
Abstract
Chronic symptoms after mild traumatic brain injury (mTBI) are common among veterans and service members, and represent a significant source of morbidity, with those who sustain multiple mTBIs at greatest risk. Exosomal micro-RNAs (miRNAs), mediators of intercellular communication, may be involved in chronic TBI symptom persistence. Exosomal miRNA (exomiR) was extracted from 153 participants enrolled in the Chronic Effect of Neurotrauma Consortium (CENC) longitudinal study (no TBI, n = 35; ≥ 3 mTBIs (rTBI), n = 45; 1–2 mTBIs, n = 73). Analyses were performed with nCounter® Human miRNA Expression Panels and Ingenuity Pathway Analysis (IPA) for identification of gene networks associated with TBI. Generalized linear models were used to analyze the predictive value of exomiR dysregulation and remote neurobehavioral symptoms. Compared with controls, there were 17 dysregulated exomiRs in the entire mTBI group and 32 dysregulated exomiRs in the rTBI group. Two miRNAs, hsa-miR-139-5p and hsa-miR-18a-5p, were significantly differentially expressed in the rTBI and 1–2 mTBI groups. IPA analyses showed that these dysregulated exomiRs correlated with pathways of inflammatory regulation, neurological disease, and cell development. Within the rTBI group, exomiRs correlated with gene activity for hub-genes of tumor protein TP53, insulin-like growth factor 1 receptor, and transforming growth factor beta. TBI history and neurobehavioral symptom survey scores negatively and significantly correlated with hsa-miR-103a-3p expression. Participants with remote mTBI have distinct exomiR profiles, which are significantly linked to inflammatory and neuronal repair pathways. These profiles suggest that analysis of exosomal miRNA expression may provide novel insights into the underlying pathobiology of chronic TBI symptom persistence.
Introduction
The long-term consequences of mild traumatic brain injury (mTBI) and especially repetitive mTBI (≥ 3, rTBI) are not yet fully understood. 1 –3 Despite increasing epidemiological evidence of a link between mTBI and late-life dementia, the mechanisms underlying this association have not been established. 3 –6 Recently, persistent or secondary central nervous system (CNS) processes involving blood–brain barrier (BBB) disruption, oxidative stress, and neuroinflammation have been implicated in the onset and maintenance of chronic persistent TBI symptoms and remote emergent neurodegeneration. 1,7,8 Increasing evidence suggests that CNS-derived exosomes modulate secondary injury processes through the transfer of macromolecules intracellularly. 9 In support of this, pre-clinical studies show that specific micro-RNAs (miRNAs) found within extracellular vesicles (EVs) are linked to neuronal injury and neuroinflammation and are thought to contribute to processes underlying secondary injury and repair, which may relate to chronic symptoms of TBI. 10 Exosomes are small (30–100 nm) extracellular vesicles that diffuse freely across the lipid bilayer. 11 Exosomes form when an endocytic multivesicular body (MVB) fuses with the plasma membrane, and the MVB's contents are exocytosed. 12 After release into the extracellular milieu, exosomes fuse with nearby cells, where their cargo (e.g., messenger RNA (mRNA), miRNA, enzymes, proteins) are released to the recipient cell, changing RNA activity from one cell to the next, thereby orchestrating cellular responses. 13 The transfer of exosomal miRNAs is of particular interest in the context of TBI, as their transfer from one type of cell to another provides a mechanism by which cell type may regulate not only protein translation in neighboring cells but inflammatory processes as well. 9
Exosomal miRNAs have been implicated in the orchestration and maintenance of persistent inflammatory responses, 14 but no studies have been published to date in chronic mTBI, particularly in the remote period years after injury, although there have been some miRNA studies in sporadic neurodegenerative disorders. Several recent studies have emphasized the role of EV-associated miRNAs in immune activation and regulation. 14,15 In a pre-clinical study of acute TBI, Harrison and coworkers reported differential expression of EV-associated miR-212, mi-21, miR-146, miR-7a, and miR-7b. 10 In a clinical study of acute TBI, Patz and coworkers identified 81 differentially expressed EV-miRNAs isolated from cerebrospinal fluid (CSF) of severe TBI patients. 16 Among the 81 EV-miRNAs identified, 23 were previously shown to be differentially regulated in pre-clinical TBI studies. 16 –18 Notably, miR-9 and miR-451 were among the 81 miRNAs identified, both of which have previously been shown to play a role in the regulation of neurological function. 10,19 To determine the role of miRNAs in chronic symptom persistence years following single and repetitive TBIs, and to gain clinical and mechanistic insights into the role of miRNA in chronic symptoms and outcomes years after mTBI, we evaluated miRNA isolated from peripherally circulating exosomes of military personnel who sustained a variable number of remote mTBIs, from none to many.
Methods
Study design
All samples included in this analysis came from participants enrolled in the Chronic Effects of Neurotrauma Consortium (CENC) Multicenter Prospective Longitudinal Study (PLS), a multi-site observational study of mTBI late effects among post-9/11 era veterans and service members, which is ongoing in the Long Term Impact of Military Brain Injury Consortium (LIMBIC), with approval by all associated institutional review boards (IRBs). In this analysis, we included samples from 153 participants with full clinical data and associated plasma for exosomal miRNA (exomiR) characterization.
Participants
LIMBIC-CENC PLS participants were recruited from four Veteran Affairs Medical Centers (VAMCs) (Richmond, VA; Tampa, FL; Houston, TX; San Antonio, TX) focusing on post-9/11 era service members and veterans who were combat deployed and were diagnosed with a spectrum of mTBI exposures (ranging from no events to multiple combat-related mTBI). Exclusion criteria included the following: (1) history of moderate or severe TBI as defined by either (a) initial Glasgow Coma Scale <13, (b) coma duration >0.5 h, (c) post-traumatic amnesia (PTA) duration >24 h, or (d) traumatic intracranial lesion on head computerized tomography; or (2) history of (a) major neurological disorder (e.g., spinal cord injury), or (b) major psychiatric disorder (e.g., schizophrenia) with “major” defined as resulting in a significant decrement in functional status or loss of independent living capacity. For these analyses, the primary independent variable was mTBI history as will be determined and measured. In this analysis, we included samples from 153 subjects with full clinical data and associated plasma for exomiRNA characterization.
Determination of TBI
The LIMBIC-CENC PLS assesses participants for lifetime potential concussive events (PCEs) and assigns an mTBI diagnosis for each event based on Department of Defense (DoD)-Veterans Administration (VA) common definition of mTBI, using the Virginia Commonwealth University Retrospective Concussion Diagnostic Interview (VCU rCDI). 20 Using these TBI determinations, participants were classified into various TBI groupings for the following analysis groups: (1) repetitive (≥ 3) mTBIs, (2) 1–2 mTBIs, and (3) controls (no TBI history).
Potential concussive event identification and TBI diagnoses
This study's structured interview process entailed screening for all PCEs during military deployments and across the entire lifetime, including childhood, using a modification of the Ohio State University TBI Identification (OSU TBI-ID) instrument. 21 Each PCE identified was evaluated to determine whether or not it was a true clinical mTBI via a detailed structured interview, the Virginia Commonwealth University retrospective Concussion Diagnostic Interview (VCU rCDI). Each VCU rCDI renders a preliminary TBI diagnosis of either mTBI with LOC/PTA, mTBI without LOC/PTA, or not mTBI, through an embedded algorithm using the structured interview data and based on the DoD/VA common definition of mTBI. Every preliminary TBI diagnosis is reviewed and vetted against the unstructured free text portion of the interview, and against any available medical documents recorded in proximity to the event (i.e., first responder, emergency department, or in-theatre documentation). Using this process, the site principal investigator confirms or overrides every preliminary mTBI diagnosis to yield the final diagnosis. The event is also assessed for TBI severity to ensure eligibility (any severity greater than mild was excluded from this study). If any doubt remains as to TBI diagnosis, the event is adjudicated by a central diagnosis committee consisting of national experts in TBI.
Assessment of post-concussive symptoms
The Neurobehavioral Symptom Inventory (NSI) was used to assess the post-concussive symptom severity. The NSI consists of a 22-item assessment with a three–four factor structure (somatic/sensory, affective, and cognitive); higher total scores indicate more severe symptoms. 22,23
Plasma sample processing and exomiR isolation
Whole blood samples were collected and centrifuged at 3000 rpm for 10 min (4°C) within 2 h of being drawn. Plasma was aliquoted and stored at -80°C until analyzed. Plasma samples were thawed on ice and centrifuged at 3000 rpm for 5 min to pellet and remove circulating cells or debris. ExomiRs were extracted from 400 mL of plasma supernatant by using Qiagen ExoRNeasy Serum Plasma Kits (Qiagen, MD, USA) as per manufacturer's instructions and eluted in 20 μL water. The RNA extract was then cleaned and concentrated using the RNA Clean and Concentrator-5 kit (Zymo Research Corp., CA, USA) and finally eluted in 10 μL of water. The concentration, purity, and integrity of the exomiR product was determined with Bioanalyzer (Agilent, CA, USA) and Qubit (Thermo Fisher Scientific, CA, USA).
ExomiR profiling analysis
Analysis was performed on all samples by using nCounter® Human v3 miRNA Expression Panels (NanoString Technologies, WA, USA), which contains 798 miRNA probes. This is currently the maximum number of probes developed and available for evaluation in humans. Probes for housekeeping genes such as beta-actin (ACTB), beta-2-microglobulin (B2M), glyceraldehyde 3-phosphate dehydrogenase (GAPDH), ribosomal protein L19 (RPL19), and ribosomal protein lateral stalk subunit P0 (RPLP0), as well as for endogenous exomiRs for Arabidopsis thaliana miR159a (ath-miR159a), Caenorhabditis elegans (cel)-miR-248 and miR254, and Oryza sativa (osa)-miR 414 and 442 are incorporated in the code sets and were used for analysis along with positive and negative controls. All hybridizations took place over 18 h, and all counts were obtained at MAX mode, creating 555 images per sample for analysis. Analysis of raw miRNA data was performed using the nSolver analysis software version 4.0 (NanoString technologies). Code count normalization was performed with the geometric median for the 50 highest expressed genes. Genes with a fold change >1.2 and an adjusted p value <0.05 after correction for multiple comparisons were considered differentially expressed. Heatmap and hierarchical cluster analyses were used to identify differentially expressed miRNAs among the TBI groups compared with controls without a history of lifetime TBI.
Network analysis with Ingenuity Pathway Analysis (IPA)
Differentially expressed exomiRNA were investigated by IPA (Ingenuity Systems Inc., CA, USA). Briefly, miRNA outcomes from nCounter analysis and the miRNA target genes defined in miRWalk (
Statistical analysis
Descriptive statistics for all demographic and clinical variables were calculated using SPSS Statistics (IBM SPSS Inc., Chicago, IL, USA; Table 1). Comparisons were made among the three groups using χ 2 tests for categorical and analysis of variance (ANOVA) for continuous variables. P values <0.05 were considered significant after adjusting for multiple comparisons using Bonferroni correction.
Demographics and Clinical Characteristics
Number of general TBIs refers to the number of TBIs caused by non-blast blunt injury. Total number of TBIs refers to the number of general TBIs plus the number of TBIs caused by blast.
Significance between noTBI and rTBI groups (≥3) or noTBI and 1–2 TBI groups (<3). p values < 0.05 are bold.
TBI, traumatic brain injury; SD, standard deviation; GED, general educational development; NSI, neurobehavioral symptom inventory.
nCounter outcomes, and demographic and clinical data were further analyzed with nSolver software (NanoString Technologies, WA, USA), RStudio (Boston, MA, USA), or SPSS v.24 (IBM Corporation, Armonk, NY, USA). A t test (p value) was performed on log2-transformed data for group comparison. The log2 transformation satisfied the t test's requirement of normally distributed data, and a heteroscedastic t test (Welch's) was used on the assumption that variance may not be equally distributed between groups. Multiple comparisons correction was adopted when multiplexed gene expression data sets were involved. Generalized linear models (GLM) were made in order to analyze the predictive value of exomiR dysregulation and persistent neurobehavioral symptoms and outcomes.
Results
Demographic and clinical characteristics
The demographic and clinical characteristics of the 153 LIMBIC-CENC participants included in this analysis are described in Table 1. The mean age of this predominately male (77%) sample was 40 years (± 10.44). Of the 153 participants included in this study, 35 had no history of TBI (control group) whereas 118 participants (mTBI
ExomiR expression profiles
After normalization, the fold change for the plasma exomiRs in the TBI group was calculated using those in the noTBI group as controls. Compared with controls, 17 exomiRs had altered regulation in the TBI group (Table 2). Among them, 1 exomiR was downregulated and 16 exomiRs were upregulated in the TBI group compared with non-TBI controls (fold change >1.2, p < 0.05, Table 2). The green and red colors in the heat map represent the up- and downregulated exomiR expression levels, respectively. The heat map shows a relative distinct separation between the TBI and noTBI groups (Fig. 1a).

Hierarchical clustering of the dysregulated exosomal micro-RNA (exomiR) profile with the pattern of expression in different TBI groups compared with the noTBI group.
Dysregulated ExomiR Profiles in Three Different Study Groups
ExomiR, exosomal micro-RNA.
The exomiR profile of the 1– 2 TBI group was compared with the noTBI group; 16 exomiRs were dysregulated in the 1–2 TBI group (Table 2). Specifically, 3 exomiRs were downregulated and 13 exomiRs were upregulated in the 1–2 TBI group (fold change >1.2, p < 0.05, Table 2). The hierarchical clustering of significantly dysregulated exomiRs showed a relative distinct separation between the 1–2 TBI and noTBI groups (Fig. 1b).
Similarly, we compared the exomiR profile between the rTBI and noTBI groups. There were 32 exomiRs dysregulated in the rTBI group (Table 2). Two exomiRs were downregulated and 30 exomiRs were upregulated in the rTBI group (fold change >1.2, p < 0.05; Table 2, Fig. 1c).
Gene network analysis for dysregulated exomiRs in TBI, 1–2 TBI, or rTBI groups
To investigate the molecular pathways associated with altered regulation in exomiRs in our cohort and explore the interconnections between these genes, we conducted network enrichment analyses of dysregulated exomiRs and their validated targets via IPA core analysis as well as network analysis within the total TBI, 1–2 TBI, and rTBI groups, respectively.
The miRNAs that were altered in regulation were associated with networks based on IPA. Enriched networks with a p value <0.05 were recorded in each TBI group. The networks of cellular movement in the TBI group, inflammatory response and organismal injury/abnormalities pathways in the 1–2 TBI group, and organismal injury/abnormalities and developmental disorder pathways in the rTBI group, were significantly activated compared to the noTBI group (data not shown).
We further investigated the biological pathways and functions of the dysregulated exomiRs and their association with related gene targets using IPA analysis. The results showed that two major exomiR hubs (hsa-miR-372-3p, and hsa-miR-762, synonym to hsa-miR-5001-5p; Fig. 2a) were found in the dysregulated exomiRs in the entire TBI group. Five miRNA subhubs were activated in the TBI group: (1) hsa-miR-3190-3p; (2) hsa-let-7e-5p; (3) hsa-miR-376b-3p; (4) hsa-miR-18a-5p; and (5) hsa-miR-567 (Fig 2a.). The hubs and subhubs were closely associated with insulin-like growth factor receptor (IGF1R) peroxisome proliferator activated receptor alpha (PPARA), extracellular signal-regulated kinase (ERK), transforming growth factor beta (TGFβ)/matrix metalloproteinase-9 (MMP-9), and tretinoin signaling pathways (Fig. 2a). IPA network analysis in the rTBI group also revealed several miRNA hot hubs including: (1) hsa-miR-762; (2) hsa-miR-1269a (synonym to hsa-miR-1269b); (3) hsa-miR-3202; (4) hsa-miR-92a-3p (synonym to hsa-miR-367-3p); (5) hsa-miR-95 (synonym to hsa-miR-1264); (6) hsa-miR-3118 (synonym to hsa-miR-134-5p); and (7) hsa-miR378a-3p (synonym to hsa-miR378f). These hubs were highly associated with TP53/cyclin dependent kinase inhibitor 2A (CDKN2A)/silent information regulator (SIRT1) mediated cell cycling/apoptosis/inflammation/neurodegeneration, proliferation/angiogenesis/apoptosis pathways, and insulin/IGF1R mediated metabolism dysregulation (Fig. 2b).
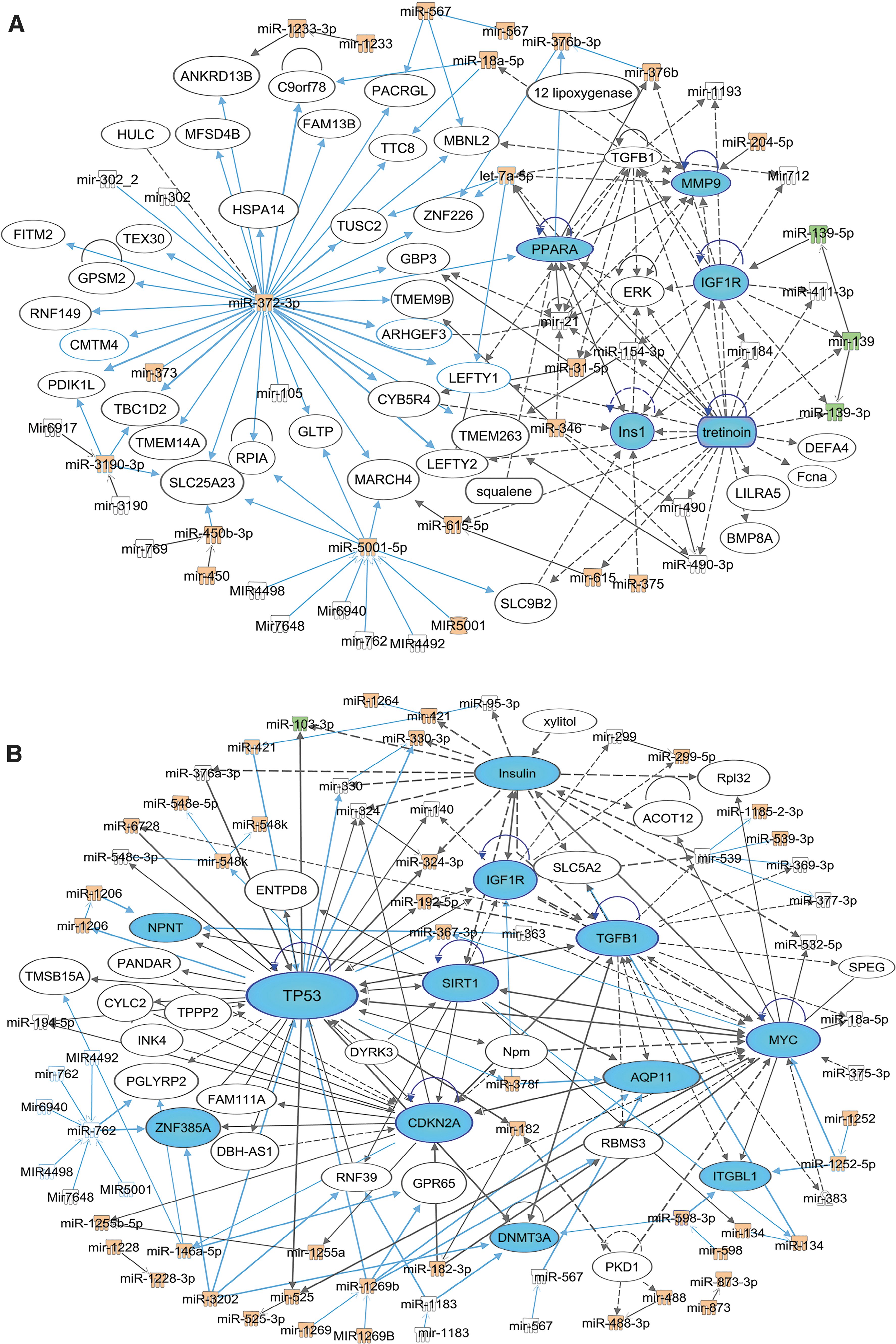
Ingenuity Pathway Analysis (IPA) network analysis for the dysregulated exosomal micro-RNA (exomiRs) in the entire traumatic brain injury (TBI) group or in the rTBI group. Analysis of the highest scored exomiRs and gene networks associated with the TBI group
miRNA expression levels are associated with the severity of post-concussive symptoms
TBI history and neurobehavioral symptom survey (NSI) scores (ρ = -0.21, p = 0.03) negatively and significantly correlated with the expression levels of hsa-miR-103a-3p (Table 3). Correlations between higher NSI scores and lower levels of hsa-miR-139-5p were significant (ρ = 0.25, p < 0.01). To further evaluate the relationship between NSI symptoms and the levels of these exomiRs in participants with a history of TBI, we used the GLM including age, gender, and time since last TBI as predictors. We observed that higher levels of hsa-miR-103a-3p and hsa-miR-139-5p were associated, respectively, with lower and higher NSI scores (Table 4).
Correlations between miRNA Levels and NSI Scores
Pearson correlation coefficient and significance levels between NSI Total Score or NSI factors (i.e., NSI Somatic, NSI Affective, NSI Cognitive, NSI Vestibular) and levels of differentially regulated miRNAs. p values < 0.05 are bold.
miRNA, micro-RNA; NSI, Neurobehavioral Symptom Inventory.
Generalized Linear Models for NSI Scores
Generalized linear models including demographics (age and gender) and TBI characteristics (Number of TBIs and time since last TBI) were built to further assess relationships between levels of the miRNAs and NSI Total scores or scores of NSI factors (i.e. Somatic, Affective, Cognitive, Vestibular). Only miRNAs that were significantly correlated with NSI total scores or scores of NSI factors were evaluated. Table represents models including both hsa-miR-103a-3p and hsa-miR-139-5p. In addition, a model including demographics and TBI characteristics, but not miRNA levels, is also shown. Results were shown for the predictors included in each model. p values < 0.05 are bold.
NSI, Neurobehavioral Symptom Inventory; SE, standard error; TBI, traumatic brain injury.
Discussion
mTBIs are common and increasingly linked to long-term outcomes and emergent neurodegeneration, especially in military personnel with repetitive injuries, but the underlying pathobiology is not completely understood. Our finding that exomiRs correlate with activity of pathways associated with inflammation and cell repair, especially in those with chronic neurological and behavioral symptoms, suggests that exomiRs may play a role in the pathophysiology of persistent TBI-related symptoms and may help identify candidates for targeted intervention. This biomarker analysis approach is novel and provides unique insights into the possible mechanisms of chronic symptoms, as exosomes are released from the CNS and can easily cross the BBB into the peripheral circulation, and carry specific contents distinct to that of free circulating miRNAs. Specifically, we report here that two miRNAs—hsa-miR-139-5p and hsa-miR-18a-5p—were significantly differentially expressed in the rTBI and 1-2 mTBI groups compared with controls. Hsa-miR-139-5p and hsa-miR-18a-5p have previously been linked to CSF exosomes in acute severe TBI, 16 as well as in Alzheimer's disease (AD) patients compared with healthy controls, 24 respectively. Additionally, in pre-clinical models, miR-18a-5p correlated with poor recovery after TBI through targeting of experimentally validated miRNAs 25 which influence processes related to neurogenesis and synaptic remodeling. 26 –28 Our findings suggest that these miRNAs that have been linked to acute and chronic neurological processes may be involved with TBIs, especially repetitive mTBIs in veterans and service members with persistent TBI symptoms.
Pathway analyses showed that within the entire TBI group, two major hubs and five miRNA subhubs were altered in regulation. These hubs and subhubs are closely associated with pathways involving insulin, IGF1R, PPARA, ERK, TGFβ1, MMP-9, and tretinoin signaling pathways. This finding may have pathomechanistic significance, as IGF-1 is a modulator of neuronal growth and function and signals predominately through IGF1R. 29 IGF1R signaling functions to control and regulate normal physiological processes in the brain related to growth, survival, metabolism, neuroprotection, and neuroregeneration. 24,30 –32 Dysregulation of this pathway may account for brain hypometabolism and several characteristic neuropathological features that overlap with several neurodegenerative diseases, including AD. 33,34 Dysregulation of IGF1R signaling has been implicated in AD. 24,30
Both IGF-1 and TGFβ promote neurogenesis, as well as clearance of amyloid-beta from the brain, and mediate the actions of growth hormone. 35 Growth factors including IGF-1 have also been implicated in recovery after TBI through promotion of neuronal regrowth and repair. Similarly, TGFβ has been shown to have similar properties and is critical to neurological recovery. ERK is one of three mitogen-activated protein kinase (MAPK) subfamilies. ERK has been shown to rapidly increase following TBI. 36,37 Sustained activation of ERK has been shown to have deleterious effects following brain injury 38 –40 possibly through upregulation of MMP-936 and its association with increased permeability of the BBB. Inhibition of the ERK pathway has been shown to decrease MMP-9 levels and mitigate tissue damage. 36 Little is known about the dynamics of MMP-9 in chronic TBI. TGFβ is a major profibrotic cytokine that regulates multiple cellular functions including differentiation, proliferation, and induction of synthesis of major extracellular matrix (ECM) proteins. Dysregulation of the TGFβ1/MMP-9 signaling pathway has been associated with interruption of the BBB. 41 –44 These findings suggest that chronic exomiR dysregulation in participants with remote mTBI may implicate these cellular pathways in repetitive TBIs.
Tretinoin, an all-trans retinoic acid, plays important roles in cell growth including neurogenesis, CNS differentiation, and apoptosis. 45 –48 Additionally, the tretinoin signaling pathway is involved in immunomodulating properties including the production of proinflammatory factors. 49,50 There are also several synthetic retinoids that are in clinical trial for the treatment of disorders with cell growth abnormality. 49,51 In aggregate, alterations in expression of miRNAs associated with these gene networks and hubs suggests a persistent interaction among developmental and inflammatory pathways for individuals with a history of remote mTBI, compared with controls. These findings suggest that these exomiRs may be candidate biomarkers of chronic neurobehavioral symptoms in remote mTBI.
Our pathway analysis for the 31 dysregulated exomiRs in the rTBI group revealed dysregulation of seven hot hubs. These hubs are highly associated with TP53/CDKN2A/SIRT1 mediated cell cycling/apoptosis/inflammation/neurodegeneration, fibroblast growth factor (FGF1) and Myc mediated cell proliferation/angiogenesis/apoptosis pathways, and insulin/IGF1R mediated metabolism dysregulation. SIRT1 is a nicotinamide adenine dinucleotide (NAD)+-dependent deacetylase and plays a role in type 2 diabetes, neurodegenerative disease, and cancer, 52 –55 as well as suppressing apoptosis. 56,57 SIRT1 has demonstrated a neuroprotective effect in pre-clinical studies of neurodegenerative disease and TBI 58,59 by preventing neuronal apoptosis. 58
TP53/Myc is also important in apoptosis pathways, further supporting the activated cell/tissue injury pathway in the remote rTBI group. Among the variety of functions regulated by the IGF-1 pathway, metabolic functions, cell proliferation/survival, myelination, and neurite outgrowth are all relevant to TBI. Altered IGF1R expression by several exomiRs after rTBI may adversely impact repair mechanisms. FGF1 is expressed in embryonic as well as adult neural tissue and is important in neuronal regeneration, 60,61 astrocyte activation, and astrocyte boundary formation after tissue injury. 62 –64
Several other exomiRs reported here have been previously related to neurological or TBI processes, including hsa-mir-103a-3p, hsa-mir-146b-5p, and hsa-mir-873, all of which were detected uniquely in the rTBI group. Hsa-mir-103a-3p has been implicated in immune regulation and neuronal dysfunction. 65,66 CSF-derived exosomal hsa-mir-873 has been shown to be differentially expressed in Parkinson's disease. 67 Hsa-mir-146bp-5p has also been identified in CSF-derived exosomes and was reported (along with hsa-mir-139-5p) to be differentially packaged into CSF EVs derived from TBI patients versus healthy controls. 16 Here, we observed higher levels of exomiR dysregulation in those with rTBI.
A major limitation of this study is that mixed severity and history of TBIs in this military cohort may cause variation in clinical detection. Well-characterized TBI histories with increased participant numbers may facilitate subgrouping for further investigation. In addition, studies at multiple molecular levels in along with miRNA investigation at multiple time points along the chronic symptom formation may greatly strengthen the current finding of regulatory miRNA functions in remote TBIs. Another limitation of this study is that we used total plasma exosomes for miRNA profiling. Brain derived-exosomes may possess distinct compositions and have specific regulatory functions to receiving cells. Dissection of neural derived-exosomes will advance our understanding of exosome biology in TBI and accelerate the development of exosome-based diagnostics and therapeutics.
This study is the first to establish an association between alterations in exomiR expression and neurological outcomes; participants with both mTBI and rTBI had distinct exomiR profiles compared with controls years after injury. Pathways most dysregulated related to changes in inflammatory regulation and neurological pathways, which were linked to neuronal repair processes. These profiles suggest that exomiR expression analysis may provide novel insights into the underlying pathobiology of chronic symptom persistence in persons with remote TBIs, and ultimately may lead to novel interventions to mitigate these symptoms.
Footnotes
Acknowledgments
We thank the military service members and veterans who participated in this study. The LIMBIC-CENC Observational Study Site principal investigators (PIs) or co-PIs also include: Risa Nakase-Richardson, PhD (Tampa), Carlos Jaramillo, MD (San Antonio), David Cifu, MD (Richmond), Melissa Guerra, MD (Fort Belvoir), Randall Scheibel, PhD (Houston), Terri Pogoda, PhD (Boston), Scott Sponheim, PhD (Minneapolis), and Kathleen Carlson, PhD (Portland).
Disclaimer
The views expressed in this manuscript are those of the authors and do not necessarily represent the official policy or position of the DoD, Department of Army/Navy/Air Force, Veterans Health Administration (VHA), United States government, or any other United States government agency. No official endorsement should be inferred.
Funding Information
This work was prepared under Contract W91YTZ-13-C-0015/ HT0014-19-C-0004 with VHA Central Office VA TBI Model Systems Program of Research/DHA Contracting Office (CO-NCR) HT0014 and, therefore, is defined as United States government work under Title 17 U.S.C.§101. Per Title 17 U.S.C.§105, copyright protection is not available for any work of the U.S. Government. For more information, please contact
Author Disclosure Statement
No competing financial interests exist.