
Abstract
Variability in recovery among concussed athletes can be attributed to several risk factors. One risk factor not definitively explored is genetic variation. Genetic variations such as variable number tandem repeats (VNTR) in the promotor region are normal in the population, and can lead to disparities in the amount of protein produced, which could be associated with neuronal recovery. Little research has been conducted to investigate promoter VNTRs within genes responsible for recovery following a concussion. The authors implemented a prospective cohort design using a standardized concussion protocol to diagnose and follow 93 athletes to full recovery at three different sites to determine the association between promotor GT(n) VNTR polymorphisms and recovery time within concussed athletes. The GT(n) VNTR within the promoter region of glutamate ionotropic receptor N-methyl-d-aspartate (NMDA) type subunit 2A (GRIN2A), potassium voltage-gated channel subfamily H member 2 (KCNH2), glutamate ionotropic receptor kainate type subunit 1 (GRIK1), and neurofilament light (NEFL) were genotyped using capillary electrophoresis. GT(n) VNTR promotor polymorphisms were dichotomized into long (L) and short (s) alleles. Using adjusted negative binomial regression models we found that athletes carrying the LL GRIN2A GT(n) VNTR within the promoter region were more likely to experience a prolonged concussion recovery, which resulted in their not being able to return to play for ∼60 days. Additionally, there was a trend toward significance, in which the ss NEFL GT(n) Caucasian athletes had prolonged concussion recovery. This could presumably be attributed to altered proteins or protein levels that disrupt neuronal recovery. This pilot study suggests that these VNTRs are associated with prolonged concussion recovery. In future studies, we plan to measure the extent to which the L or s alleles alter the level and the activity of the GluNR2a and NEFL proteins that GRIN2A and NEFL produce, respectively.
Introduction
Concussions can be transient injuries that are usually relieved within 10–14 days; however, many athletes experience concussion signs and symptoms (s/s) for ≥30 days. 1 –4 This wide variability in recovery times is important to note because if an athlete returns to activity prior to the recovery, sustaining additional head impacts may lead to a protracted recovery, or risk of other musculoskeletal injuries. 5 Moreover, repetitive or multiple concussions have been associated with longer s/s recovery time for future concussions and the development of chronic diseases (e.g., depression, anxiety, chronic traumatic encephalopathy, cognitive deficits) later in life. 6 –8 Understanding the risk factors for prolonged concussion recovery can support medical professionals who are managing an athlete's safe return to play post-concussion. Moreover, greater understanding of risks factors could in turn prevent or lessen both short- and long-term negative outcomes.
There are several proposed risk factors that may contribute to prolonged concussion recovery. Risk factors include specific populations (e.g., children), 9,10 sport type (e.g., American football), 11,12 persistent concussive s/s, 13 history of previous concussions, 14,15 and biological sex. 16,17 Additionally, some researchers are focusing on intrinsic risk factors, such as genetics, to further understand concussion pathophysiology as well as help to individualize concussion care. Many researchers have investigated the genetic associations between concussion risk and poor recovery outcomes. 18 –22 In this context, apolipoprotein E (APOE) is one of the most studied genes in this field, and has been linked with concussion risk and poor recovery; however, the findings are inconclusive. 21,23 –25 Therefore, to expand our understanding of the potential genetic risk factors for poor outcome, the identification of other genes that code for proteins responsible for neuronal recovery such as proteins that regulate the ionic flux (e.g., efflux of potassium, influx of calcium) and glutamate release is warranted. Changes in these signaling events (e.g., indiscriminate release of glutamate, increase in intracellular calcium) can occur after a neuron has been stretched following a concussive event, and altered levels or abnormal proteins could alter the ability to restore neuronal homeostasis. Therefore, genes such as the potassium voltage-gated channel subfamily H member 2 (KCNH2; 7q36.1) that codes for the potassium channel responsible for the efflux of potassium, and the N-methyl-d-aspartate receptor 2A subunit (GRIN2A;loci 16p13.2) and glutamate ionotropic receptor kainate type subunit 1 (GRIK1; 21q21.3) that are responsible for the coding glutamate receptors that aid in synaptic plasticity and efficiency are examples of target genes that could alter an athlete's concussion recovery time. Similar to how some regard APOE as a target because of its role in the response and repair of neuronal tissue, 26 neurofilament light polypeptide (NEFL; 8p21.2) codes for the NFL protein that is necessary for axonal and dendritic growth, function, and maintenance, 26 which is needed for neuronal repair and recovery. Lastly, these genes were chosen because of polymorphisms found to be associated with poor outcomes such as schizophrenia 27 and bipolar. 28 Therefore, DNA sequence variations in genes including KCNH2, GRIN2A, GRIK1, and NEFL could alter response to stress in the context of concussion and thereby influence outcome post-injury.
An individual's genotype determines the structure and function proteins, and ultimately, how the body will respond to injury or stressors (e.g., head impact). The amount of the protein produced may be altered, possibly altering the function of the protein as well. Altered levels or abnormal proteins can occur because of variations in the promoter region of the gene. One common variation that could cause alterations in protein production and function is a repeat polymorphism. Repeat polymorphisms are a type of sequence variation, in which two or more nucleotides are repeated, altering the sequence length of an individual (e.g., GTGTGTGTGTGTGT vs GTGTGT). If this sequence variation occurs within the promoter region, transcription may be affected, which can affect the amount or functionality of protein being produced. 28 Individual structural differences in neuronal proteins, such as channel proteins that allow ion flow and synaptic efficiency, could influence the way an athlete's brain responds to an applied force, and may represent a predisposing factor to concussions. However, much of the research in this field measures protein's activity caused by single nucleotide polymorphisms (SNPs).
Modern genome analysis techniques such as genome wide association studies (GWAS) provide details regarding SNPs; however, they are unable to target short tandem repeats. Therefore, possible associations with short tandem repeats can be missed if GWAS is solely implemented to identify genetics associated with conditions such as concussions. Approximately 3% of the human genome is made up of repeat polymorphisms, and the GT(n) motif is a common repeat polymorphism. 28 Additionally, there are more repeat polymorphisms within promoter regions than in other regions of the gene, and those that lie in the promoter region near the transcription start site are highly conserved. 28 Many promoter repeat polymorphisms have the potential to alter phenotype by generating mutations in regulatory elements, which could ultimately result in disease. For example, the variable number of repeats in the GRIN2A promoter region alters the expression level of GRIN2A. 27,29 –31 However, little is known regarding other genes that have a GT(n) motif within promotor, which also produce proteins that are important for neuronal recovery. Therefore, this study sought to determine the genetic variability of the GT(n) variable nucleotide tandem repeat (VNTR) within the promoter regions of KCNH2, GRIN2A, GRIK1, and NEFL, and the association with concussion recovery time. We hypothesized that there would be a difference in allele and/or genotype distribution between two groups of athletes with normal versus prolonged recovery from concussion.
Methods
Participants
University institutional review boards approved this study. Enrolled participants were recruited from two different universities (Division I and Division II) as well as from an orthopedic clinical setting. The orthopedic setting consisted of a clinic where athletes were referred to one of two physicians who specialized in concussion assessment. Athletes who sustained a concussion between September 1, 2011 and April 25, 2015 who met inclusion criteria, and signed the informed consent or assent were included in the study. Inclusion criteria were a Glasgow Coma Scale of 15, not having been sent for imaging studies (e.g., no suspicion of swelling, lesion), and a concussion sustained from a sport-related mechanism. Athletes went through the concussion assessment battery, provided salivary samples for isolation of DNA, and were followed prospectively until full return to play clearance date was determined by the treating physician. A concussion case for this study was defined as the patient having a sports-related pathomechanical event followed by concussive s/s (e.g., headache, dizziness) within 48 h of injury. The final analysis included 93 participants.
Concussion assessment battery
Participants completed a standardized initial concussion evaluation in which the following parameters were assessed: injury characteristics including the date and time of injury, mechanism of injury, acute s/s (e.g. loss of consciousness, dizziness), patient history including self-reported prior concussion history, migraine, hyper-attention deficit disorder, and psychiatric history (e.g. depression). Initial evaluation was followed by an objective screening that included vestibular ocular assessments, the Balance Error Scoring System (BESS) test, and an online Immediate Post-Concussion Assessment and Cognitive Testing (ImPACT). These assessments were also used to determine the athletes' readiness to return to play. 2,32
DNA collection, purification, and estimation
DNA collection
Following the concussion assessment, DNA was collected using Oragene®•DNA Self Collection Kits (DNA Genotek, Ottawa, Canada) for genotyping. For a valid saliva sample collection, athletes must refrain from eating, drinking, and smoking 30 min prior to collection. Therefore, samples were collected at the end of the concussion evaluation, because the evaluation took ∼30 min. The saliva was transported and stored at room temperature.
DNA extraction
DNA was extracted from saliva collection containers according to the manufacturer's standard methodology. In brief, saliva sample tubes were incubated in an air incubator at 50°C for 1 h. Next, 400 μL was transferred into a 1.5 microcentrifuge tube labeled with the corresponding participant identification number. Sixteen μL of Oragene Purifier was added to the sample and mixed by inversion for 3 min by hand. Following the inversion, the tubes were incubated on ice for 10 min. The samples were centrifuged for 3 min at 13,000g. Two hundred microliters of the clear supernatant was transferred into a new 1.5 microcentrifuge tube labeled with the corresponding participant identification number, and the remaining sample was discarded according to Occupational Safety and Health Administration (OSHA) regulations. The samples were withdrawn for DNA purification using the DNA Tissue Kit protocol implemented in EZ1 Biorobot software (Qiagen, CA). Samples, elution tubes, and cartridges were loaded into the BioRobot EZ1 workstation and the extraction process, which took ∼20 min to process. The BioRobot EZ1 uses magnet bead technology for extraction of DNA from different sources including saliva (our tissue source). The last step of the protocol DNA was eluted to a final volume of 200 μL. The DNA solution was stored at 4°C.
DNA estimation
Purified DNA was quantified fluorometrically using a Quant-it PicoGreen dsDNA assay kit (Invitrogen, CA), according to manufacturer's instructions with modifications. Briefly, the sample DNA diluted to a final volume of 100 μL with 1 × TE buffer was mixed with 100 μL of 1 × PicoGreen dsDNA reagent buffer in a 96 black well plate. Fluorescence data were collected by a SpectraMax M2 (Molecular Devices, CA) fluorescence plate DNA concentration in samples were calibrated from calibration curve prepared from DNA standard provided with the Quant-it PicoGreen dsDNA assay kit (Invitrogen, CA). Samples and standards were measured with the excitation and emission wavelengths set at 480 nm and 520 nm, respectively.
DNA amplification
The DNA region surrounding the VNTR within the promoter of each of the genes was amplified by polymerase chain reaction (PCR) with the forward (FWD) or 5’-FAM-labelled forward (FAM-FWD) and reverse (REV) primers. Primers sequences used in this study can be found in Table 1. PCR reaction mix contained 20–40 ng genomic DNA, 0.1 mM of each primer and 1.25 units of AmpliTaq Gold DNA polymerase (Applied Biosystems, CA) in 3.5 mM MgCl2 at pH 8.5. The cycling conditions were 94°C for 2 min, 35 cycles (94°C for 1 min, 55°C for 2 min, 72°C for 1 min), followed by incubation at 72°C for 7 min.
Gene Selection and Primers
NMDA, N-methyl-d-aspartate.
Purification of the reaction mix and genotyping
The PCR products (5 mL) were mixed with 2 mL of ExoSAP-IT reagent (Affymetrix, Santa Clara, CA) and then were incubated at 37°C for 15 min followed by thermoinactivation at 80°C for 15 min. The reaction mix was analyzed by capillary electrophoresis (Genewiz, South Plainfield, NJ). Fragment length polymorphism analysis (FLP) was performed by measuring the migration time of a PCR product and extrapolation to the known fragments in the DNA standard ladder using Peak Scanner software vs1.0 (Applied Biosystems) using internal standards. The number of GT VNTR was calculated. The genotyping success rate was 100% for GRIK1 and GRIN2A, and 99% for NEFL and KCNH2. The amplification products were inserted into pCR4-TOPO cloning vector (Invitrogen). The recombinant constructs were used for transformation of bacterial strain TOP10. The identity of amplified products was confirmed by sequence analysis.
Pedigree analysis of (GT)n polymorphism
To validate the analytical method for (GT)n VNTR genotyping, a pedigree analysis of (GT)n alleles in a three-generation family was performed. To this end, saliva was collected from six members of the three-generation family, and genotyping analysis was performed using the established protocols described. The pedigree chart analysis confirmed that the protocol established in this work accurately detected the presence of allelic variants both in homo- and heterozygous individuals and that the corresponding alleles are inherited according to the Mendel's laws. Therefore, the pedigree analysis validated the protocol for (GT)n genotyping both in homo- and heterozygotes.
Statistical analyses
To test the association between (GT)n VNTR genotypes and recovery time in the overall sample, we first conducted an adjusted negative binomial regression analysis followed by an adjusted model controlling for race (Caucasian vs. not) and assuming a (co-dominant; LL vs. Ls vs. ss) genetic model. Models did not additionally adjust for sex, age, prior concussion, and contact sports, as these were not deemed confounders (e.g., not significantly associated with (GT)n VNTR genotypes and recovery time) of any significant associations between (GT)n VNTR genotypes and recovery time. SAS vs 9.2 (SAS Institute, Cary, NC) was used for all analysis. Significance was set at p < 0.05. An adjusted Bonferroni threshold setting at 0.05/4 = 0.012 were made for multiple testing (four tests conducted) illustrating the potential clinical significance of the associations between (GT)n VNTR genotypes and recovery time.
Results
Ninety-three athletes sustaining a concussion were between 12 and 53 years of age (median = 20, Interquartile range [IQR] = 4), primarily Caucasian (71%), and male (68%; Table 2). More than half of the athletes had sustained a previous concussion (55%), and the majority engaged in contact sports including American football, soccer, rugby, and lacrosse (76%).
Demographic Characteristics of 93 Athletes Sustaining a Concussion
SD, standard deviation; IQR, interquartile range.
Allele distribution and recovery time
In agreement with the previous findings, the allele distribution found that patients with poor brain injury outcomes were carriers of the the longer GRIN2A (GT)n alleles.
27,29
–31
Because of the suggested length-dependent suppressive effect of the GRIN2A (GT)n repeat on gene transcription and the observed allele distribution patterns, we dichotomized the VNTR alleles into two classes according to length: short (≤ 24 repeats) and long (> 25 repeats) as previous authors have done.
27,29
–31
Currently, there is no standard rational criterion for determining the (GT)n repeat allele. The ambiguity arises from not knowing the exact in vivo activity that is the result of each allele by the specific repeat length.
27
As this is the first study to determine variable (GT)n repeat lengths for NEFL, KCNH2, GRIK1, we determined cut points using the mean allele lengths determined from the Peak Scanner Software to dichotomize long allele (L) and short allele (s) within each gene, as described previously in similar studies.
33,34
Allelic groups for each of the genes are as follows. GRIN2A: L allele
GRIN2A is the most variability of the promoter (GT)n ranging from 19 to 36 repeats (19 = 0.5%, 20 = 1.6%, 21 = 29%, 22 = 9.1%, 23 = 6.5%, 24 = 5.9%, 25 = 4.8%, 26 = 9.7%, 27 = 11.8%, 28 = 5.9%, 29 = 5.4%, 30 = 6.5%, 31 = 2.2%, 36 = 1.1%). KCNH2 promoter (GT)n ranged from 10 to 16 repeats (10 = 1.1%, 11 = 35.9%, 12 = 22.3%, 13 = 3.8%, 14 = 32.1%, 15 = 0.5%, 16 = 4.3%). GRIK1 promoter (GT)n ranged from 24 to 28 repeats (24 = 5.4%, 26 = 2.7%, 27 = 88.7%, 28 = 3.2%). NEFL promotor (GT)n ranged from 3 to 11 repeats (3 = 0.5%, 4 = 21.2%, 5 = 37%, 6 = 10.3%, 7 = 10.9%, 8 = 4.9%, 9 = 8.7%, 10 = 5.4%, 11 = 1.1%).
Genotype distributions were in Hardy–Weinberg equilibrium for the entire sample (ps > 0.05) for all genes except KCNH2 (χ2 = 32.72, p < 0.000; Table 3). With respect to the time to recovery, the median recovery time was 23 days (IQR = 44). Only 10% of the athletes recovered within 10 days, 25% recovered within 14 days, 50% recovered within 24 days, 75% recovered within 61 days, and 90% recovered within 150 days (Fig. 1).

Distribution of days to recovery among the 93 athletes sustaining a concussion.
GT Genotype Distributions in a Sample of Athletes Sustaining a Concussion
One missing KCNH2 and NEFL genotype data.
p < 0.05.
GT(n) VNTR association to prolonged recovery
In the analyses in the overall sample, only GRIN2A was significantly associated with time to recovery (χ2 = 7.91, p = 0.010). Specifically, the predicted number of days to recover was 63.4 days (standard error [SE] = 12.7) for LL carriers; whereas, ss carriers were predicted to recover in 43.6 days (SE = 5.2), and Ls carriers were predicted to recover in 29.4 days (SE = 5.3; Fig. 2). LL carriers took twice as long (estimated mean = 2.1, SE = 0.58) to recover than Ls carriers, χ2 = 8.1, p = 0.004; the trend was for for LL carriers to need a longer recovery time than ss carriers (mean estimate = 1.5, SE = 0.32, χ2 = 3.3, p = 0.06). After race adjustment, LL carriers still showed a slower recovery time than Ls carriers, χ2 = 6.1, p = 0.010. Re-running the analysis using the dominant unadjusted model in the overall sample, GRIN2A continued to be associated with time to recovery, with LL carriers taking 1.6 times longer to recover (SE = 0.37) than Ls and ss carriers combined, χ2 = 4.8, p = 0.03. The same pattern remains after adjusting the model for race, χ2 = 5.0, p = 0.02. There were no differences detected between recovery and the KCNH2, NEFL, or GRIK1 (GT)n VNTR genotypes in the total population.
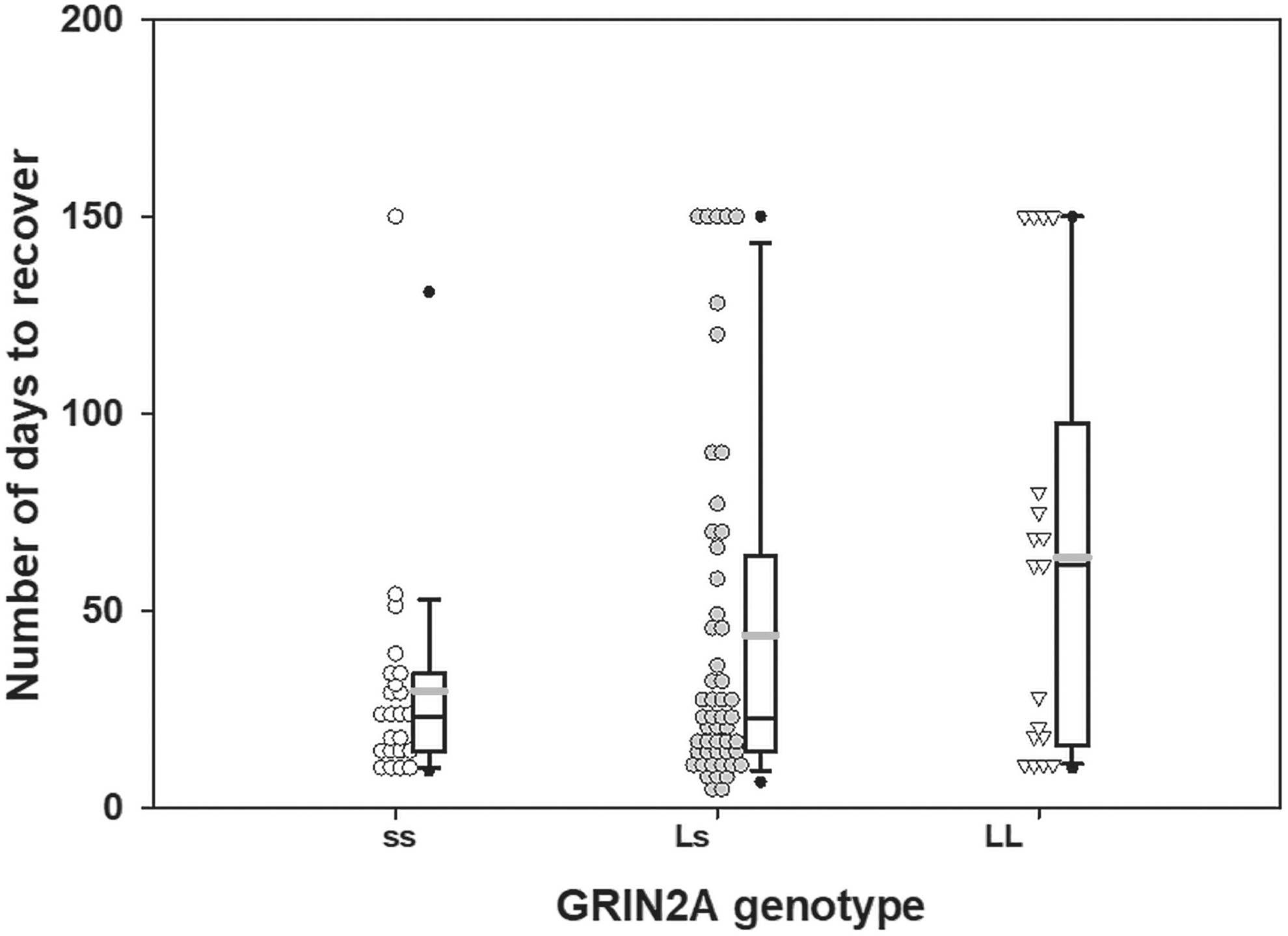
Effect of the glutamate ionotropic receptor N-methyl-d-aspartate (NMDA) type subunit 2A (GRIN2A) genotype (ss, homozygous short genotype; Ls, heterozygous genotype; LL, homozygous genotype) on the number of days to concussion recovery in the overall sample. The gray line indicates the mean and the black line indicates the median.
Association with recovery in Caucasians
A second series of binomial regression models analyses were conducted in Caucasians only (n = 66; 71% of the sample). For GRIN2A, the same pattern of a trending association remained, χ2 = 7.7, p = 0.020. The pairwise differences were stronger, whereby, LL carriers needed longer to recover than both Ls (mean estimate = 2.7, SE = 0.93, p = 0.004) and ss carriers (mean estimate = 1.8, SE = 0.50, p = 0.03). In contrast to the overall sample, NEFL was trending toward association with time to recovery, χ2 = 6.3, p = 0.04. Specifically, the predicted number of days to recover was 28.6 (SE = 8.9) in LL carriers, 40.0 (SE = 6.0) in ss carriers, and 64.3 (SE = 11.7) in Ls carriers. Ss carriers took 2.2 times (SE = 0.81) (Fig. 3) longer to recover than LL carriers, χ2 = 5.0, p = 0.02, and ss carriers took 1.6 times (SE = 0.38) longer than Ls carriers to recover, χ2 = 4.0, p = 0.04.
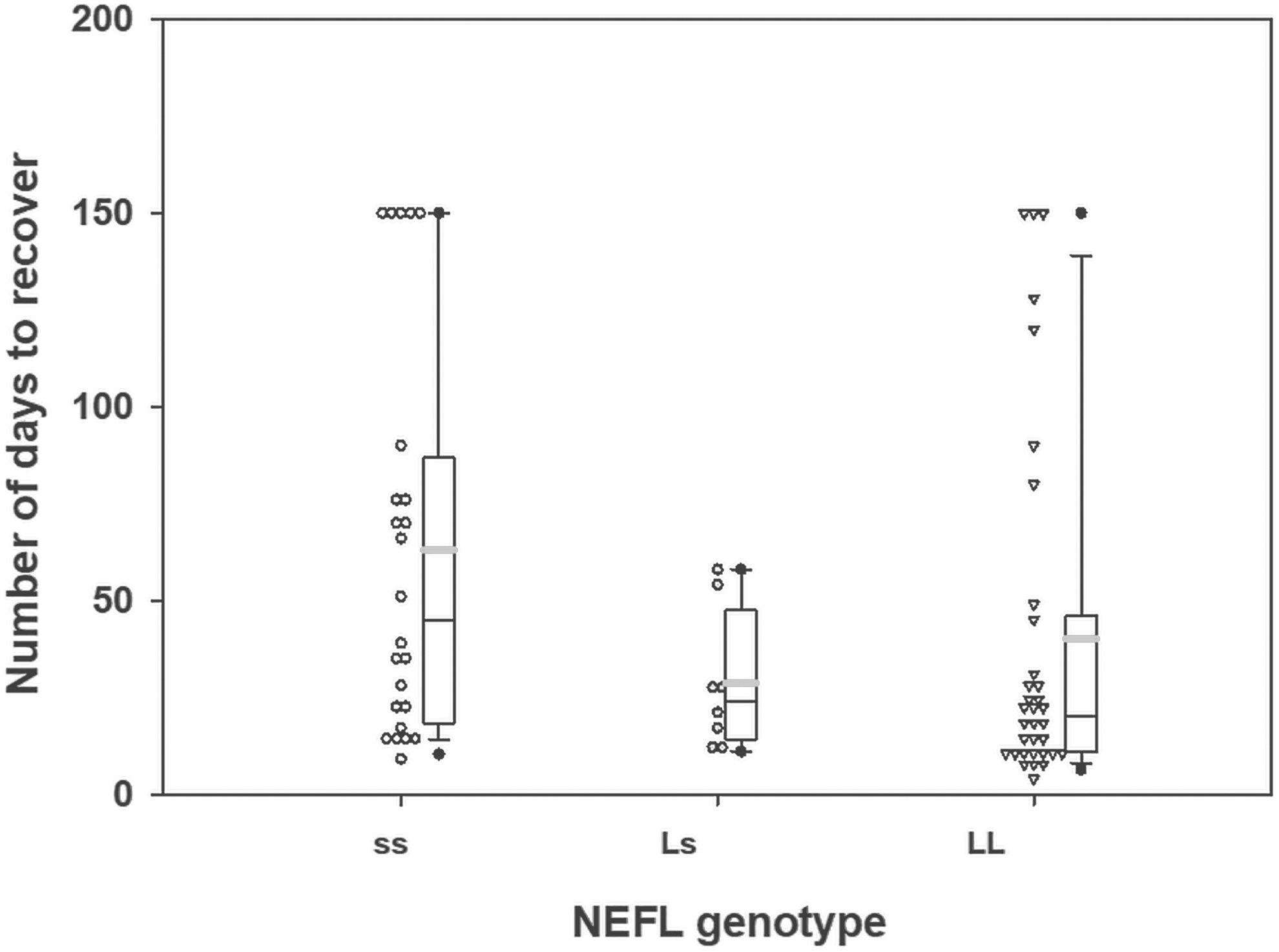
Effect of the neurofilament light (NEFL) genotype (ss, homozygous short genotype; Ls, heterozygous genotype; LL, homozygous genotype) on the number of days to concussion recovery in Caucasians. The gray line indicates the mean and the black line indicates the median.
Discussion
In the present study, we analyzed four different genes with the promotor (GT)n VNTR polymorphism from 93 participants with concussions. GRIN2A LL (GT)n VNTR was associated with prolonged concussion recovery, and NEFL ss was trending toward prolonged recovery within the Caucasian population. In particular, carriers of the GRIN2A LL genotype were nearly two times slower to recover from concussion than Ls and ss carriers. Additionally, after accounting for race, carriers of the NEFL ss genotype were twice as likely to have a prolonged concussion recovery as Ls and LL carriers. Both GRIN2A LL and NEFL ss genotype carriers were more likely to recovery in >60 days. Although concussion position and consensus statements convey that return to play following a concussion is typically between 10 and 14 days, 2,32,35 in our sample, only 25% of the athletes recovered within this time frame. We found that 50% of our participants needed ∼24 days to return to play, and 71% of our cohort were fully recovered in 61 days. Thirty-five percent of our participants were recruited from an orthopedic clinical setting concussion program, which may contribute in part to the longer recovery time. It could be suggested that athletes taking longer to recover from concussion s/s may seek out further healthcare assistance. It could also be suggested that current guidelines are focused on the mean time to return to play rather than accounting for what total number of athletes are actually returning within that time frame. Further, Vagnozzi and coworkers demonstrated that following concussion, metabolic syndrome can last up to 30 days post-injury. 36 Given this finding, the “normal” 7–10 days to recovery may not be the best window to begin the return to play progression to activity for all cases. Because 50% of the participants in our study returned to play within 30 days, this may coincide with metabolic syndrome dissipation and represent a normal and safe return to play period.
Using what is known regarding the pathophysiological pathway following a concussion, 37 –39 four candidate genes were chosen because of the proteins they produce that are active during neuronal recovery. The (GT)n VNTR in the promoter region has been earlier associated with expression level of GRIN2A. 27,29 –31 The length of (GT)n repeats modulates GRIN2A expression level, with longer alleles (> 25 repeats), associated with lower transcription of GRIN2A mRNA. 27 Additionally, studies attribute poor brain outcomes to the GRIN2A (GT)n LL genotype. Specifically, carriers of this genotype demonstrate a greater risk for smaller brain volumes 30 and psychiatric disorders including bipolar disorder 29 and schziphrenia. 27,31 Findings could be attributed to the decreased transcription of the GluNR2a subunit, which is necessary for N-methyl-d-aspartate (NMDA) channel functioning. The results of this study demonstrated that carriers of the LL genotype are almost twice as likely to recover in ≥60 days. These findings suggest that the expression level of GluNR2a may be altered because of this varied promotor sequence motif, which could alter neuronal recovery and prolonged concussion s/s and return to play time for athletes.
This was the first pilot study to identify a trend toward a significant association between NEFL ss genotype and concussion recovery within the Caucasian population. NEFL is a protein that is preferentially expressed to maintain the structural integrity of a neuron. Much of the research surrounding NEFL is in regard to its use as a blood biomarker. Axonal injury can be detected immediately after injury when forces of sufficient magnitude are applied to the brain. Microtubule disruptions can occur within 2 min after rapid stretch of the axons of cultured neurons. 42 Additionally, abnormal accumulation of NEFL in degenerated motor neurons are seen in patients with amyotrophic lateral sclerosis. 33 NEFL abnormalities are associated with decreased axon conduction rates, secondary demyelination, and profound structural alteration in the cytoskeleton. 42 Structural alteration of the cytoskeleton was brought on by a polymorphism within the NEFL gene, which caused an axonal neuropathy that lead to dysfunction within intermediate filaments organization. 42 It should also be noted that NEFL is more abundant than the heavy and medium neurofilaments and is capable of homologous assembly, in which heavy and medium neurofilaments are not able to assemble in the absence of NEFL. 33 This is the reason that NEFL's primary function is neurofilament assembly, because it is the only neurofilament protein capable of organizing the filament by itself. 43 Therefore, it is not surprising that an alteration in the promoter region could be associated with prolonged recovery in our population.
It was interesting to note that NEFL (GT)n ss carriers were trending toward a significant association with prolonged recovery as opposed to the LL carriers with GRIN2A. There are numerous genes with (GT)n polymorphisms, and they vary in which L or s allele carriers are associated with protein transcription. For example, GT VNTR of 13 within the promoter of signal transducer and activator of transcription 6 (STAT6) was associated with atopic asthma, in which longer repeats showed a decreased risk for asthma. 44 Conversely, long GT repeats within cyclin dependent kinase inhibitor (CDKN)1A/CDKN1C/CDKN2A were associated with increased risk of artheroscerlosis. 34 Therefore, it is not surprising that in our study we found different genotypes (i.e., GRIN2A, LL and NEFL, ss) to be associated with prolonged recovery.
This research also contributes to the greater knowledge of genomic study protocols. Genomic analysis techniques have excellent tools to deal with heterogenous stretching of DNA. However, these techniques are not as effective when the target regions are homogeneous or tandem repeats. Nevertheless, we provided a validated protocol to perform such genetic association analyses. Currently, under investigation we plan to identify if these genetic findings are consistent with the levels and function of the proteins being produced.
It should be noted that there are some of limitations in this study. First, the strongest finding in this pilot study is that there is an association of prolonged recovery between carrying the LL GRIN2A (GT)n VNTR in the promoter region. This novel finding is based on using saliva samples obtained following a standardized concussion assessment, in which days of recovery were followed prospectively. The limitations of this study include that KCNH2 was not in Hardy–Weinberg equilibrium; therefore, we did not perform further analysis with this gene. Our small population is likely the reason why KCNH2 was not in Hardy–Weinberg equilibrium. A small population can cause a random change in allele frequencies, and we did have a lower frequency of the less common VNTRs within KCNH2 (i.e., 10, 13, 15, 16). However, GRIK1 was within Hardy–Weinberg equilibrium and the range of (GT)n genotypes was similar within our population, and was deemed that this microsatellite may not be variable in this population (i.e., everyone has some number of repeats). However, it should be noted that this was the first time that either of these genes were genotyped for the (GT)n VNTR within the promoter region. Lastly, given the hypothesis-driven nature of this research endeavor, the number of concussed athletes is relatively low, but despite this limitation the authors were able to find significance, although we were not able to adjust for multiple corrections. Future studies should incorporate a larger cohort with equal demographics being represented (e.g., age, sex, race) within each recovery group to identify potential genetic associations to concussion risk and prolonged recovery.
With these limitations noted, information regarding GRIN2A and NEFL genotype may guide future therapeutic agents, allowing stratification of patients with different concussion recovery points, which could be attributed to altered proteins or protein levels that disrupt neuronal recovery. Lastly, the prospective genotyping of athletes could help medical professionals improve the monitoring and managing of athletes who experience concussion injuries, which would individualize and enhance the patient care that they are able to provide.
Footnotes
Acknowledgments
We thank all of the participants, universities, and medical professionals for their time and effort given to our study. We also thank Dr. Evgeny Krynetskiy for direction and expertise as well as the Jayne Haines Center for Pharmacogenomics for allowing us to use the laboratory space and equipment.
Funding Information
No funding was received to perform this study.
Author Disclosure Statement
No competing financial interests exist.