
Abstract
Cell transplantation constitutes an important avenue for development of new treatments for spinal cord injury (SCI). These therapies are aimed at supporting neural repair and/or replacing lost cells at the injury site. To date, various cell types have been trialed, with most studies focusing on different types of stem cells or glial cells. Here, we review commonly used cell transplantation approaches for spinal cord injury (SCI) repair, with focus on transplantation of olfactory ensheathing cells (OECs), the glial cells of the primary olfactory nervous system. OECs are promising candidates for promotion of neural repair given that they support continuous regeneration of the olfactory nerve that occurs throughout life. Further, OECs can be accessed from the nasal mucosa (olfactory neuroepithelium) at the roof of the nasal cavity and can be autologously transplanted. OEC transplantation has been trialed in many animal models of SCI, as well as in human clinical trials. While several studies have been promising, outcomes are variable and the method needs improvement to enhance aspects such as cell survival, integration, and migration. As a case study, we include the approaches used by our team (the Clem Jones Centre for Neurobiology and Stem Cell Research, Griffith University, Nathan, QLD, Australia) to address the current problems with OEC transplantation and discuss how the therapeutic potential of OEC transplantation can be improved. Our approach includes discovery research to improve our knowledge of OEC biology, identifying natural and synthetic compounds to stimulate the neural repair properties of OECs, and designing three-dimensional cell constructs to create stable and transplantable cell structures.
Background
Although our understanding of spinal cord injury (SCI) has increased significantly over the past decades, the development of new clinically translatable therapies has been hampered by variable outcomes, limited improvement from surgical interventions, and the cost of therapeutic developments. The global prevalence of SCI has been reported to be 236–1009 cases per million persons, but it is difficult to accurately standardize such estimates. 1 –4 In Australia, prevalence of traumatic SCI alone has been estimated to 490–886 cases per million (approximately 10,000–20,000 persons), 5 with the highest prevalence in males and in the age group of 46–60 years; in Australia, high-level SCI resulting in quadriplegia constitutes the most common SCI outcome. 5 Recent Australian data indicate an annual incidence of 21–32.3 cases per million of traumatic SCI alone in Australia. 5 Fall-related incomplete quadriplegia and complete paraplegia, in particular in elderly persons, is increasing in incidence in Australia 5,6 and in other countries. 7 In addition to the personal suffering, the socioeconomic cost to society is high because of the numerous associated factors, including the high level of assistance provided by carers and family and the low of income earning potential of many persons living with SCI.
In 2009, the socioeconomic lifetime cost to Australia for persons living with paraplegia was $5 million (Australian dollars; AUD) and quadriplegia $9.5 million (AUD), which equated to approximately $2 billion (AUD) annually. 8 With improvements in healthcare and life expectancy, the socioeconomic costs in 2019 are likely to be much higher. Thus, novel therapeutic interventions for SCI are urgently needed.
Most SCIs are traumatic in nature. The period after primary injury (the injury caused by physical forces of the injury event) is followed by a cascade of secondary injury events, during which the injury typically expands and becomes more severe. Secondary injury is characterized by infiltration of immune cells, release of proinflammatory molecules and cytokines, as well as hypercalcemia and oxidative damage. Together, these events exacerbate cell death. 9,10 Several barriers prevent regeneration of the injured central nervous system (CNS). CNS neurons do not have a strong capacity for regeneration, attributable to both intrinsic and environmental factors such as accumulation of myelin-associated proteins, which inhibits axonal regeneration. After CNS injury, reactive astrocytes undergo hypertrophy and form a glial scar, the physical and chemical barrier which seals off the injury site from healthy surrounding tissue and prevents axons from extending across the injury site. Over time, extracellular matrix (ECM) becomes replaced with fibrous tissue, and further growth-inhibiting molecules are deposited at the injury site. 11
Current acute management strategies largely focus on symptomatic care with surgical decompression and blood pressure augmentation. 11 In the long term, management is primarily focused on living as well as possible with SCI, such as pain management and return to work. Activity-based rehabilitation has shown significant promise in improving function 12,13 ; however, there is currently no satisfactory SCI treatment available. Novel treatment approaches tackling SCI repair, most of which are currently in the pre-clinical phase, include the use of biomaterials, pharmacological approaches, and cell transplantation. Biomaterials such as hydrogels, which can be synthetic, biosynthetic, or biological, 14 can mimic ECM and stimulate repair. 15,16
Pharmaceutical approaches are typically targeted to ameliorate the inflammatory environment at the SCI site or aimed at stimulating neuronal survival and axonal regrowth. 17 –19 While the use of biomaterials and pharmaceutical management strategies have been reviewed elsewhere, the current review covers cell transplantation, with particular focus on transplantation of olfactory ensheathing cells (OECs), the glial cells of the primary olfactory nervous system. The work in this field, conducted by our group at the Clem Jones Centre for Neurobiology and Stem Cell Therapies at Griffith University (Nathan, QLD, Australia), is included as a case-study example of how OEC transplantation is being optimized toward improved therapeutic potential.
Cell Transplantation Approaches
The two core objectives of cell transplantation for SCI repair are to 1) modulate the wound environment in favor of intrinsic repairs and 2) provide a source of stem or progenitor cells to replace the lost tissue. To date, a range of cell types have been transplanted into animal models of SCI, and some of these have progressed to phase I/II clinical trials in humans. Cell transplantation therapies have resulted in highly variable results in terms of minimizing secondary injury progression and promoting tissue repair, accompanied by a vast range of functional outcomes. There are many factors which contribute to this variability, including the type and purity of cells; cell source (autologous, allogeneic, or xenogeneic); use of a support matrix; implant location; and transplant modality. 20 –23
A diverse array of cell types has been investigated for their potential use as therapeutics for SCI (Fig. 1). Further, these different cell types have been used at various developmental stages, including multi-potent cells (neural stem cells, 24 –29 mesenchymal stem cells [MSCs] 30 –36 ), unipotent progenitor cells (oligodendrocyte progenitor cells 37 ), or terminally differentiated cells (OECs, 38 –44 Schwann cells 45 –52 ). Alternatively, two or more cell types have been transplanted together, for example OECs cotransplanted with adipose-derived stromal cells. 53 However, regardless of cell type, there is considerable variation in survival and subsequent ability of transplanted cells to inhibit secondary injury and promote regeneration. 54 –57 However, in some instances, where mature peripheral glial cells have been transplanted to regulate the wound environment and encourage regeneration, long-term cell survival may not be necessary, 41 given that once the transplanted cells have initiated regeneration, endogenous cells can take over the role of the transplanted cells. Comparatively, the goal of stem/progenitor cell transplantation is often to replace damaged tissue, and thus long-term cell survival within the wound environment is required. 54,55,58,59
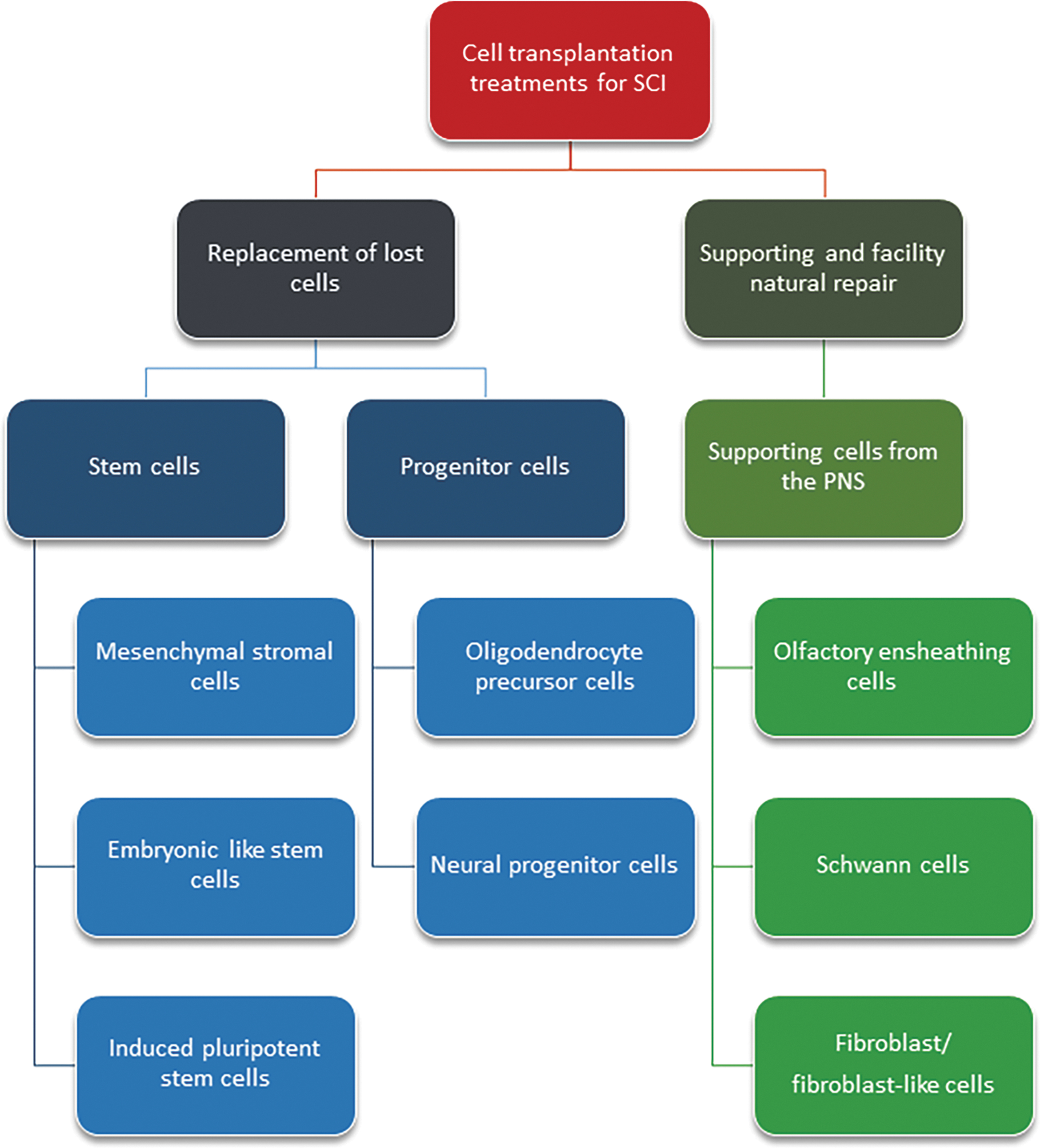
Potential therapeutic cell types for repair of spinal injury. PNS, peripheral nervous system; SCI, spinal cord injury.
Improving Cell Survival after Transplantation: Immunomodulation and Three-Dimensional Constructs
Recently, novel approaches have been developed to mitigate the impact of the wound environment on transplanted cells, with the aim to either improve cell survival or modulate the injury site environment, particularly the use of 1) immunosuppressants and 2) supporting materials allowing cells to be organized in three dimensions.
One major cause of this often variable and limited cell survival after transplantation is the hostile environment at the injury site, 60 attributable to the astrocytic scar which develops after injury, accumulation of cytokines, chemokines, cell debris, and immune cells, in combination with limited or absent vascular supply. 61 In animal models, it is uncommon for immunosuppressants to be used when transplanting allogeneic or autologous cells such of OECs, given that the assumption is that the cells themselves are unlikely to be immunogenic, although when using donor stem cells/progenitor cells, immunosuppressants are often used. However, for cell transplantation into traumatic CNS injury sites, including SCI, the use of transient systemic or local immunosuppression may improve long-term viability of the implanted cells regardless of source. Addressing the inflammatory factors by use of growth factors or steroids may elicit further improvements.
Another avenue for improving cell survival is to transplant them in a three-dimensional (3D) construct. In most studies, cells are injected as a suspension either directly into the wound site 38,47,50,62 –64 or into areas adjacent to the wound site, such as within the spinal cord parenchyma 65 –68 or the intrathecal space, 24,69,70 all of which have been associated with poor survival of transplanted cells. 57 In attempts to improve survival of transplanted cells, a range of biological 35,36,46,71 and biosynthetic materials, 27,48,72 –74 in particular hydrogels, have been tested. It is unclear whether the biomaterials actually directly improve cell viability and therefore improve neural repair, or whether the biomaterials purely function as a physical barrier which retains the cells within the SCI lesion, thus increasing their direct impact on the SCI. However, introducing an additive component increases the complexity of the treatment regime, which will lead to more-extensive experimentation to determine mechanisms of action and also for safety testing.
Another interesting approach to improve cell survival is to transplant cells which have already formed stable 3D constructs, as has been trialed with different types of cell-based transplantation approaches using mesenchymal stem cells. 75 –78 3D MSC spheroids have shown increased post-implantation survival in rat kidneys when compared to cell suspensions. 79 The 3D spheroid environment for MSCs has also been reported to increase the anti-inflammatory and angiogenic properties of transplants. 77,78 It is thought that a key factor which allows for 3D spheroids to improve not only cell survival, but also improve integration and repair is attributed to the cells being able to produce their own ECM and thus establishing an optimized microenvironment facilitated through cell-cell contact. 80
Autologous Transplantation of Olfactory Ensheathing Cells
Transplantation of OECs is emerging as a promising therapeutic approach for SCI repair. The primary olfactory nervous system is comprised of the olfactory nerve and outer layer of the olfactory bulb (the nerve fiber layer). Cell bodies of olfactory sensory neurons are found in the olfactory neuroepithelium at the roof of the nasal cavity, and their axons extend to the olfactory bulb in the brain. The axons are organized in multiple fascicles surrounded by OECs; together, these fascicles constitute the highly branched olfactory nerve 81,82 (Fig. 2). The primary olfactory nervous system is unique in that it naturally regenerates throughout life. Primary olfactory receptor neurons (ORNs) have evolved to naturally undergo apoptosis every ∼30 days (shown in rodents; the exact life span of human ORNs is not known). 83 Constant regeneration of these nerves is attributed, at least in part, to OECs. 83 –91 OECs are present in close contact with olfactory axons all the way from the lamina propria layer beneath the olfactory neuroepithelium to the nerve fiber layer of the olfactory bulb and are thus considered transitional glia present in both the peripheral nervous system (PNS) and CNS. 92 –94 Although one more-recent study challenges this paradigm and suggests that OECs are PNS glia, 95 the fact that these cells can exist in both the PNS and CNS constitutes one reason OECs have been considered good candidates for CNS transplantation. OECs can interact with CNS glia 90,96 and exhibit greater capacity for intermingling and integration with astrocytes than purely peripheral glia, such as Schwann cells. 97 –99 This is, in part, attributable to OECs having a unique heparan sulfate profile, which allows them to mingle with astrocytes whereas Schwann cells cannot. 97

The olfactory system comprises the olfactory mucosa and olfactory nerve that are located in the peripheral nervous system (PNS) and the olfactory bulb, which is located within the central nervous system (CNS). The olfactory sensory neurons project axons from the mucosa up to the olfactory bulb. OECs surround the axons to form the fascicles. OECs can be obtained from either the olfactory mucosa lining the nasal cavity or the olfactory bulb within the cranial cavity. OECs, olfactory ensheathing cells.
OECs also promote neural regeneration by providing physical support, forming a channel-like structure around axon fascicles, and by secreting a range of growth and guidance factors. 84,100,101,103 Further, OECs are highly phagocytic cells with innate immune functions. 104 –107 OECs are also considered good candidates for SCI repair because of their capacity for migration. During development of the primary olfactory nervous system and after injury, OECs migrate ahead of axons towards the olfactory bulb. 108 –111 Thus, OECs exhibit strong neural repair properties, can migrate well, and interact with astrocytes. For these reasons, in combination with the possibility of isolating OECs from nasal biopsies without the need for invasive neurosurgery, OECs are interesting candidates for transplantation into the injured spinal cord, particularly for autologous transplantation.
In the past few decades, OEC transplantation has shown strong promise for promoting regeneration in many animal models of SCI. In rodents, transplantation of OECs into the spinal injury site has led to reduced scar and cavity formation, 112 attenuated immune-cell–induced damage, 113 and reduced tissue loss, 113,114 accompanied by improved axonal extension across the scar. 113,115 OEC transplantation has also led to functional improvement in rodents, including improved locomotion and hindlimb movement, 116 –118 restoration of breathing and climbing, 119 and reduced autonomic dysreflexia. 65,120 In some studies, significant functional improvements have occurred even after complete spinal cord transection. 118,121 OECs have also shown strong promise for spinal cord repair in canines. 122
The first clinical trial using autologous transplantation of OECs in humans with SCI, led by Professor Emeritus Alan Mackay-Sim (Australian of the Year 2017; awarded particularly for his work with OECs and SCI), was performed in Australia in the early 2000s. This trial demonstrated that transplantation of OECs is feasible and safe, and although patient numbers were insufficient to determine statistical significance, some patients showed minor/moderate functional improvements. 123,124 Since then, several human clinical trials of OEC transplantation have been undertaken worldwide, resulting in a vast range of outcomes, 11,125 –131 with one particularly encouraging case of a patient who regained considerable function. 132 Despite these encouraging findings, the outcomes of OEC transplantation are highly variable, ranging from no improvement at all to significant regain of function. Thus, the method needs improvement. 131,133,134
Discrepancies in outcomes are largely attributable to differences in OEC purity and yield between studies, 135 –139 as well as time in culture before transplantation. 140 –142 In addition, OECs for transplantation have been isolated from distinct anatomical sites; whereas olfactory mucosa is the preferred site, given that it can easily be reached for biopsies, OECs can also be obtained from the olfactory bulb within the cranial cavity. Isolation of OECs from distinct sites is associated with variability in cell purity and cell functions. 143 –146 Combined with the use of different SCI models, it is therefore very difficult to compare outcomes of OEC transplantation studies. 147
In addition to development of standardized protocols for transplantation to reduce variability between studies, there are also key aspects of the method that need to be particularly focused on for improvement. First, it is essential to improve the OEC production protocol to minimize the time needed to culture sufficient cells for transplantation, given that the longer the cells are kept in culture, the more likely it is that their morphology, phenotype, or functional capacity may change. Currently, for human autologous transplantation, OECs must typically be cultured for a long time before being transplanted (2.5–10.0 weeks). 123,124,148 A second key aspect is to improve cell survival, which is not only typically very poor, but has also not been well documented in animal studies. 57 The third key aspect is to improve OEC migration within the injury site so that the transplanted cells can better integrate with the host tissue and thereby promote regeneration.
Currently, transplanted OECs display highly variable migration, with some studies demonstrating strong capacity for migration 149,150 and others showing limited migration. 151,152 Thus, in summary, improvements to the use of OECs to repair the injured spinal cord could be achieved by enhancing the cell-production process to generate higher-quality cells in a shorter period, by using 3D cell constructs to improve cell survival post-transplantation and then to encourage the migration and integration of cells into the host tissue.
Optimizing Olfactory Ensheathing Cell Transplantation for Spinal Cord Injury Repair: Example of an Australian Approach
Following on from the work by Professor Emeritus Alan Mackay-Sim, our team at the Clem Jones Centre for Neurobiology and Stem Cell Research at Griffith University, Australia, is working toward optimizing the preparation and therapeutic use of OECs to improve outcomes after transplantation to repair SCI. We are pursuing three main areas: 1) discovery biology to understand more about the roles of OECs; 2) compound screening to identify compounds (natural and synthetic) that can stimulate the neural repair functions in OECs; and 3) creation of 3D cell structures suitable for transplantation, which, we anticipate, will enhance cell survival post-transplantation (Fig. 3).
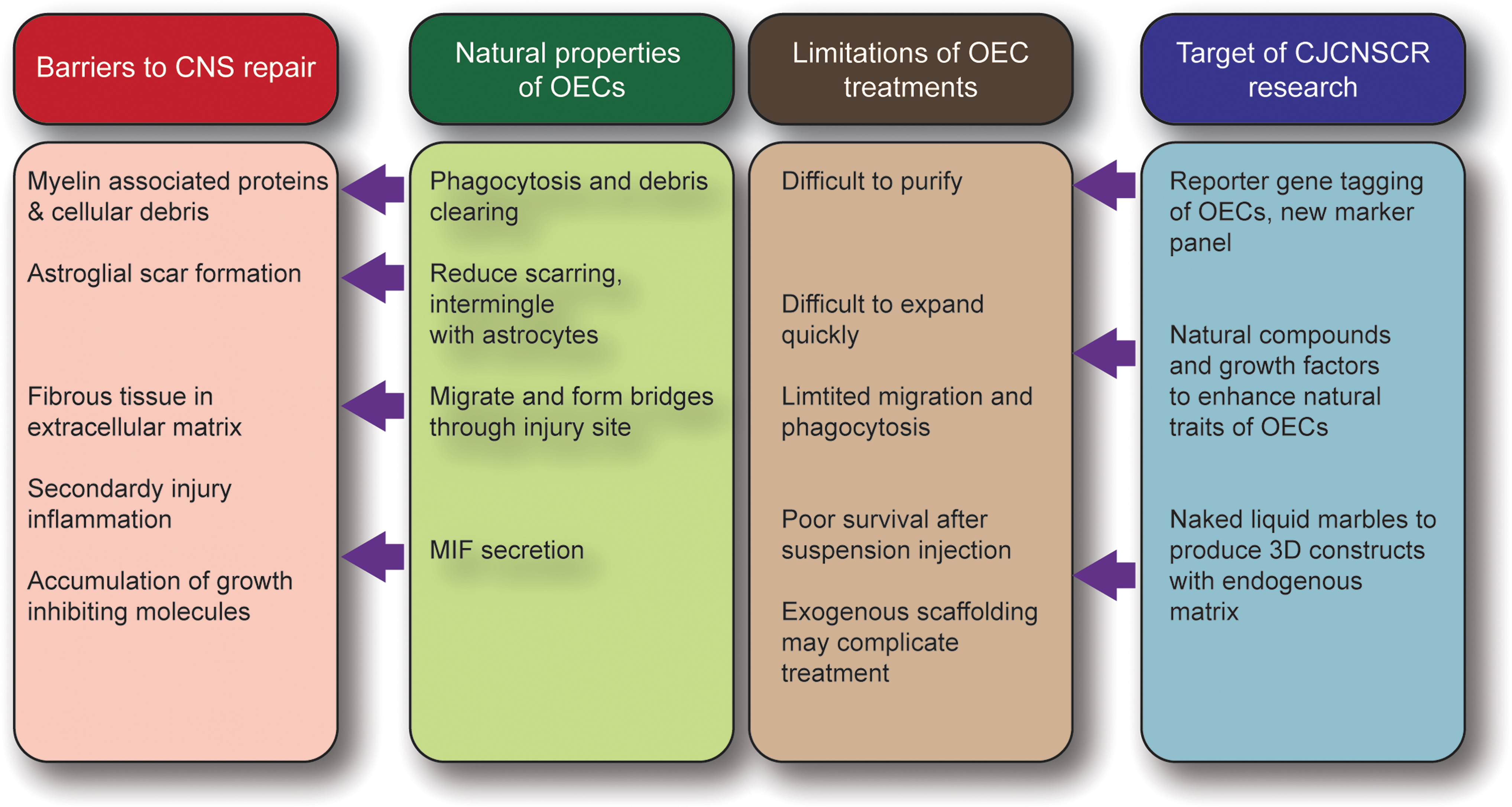
The approach to solve the limitations of the OEC transplantation therapy. OECs have properties that aid neural repair, but limitations in their use reduce the likely therapeutic success. The Clem Jones Centre for Neurobiology and Stem Cell Research is addressing key limitations with the use of OECs with the aim of improving the therapeutic outcomes for repairing spinal cord injury. 3D, three-dimensional; CJCNSCR, Clem Jones Centre for Neurobiology and Stem Cell Research; CNS, central nervous system; MIF, macrophage migration inhibitory factor; OECs, olfactory ensheathing cells.
Understanding why olfactory ensheathing cells are useful for neural repair
As mentioned above, the isolation and unambiguous identification of OECs is difficult because of the lack of specific markers for OECs. To overcome this, we generated a reporter transgenic, the S100 calcium-binding protein B (S100β)-DsRed line, in which the S100β promoter drives expression of DsRed in OECs. 153 Given that S100β is also expressed in all glia and some other cell types, such as macrophages and chondrocytes, fluorescence is not exclusively restricted to OECs. However, in combination with a panel of cell markers, however, this mouse model enables improved identification of OECs.
Despite the fact that OECs are known to promote neural regeneration, the cellular and molecular mechanisms behind OEC-mediated regeneration are largely unknown. 86,131,154,155 Over the past decade, we have worked toward an increased understanding of the biological roles of OECs. For example, we have shown that OECs exhibit highly motile lamellipodia, which are crucial for contact-mediated migration and cell-cell interactions, with more cell-cell contact between OECs leading to increased migration of the cells in vitro. 153 Migration of OECs is also an important regulator of axon extension, with in vitro studies demonstrating that increased OEC migration drives higher axon extension. 156
We have also demonstrated that mass migration of OECs in vivo is important within the olfactory system. Surgical removal of the olfactory bulb leads to both OECs and primary sensory olfactory axons growing into the cavity left by the bulbectomy. In an attempt to aid the repair of the traumatic injury, we have tested how transplantation of OECs into the cavity left after bulbectomy affects axon regeneration. 157 Thanks to the S100β-DsRed mouse line, OECs obtained from these mice express DsRed, which facilitates easy visualisation of OECs after transplantation. Transplanted OECs improved axon growth into the traumatic injury site, indicating that adding in more glial cells can enhance regeneration. 157 However, injection of a suspension of OECs led to the OECs being distributed across various regions without uniform contact between the cells. We therefore implemented an alternative approach, in which the axon regeneration was delayed because of chemical insult, which selectively affected the nasal mucosa, but which did not affect OEC migration to the injury site. In this model, OECs were able to migrate into the cavity without axons, with the OECs subsequently forming a more uniform mass of cells within the injury site. Several days later, the neurons within the nasal mucosa regenerated and extended axons to the injury site, where they were able to migrate over the extensive OEC mass that had been formed.
The subsequent result was that the axons spread out over a greater area compared to when OECs and axons migrate in together. 157 In vitro studies examining the combined effect of OEC migration and axon extension demonstrated that OEC migration is superior when axons are not present, and that axon extension is much greater when an OEC monolayer is pre-formed. 157 Together, these results indicate that a uniform mass of OECs may lead to higher migration of OECs, and that this may lead to subsequent enhanced axon extension.
This is consistent with the in vitro data which demonstrated that OECs migrate faster when there is sufficient cell-cell contact. Thus, when considering OECs for therapeutic treatment of the injured spinal cord, it would be important to design mechanisms to enhance cell-cell contact between OECs given that it may lead to improved migration within the injury site.
However, OECs are not a uniform population of cells, and thus predicting cell responses is more complicated. It has previously been shown that OECs from the olfactory nerve and olfactory bulb have distinct functions. 136,158 –161 We have shown that OECs derived from the olfactory nerve promote axonal fasciculation, while axons with OECs from the olfactory bulb exhibit a more disorganized extension pattern. 161 This is consistent with their roles in vivo, where OECs of the olfactory nerve ensheath the axons and guide them to the outer layer of the olfactory bulb, whereas OECs of the olfactory bulb contribute to the defasciculation and sorting of axons as they project to their diverse targets around the olfactory bulb. Yet, within the olfactory bulb itself, OECs may have different roles depending on their location. In the rostral-ventral nerve fiber layer, the incoming fascicles undergo extensive defasciculation and sorting, whereas in the dorsal olfactory bulb the axons have already been largely sorted and are refasciculated and projecting to their targets.
When OECs were obtained from these different regions and cultured in vitro, they exhibited distinct differences in their attraction versus repulsion behaviors with other OECs. 161 If these characteristics are retained after biopsy and purification for transplantation into the injured spinal cord, then these separate distinct populations of OECs may confer different outcomes for regenerating axons within the spinal cord depending on the anatomical site of cell harvest.
OECs not only help promote the growth and guidance of axons, but they are also highly phagocytic cells. Within the olfactory system, constant turnover of axons means that the dead axons need to be removed on a daily basis. Rather than macrophages, it is the OECs that are the primary cells responsible for phagocytosis of cell debris in the olfactory nerve. 104,106 We found that macrophages are largely absent from the inside of axon fascicles, and instead, OECs phagocytose the large amounts of debris derived from turnover of olfactory neurons. 104,105 We and others have also shown that OECs can respond to and phagocytose bacteria. 162 –165 More recently, we have now identified that OECs express macrophage migration inhibitory factor (MIF), which likely repels the macrophages and thereby prevents them from intermingling with the OECs. 166 From a therapeutic point of view, this adds to the suite of tools that OECs may utilize on the injury site when transplanted into the spinal cord. They can potentially phagocytose the debris from the dead cells that arise because of the injury, and they may regulate the recruitment of microglia or macrophages because of their natural expression of MIF.
The safety of OEC transplantation into the injured spinal cord is of high priority, and one consideration is not just the subpopulation of the OECs, but also the other cell types that are included in the cell preparation given that these can affect cell survival and integration. 57 Sourcing OECs from different anatomical sites also results in different contaminant cells being present in cell harvests. OEC cultures derived from the olfactory nerve frequently contain fibroblasts, whereas olfactory-bulb–derived cell cultures may contain both fibroblasts and astrocytes. OEC purification is notoriously difficult; however, significant advances in purification techniques have been made over the past decade. 145,158,159 It is crucial that the composition of cells in transplants is highly controlled. Presence of other cell types than OECs in transplants can have significant effects on outcomes, and accidental transplantation of respiratory epithelium has resulted in the formation of mucous-producing cysts in the spinal cord. 167,168 Purification commonly involves immunolabeling-based methods, 113,115 such as fluorescence-activated cell sorting, which is complicated by the fact that there are currently no markers exclusively expressed in OECs, and that marker expression can differ between rodent and human OECs. Thus, it is important that purification and identification of OECs rely on a panel of several markers than just one or two markers (commonly the p75 neurotrophin receptor, which is also expressed by Schwann cells, and S100, which is expressed in all glia). 169 Our team is currently developing suitable panels of markers for purification of mouse and human OECs.
Apart from the potential of contaminating cells being present within the OEC cell preparations, the risk of uncontrolled cell growth leading to tumour formation needs to be carefully considered. One characteristic of OECs in this regard is that there are very few reports of OECs forming tumors in their natural environment within the olfactory system of humans. 170 Indeed, of the few reports of potential OEC tumors, the lack of robust identification markers casts doubt on whether tumors arose from OECs or from other glial cells, such as Schwann cells. We have speculated that one reason why OECs may be less prone to forming tumors is the relatively harsh environment in which OECs naturally live. They are constantly tending to dead and dying olfactory sensory axons to remove the cell debris, 104 and they are under threat of bacterial attack given that pathogens can quite easily penetrate the olfactory nerve. 171 –173
In contrast, other glial cells typically only remove dead cells only after injury, and they are less exposed to the risk of pathogens, and gliomas involving Schwann cells, astrocytes, or oligodendrocytes are more common, although the safety of Schwann cell transplantation has been demonstrated in a human clinical trial with no adverse effects reported in the 12 months after transplantation. 52 Whether the lack of tumor-forming potential of OECs is maintained after extraction, purification, and transplantation into the injured spinal cord is uncertain and needs to be carefully monitored and assessed.
Stimulation of olfactory ensheathing cell functions relevant for neural repair
It can be difficult to harvest enough OECs for transplantation from biopsies, and the harvested cells must typically be cultured for several weeks in vitro before transplantation, 124,148,174 which can result in dedifferentiation. 175 Further, as discussed earlier, survival of OECs after transplantation is often poor. Thus, compounds or factors that can stimulate OEC proliferation or viability may enhance the therapeutic potential of cells. It may also be useful to modulate the activity of OECs to improve their migration or phagocytosis after transplantation. Although OECs are naturally migratory cells, results regarding OEC migration in the SCI site after transplantation are highly variable. 149 –151 It is also possible that stimulating the phagocytic activity of OECs (without causing a proinflammatory response) can be beneficial for debris clearance and therefore neural repair. 84,134
We and others have shown that OEC migration can be stimulated using growth factors, such as glial-derived neurotrophic factor and brain-derived neurotrophic factor, 134,153,176 which can result in better axonal extension. 156 We have also shown that natural compounds can promote neural repair properties of OECs. For example, we have found that several natural compounds can enhance OEC viability, including curcumin from turmeric, 177 linckosides isolated from starfish, 178 and serrulatane diterpenoids from the Australian plant Eremophila microtheca. 179 We have also shown that curcumin, in addition to stimulating viability and migration, at low dose strongly simulates OEC phagocytosis 177 ; the effect on phagocytosis has since also been confirmed by another group. 180 These isolated examples of how compounds can stimulate selective activity of OECs has led us to perform a large screen of several thousands of natural and synthetic compounds from Compounds Australia and NatureBank, two major compound resources which are located at the Griffith Institute for Drug Discovery, Griffith University. We are now in the process of further characterizing how the successfully screened compounds modulate OECs with a view to identify compounds that can stimulate neural repair-related OEC properties.
Three-dimensional culture
As for most other cell transplantation therapies, the current standard approach for OEC transplantation into the injured spinal cord, in both pre-clinical models and in human clinical trials, is to inject the cells as a suspension. 11,125,131,133 With this transplantation method, each individual cell needs to incorporate itself into the host tissue, migrate to the appropriate region, and form stable cell-cell connections. However, as discussed above, OECs are highly dependent on cell-cell contacts for migration, with increased migration occurring when OECs are at higher density, 153 and the migration of OECs positively regulating axon guidance. 156,161 Therefore, injecting a suspension of OECs into the injured spinal cord is unlikely to result in OECs maintaining sufficient cell-cell contact with each other, which may lead to reduced ability to migrate and interact with axons. Rather than injecting a suspension of cells, an alternative approach is to transplant 3D constructs of OECs in which the cells have already formed stable cell-cell connections with each other. While biological scaffolds are currently being trialled to support OECs in vitro or in vivo, 181 –184 the introduction of additional components may lead to unexpected outcomes or complicate the interpretation of results.
In our laboratory, we have focused primarily on generating 3D OEC constructs, in which the cells form stable 3D structures using only regular media formulation without the need for any additive components, artificial substrates, or scaffolds. We have recently developed a 3D spheroid culture system, termed the Naked Liquid Marble (NLM) platform, 185 which stems from our earlier work on culturing cells in floating liquid marbles. 187 OECs are cultured in a liquid drop (the NLM) on a superhydrophobic coated surface, causing cells inside the NLM to naturally and rapidly (within hours) form 3D spheroids. This low-cost system, which can be produced using automated laboratory systems, enables highly reproducible production of 3D spheroids. The size of the spheroids can be tailored using different cell numbers/density or by using vibration at different frequencies during spheroid formation. 187
The benefit of the 3D spheroid formation is demonstrated by the robust migration of OECs from the spheroids after they are transferred to a regular two-dimensional culture plate. Compared to dissociated cells or small spheroids, the larger spheroids led to uniform migration of OECs out from the spheroid over larger distances. 187 If these in vitro characteristics of the 3D naked liquid marble spheroids were retained after transplantation into the spinal cord, then enhanced integration and migration may be achieved. We are currently trialing transplantation of OEC spheroids produced using this system of transection and contusion rodent models of acute, subacute, and chronic SCI.
Importantly, the NLM 3D culturing system is suitable for coculturing different cell types to determine how cells interact and sort out. The ability of the cells to freely move within the liquid marbles enables them to naturally arrange themselves in a manner that resembles in vivo cell relations. For example, if OECs and fibroblasts are combined, the core of the spheroids consists of OECs and the outer layer of fibroblasts, resembling the organization in the olfactory nerve where OECs are surrounded by perineural fibroblasts. When OECs and astrocytes are cocultured, the cells loosely organize into spheroids consisting of OECs surrounded by astrocytes, mimicking the glia limitans layer of the olfactory bulb. 95,185 Thus, in addition to being suitable for transplantation into the SCI injury site, the NLM system is excellent for in vitro studies of OEC behavior in a 3D multi-cellular environment.
Conclusion
The transplantation of OECs is a promising approach for treating SCI, but the methods need improvement. We have identified that improving our understanding of the biology of OECs and their potential roles can aid the design of strategies to improve the therapeutic use of the cells. In particular, the current major bottlenecks in the therapy include 1) the difficulty in purifying OECs from the olfactory mucosa, which is the preferred source, 2) amplifying the numbers of cells while maintaining their phenotype, 3) maintaining and/or stimulating cell behaviors, such as migration and phagocytosis, and 4) survival of cells after transplantation. We have demonstrated that exploration of compounds/drugs is revealing that the viability, proliferation, and activity of OECs can be selectively enhanced. In addition, the advent of 3D cell culture enables the creation of cell constructs suitable for transplantation that are likely to result in better cell survival and integration after transplantation in an injury site. Overall, the combined approaches of improved cell purification, stimulation of OEC activities suitable for neural repair, and the 3D production of cell constructs are likely to lead to enhanced therapeutic outcomes for spinal cord repair.
Funding Information
This work was supported by a Motor Accident Insurance Commission (Queensland State Government) grant to J.S.J. and J.E., a Perry Cross Spinal Research Foundation grant to J.E. and J.S.J., a Clem Jones Foundation grant to J.S.J. and J.E., and a Catwalk Trust Grant to J.S.J.
Footnotes
Author Disclosure Statement
No competing financial interests exist.