
Abstract
Early decompression of the traumatically injured and persistently compressed spinal cord is intuitively beneficial for neurological outcome. Despite considerable pre-clinical evidence of a neurological benefit to early decompression, the effect of early surgical decompression in clinical spinal cord injury (SCI) remains less clear. The discrepancy between pre-clinical and clinical results may be due to differences between the biomechanical variables used in pre-clinical animal models and the biomechanical conditions occurring in clinical injuries. These pre-clinical variables include region of spinal cord, velocity of impact, and injury mechanism.
In this study, the effect of velocity and duration of residual compression on injury severity were evaluated using a novel, rodent model of cervical dislocation SCI. Fifty-two male Sprague-Dawley rats were included in five groups: two timings of decompression (24 min, 240 min), two velocities (10 mm/sec, 500 mm/sec), and a sham group. All injuries involved a 1.45-mm dorsal dislocation of the C6 vertebra relative to C5 with subsequent residual compression of 0.8 mm. Animals were evaluated for motor function using the Martinez open field, grip strength, and grooming tests for 6 weeks post-injury. Immunohistochemistry and histology following sacrifice were conducted with counts for NeuN- and choline acetyltransferase (ChAT)-positive neurons, and length of cavitation. Behavioral testing and histological analysis revealed that injuries induced by the high velocity were consistently more severe than those induced by the low velocity, with behavioral correlations ranging between 0.46 and 0.58 (p < 0.05). Longer duration of residual compression did not produce significantly more severe injuries as measured by functional tests and histology. These findings demonstrate that the velocity of the initial traumatic impact may be a more important factor than duration of residual compression in determining SCI severity in a dislocation model of SCI.
Introduction
Traumatic spinal cord injury (SCI) is a life-altering condition, disrupting the central nervous system and often causing irreversible functional deficits. 1 The societal effects of this condition measure in the billions of dollars per year in Canada with more than 1700 new cases occurring annually. 2,3 Acute SCI most commonly occurs from damage to the spine when vertebrae and soft tissue dynamically impinge upon the spinal cord. 4 Following the initial injury event, a certain level of spinal cord deformation can remain and is typically referred to as residual compression. Residual compression is often treated by surgical decompression, which involves removing the impingement on the cord. 5 The effect that the timing of surgical decompression has on functional outcomes is a clinically controversial question. 6 –8 The most extensive clinical study to examine the timing of surgical decompression in cervical SCI reported that patients treated with early decompression (within 24 h of injury) were more likely to achieve two or more grades of improvement from their baseline American Spinal Cord Association (AIS) grade. However, even in this study, the majority of patients who underwent early decompression did not improve. In particular, for those with the most severe injuries (AIS Impairment Scale grade A), AIS grade improvement was observed in 43% with early decompression, and 37% with late decompression. This highlights the fact that although early surgical decompression seems to be biologically rational, it is unclear which patients will actually benefit from this, particularly when the injury is severe.
Within the pre-clinical literature on the effects of timing of decompression, there is strong evidence for the benefits of early decompression. For example, studies have been conducted with differing means of compressing the cord, including static weights, balloon catheters, shims, pistons, ties, and aneurism clips. 9,10 These injuries were primarily conducted statically or at low velocity, ignoring the high-velocity nature of most clinical injuries. Although high-velocity studies have been conducted, using a shim placed within the spinal canal following contusion, the methods used require a brief decompression of the cord between dynamic and residual compression, a potentially sensitive acute time period for secondary injury mechanisms. 11
Pre-clinical studies investigating time of residual compression have observed statistically significant differences between early and late decompression ranging from seconds to multiple days depending on the methods and severity of the cord compression. 9 Because of the many challenges in translating the findings of animal models to clinical practice, determining clinically relevant decompression times is not feasible with animal studies, but rather the effect and interaction of variables in an SCI can be demonstrated. To date, there are no pre-clinical studies examining the effect of timing of decompression using a dislocation model of injury. This is somewhat surprising given that fracture-dislocation injury is one of the most common clinical injury patterns in traumatic SCI. 12 Additionally, most injuries occur in the cervical region of the spine, yet this area is often neglected in pre-clinical research. 13,14 Evidence suggests that the progression of damage in the cervical region occurs at a quicker rate than the thoracic, which is commonly used in pre-clinical models. 15 This highlights the need for clinically relevant studies to ensure that results can better translate to clinical findings.
The effect of velocity on injury outcomes is a relevant factor as there is potentially a difference in the velocity of spinal cord impact between two of the most common causes of SCI: motor vehicle accidents and falls from a standing height. 5 Previous studies have demonstrated histological and behavioral differences in contusion injuries conducted between 80 mm/sec and 800 mm/sec and between 3 mm/sec and 300 mm/sec, in addition to correlations with somatosensory evoked potential (SSEP) recovery between 1500 mm/sec and 6000 mm/sec. 16 –18 These results are intuitive as the spinal cord is a viscoelastic material, meaning that a higher velocity will result in greater forces and energy transfer to the cord. Velocity has also been shown to have interaction effects with depth of compression, indicating the importance of studying more than one biomechanical variable at a time to control for confounding effects. 17
The only study examining the effects of velocity in a dislocation model showed no effect of increasing velocity between 100 mm/sec and 250 mm/sec. 19 The counter-intuitive results from this study involving velocity, and the lack of residual compression studies using a dislocation injury mechanism, represent a real gap in knowledge surrounding which factors most affect neurological recovery after SCI. The motivation for this study is to understand the effects of both duration of residual compression and velocity of injury in a clinically relevant, cervical dislocation model of SCI.
Methods
Overview
All experimental procedures were approved by the University of British Columbia's Animal Care Committee (Vancouver, BC, Canada) in accord with the guidelines published by the Canadian Council on Animal Care. Fifty-two male Sprague-Dawley rats between 325 and 350 g were split between four injury groups and one surgical sham group. Each group was injured with a maximum dislocation velocity of either 500 mm/sec or 10 mm/sec with residual compression held for either 240 min (n = 12 for each velocity) or 24 min (n = 11 for each velocity). These parameters were determined from preliminary tests conducted prior to beginning this study. 20 Sham animals (n = 6) had the complete surgical procedure including a pre-load added, but no dislocation was completed.
Behavioral assessments
Animals arrived 2 weeks prior to injury and were acclimated to the facility and the behavioral tests. The Martinez locomotor rating scale was used to assess forelimb and hindlimb locomotor functions using blinded (to the group assignment) observers in an open-field environment. 21 This test characterizes specific limb movements in addition to overall gait quality, giving each rat a total score out of 40. The grip strength test was used to measure forepaw maximum grip strength as the animal was pulled away from a force-sensing bar. 22 Each grip strength measure was achieved by taking the maximum value of three consecutive trials to account for rodent motivation and variance. The grooming test was used to assess forelimb range of motion and was filmed and later scored by a blinded observer. 23 Scores were given for highest paw placements during grooming, with a 1 for each paw to reach the tip of the nose and a 5 for each paw to reach behind the ear. All tests were performed on the same days, beginning on Day 7 and continuing weekly, with the grip strength test always performed last and the Martinez locomotor rating scale being used additionally on Day 4 post-injury. Prior to injury, all animals were assessed three times on the Martinez locomotor rating scale, requiring a perfect score to be included in the study. In addition, animals were tested twice for grip strength to acclimate and record a baseline assessment.
Injury production
Rats were anesthetized to a surgical plane of anesthesia with 4%/L O2 inhalational isoflurane, which was then maintained between 1 and 2% throughout the procedure to ensure no pain, determined by clinical signs of nociceptive reflexes. The animals were then given a subcutaneous injection of buprenorphine (0.05 mg/kg) and 5 mL of lactated Ringer's solution, which was continued at a minimum of 10 mL/kg/h throughout surgery. In addition, bupivacaine was administered as a local anesthetic via a subcutaneous line block (0.5 mL at 0.25 mg/mL) and preventative enrofloxacin was given (10 mg/kg) subcutaneously. Corneal ointment was applied to the animals' eyes to prevent drying. The animals' temperature was maintained between 36.5 and 37.5°C throughout using a rectal temperature probe coupled to an electric heating pad, and they were monitored with a pulse oximeter for blood oxygenation and heart rate.
Under anesthesia, the dorsal side was shaved in a region surrounding the cervical spine, and then prepared using aseptic technique with 2% chlorhexidine disinfectant and 70% ethanol. An incision was made in the skin above the cervical spine, and the muscle layers were separated along the midline for spinal column exposure. A facetectomy was performed between C5 and C6, whereby the facet joint was removed with a surgical drill to allow for vertebral dislocation. Vertebral clamps were attached as described previously, with the rostral clamp on C4 and C5 and the caudal clamp on C6 and C7. 24
The injuries were conducted according to the assigned group, high velocity: 240 min (n = 12), high velocity: 24 min (n = 11), low velocity: 240 min (n = 12), low velocity: 24 min (n = 11), and sham (n = 6). The UBC multi-mechanism injury device has been previously described and consists of an electromagnetic actuator, custom stereotaxic frame, and implantable injury clamps, which also act as a spinal fixation device. 20 The injury clamps attached to the electromagnetic actuator and an initial pre-load of 2 N was applied for consistent starting conditions. Injury group animals had dislocations performed by moving the caudal clamp dorsally and holding it in residual compression for the assigned period of time before the dislocation was reduced and the spinal column fixated with the injury clamps implanted. This dorsal displacement of the caudal vertebrae simulates the typical ventral displacement of the rostral vertebrae in a fracture dislocation. The dynamic injury depth was 1.45 mm and the residual compression was held at 0.8 mm. This injury was intended to produce a moderate SCI and the residual compression depth was chosen to represent the static deformation following dynamic injury. These parameters were based on a previous study using electrophysiology, which found that 0.8 mm compression depth was sufficient to reduce SSEPs by at least 50% in most animals. 20 All animals remained on isoflurane for 4 h following injury (or sham) as a control for possible anesthetic effects.
Animals continued to receive buprenorphine and lactated Ringer's solution three times per day for 3 days post-surgery and enrofloxacin (5mg/kg) for 4 days post-injury. Animals were monitored closely for signs of medical complications, including bladder dysfunction (in which case expressions were performed), and were kept for 6 weeks post-injury with weekly behavioral tests.
Histology
After 6 weeks of survival post-injury, the rats were euthanized to harvest tissue for analysis. The animals were given an anesthetic overdose of 5% chloral hydrate (1 mL/100 g) before a cardiac perfusion of phosphate-buffered saline was given followed by 4% paraformaldehyde. After perfusion, the cervical region of the spinal cord was harvested and post-fixed overnight in paraformaldehyde. The cords were placed in increasing concentrations of sucrose (12%, 18%, 24%) for 24 h each before being frozen in optimal cutting temperature compound (OCTR). They were then cut on a cryostat at a thickness of 20 microns in the transverse plane and separated into 10 sets.
Spinal cords were immunostained by first blocking with normal donkey serum and then staining for ventral horn motorneuron counts, using primary antibodies against choline acetyltransferase (ChAT) (goat host, 1:500, Millipore, Burlington, MA), NeuN for all neurons (mouse host, 1:500, Millipore), and neurofilament-200 (NF-200) (rabbit host, 1:1000, Sigma-Aldrich, St. Louis, MO) in combination with secondary antibodies conjugated with DyLight 488 donkey anti-goat, DyLight 594 donkey anti-mouse, DyLight 647 donkey anti-rabbit (1:200 Jackson ImmunoResearch Laboratories, West Grove, PA), and Hoechst nucleic acid stain (1:1000, Invitrogen, Carlsbad, CA).
All images were taken with a fluorescence microscope (Axioplan 2, Carl Zeiss, Jena, Germany) (10 × objective lens) with brightness and contrast optimized for obtaining cell counts. The injury epicenter was identified through areas of greatest cavitation and lack of NeuN-positive cells. Cell counts were conducted for both NeuN and ChAT in a 600 μm diameter circle in the ventral horn by a blinded observer to indicate total surviving neurons and surviving motor neurons. Cavitated regions were identified by an obvious lack of gray matter tissue (lack of NF200) and the length of continuous rostral/caudal cavitation was measured from the epicenter.
Statistical analysis
Using the Shapiro-Wilks test (p > 0.05), the behavioral and histological data were found to be not normally distributed, requiring the use of non-parametric statistics. Kruskal-Wallis tests (p < 0.05) were run to examine statistical significance of histological data and behavioral data at 1, 4, and 6 weeks. At each time-point a multiple comparisons post hoc test was run to examine between-group differences. A Spearman rank test (p < 0.05) was run at Week 1 and Week 6 on behavioral data, and on cavitation length to examine the correlations between velocity and duration of residual compression with injury severity. The Benjamini-Hochberg method was used to control for multiple comparisons and ensure a false discovery rate below 0.05 for the study.
Results
The velocity and force were measured for each test and are shown in Table 1. The maximum velocities had standard deviations of less than 4% and the peak force was higher for the high-velocity groups than the low.
Summary of Biomechanical and Group Demographic Data
Behavior
Similar trends were observed between groups across all three behavior tests. The high-velocity injury groups demonstrated a decrease in Martinez score, which recovered over time (Fig. 1). This same trend was not seen in the low-velocity groups, which had minimal functional losses. Starting from Day 7, significant differences were observed between high- and low-velocity groups and continued to be seen at Day 28. However, these were no longer significant by Day 42 as animals in the high-velocity group recovered function. No differences were seen between 24-min and 240-min residual compression at either high or low velocity.
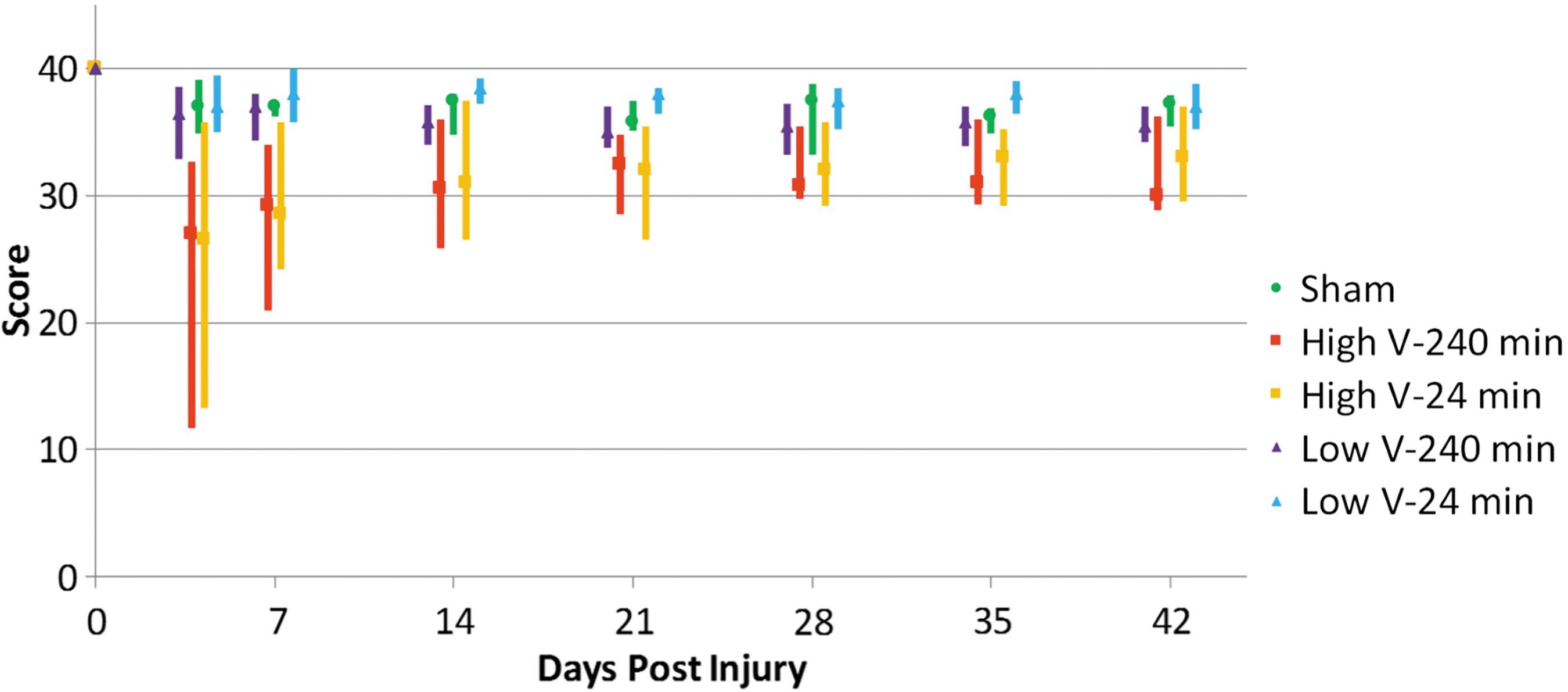
Median Martinez open field scores with quartiles. High-velocity groups have lower median values and greater variance, with significant differences between the low-velocity 24-min group and both high-velocity groups at Days 7 and 28 and between the high- and low-velocity 240-min groups at Day 7, with no significance at Day 42. Color image is available online.
Rats gained grip strength over time in all groups as weeks progressed (Fig. 2). Obvious deficits were seen in high-velocity groups after injury but not in low-velocity groups. Significant differences between high- and low-velocity groups were seen from Day 7 through to Day 42. No differences were seen between 24 min and 240 min of residual compression at either high or low velocity.
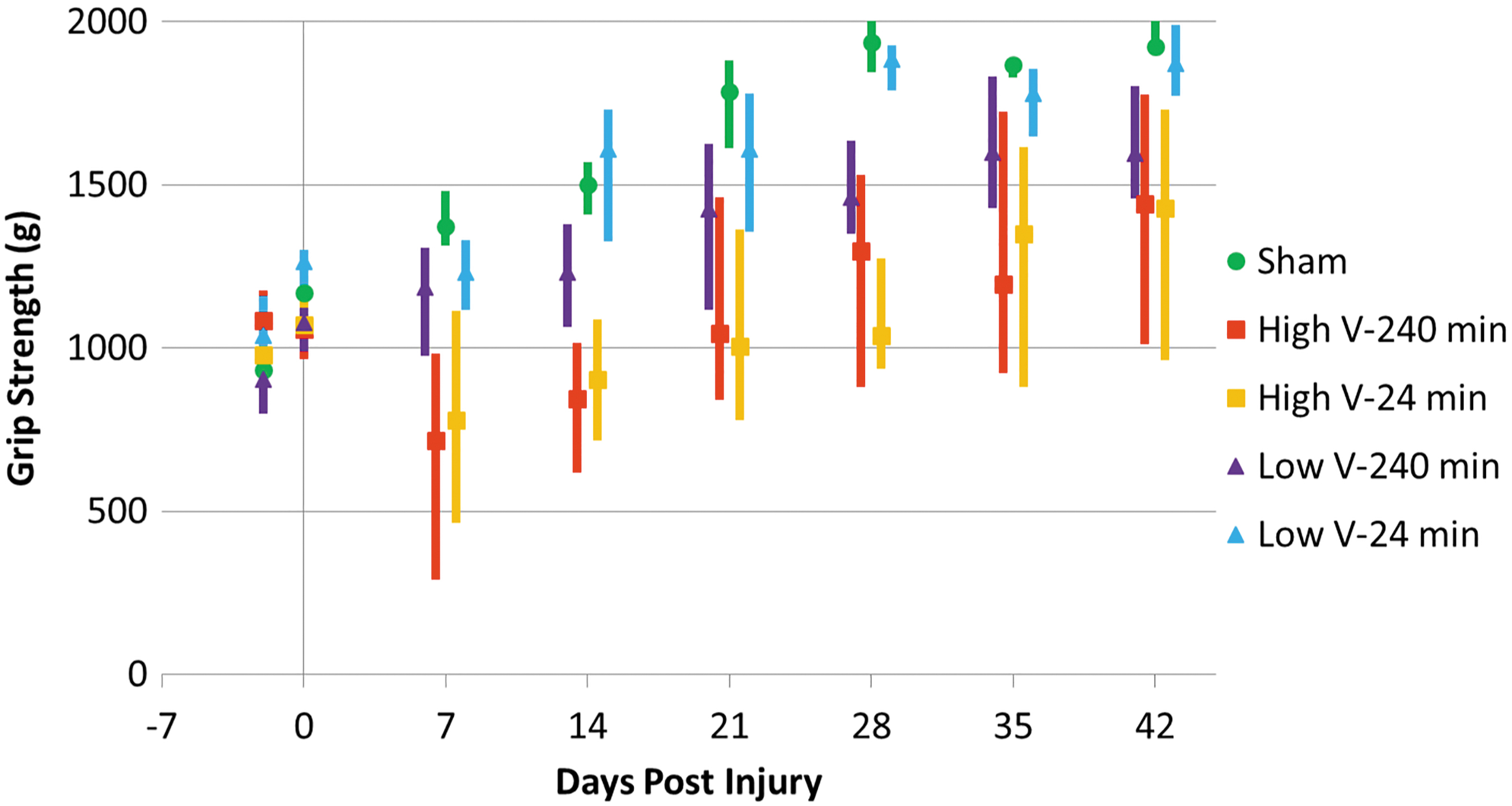
Median grip strength scores with quartiles. High-velocity groups have lower median values and greater variance, with significant differences between both high-velocity groups and the low-velocity 24-min group at Days 7, 28, and 42 and between high- and low-velocity 240-min groups at Day 7. Color image is available online.
In the grooming test, deficits were seen in both high-velocity groups, which did not recover over time (Fig. 3). Once again, significant differences were seen between high- and low-velocity groups, but not between times of residual compression.
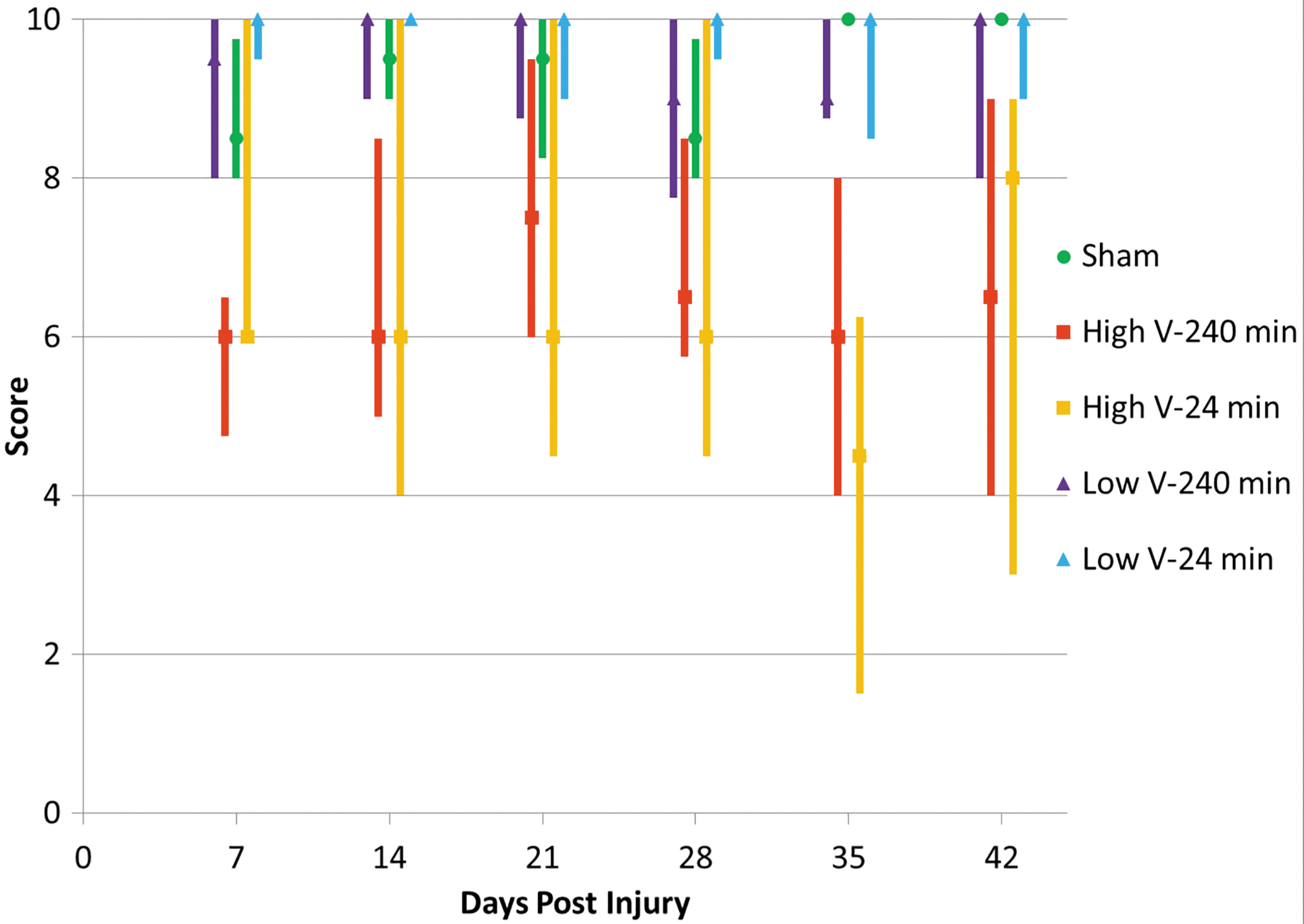
Median grooming scores with quartiles. High-velocity groups have lower median values and greater variances, with significant differences between high-velocity 240-min and both low-velocity groups at Day 7, between both high-velocity groups and the low-velocity 24-min group at Day 28, and between high- and low-velocity 24-min groups at Day 42. Color image is available online.
Table 2 shows all correlations with duration of residual compression were not significant (n.s.), and all velocity correlations were significant (Figs. 4 and 5).
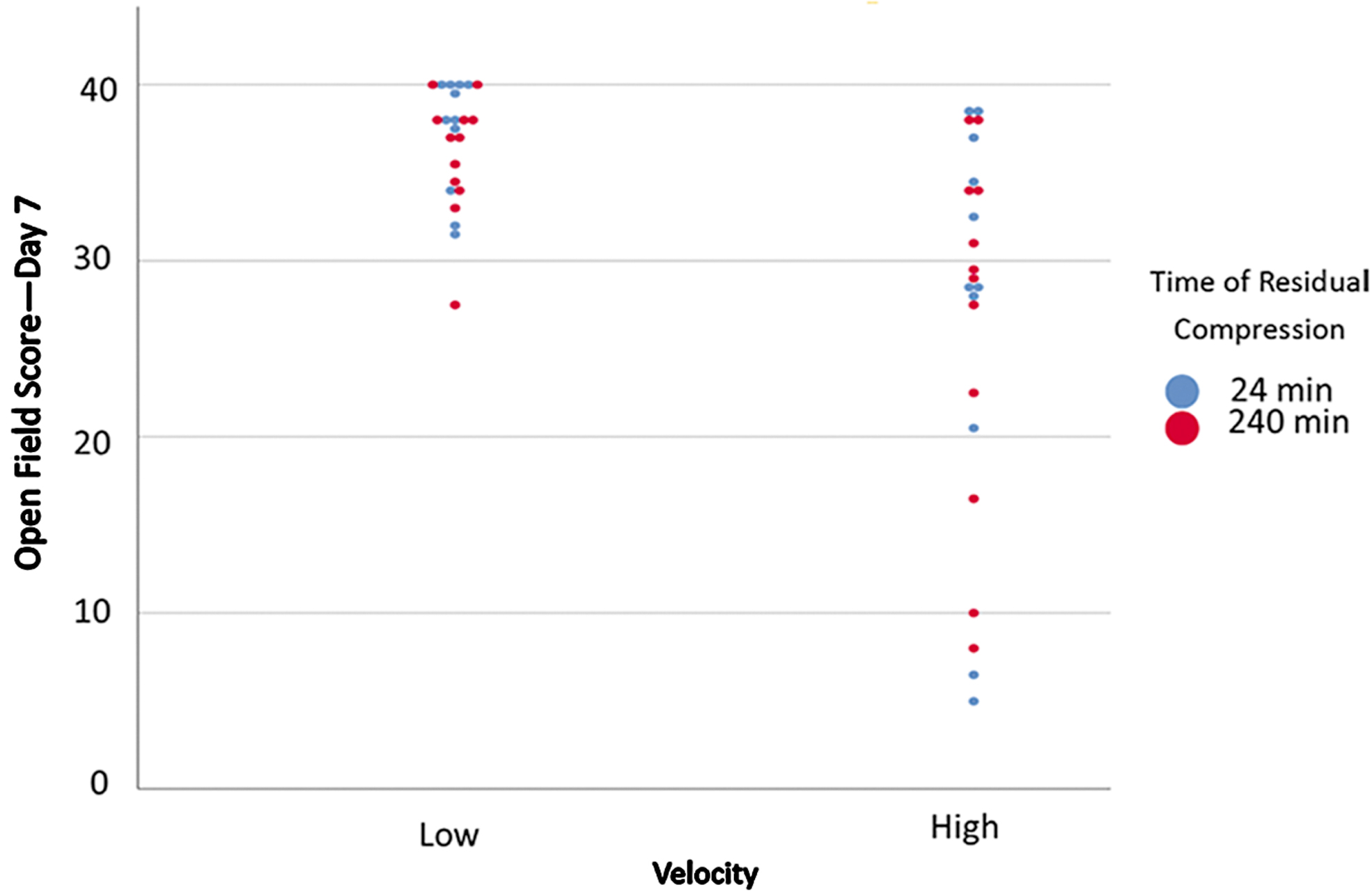
Day 7 Martinez open field score by velocity and time of residual compression. Significant correlation between velocity and score exists, but not between time of residual compression and score. Color image is available online.

Day 42 Martinez open field score by velocity and time of residual compression. Significant correlation between velocity and score exists, but not between time of residual compression and score. Color image is available online.
Spearman Rank Correlations between Velocity, Timing of Residual Compression, and Behavioral Outcomes
n.s., not significant.
Histology
Counts of NeuN-positive cells and ChAT-positive cells showed similar results with all injury groups experiencing a loss of neurons (Fig. 6). Despite consistent differences in mean and median NeuN counts between high- and low-velocity groups, they only reached statistical significance in a subset of locations (Fig. 7). Significant differences between the low-velocity 240-min group and both high-velocity groups were observed at +0.2 mm and with the high-velocity 24-min group at +0.6 mm. ChAT counts showed the same trends; however, no counts reached significance (Fig. 8).
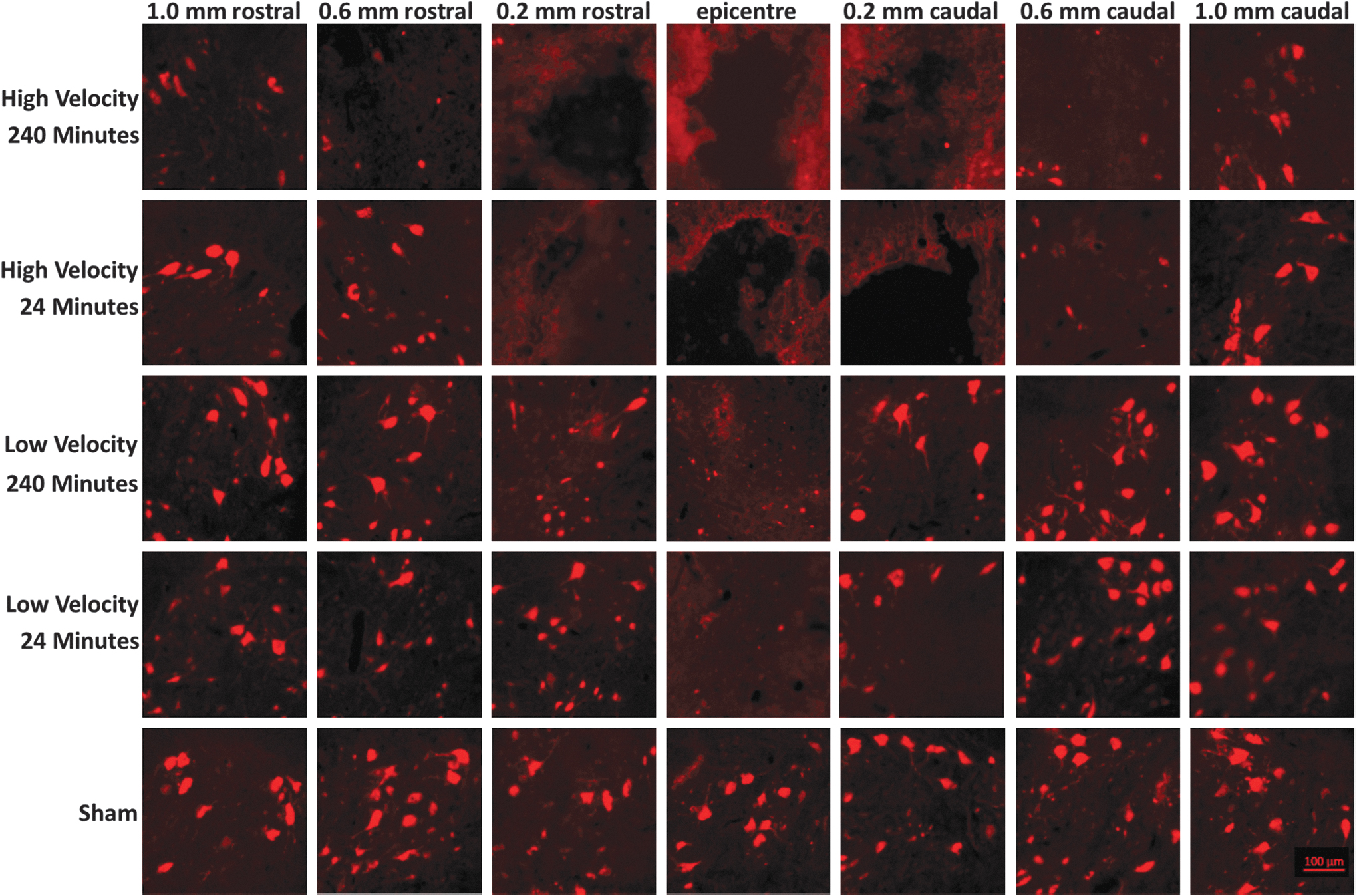
Micrographs of ventral horn neurons stained with NeuN. Representative images in the region used for total neuron counts by injury group. Color image is available online.
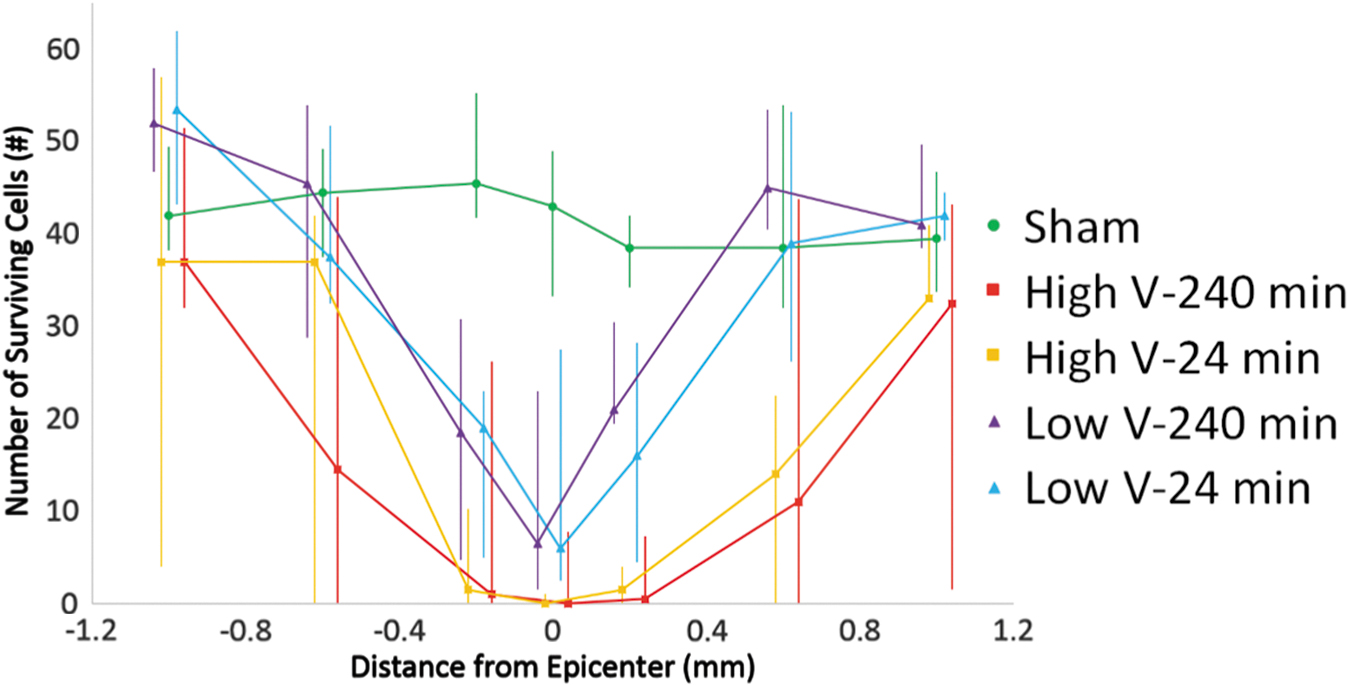
Median NeuN counts with quartiles. High-velocity groups have lower median values, with significant differences between low-velocity 240-min and both high-velocity groups at both +0.2 and +0.6 mm, and between low-velocity 24-min and high-velocity 24-min at +0.6 mm. Color image is available online.
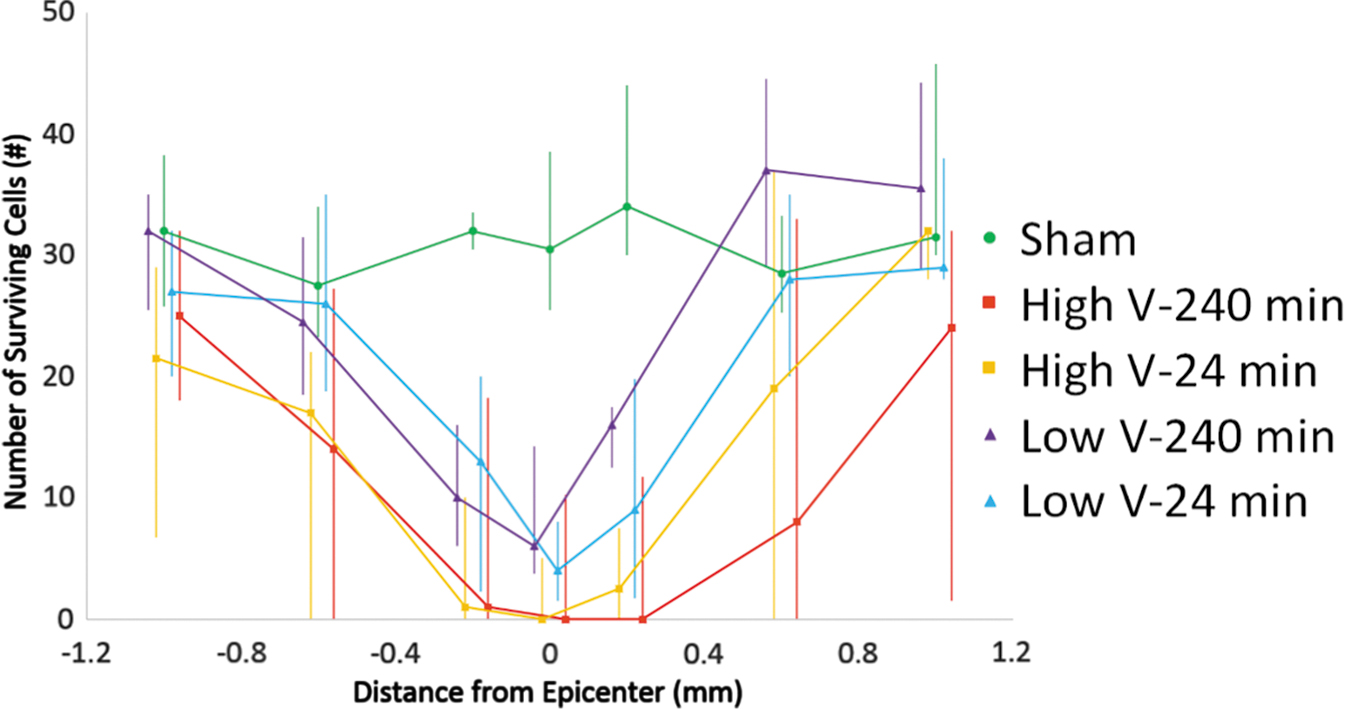
Median choline acetyltransferase (ChAT) counts with quartiles. High-velocity groups have lower median values. Color image is available online.
The extent of cavitation in the rostral-caudal direction was greater in the high-velocity groups than low (Fig. 9). A Spearman correlation coefficient of 0.474 was found between velocity and cavitation length (p < 0.01), whereas no significant correlation was seen between time of residual compression and cavitation length.
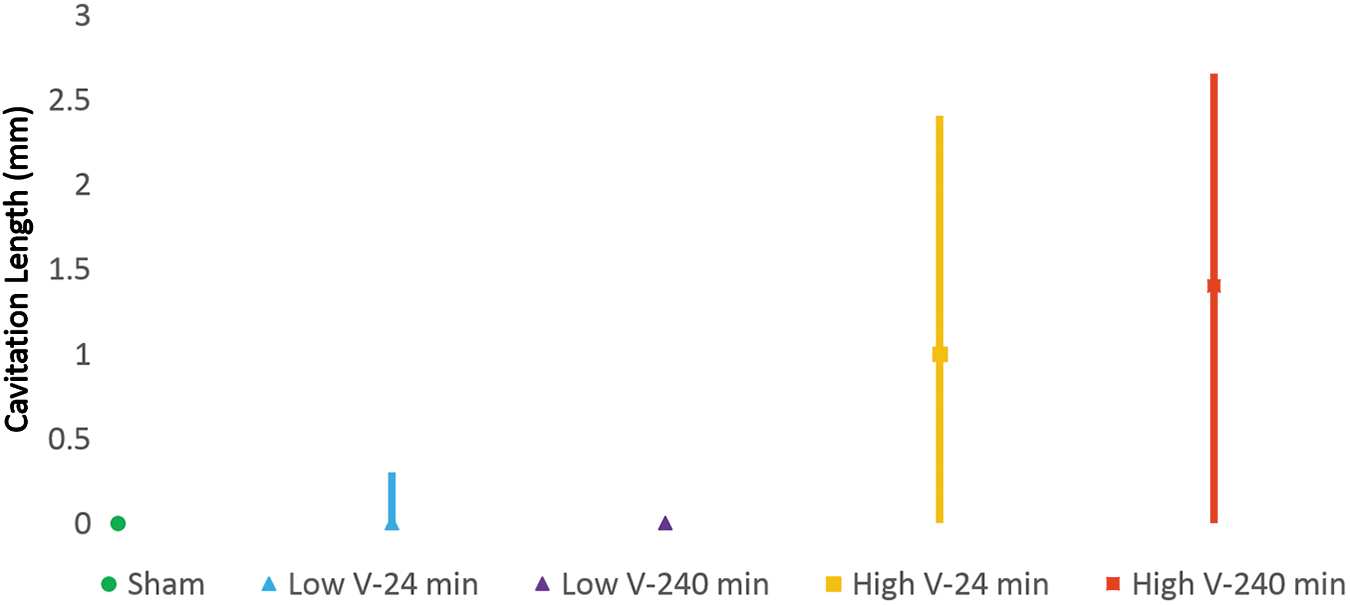
Median cavitated length with quartiles. High-velocity groups have greater cavitated length than low-velocity groups. Significant differences are seen between the low- and high-velocity 240-min groups. Color image is available online.
Significant correlations (p < 0.01) between cavitation length and open field score at Day 7 and Day 42 were observed (Figs. 10 and 11).
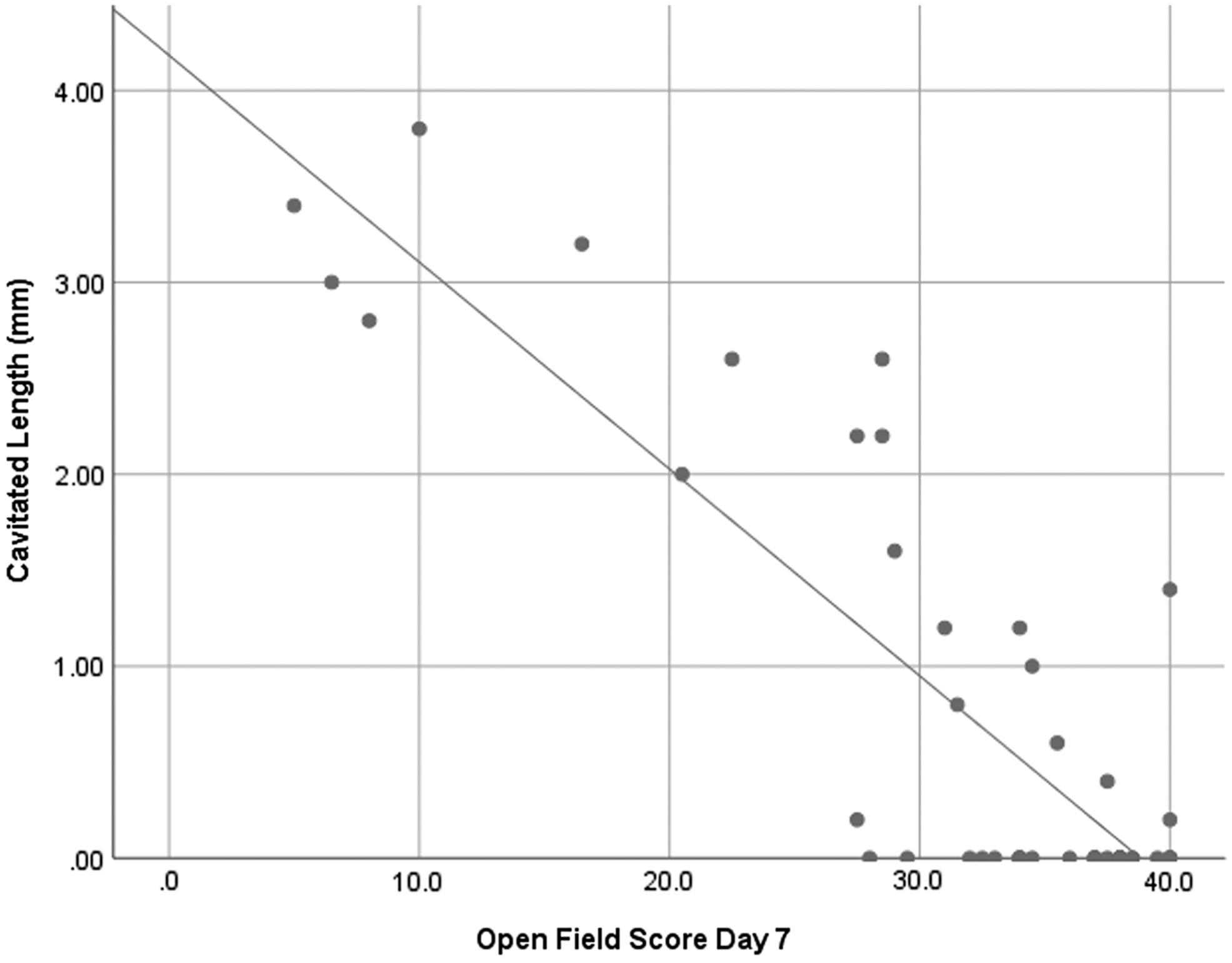
Cavitated length versus open field score Day 7. A significant correlation was seen between the two factors and a linear fit R2 value of 0.736 was found.
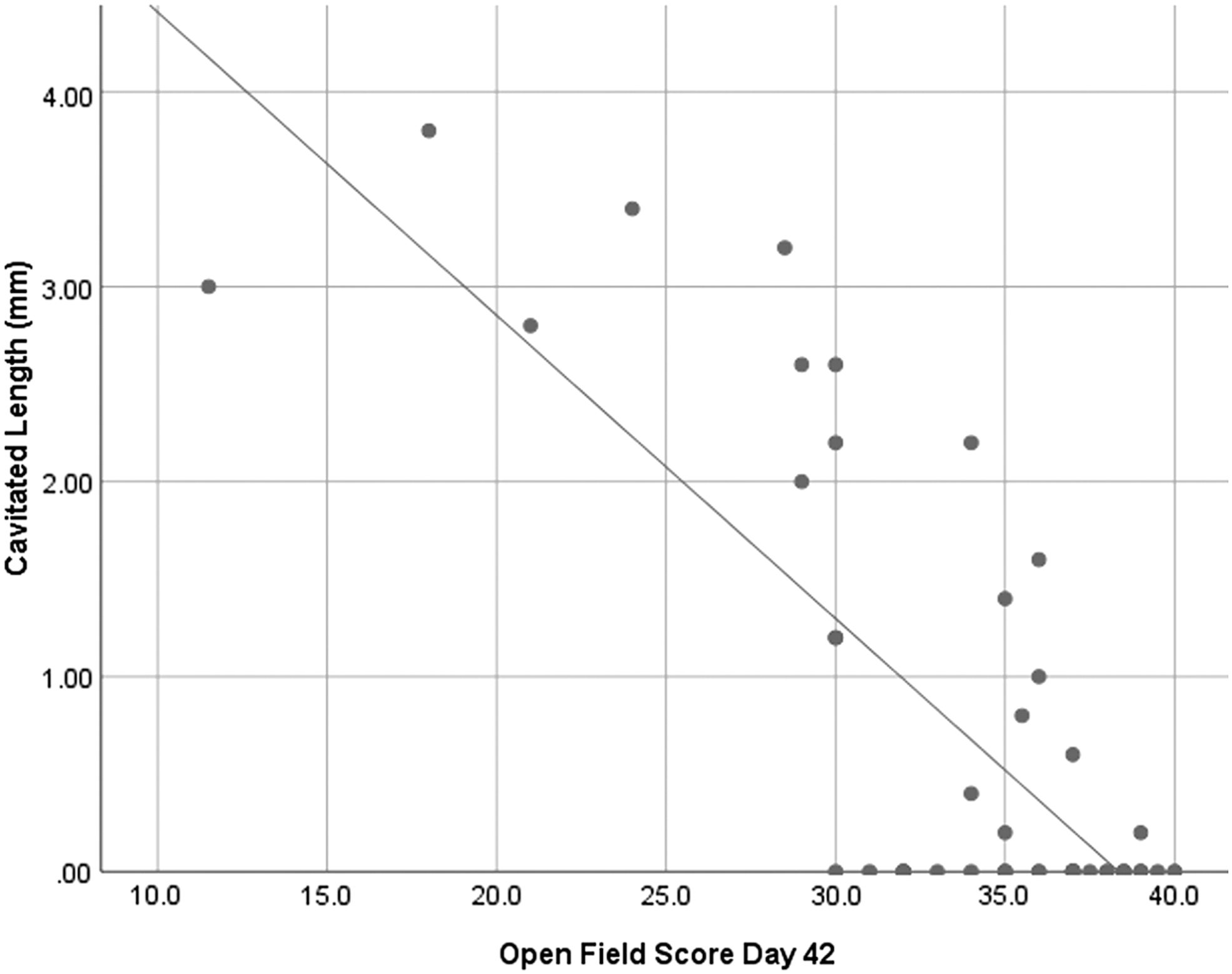
Cavitated length versus open field score Day 42. A significant correlation was seen between the two factors and a linear fit R2 value of 0.607 was found.
Discussion
This study is the first to investigate the effect of timing of decompression in a dislocation model using clinically relevant biomechanical variables. This study showed both the significant influence of velocity and the importance of using high-velocity pre-clinical models to better replicate clinical injury conditions. The impacts of increasing velocity were seen in all three behavior tests, NeuN counts, and cavitation length, and correlations remained even at 42 days post-injury.
The cavitation length was highly correlated with behavior tests, more so than any neuron count with behavior. This shows the importance of the rostral-caudal extent of gray matter injury to functional outcomes and the usefulness of the cavitation length measure. Previous dislocation studies have demonstrated significant white matter loss, which although not quantified here, is also important in explaining functional losses.
Effect of velocity
The variety of behavioral and histological tests showed both the behavioral deficits incurred with greater velocity, but also confirmed that injury was still occurring at lower velocities.
These results differ from a previous dislocation-velocity study in which lateral dislocations in the thoracic spine were performed at velocities between 100 mm/sec and 250 mm/sec before sacrificing at 5 h post-injury. 19 No differences between groups were reported. A likely explanation for this discrepancy is the magnitude of velocities tested. Given the variability of outcomes in dislocation models, a larger difference in velocities may be required to find significance.
In a contusion model, behavioral differences have been shown between 800 mm/sec and both 8 mm/sec and 80 mm/sec, but not between 8 mm/sec and 80 mm/sec. These results indicate that a velocity threshold may exist at which additional damage is caused. 17
Increasing velocity causing further injury to the spinal cord is consistent with our understanding of tissue mechanics, as a greater amount of energy can be transferred to the cord, causing damage. This also matches with clinical findings, which have shown velocity to be an important predictive factor in recovery following SCI. 25 However, understanding whether the threshold and interaction of velocity and depth of injury exist in dislocation injuries similar to contusions would require further study of these factors.
Effect of time of residual compression
No behavioral or histological differences were seen between animals decompressed at 240 min and 24 min post-injury at either high or low velocity. These results differ from most pre-clinical studies, which have generally seen an influence of timing of decompression, especially in low-velocity injury models. 9 Although there were (non-significant) trends at a small number of individual measures, they were not consistently seen and not always in the same direction. At the vast majority of measures, differences were not apparent. Of course, we may not have observed differences in decompression time due to low statistical power and thus further studies with larger sample sizes may be worthwhile.
Compared with other residual compression studies, there are many important factors varying between studies including: mechanism of injury, depth of dynamic injury, depth of residual compression, region of cord, velocity of injury, time between injury and sacrifice (for histology results), and outcome measures. A recent meta-analysis examined some of these factors in pre-clinical studies, attempting to determine effect sizes for each. 9 One of the most significant factors found was compressive pressure, which, depending on the injury model, likely correlates with depth of residual compression. In this analysis, a power law distribution was seen between compressive pressure and time to severe neurological injury. This indicates that using a greater depth of residual compression in our study may have shown differences between durations of residual compression, as the increased compressive pressure would lead to a shorter time to severe neurological injury. Although our compression depths were validated using SSEP, this was the first study to examine the behavioral and histological outcomes using these values, and further study at differing depths would serve to elucidate the impact of this variable in a dislocation injury model.
When comparing between studies, there exists a trend by which models involving static deformation following dynamic injury employ lower pressures and thus see longer compression times required for neurological injury. 9 These models better represent the biomechanics of the clinical injury scenario, which typically involve a dynamic compression of the cord, immediately followed by static residual compression. 12 This concept of pressure may also explain some of the variability found in our model, as differing vertebral morphology may result in variations in pressure between animals despite the same dislocation displacement. Compressive pressure has not been measured in our rodent dislocation injury, thus preventing a direct comparison of values.
The region of cord is relevant as cervical and high-thoracic pre-clinical studies in particular have seen less benefits from early decompression indicating that duration of residual compression may be more important in mid- and low-thoracic injuries; however, few cervical studies exist. 9,26,27 In a cervical clip compression model, early decompression was only beneficial when lighter injuries were produced, and had no effect when a higher force was applied to the cord, once again highlighting the importance of interaction effects between biomechanical variables. 27 The lack of studies examining injuries to the cervical region of the spinal cord is a significant gap as this is the most common region for clinical injuries, and may respond differently due to its unique anatomy, including the increased vasculature due to greater neuronal pools and more gray matter. 14 In addition, although both cervical and thoracic injuries affect both long tracts and motor neurons, local motor neuron disruption in thoracic injuries is not manifest clinically in the lower extremities, whereas it is in the upper extremities for cervical injuries. Thus, any impacts of residual compression on motor neurons and their related interneuronal networks will have distinct outcomes between a cervical and thoracic model.
Another important variable is the method of compression. When grouped by the device causing the injury, significant differences were seen on effect size of timing of residual compression. 9 These results provide motivation for understanding how different methods of producing injury impact the effects of residual compression.
In addition to being the first dislocation study with residual compression, this study is also the first closed-column model with residual compression. These methods eliminate many of the variables that other models have, such as foreign objects interacting with the cord, laminectomies performed, and non-clinically relevant geometry of the impactor. Another differentiating factor of the dislocation model is the use of a constant displacement rather than a constant force, which is common in other studies using clips or weights. A constant force model will continue to increase displacement on the cord over time, a phenomenon not likely occurring in clinical scenarios. A significant limitation of this dislocation, and all closed-column models, is the inability to directly measure or observe the deformation of the cord. Because of this limitation, inevitable variations in rat spinal column and spinal cord morphology will result in differing levels of cord compression.
Similar to previous studies, grooming behavioral deficits do not recover to the same extent as other behavioral tasks following dislocation injury. 28 This has been ascribed to motor neuron death between C5 and C6, which is largely responsible for forelimb range of motion, whereas forepaw grip strength is controlled over a larger rostral-caudal extent of the spinal cord. 29 This explanation is consistent with our results as surviving motor neurons would allow for some recovery of gross movement; however, the loss of gray matter between C5 and C6 may result in permanent deficits to forepaw range of motion. As seen in the sham animals, grip strength also increased as the animals grew older and gained weight over time, thus not all gains following injury can be attributed to neurological recovery.
The variability seen in dislocation studies has been well documented and may obscure significant differences between groups; however, trends are still clearly seen and the effects of each variable are visible. 24,28,30,31 A possible explanation for the variability seen is the inherent differences in animal spinal canal morphology. Differing canal and spinal cord dimensions would cause the same vertebral displacement to cause varying degrees of both dynamic shear force and static cord compression following. In addition, although the 2 N pre-load applied prior to injury is vital to defining a consistent starting location for vertebral displacement, it may cause minor differences based on disc material properties and morphology.
A challenge exists when attempting to compare timing of compression between rodents and humans, ensuring that the values used are of clinical relevance. Although current clinical guidelines recommend decompression to occur within 24 h of SCI, this may not be an appropriate timescale to use in a rat based on different rates of biological processes. 32 Taking processes such as metabolic rate and messenger RNA (mRNA)/protein turnover into account estimates the equivalent of 24 h in a human at between 2.5 h and 9.6 h in a rat. 33 These indicate that the values used in our study of 4 h and 0.4 h are likely relevant to demonstrate overarching differences. We would acknowledge, however, the speculative nature of these extrapolations of biological time courses along the lifespan between human and rodents may no longer apply for acute tissue-injury responses. This study was not designed to determine timing windows for clinical decompression, but rather to determine the effect of residual compression in a dislocation model. As the first study to use a dislocation injury model with residual compression, there was significant concern regarding rodent survival in a longer time of compression. Thus, the 4 h choice was a balance of feasibility and relevance, understanding that previous studies have seen significant results using compression durations much shorter than 4 h.
Conclusion
The model used in this study attempts to mimic the biomechanical variables of a clinical dislocation injury more closely than previous studies. Importantly, this study used a clinically relevant velocity, involved immediate residual compression, and the injury was caused by vertebral dislocation. Across behavioral and histological tests, velocity was seen to have a significant influence on injury severity, whereas timing of residual compression did not, even in the low-velocity injury. Identifying velocity as a significant factor in SCI severity, both initially, and weeks after injury provides further incentive for this injury parameter to be taken into account clinically. Timing of residual compression was not significant in this study, indicating that, perhaps, cervical dislocations are not as sensitive to this variable as other injuries and highlighting an area for further research.
Footnotes
Acknowledgments
We wish to acknowledge the technical assistance of Caron Fournier, Nicole Janzen, Elizabeth Raffaele, Aaron Moulson, and Alexander Burden in this project.
Funding Information
The Canadian Institutes for Health Research (CIHR) and the Blusson Integrated Cures Partnership provided funding support (CIHR Grant #133574).
Author Disclosure Statement
No competing financial interests exist.