight at 4°C. The following antibodies were used: mouse anti-nestin (1:1000, Millipore), rabbit anti-sox2 (1:1000, Cell Signaling), and mouse anti-βIII−Tubulin (1:1000, Promega). Membranes were washed three times for 5 min each time with T-TBS and incubated with the corresponding horseradish-peroxidase-conjugated secondary antibody (1:2000, Cell Signaling) diluted in 0.5% non-fat dry milk in T-TBS for 1 h at room temperature. The signal was detected using BioRad ECL and captured using the ChemiDoc imaging system (Biorad). Horseradish peroxidase (HRP) conjugated glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (1:3000, Thermo Fisher) was used as loading control.
In vitro treatment of hippocampus neural stem cells with NPLT
In order to analyze the effect of the NPLT treatment directly on NSC, without other perturbating factors present in vivo, cells were treated in vitro and the expression of select microRNAs known to regulate NSC proliferation, differentiation, and migration was analyzed by qRT-PCR, 24 h later. NPLT was delivered through an optical parametric oscillator probe (3 mm diameter) directly to NSC plated in a 96-well plate. We used a 96-well plate because the diameter of each well perfectly fit with the optoacoustic probe. Five × 105 of cells in suspension were plated in a single well only for the time of the treatment (5 min) and then transferred to a T25 flask at the appropriate density for growth (5 × 105 cells/5 mL) in proliferating media. Because a key feature of NPLT is the production of ultrasound waves (US), generated in the brain by each nanopulse of NIL upon absorption by hemoglobin, 35 we used a black nonporous polyvinyl chloride (PVC) film attached to the bottom of the plate with an ultrasound gel (Aquasonic 100, Parker Laboratories), to act as chromophore for NIL-stimulated US production. The probe for NIL delivery was positioned at the top of the plate, and the US were generated when the laser reached the black nonporous PVC film at the bottom of the plate. The treatment lasted for 5 min for a total energy delivery of 4.3 mJ. Total RNA was extracted 24 h after treatment for qRT-PCR analysis of select miRNAs known to regulate proliferation, differentiation, and migration of hippocampal NSC.
Laser capture microdissection (LCM) and total RNA isolation
Frozen brains were embedded in OCT mounting medium and then sagittal sections (10 μm thick) were cut through the hippocampus (bregma -3.16 mm to bregma -5.16 mm) on a cryostat (Leica Microsystem CM1850) and mounted on superfrost clean glass slides (Superfrost Plus, Thermo Fisher Scientific) The sections were counterstained for 20–30 sec with 1% cresyl violet, and the SGZ of the DG was captured on a thermoplastic film of CapSure Macro LCM caps (1 cap/brain) using a PixCell IIe laser capture microscope with an infrared laser diode. The samples were placed into a 0.5 mL tube containing 100 μL of Lysis buffer, vortexed, and stored at -80°C until isolation of RNA.
Total RNA was isolated from LCM samples using the RNA Aqueous Micro kit (Ambion/Thermo Scientific) according to the manufacturer's protocol before DNase treatment at 37°C for 20 min to remove any traces of genomic DNA. The concentration and quality of total RNA was assessed using an Agilent Bioanalyzer with the RNA6000 Pico Lab Chip (Agilent Technologies).
MicroRNAs retrotranscription
Ten nanograms of total RNA were used for the retrotranscription of the pool of miRNA by using TaqMan Advanced miRNA Assay kit (ThermoFisher Scientific) performed in a thermocycler following this step: Poly (A) tailing reaction one cycle for 45 min at 37°C, then 10 min at 65°C, and held at 4°C. Ligation reaction one cycle for 60 min and 16°C, and held at 4°C. Reverse transcription reaction one cycle for 15 min at 42°C, then 5 min at 85°C, and held at 4°C. miR-Amp reaction one cycle for 5 min at 95°C and then two-step PCR reaction with 14 cycles each for 3 sec at 95°C and 30 sec at 60°C, then one cycle for 10 min at 99°C and held at 4°C.
qRT-PCR
qRT-PCR was performed on a MX3000P system (Stratagene, Santa Clara, CA) using Taqman reagents from Applied Biosystems (Foster City, CA). Twenty microliters of PCR reaction were made by mixing 10 μL of 2X Fast Advanced MasterMix, 5 μL of the product of the retrotranscription diluted 1:10, 1 μL of pre-designed Taqman Advanced miRNA assay primers from Applied Biosystems (Foster City, CA), and 4 μL of nuclease-free water. The thermal profile setup used for the PCR reaction was one cycle of 20 sec at 95°C and a two-step PCR with 40 cycles each for 3 sec at 95°C and 30 sec at 60°C. All data from the PCR was collected and analyzed by the MXPro software (Stratagene) and the ΔΔCT fold changes compared to the calibrator (GAPDH).
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). Incidence of DCX+/NeuN+ and DCX+/NeuN- cells were each modeled by generalized estimating equation with logit link, blocking on animal to control for repeated measures. Differences among groups were assessed by Tukey-adjusted contrasts. Other outcomes were modeled by analysis of variance (ANOVA), with heterogeneous variance controlled by weighting. Differences among groups were assessed by Tukey-adjusted contrasts. A 95% level of confidence was assumed (α = 0.05). Statistical analyses were performed using R statistical software (R Core Team, 2019, version 3.6.0 and GraphPad 7 Prism software).
For the Neuroscore, beam balance, and beam walk assessments, data were expressed as mean ± SEM and were analyzed using a 4 (Injury: SHAM, TBI, SHAM+NPLT, TBI+NPLT) × 5 (Days: 1–5) two-factor ANOVA with replication followed by Student's t tests for differences between the means. Differences were considered significant when p < 0.05.
Working memory water maze results are expressed as the overall mean latency + SEM of Trial 1 minus Trial 2 (Trial 1-Trial 2) for each group for days 11–15 post-injury. Two-factor ANOVA (treatment group × day) followed by post-hoc Student's t test was performed. Differences were considered significant when p < 0.05.
Results
NPLT prevents TBI-induced deficits in fine vestibulomotor coordination
To evaluate the effect of NPLT on vestibulomotor coordination, on days 1–4 after surgery, rats were evaluated on the beam walk, beam balance, and neuroscore tasks as previously described. 40 Because of large variances, there were no differences among groups in the neuroscore task. In the beam-balance task, there was a significant within-group effect of surgery on day 1 compared with day -1 in all groups. This continued for day 2 compared with day -1 for both TBI and TBI+NPLT groups. Therefore, neither the neuroscore task nor the beam-balance task were sensitive to effects of NPLT. However, the beam walk task indicated a protective effect of NPLT treatment in both SHAM+NPLT and TBI+NPLT groups. There was a significant within-group difference between day 1 and day -1 for both untreated SHAM and untreated TBI. For untreated TBI this difference continued for days 2–4. Because there were no differences between any days and day -1 for either NPLT-treated group (SHAM+NPLT, TBI+NPLT), this demonstrates that treatment with NPLT protected against deficits in vestibulomotor coordination (Fig. 2).

Nano-pulsed laser therapy (NPLT) does not improve performance in the neuroscore or beam-balance tasks, but prevents impairment on the beam-walk task. All rats were pre-trained for the neuroscore, beam-balance and beam-walk tasks, and baseline assessments were performed on day -1. There were no significant effects of day or injury group for the Neuroscore task (upper). Significant differences were observed within each group for day 1 compared with day -1, and for both treated and untreated traumatic brain injury (TBI) groups on day 2 compared with day -1 on the beam-balance task (middle). Significant differences were detected for both untreated groups (SHAM, TBI) on day 1 compared with day -1, and for untreated TBI on days 2–4 compared with day -1, indicating that both NPLT-treated groups (SHAM+NPLT and TBI+ NPLT) were protected from impairment on the beam-walk task (lower). * p < 0.05 versus day -1 (within group).
NPLT prevents TBI-induced working memory impairment
To evaluate the effect of NPLT on cognitive function, on days 11–15 after surgery rats were subjected to the MWM test (working memory paradigm) as previously described. 40 The difference between the latency to find the platform during Trial 1 and the latency during Trial 2 (Trial 1 – Trial 2) was significantly less for untreated injured rats (TBI) than for uninjured rats (SHAM), indicating that injured rats did not remember the location of the platform from Trial 1 when undergoing Trial 2 for each session. The greater the difference between Trial 1 and Trial 2 the better the rat has remembered the location of the platform learned during Trial 1, such that the latency to the platform during Trial 2 will be shorter (Trial 1 – Trial 2 will be greater). Injured rats treated with NPLT were not significantly different from SHAM or SHAM+NPLT rats. Taken together, these results show that NPLT treatment protected working memory function in injured rats (Fig. 3).

Nano-pulsed laser therapy (NPLT) treatment prevents the impairment in working memory caused by traumatic brain injury (TBI). Rats received fluid percussion injury (TBI) or sham-injury (SHAM) followed by either NPLT treatment (+NPLT) or no treatment. On post-injury days (PIDs) 11–15, rats were tested using the working memory water maze test (n = 14/group). Working memory water maze results are expressed as the average delta (Trial 1 – Trial 2) + standard error of the mean (SEM) for each group. A larger number indicates a better working memory because of a shorter latency in the second trial compared wit the first trial (i.e., better memory retention of the platform location from the first trial). Two-factor analysis of variance (ANOVA) (treatment group × day) overall F 3,260 = 3.30, p = 0.021, post-hoc t test *p < 0.05 versus day -1 (within groups).
NPLT does not affect TBI-induced changes in NSC proliferation in the hippocampus DG
In order to evaluate the proliferation rate of hippocampal NSC in the SGZ of the DG, 24 h or 2 weeks post-surgery, rats subjected to TBI or sham injury and treated or not treated with NPLT, received BrdU (four ip injections) and were euthanized 2 h after the last injection. We found that 24 h after TBI, the number of BrdU+ cells were significantly higher in the SGZ ipsilateral to the injury site (Fig. 4C). No significant differences in the number of BrdU+ cells in the SGZ were observed in the contralateral side 24 h after TBI or sham injury with or without NPLT (Fig. 4D). At 2 weeks after TBI, a small reduction of BrdU+ cells was observed in the SGZ, in both ipsilateral and contralateral sides of the brain, but only reached statistical significance in the contralateral side compared with sham rats (Fig. 5C, D). NPLT administered after TBI or sham injury did not significantly affect the number of BrdU+ cells at 24 h or 2 weeks compared with untreated TBI and sham groups.

Nano-pulsed laser therapy (NPLT) does not prevent increased cell proliferation of neural stem cells (NSC) in the subgranular zone (SGZ) of the hippocampus dentate gyrus 24 h after traumatic brain injury (TBI).

Nano-pulsed laser therapy (NPLT) does not prevent decreased cell proliferation of neural stem cells (NSC) in the subgranular zone (SGZ) of the hippocampus dentate gyrus 2 weeks after traumatic brain injury (TBI).
TBI increases neuronal progenitor cells in the hippocampus DG but decreases their maturation, an effect prevented by NPLT
Neuronal maturation of DCX+ neuronal progenitor cells requires an intermediate stage in which the mature neuronal marker NeuN is co-expressed with the immature progenitor marker DCX. 41 Therefore, in order to evaluate the effect of TBI and NPLT on neural progenitor cells' maturation, we quantified, using sterological analysis, the number of DCX+/NeuN- cells (immature neural progenitor cells only expressing DCX) and of DCX+/NeuN+ cells (representing an intermediate stage of maturation between immature neural progenitor and fully mature neurons only expressing NeuN) in the hippocampus DG 2 weeks after FPI or sham injury (Fig. 6). Our data show that the total number of DCX+ cells was significantly increased in the DG of TBI rats ipsilateral to the injury side, compared with sham rats (Fig. 6B). Among all DCX+ cells, the proportion of DCX+/NeuN- cells was also significantly increased after TBI (Fig. 6C), whereas the proportion of DCX+/NeuN+ cells (cells that have started the transition from progenitor cells to mature neurons) was significantly decreased (Fig. 6D). On the other hand, the proportion of DCX+/NeuN+ cells and of DCX+/NeuN- cells in the TBI+NPLT group was not different from that in the sham group (Fig. 6C, D). No significant differences were observed in the ipsilateral DG between sham rats and sham rats treated with NPLT (Fig. 6 B–D). No significant changes in the number of DCX+ cells, the proportion of DCX+/NeuN+ cells and DCX+/NeuN- cells were observed in the contralateral side (Fig. 6 E, F). These data suggest that, although the number of DCX+ cells is increased after TBI, an event that is prevented by NPLT, fewer progenitor cells become mature neurons.

Traumatic brain injury (TBI) increases neuronal progenitor cells in the hippocampus dentate gyrus (DG) but decreases their differentiation, an effect prevented by nano-pulsed laser therapy (NPLT).
NPLT prevents aberrant migration of neural progenitor cells in the hilus and in the granular layer of the DG after TBI
Ectopic migration of neural progenitor cells, in particular in the hilar region of the DG, has been previously reported after TBI and is known to be associated with the onset of epileptic seizures. 42 –44 To assess the number of neuronal progenitor cells in the hilus, rats subjected to FPI or sham injury and treated or not treated with NPLT, were euthanized 2 weeks after surgery, and coronal brain sections were immunostained for DCX (Fig. 7A, B). Quantification analysis showed a significant increase in DCX immunoreactivity in the ipsilateral hilus after TBI, which was significantly reduced by NPLT treatment (Fig. 7C). No significant differences between sham rats, treated or not treated with NPLT, were observed. The contralateral side showed no significant differences among the groups (Fig. 7D).
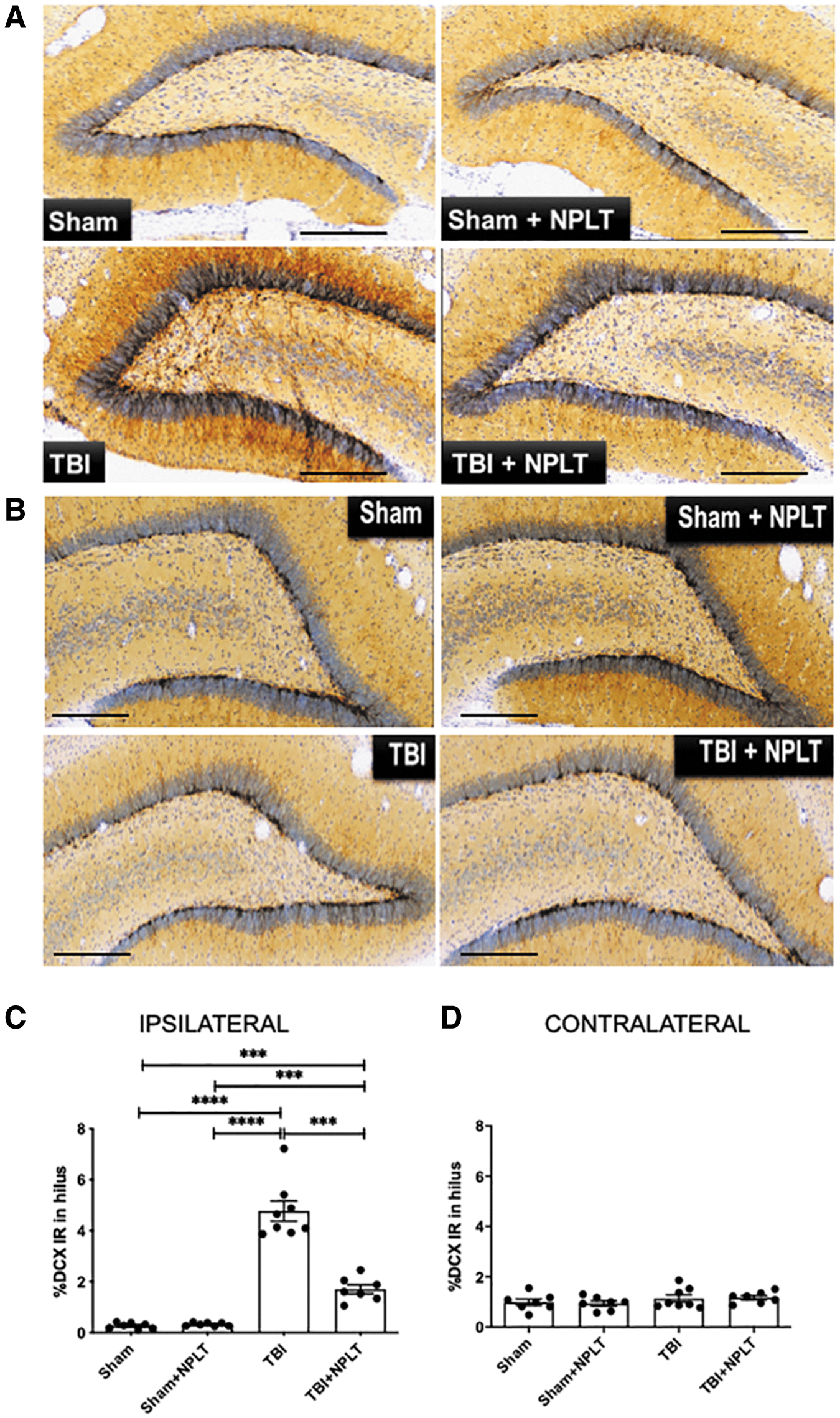
Nano-pulsed laser therapy (NPLT) prevents ectopic migration of neural progenitor cells in the hilus of the hippocampus dentate gyrus 2 weeks after traumatic brain injury (TBI).
The same sections were used to assess the number of DCX+ neuronal progenitor cells in the granular layer migrating toward the molecular layer. The granular layer was arbitrarily divided into two sublayers (namely layer I and II starting from the SGZ) (Fig. 8A). Stereological analysis showed a significant increase of DCX+ neuronal progenitor cells in the DG layers I and II ipsilateral to the injury in TBI rats compared with sham rats and with sham rats treated with NPLT (Fig. 8B). Although the number of DCX+ cells in layers I and II of TBI rats treated with NPLT was significantly increased compared with sham rats, they were significantly lower than in TBI rats. No significant differences were observed between sham rats treated or not treated with NPLT. In the contralateral side, DCX+ cells were significantly increased after TBI as compared with both SHAM and SHAM+NPLT rats in layer I and as compared with only sham rats in layer II. In layer I, TBI+NPLT rats showed a significant increase of DCX+ cells compared with SHAM but not SHAM+NPLT or TBI rats. No significant differences between sham rats, treated or not treated with NPLT, were observed in the contralateral side of the injury (Fig. 8C).
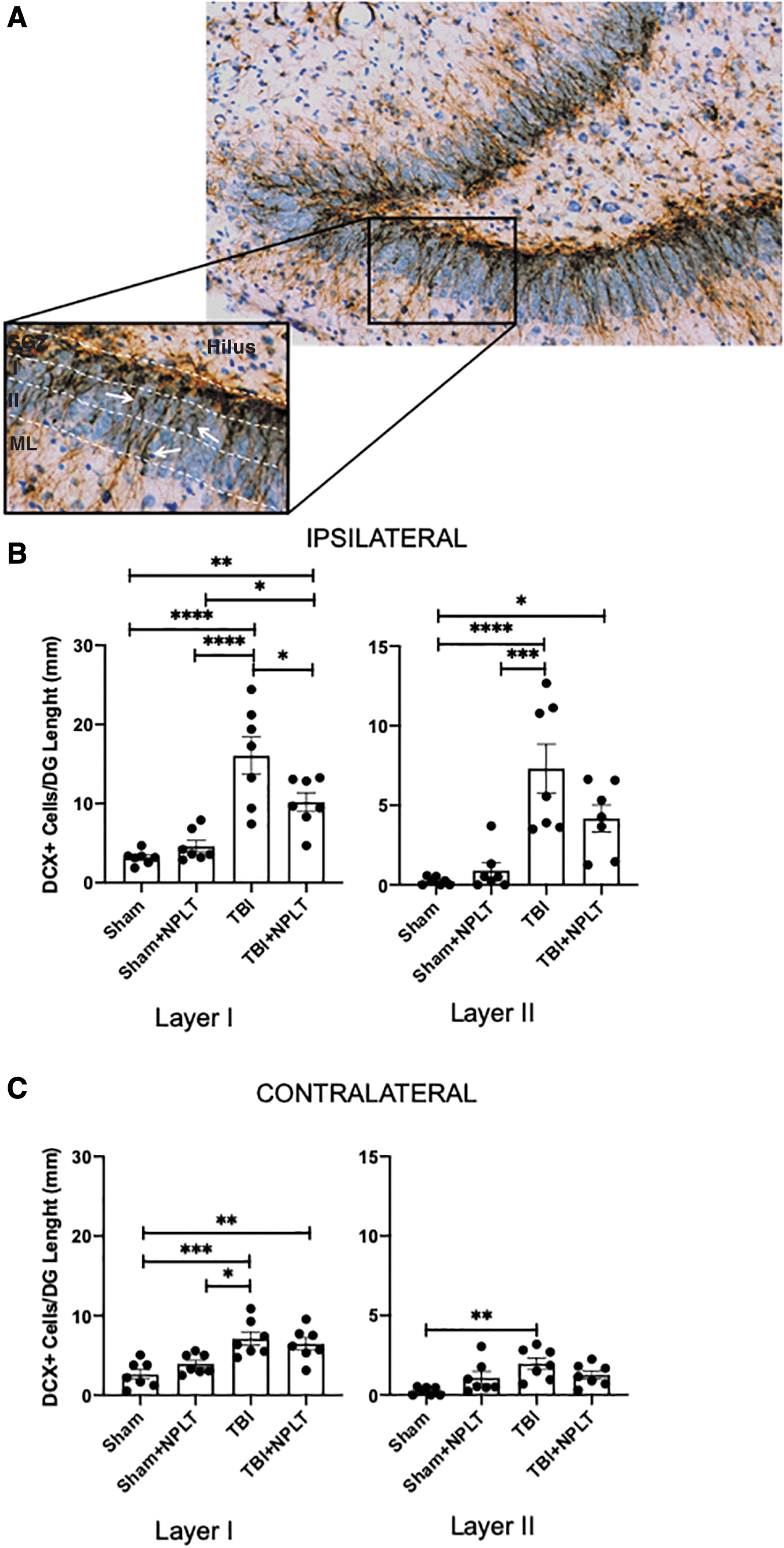
Nano-pulsed laser therapy (NPLT) prevents ectopic migration of neural progenitor cells in the granular layer of the hippocampus dentate gyrus 2 weeks after traumatic brain injury (TBI).
NPLT does not prevent TBI-induced microglia activation in the hilus and in the granular layer of the DG
Microglial activation has been shown to contribute to aberrant migration of neuronal progenitor cells. 45,46 Rat brain sections were stained with a specific antibody against CD68, a marker for phagocytic microglia, and for Iba1 expressed on both resting (ramified) and active (amoeboid) microglia. Stereological quantification showed a significant increase of both resting and active Iba1+ cells in the hilus of the hippocampus 2 weeks after TBI, as compared with SHAM and SHAM+NPLT rats, which was unchanged in NPLT-treated rats (Fig. 9). Moreover, we found that TBI resulted in a significant increase of CD68+ microglia in the hilus and dentate gyrus (Fig. 10). Similarly to what we observed for Iba1+ microglia, NPLT did not reduce the number of activated CD68+ microglia (Fig. 10). These results suggest that the observed effect of NPLT on preventing TBI-induced aberrant migration of DCX+ progenitor cells in the hilus and granular layer of the DG is not likely to be mediated by reduced microglia activation, but it could possibly be the result of a direct effect of NPLT on neuronal progenitor cells.

Nano-pulsed laser therapy (NPLT) does not prevent traumatic brain injury (TBI)-induced ionized calcium binding adaptor molecule 1 (Iba1)+ microglia activation in the hippocampus hilus.
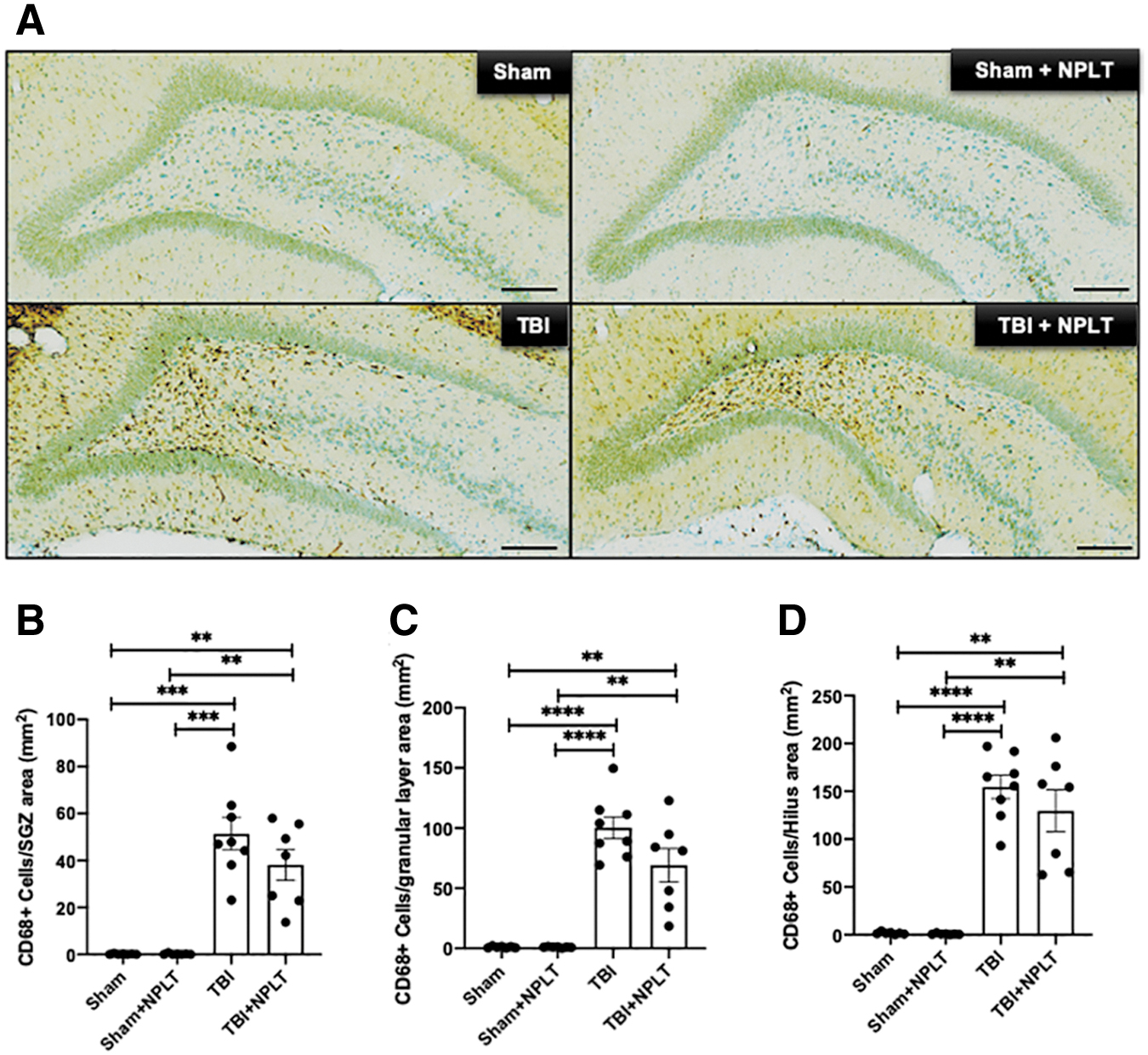
Nano-pulsed laser therapy (NPLT) does not prevent traumatic brain injury (TBI)-induced upregulation of cluster of differentiation (CD)68+ (active) microglia in the hippocampus hilus.
NPLT modulates the expression of specific miRNA in hippocampal neural stem/progenitor cells
Neurogenesis is a process tightly regulated by epigenetic mechanisms. To determine whether NPLT can modulate the expression of specific miRNAs, known to regulate proliferation, differentiation, and migration of hippocampal neural stem/progenitor cells, 47,48 we used LCM to selectively isolate the SGZ of the DG of rats 2 weeks after TBI or sham injury with or without NPLT. We found that the expression of miR9, miR25, miR29, miR124, and miR137, assessed by qRT-PCR, was significantly increased in the SGZ of TBI rats compared with sham rats (Fig. 11). Interestingly, after TBI, NPLT significantly decreased the expression of the same miRNAs (Fig. 11).
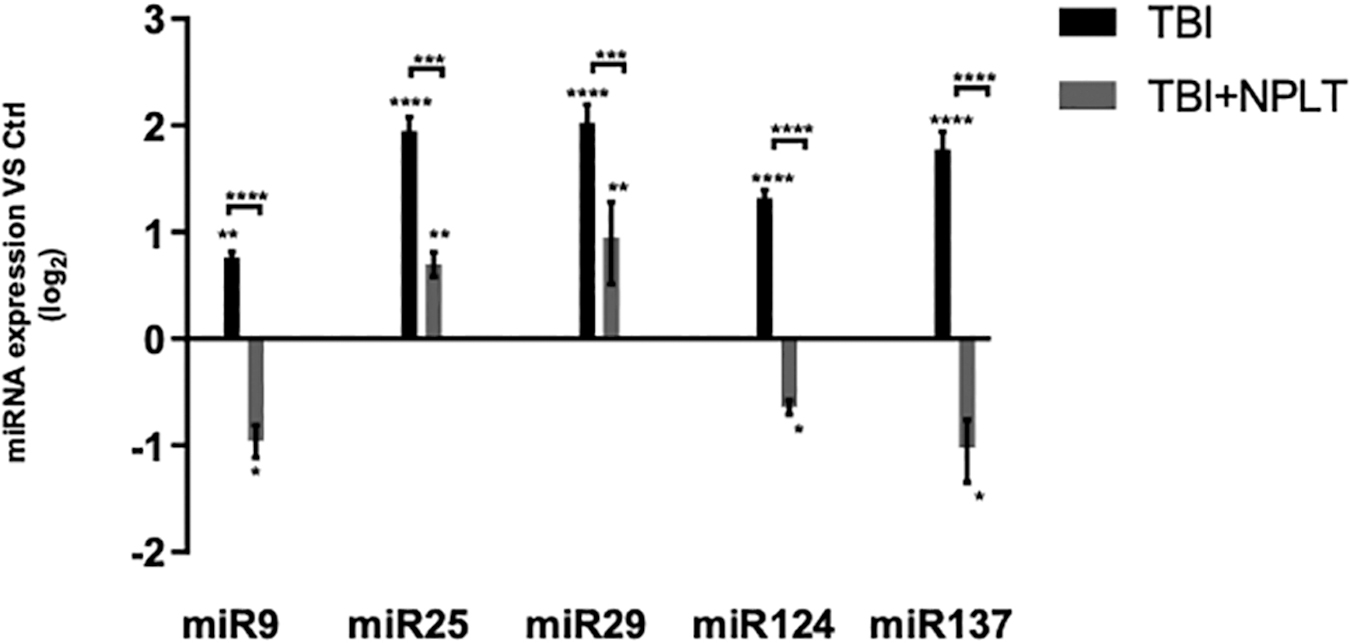
Nano-pulsed laser therapy (NPLT) significantly reduces the expression of microRNA (miRNA) in the subgranular zone (SGZ) of the hippocampus dentate gyrus (DG) that are upregulated following traumatic brain injury (TBI). Quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis of the expression of miRNA known to regulate neurogenesis in the SGZ of the hippocampus DG isolated by laser capture microdissection (LCM). Data are expressed as Log2 fold change versus sham-injured rats (Ctrl). n = 8; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 two-way analysis of variance (ANOVA) with Tukey's post-hoc test.
In order to assess whether NPLT can directly modulate miRNAs in NSC, we first isolated NSC from the hippocampus of rats 1 month after FPI (TBI-NSC) and assessed their stemness with specific markers (Fig. 12A), then we treated them with NPLT for 5 min. The expression of miRNAs was assessed 24 h later by qRT-PCR. We found that the expression of miR9, miR25, miR29, miR124, and miR137 in TBI-NSC was significantly increased as compared with NSC isolated from naïve rat hippocampus (controls). NPLT significantly reduced the expression of all miRNAs in TBI-NSC as compared with untreated TBI-NSC (Fig. 12B).

Nano-pulsed laser therapy (NPLT) reduces the level of microRNA (miRNA) in hippocampal neural stem cells (NSC) isolated from traumatic brain injury (TBI) rats.
Discussion
TBI is characterized by permanent structural and physiological changes that compromise the normal function of the brain. 1,49 One of the regions most affected by TBI is the hippocampus, which is located in the temporal lobe and plays an essential role in learning and memory. 10,20,50 Indeed, memory impairment and disorientation are among the most important symptoms reported by TBI patients. 1 The hippocampus is also a region of the brain in which neurogenesis continues throughout life. 51 In this work, we report that a highly innovative, non-invasive therapy, which combines NIL (808 nm) and US, optoacoustically generated with each short (10 ns) high-energy (15 mJ) laser pulse within the brain, prevents TBI-induced impairments in cognitive function and in neurogenesis in the hippocampus of rats subjected to FPI. 36
We first assessed the effect of NPLT on hippocampus-dependent cognitive function with the working memory paradigm of the MWM. Our data show a significant reduction of latency in the TBI group, demonstrating a memory impairment induced by the injury. After treatment with NPLT, the injured animals took on average the same time to reach the platform as the sham animals, indicating a beneficial effect and recovery of the working memory induced by NPLT. There were no differences between NPLT-treated and untreated sham rats.
Then we assessed the proliferation of NSCs by measuring the incorporation of BrdU in the SGZ of the DG of the hippocampus. Our data show that 24 h after TBI (consistent with published literature), the proliferation of NSCs is significantly increased in the DG ipsilateral to the injury site but not in the contralateral site. 52,53 Interestingly, 2 weeks after TBI, the number of proliferating cells in the SGZ was slighlty reduced as compared with the sham group, suggesting a depletion of the pool of NSCs (that were initially stimulated in response to the injury). These results are consistent with other reports showing that NSC proliferation decreases 1 and 3 months after TBI. 52 NPLT did not change the increase in NSC proliferation 24 h after TBI.
Neuronal maturation of NSCs leads to the generation and integration of newborn neurons into the pre-existing circuit in the hippocampus DG (a critical event for memory formation and consolidation). 54 In order to evaluate how TBI alters this fundamental aspect of neurogenesis, we performed double immunostaining for DCX and NeuN and used standard stereological methods to quantify the number of newborn granular neurons in the hippocampus DG. 41,55 –57 We show that 2 weeks after TBI, whereas the number of DCX+ progenitor cells is significantly increased in the ipsilateral DG, most likely as result of the increased proliferation of NSCs after injury, the proportion of neural progenitor cells that become mature neurons (DCX+/NeuN+ cells) is reduced. Interestingly, NPLT treatment prevents TBI-induced increase in DCX+ progenitor cells, thus suggesting that, although not affecting TBI-induced proliferation of NSC, NPLT might be affecting their neuronal maturation. Because this is only observed in the ipsilateral side, it suggests a direct effect of injury and NPLT on neural progenitor cells in the DG.
In addition to inducing an abnormal proliferation and differentiation of neuronal progenitor cells, TBI can also influence the migration of neural progenitor cells. 58 Indeed, the aberrant migration of immature neurons in areas outside the granular layer of the hippocampus DG, such as the hilus, has been previously reported in rodent models of TBI, and has been shown to strongly correlate with the onset of seizures and increased incidence of epilepsy after injury. 44,59 Specifically, seizures have been reported to occur as early as few weeks following TBI with some manifesting months to years after injury. 9 Therefore, preventing aberrant migration of neuronal progenitors and the presence of displaced neurons in the hilus of the DG is an important therapeutic goal for reducing TBI-induced epilepsy. 21,44 In the present work, we have shown that, in the rat FPI model of TBI, neural progenitor cells (DCX+) aberrantly migrate in the hilus of the hippocampus, where they are found 2 weeks after injury. This aberrant migration is prevented by NPLT applied 1 h after TBI. Moreover, the NPLT treatment corrects abnormal migration of immature neurons in the granular layer of the DG induced by TBI. Taken together, these results strongly support the therapeutic potential of NPLT for preventing the pathological migration of immature neurons, and the associated onset of seizures, following TBI.
In order to assess whether the effects of NPLT on neuronal progenitor cell migration could be, at least partially, the result of its anti-inflammatory properties, we performed immunostaining for Iba1 (a marker of microglia) and CD68 (a more specific marker of activated/phagocytic microglia). 60 Interestingly, we found that NPLT did not reduce the numbers of CD68+ microglia (that was significantly increased after TBI as compared with sham), in the hippocampus SGZ, granular layer, and hilus. Therefore, our data strongly suggest that NPLT prevents aberrant neuronal migration by directly affecting stem cells and neuronal progenitor cells in the hippocampus. Moreover, the quantification of Iba1 (a marker for total microglia) shows, as expected, an increase of both resting and activated (hypertrophic-ameboid) microglia after injury. 61
Finally, to further study the changes induced by NPLT on neural progenitor cells, we used LCM to selectively isolate the SGZ of the DG and to analyze the expression of specific miRNAs (miR9, miR25, miR29, miR124, and miR137), known to be involved in the regulation of neurogenesis (differentiation of stem cells, neurite formation, migration, and integration). 38,62 Our data show that NPLT prevents or significantly mitigates TBI-induced increase in the expression of regulatory miRNAs. To further confirm a direct effect of NPLT on NSC, we isolated NSC from the hippocampus of TBI rats and we cultured them for 1 month. Similarly to what we observed in the LCM samples, TBI-NSC showed a significant increase of the same regulatory miRNAs, indicating a persistence of TBI-induced epigenetic changes in daughter cells. This increased expression is reduced after in vitro treatment with NPLT, thus further supporting a direct effect on NSC.
Conclusion
Taken together, our data demonstrate that, in the rat FPI model of TBI, treatment with NPLT improves cognitive function and prevents TBI-induced impaired maturation of neural progenitor cells in the hippocampus DG and their aberrant migration, one of the key factors that, in addition to seizure and epilepsy, leads to the failure of recovery after brain trauma. Moreover, our data show that, in neural stem cells, NPLT corrects TBI-driven dysregulation of specific miRNAs known to modulate their neuronal maturation and migration in the hippocampus. Although further studies are needed in order to firmly establish the role of TBI and NPLT on neuronal maturation and migration of progenitor cells, our work provides strong evidence for a protective effect of NPLT when applied in the acute phase after injury, and it prompts future studies aimed at testing the therapeutic potentials of this promising non-invasive treatment for TBI patients.
Footnotes
Acknowledgment
We thank Clark Anderson for assistance with the statistical analysis of the data.
Funding Information
These studies were completed as part of an interdisciplinary research team funded by the Moody Project for Translational Traumatic Brain Injury Research.
Author Disclosure Statement
No competing financial interests exist.