
Abstract
Bowel dysfunction after chronic spinal cord injury (SCI) is a common source of morbidity and rehospitalization. Typical complications include constipation, fecal impaction, incontinence, abdominal distention, autonomic dysreflexia, and the necessity of interventions (i.e., suppositories, digital stimulation) to defecate. Numerous surveys have confirmed that the remediation of bowel complications is more highly valued for quality of life than improvements in walking. Much of what is known about bowel function after SCI for diagnosis and research in humans has been gained using anorectal manometry (ARM) procedures. However, ARM has been underutilized in pre-clinical animal work. Therefore, a novel combination of outcome measures was examined in the current study that incorporates functional output of the bowel (weekly fecal measurements), weight gain (pre-injury to terminal weight), and terminal ARM measurement with external anal sphincter electromyography under urethane anesthesia. The results indicate higher fecal output after contusion during the sub-acute period (4–7 days) post-injury, changes in the composition of the feces, and functionally obstructive responses in a specific section of the rectum (increased baseline pressure, increased frequency of contraction, and reduced ability to trigger a giant contraction to a distension stimulus). These results demonstrate significant bowel dysfunction in the rodent SCI contusion model that is consistent with data from human research. Thus, the combined measurement protocol enables the detection of changes and can be used, with minimal cost, to assess effectiveness of therapeutic interventions on bowel complications.
Introduction
Spinal cord injury (SCI) causes a plethora of complications that impact nearly all bodily functions. When surveyed, bowel complications are a top priority quality-of-life issue in the SCI population and the restoration of this function is more highly rated than improvements in walking.1– 6 Further, bowel complications are a significant source of morbidity and rehospitalization. 7
Constipation, fecal incontinence, abdominal distension, bloating, difficulty with evacuation, diverticulitis, hemorrhoids, and fecal impaction are common consequences of SCI. 8 –14 Additionally, the excessive time necessary to complete a bowel program, which is often over an hour, coupled with the risk of fecal incontinence severely limits participation in daily life. 15,16 These limitations often lead to bowel programs being conducted every other day, which can exacerbate ongoing dysfunction (i.e., constipation). 16
Upper motor neuron (UMN) injuries occur in humans when damage to the spinal cord is localized above the conus medullaris (L1–L2) and spares the sacral reflex circuits. 12,13 Rectal tone, baseline colonic activity, and external anal sphincter (EAS) activity are all elevated, with the EAS becoming dyssynergic and spastic. 12 –14, At the same time, the internal anal sphincter (IAS) has reduced tone 12 and therefore there is reduced ability to maintain continence. Typically, excessive segmental activity and reduced propulsive activity cause slower movement of material through the organ. 12 Additionally, the complex coordination between the IAS, EAS, and rectum is disrupted causing difficulty in evacuation and necessitates intervention for successful defecation. 12,13 A common intervention is manual/digital stimulation of the anus which triggers relaxation of the IAS and contraction of the rectum to induce defecation. 1 –3 Therefore, SCI in humans causes numerous functional changes that combine to impair the storage and evacuation functions of the gastrointestinal tract.
Many studies have investigated colonic motility after SCI in humans and overall the results show a reduction in motility after injury. 17 –23 Although the findings can be mixed in any one study, the overall picture is that there is a reduction in the movement of material through all major divisions of the colon. This finding is consistent with the common complications of constipation and fecal impaction found in clinical settings.
There are several categories of contractions in the bowel that have been observed in rodents: phasic contractions, which are small amplitude and related to segmental peristalsis; giant contractions, which are very high amplitude contractions primarily for mass movement of material through the organ; and tonic contractions, which are related to the resting tone of the organ. 12,13,24,25 The presence and frequency of giant contractions are species-dependent (humans, ∼2/day; rat, ∼40/h) and in humans are related to the urge to defecate. 25
Anorectal manometry (ARM) is often used clinically to determine colonic dysfunction (i.e., diabetic diarrhea, irritable bowel syndrome) and has been used extensively to investigate dysfunction after SCI in humans. 26 –32 Indeed, much of the characterization of dysfunction after SCI are results from human manometry studies. Although ARM has been used extensively in the clinic to investigate the deficits that occur after SCI, there are relatively few animal models which utilize this technique. 29,32 –46
Although few in number, pre-clinical studies utilizing animal models of bowel dysfunction have added to the understanding of this complication. Much of the pre-clinical work utilizes a high level (T3-4) injury model to study gastrointestinal complications after SCI. In a complete transection model, it is clear that colonic motility is reduced 29 and cholinergic responsiveness of ex vivo colonic muscle strips is reduced. 37 Further, reduced giant contractions, elevated inflammatory markers, histological disturbances, and reduced enteric neuron density have been found after severe contusion (7% white matter sparing). 34
Colonic contractions can be stimulated by ghrelin agonist stimulation in rats 35,36 as well as neurokinin receptor 2 stimulation in minipigs, 38 dogs, 40,41 macaques, 42 and rats. 39,43 –45 Importantly, NK2 activation has similar action after high-level spinal cord transection acutely and chronically and may represent an intervention target and possible compromised mechanism after SCI. Further, remodeled changes in afferent signaling have been found after a luminal irritant inflammatory stimulus. 46 Taken together these studies provide insight into some of the changes that occur after chronic severe SCI in rat models, as well as highlighting neurochemical targets including acetylcholine, ghrelin, tachykinins, and nitric oxide.
Above studies notwithstanding, numerous gaps in our knowledge remain regarding compromised bowel function after SCI including enteric remodeling, influence of enteric and autonomic systems on the neuromuscular junction and motility, multiorgan dysfunction after injury, inflammatory mediators, and mesenteric perfusion. 45 The efforts undertaken to fill in these gaps would be aided by a testing model which is simple to use, sensitive to disturbances, and relatively inexpensive.
Therefore, we undertook the development of such a model using a clinically relevant T8 moderate-severe contusion injury. This model utilizes ARM as a tool for investigators to explore mechanistic, pre-clinical, treatment development, and translational endpoints. The set of outcome measures was designed to provide a comprehensive picture of bowel function (spinally intact) and dysfunction (spinal contusion), by combining functional output (weekly metabolic and home cage collection), weight gain (preinjury to terminal difference), and terminal ARM procedures (measurement of baseline activity, distension-induced responses, and EAS electromyography [EMG]) to detect the interaction between fecal output and rectal/colonic dynamics.
Methods
Animals
All animal procedures were performed according to National Institutes of Health guidelines after review and approval by the Institutional Animal Use and Care Committee at the University of Louisville, School of Medicine. Twenty-four male Wistar rats (Harlan; Sprague Dawley, Inc., Indianapolis, IN) were housed in a 12-h light-dark cycle with ad libitum food (Laboratory Rodent Diet) and water with soft paper home cage bedding (∼4 mm2 squares; AlphaDry). Functional assessments occurred prior to (baseline) and throughout (daily Days 2–7, then weekly) the recovery period. Terminal procedures occurred in the 10th week after injury.
Spinal cord injuries
Surgical protocol has been published previously. 47 –49 Briefly, animals were anesthetized with a ketamine (80 mg/kg; Ketoset; Fort Dodge Laboratories, Fort Dodge, IA) and xylazine (10 mg/kg; Anased; Akorn Inc, Lake Forest, IL) mixture. Chlorohexiderm scrub (Dermachlor 4%; Henry Schein, Dublin, OH) was used to cleanse the incision site after the fur was shaved and eye lubricant (Optixcare; CLC Medica, Ontario Canada) was applied to the eyes. Antibiotic injections included gentamicin (subcutaneously [SC], 5 mg/kg, once daily for 5 days; GentaFuse, Sparhawk Laboratories Inc, Lenexa, KS) and penicillin G (SC, 0.5 mL, once at surgery as a prophylactic measure; Penject; Bimeda Inc, Le Sueur, MN). The anti-inflammatory/analgesic meloxicam (0.2 mL SC; Eloxiject; Henry Schein, Melville, NY) was given prior to surgery and twice daily for 3 days for pain management. Fluids were given (10 mL, 0.9% sterile saline SC, one bolus of 5 mL over each hip) once at surgery and as necessary in the recovery period.
A T9 contusion was done on 12 rats using the Infinite Horizon impactor with a force of 215 kDyne (moderate/severe). 47 –49 A midline incision overlaying the mid-thoracic vertebra and T8 laminectomy was performed. After exposure, the T7 and T9 spinous processes were clamped to stabilize the spinal column and the contusion applied. Twelve sham surgical rats received the same procedures, except for the contusion. The muscle was then sutured with 4-0 Ethicon non-absorbable surgical suture, the skin closed with Surgical Autoclips (Mikron Precision Inc., Gardena, CA), and antibiotic ointment applied to the incision site. Heating pads were used to maintain body temperature during the surgery and recovery period. In addition to medication, post-surgical care included manual emptying of bladders (Credé maneuver, three/day for 5 days until fully reflexive), additional food access via several food pellets placed on the cage floor, visual inspection of the incision site (when bladder emptied), assessment of signs of distress (porphyrin staining, poor grooming, grimacing, and overgrooming/autophagia), and cleaning of soiled fur to prevent urine scald.
Basso-Beattie-Bresnahan testing
Basso-Beattie-Bresnahan (BBB) testing was conducted in an open field for observations of hindlimb function according to published criteria. 50 Testing was conducted and confirmed between two observers and the data used as a supplement to histological extent of lesion measurements regarding severity and consistency of the contusion.
Feces collection
Feces data was collected using two different methods each for 24 h: C.L.A.M.S. metabolic cages (Oxymax; Columbus Instruments, Columbus, OH) and by swapping home cages. Metabolic cage measures were collected twice prior to injury (baseline) and once weekly after injury. Home cage collections were started on Day 2 of recovery and continued daily through Day 7 to capture output during the acute recovery period. Further, home cage collections continued once weekly throughout the recovery period. Metabolic cages collect particulates (feces and food crumbs) over the 24-h period and both food crumbs and fecal pellets are quantified (counted, weighed). Feces from home cages were collected by hand and then counted and weighed. All metabolic cage fecal samples were air dried in a fume hood for 2 h prior to quantification due to moisture content. Home cage fecal samples are far more desiccated by the paper bedding and therefore could be quantified immediately.
ARM procedure
Prior to terminal testing there was no food restriction or overnight fasting; therefore, these data were collected from a replete system. On the testing day, rats were anesthetized with urethane (1.2 g/kg, intraperitoneally; Aldrich Chemistry, St Louis, MO) and placed on a heated water pad (Gaymar) to maintain body temperature. The animal was placed in the prone position on the heating pad and the tail elevated using a tail clamp to allow access to the anus for testing. Fine wire hook electrodes were fabricated from stainless steel wire (0.003″ diameter coated wire; A-M Systems, Sequim, WA) and a 27-gauge needle. Electrodes were implanted bilaterally into the EAS using an oblique angle starting at the midline and moving laterally through the muscle. The EMG signal was amplified (A-M Systems, four-channel differential AC amplifier), then captured using a CED Micro1401 acquisition unit and Spike 2 software (Version 8.15).
The ARM testing procedure is illustrated graphically in Figure 1 alongside normative example data traces. EAS EMG testing (Fig. 1A) consisted of four distension trials by inserting a balloon catheter 1–2 cm into the anus, inflating the balloon with 0.8 mL water (approximates the size of a large fecal pellet), and slowly (over approximately 7–10 sec) pulling the balloon through the anus. Subsequent EMG activity during the closure of the EAS after distension was recorded and analyzed (details below).
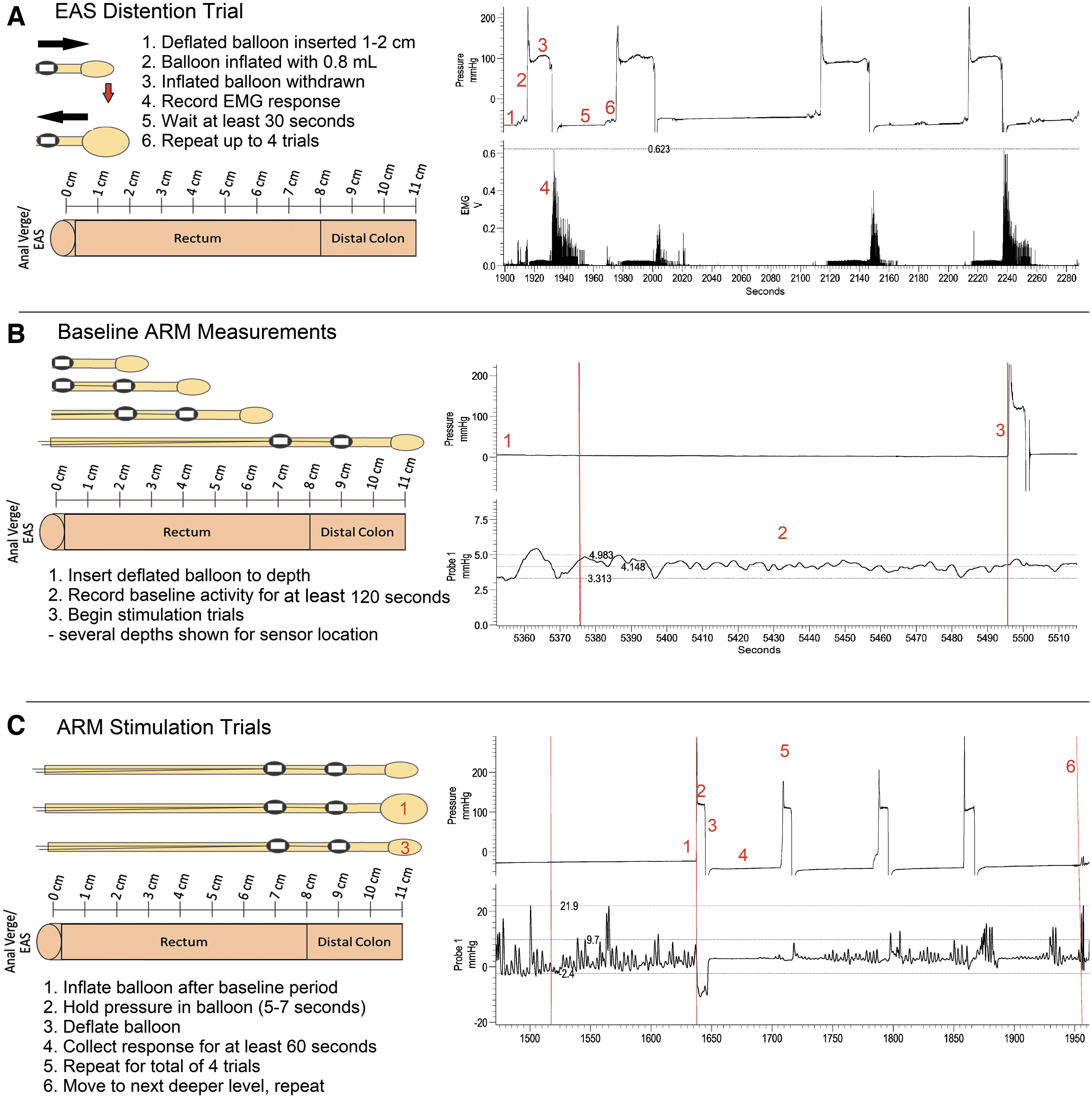
Methodology illustration showing a balloon probe diagram and anorectal manometry (ARM) testing examples.
Manometry pressure recordings were collected using an in-house fabricated balloon catheter 51 (modified butterfly catheter with a 1 cm balloon made from a non-lubricated latex condom) that had two pressure sensors (Millar, SPR-524, 3.5F; recommended by G.M. Holmes) affixed at 2 cm and 4 cm from the end of the catheter tubing using Parafilm (Bemis, USA). At each of nine levels tested, starting with 2 cm and going as deep as 11 cm, the probe was put into position and fixed to the tail with tape to prevent movement during testing. Two minutes of baseline pressure activity was recorded (Fig. 1B), followed by stimulation trials (Fig. 1C) that began with inflation of the balloon with 0.8 mL water for approximately 5–7 sec and then deflation. Response to the balloon distention stimulus was recorded for approximately 60 sec. The stimulation trial was repeated for a total of four trials. Once the 60-sec response period was over for the last stimulation trial, the probe was moved to the next level (i.e., from 2 to 3 cm depth) and the procedure repeated until all nine levels (ending at 11 cm depth) had been measured. Non-spermicidal lubricant (Surgilube; HR Pharmaceuticals Inc, York, PA) was used as needed during the procedure.
EMG data quantification and filtering
Signals were acquired with a four-channel differential AC amplifier (AM-Systems) and acquisition unit (CED Micro4 1401). EMG signals were filtered using a 50–500 Hz band-pass filter. Signals were then rectified and quantified for latency (using the balloon pressure channel as a marker for balloon exit from the anus), duration of response, maximum amplitude, and area under the curve (or integral).
Transcardial perfusion and tissue collection
Animals were perfused after testing by first being given an overdose bolus of urethane anesthesia. Following the injection, animals were rapidly perfused as previously described. 47 –49 A total of 300 mL heparinized phosphate buffered saline was used for exsanguination followed by 300 mL 4% phosphate buffered paraformaldehyde.
Distal colon, rectum, and anus tissue were collected fresh (after exsanguination but before paraformaldehyde) and frozen for future biochemical analyses. During the exsanguination, a laparotomy was performed, and the skin incision is continued along the perineum. Incisions are made around the anus and a combination of blunt and sharp dissection is used to free the rectum and anus from ligaments. After exsanguination was complete, the colon was then separated from the mesentery using blunt dissection and the anus pulled through the pelvis into the abdominal cavity. The colon was clamped with a hemostat just distal to the cecum and removed. Because the removal preparation was done during the exsanguination, the perfusion pump was off for approximately 15–30 sec before resuming, after which paraformaldehyde was infused for fixation of the remaining tissues.
Histology and sparing quantification
SCI histology was done following published protocols 47 –49 by first collecting a block of tissue 1 cm rostral and caudal to the lesion site. Tissue was later cut on a cryostat into coronal sections 35 μm thick and thaw-mounted for staining. Staining was done using cresyl violet (gray matter) and luxol fast blue (white matter) for sparing quantification. Micrographs were taken (SPOT RT3, 2MP microscope camera, Nikon Eclipse E400 microscope, 40 × magnification) and quantified (SPOT software 5.1) following established protocols. 47 –49 Briefly, sparing percentages were calculated by averaging the area (mm2) of the white and gray matter in the intact sections taken rostral and caudal to the injury. This average was then compared to the area of tissue at the epicenter and a percentage of sparing was calculated for both white and gray matter.
Statistical analysis
Data were analyzed using a two-way (Sham vs. Contusion; Distance) repeated measures analysis of variance with the Holm-Sidak post hoc analysis, except weight gain which required an independent sample Student's t-test. Variability is depicted in figures as standard error of the mean (SEM).
Results
Animals
Due to repeated measures statistics requirement, only rats with complete data sets were included in the analysis (18 of 24; nine per group). Reasons for exclusion from analysis were animal death from terminal testing anesthesia (n = 4; 2/group), non-responsive EAS in EMG trials (n = 1, sham group), and fecal blockage that prevented 11 cm testing (n = 1, contusion group).
Injury
For the contusion group, mean force of impact was 219.2 (± 5.3) kDyne with a mean displacement of 1421.1 (± 497.2) μm. Mean BBB scores (Fig. 2) were significantly reduced after injury (p < 0.001) with the contusion group having a mean score of 10.1 (± 0.4) after Day 14. A BBB score of 10 indicates occasional weight supported plantar steps with no forelimb-hindlimb coordination. White matter sparing (mean = 16.0 ± 4.2 %) and gray matter sparing (1.5 ± 1.6 %) extent after injury are indicated with a representative example (provided in Fig. 3).

Average (± standard error of the mean) Basso-Beattie-Bresnahan (BBB) scores of sham (n = 9) and contusion (n = 9) animals throughout the recovery period. At 7 days post-injury, the Contusion group shows severe and significant hindlimb impairment that is present at all time-points after injury (*significant, p < 0.001). The # indicates several instances of a 20 score in shams, which means some animals had consistent and coordinated stepping with some trunk instability.
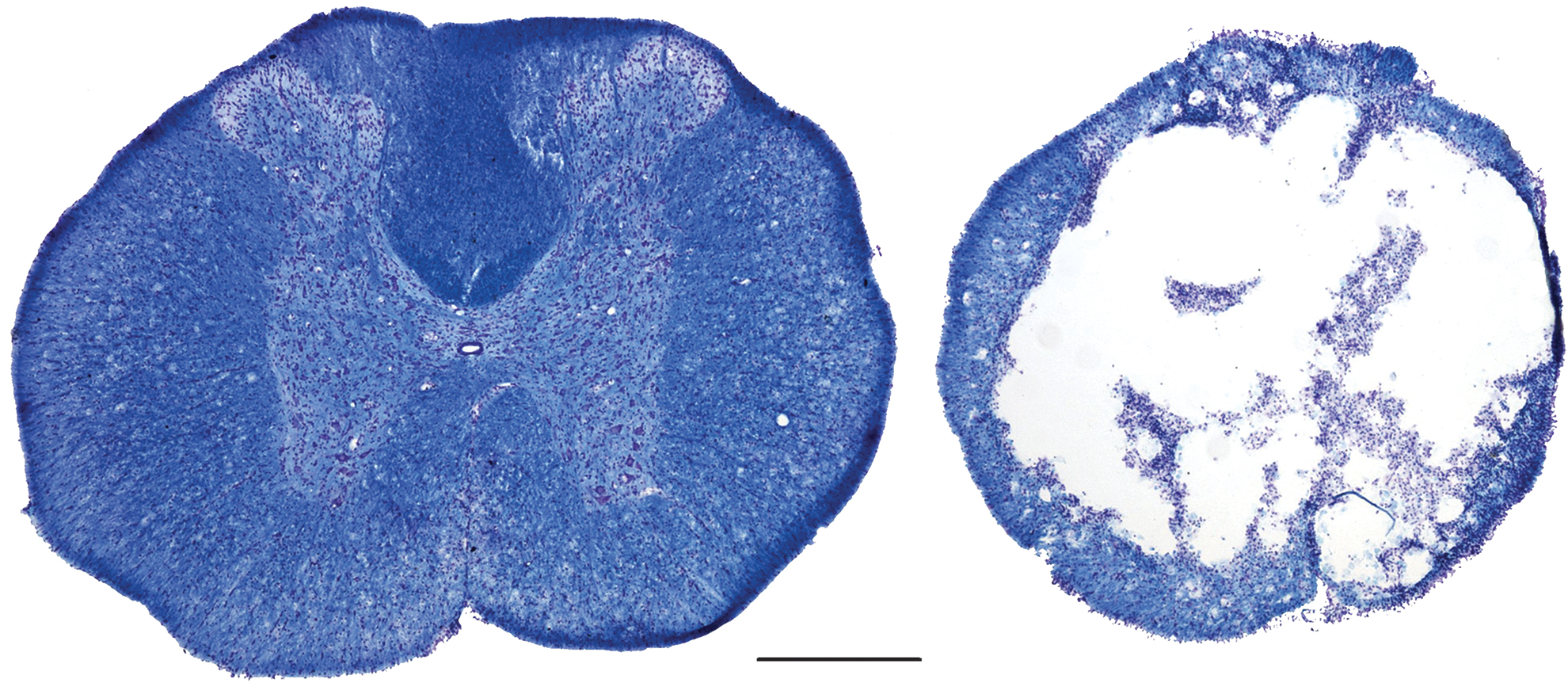
Histological sections showing an intact spinal cord section (left image, 1 cm rostral of epicenter) with a representative section (right image, ∼16% white matter sparing) of the injury epicenter from the same animal. Images taken at 40 × magnification; scale bar is 500 μm.
Body weight gain
The amount of weight gained during the recovery period was not significantly different between contusion (mean = 179 ± 35 g) and sham (mean = 178 ± 22 g) groups (p > 0.05).
Feces data
For the fecal pellet count (Fig. 4A), a significant interaction was found between group (Contusion vs. Sham) and post-injury time-point (p < 0.001). There were no significant differences detected in pellet count for the sham group across the recovery period. At Days 5 (p < 0.001), 6 (p < 0.05), and 7 (p < 0.05), the pellet output of the contusion group was significantly higher than in the sham group. Additionally, there were numerous differences within the contusion group across the recovery period (all at the p < 0.001 level), which are enumerated in Table 1. With respect to fecal wet weight (Fig. 4B), a significant interaction occurred with time post-SCI (p < 0.001). The contusion group was different from the sham group at the 2-, 3-, 21-, 28-, and 42-day time-points. Only the between-group differences are shown in Figure 4B; significant within-group differences are found in Table 1.

Feces measurement results.
Fecal Pellet and Wet Weight within Group Differences
Significant difference from pre-injury, 3, 4, 5, 6, 7, . and 14 days, respectively (p < 0.05).
SCI, spinal cord injury; SEM, standard error of the mean.
EAS EMG data
EMG data quantification (Fig. 5A) and representative EAS responses are shown (Fig. 5B). For response latency, a significant main effect for group (p < 0.05) was found with the contused rats having a longer latency to respond relative to shams. For duration of the EMG excitatory response (Fig. 5A), a significant main effect occurred whereby the contusion group had a shorter mean duration of response than the sham group (p < 0.05). Further, there is a significant main effect of trial (p < 0.001) showing a longer duration in trial 1 than in trials 3 and 4, indicating potential habituation.
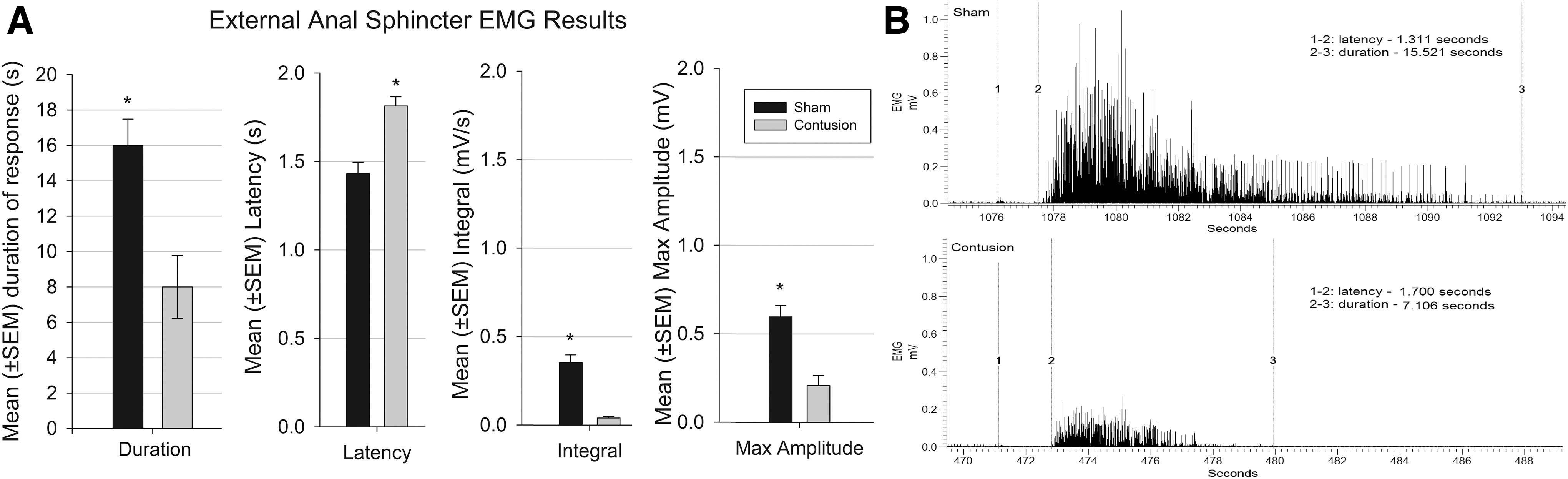
Response of external anal sphincter (EAS) to distension.
Area under the curve (Integral)
A significant main effect (p < 0.01) was present where the contusion group has a smaller integral than the sham (Fig. 5A). For maximum amplitude (Fig. 5A), a significant main effect of group (p < 0.05) with the contusion group having significantly lower maximum amplitude than the sham group.
Anorectal manometry terminal testing
Data quantification results are presented in Figures 6A, 6B, and 6C, with representative traces provided in Figure 7. The baseline pressure (Fig. 6A) for the contused rats was found to be significantly higher relative to surgical shams (p < 0.01). A significant interaction between group and distance (p < 0.001) shows that the significant main effect is due to elevated baseline pressure at 2-, 3-, 4-, and 5-cm depths in the contusion group. With respect to baseline contraction frequency (Fig. 6B), a significantly higher mean baseline frequency was also found for the contused animals. A significant interaction between group and distance (p < 0.001) shows that the significant main effect is due to elevated baseline frequency at 3- and 4-cm depths in the Contusion group. For baseline area under the curve (Fig. 6C), a significant main effect of group (p = 0.001), distance (p < 0.001), and interaction (p < 0.001) were found for depths of 3, 4, 5, and 6 cm. At each of these levels there was a substantial increase in the area under the curve (AUC) after SCI, suggestive of a higher baseline rectal tone at these levels.

Manometry data.
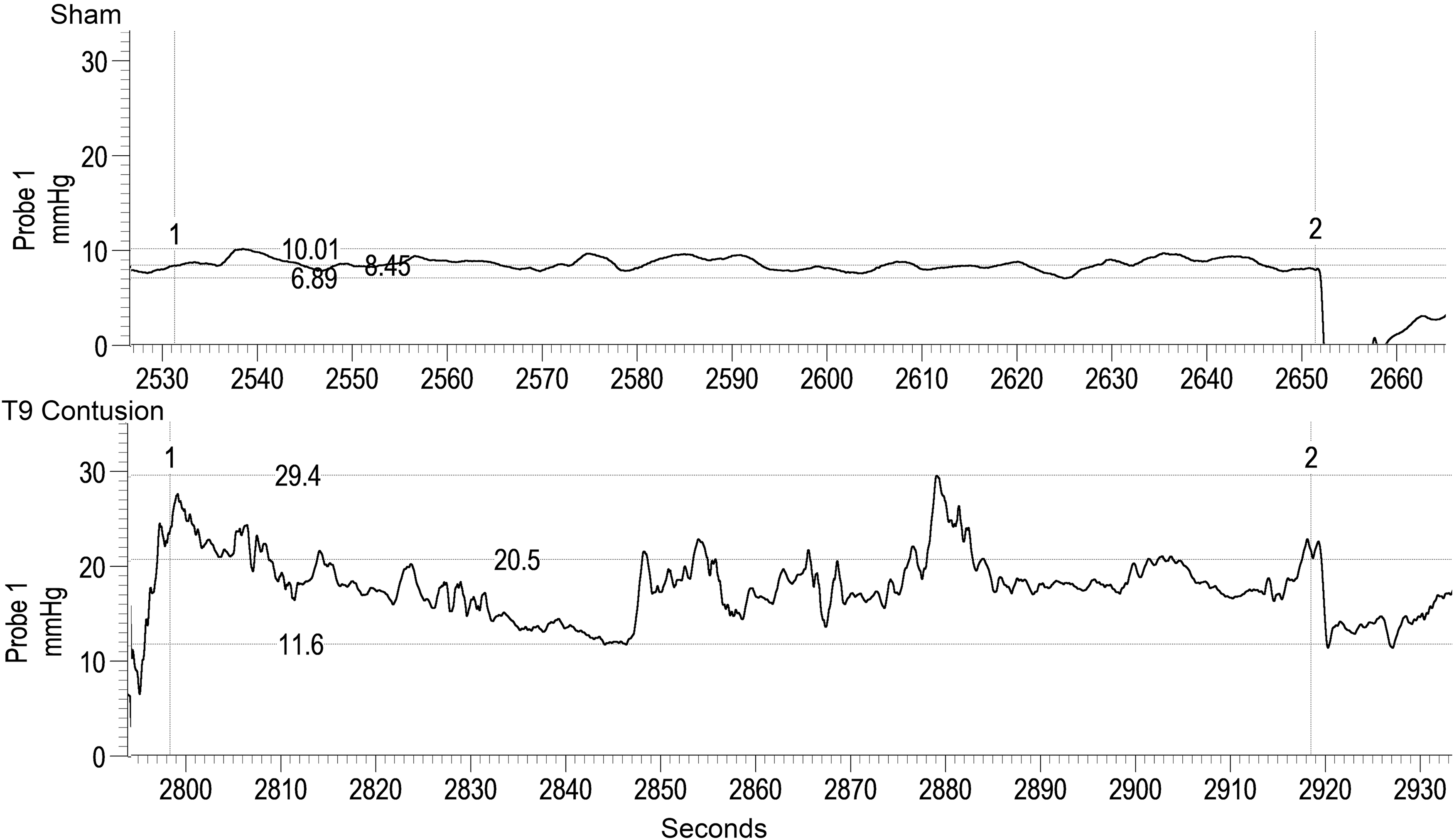
Representative traces of baseline differences between groups during anorectal manometry (ARM) testing. Representative traces were obtained at the 4 cm distance from the anal verge. The baseline period is between the vertical cursors (1 and 2) with the upper and lower values marked by horizontal cursors. The mean baseline pressure is marked by the middle horizontal cursor in each trace. After chronic injury there is increased mean baseline pressure (20.5 vs. 8.45 mm Hg) and increased mean contraction frequency (0.31 vs. 0.16 Hz) respectively, compared to shams.
With respect to pressure effects in response to distension, a significant interaction between group and distance (p < 0.05) occurred due to a high degree of change after distension at 4 cm depth in the contusion group. There were no within-group differences for the sham group, suggesting a consistent response across the organ to distension stimulation. The SCI group had inconsistent responses, which were the most pronounced at 4 cm.
The number of occurrences of a giant contraction (GC; Fig. 8A) was greater (p < 0.001) in the shams, with a significant main effect of distance (p < 0.001). A significant interaction (p < 0.001) between group and distance indicates a greater number of GCs in the sham group at 1-, 2-, 3-, and 5-cm distances from the anal verge. In terms of GC maximum amplitude (Fig. 8B), a significantly higher amplitude was measured for shams, with a significant main effect of distance (p < 0.001) and interaction (p < 0.001) whereby the sham group had higher GC maximum amplitude at 1-, 2-, and 3-cm distances from the anal verge. Representative traces are provided in Figure 9.

Giant contraction (GC) data:
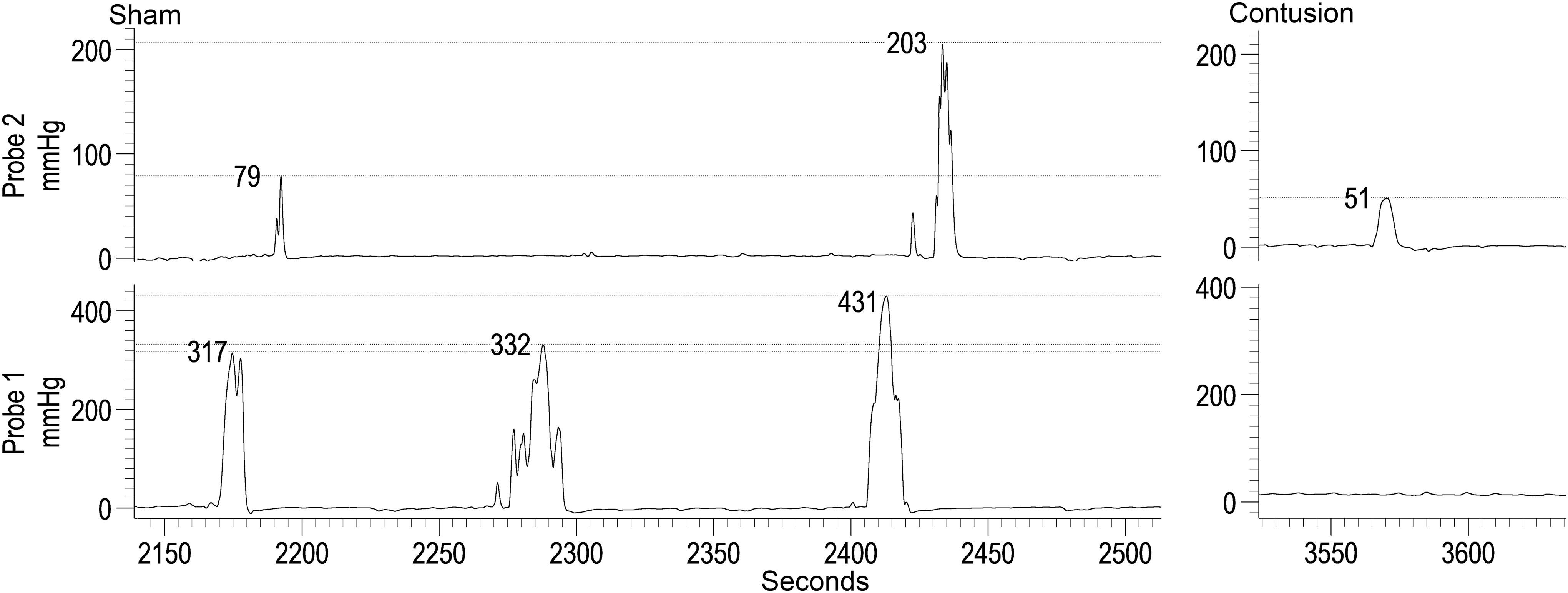
Representative traces showing a series of giant contractions (GCs) in a sham rat (probe 1 is 3 cm and probe 2 is 1 cm from the anal verge). The contusion trace is one of the two examples of GCs from an injured rat (Probe 1 is 4 cm and Probe 2 is 2 cm from the anal verge). The amplitude of each contraction is marked with a horizontal cursor. Note in the shams the reduction in size of the GCs as it propagates from Probe 1 to Probe 2.
Discussion
The data from the present group of experiments are consistent with what has been found in humans after chronic SCI with regard to spastic rectal dynamics at baseline (increased pressure/tone, increased contraction frequency), and is therefore confirmation that the ARM model is appropriate and sensitive to the deficits that occur in the rat contusion model post-SCI. Further, these are the first data showing a disruption in the occurrence and amplitude of colonic giant contraction after chronic T8 moderate/severe contusion SCI in the rat. Note that the injury parameters, BBB scores, and histological sparing percentages are consistent with a moderate to severe contusion per our previously published studies. 47 –49
Assessment of fecal output indicates that there is a significant increase in the number of pellets excreted by the injured animals during a portion of the sub-acute injury period (5–7 days post-injury). These functional data are at odds with previous myoelectric data showing a substantial reduction in activity for the first week after injury. 29 Methodological differences between studies could account for this difference because the previous study utilized a T4 complete transection, while this study investigated a more clinically-relevant T9 incomplete contusion injury. Importantly, the motility index in the previous experiment did not reduce to zero, so the functional outputs shown here may be possible with lowered motility but a substantial increase in the material being ingested.
Fecal pellets from home cages were collected by hand and it was noted that the fecal pellets had a substantial amount of bedding as a component. This suggests that pica, 52,53 ingestion of non-nutritive substances in response to gastrointestinal distress, 54 is a significant component of the post-injury recovery period in rats. Because rats lack the ability to vomit, 54,55 the behavioral response to gastrointestinal distress is to engage in pica. The neural circuits implicated thus far include: vagal afferents, detection of circulating factors via the area postrema and the nucleus of the tractus solitaries (NTS). 55 The role of the NTS seems to be integration of the various inputs and coordination of the numerous components of the emetic response (i.e., esophageal contraction, diaphragm contraction). 55 Evidence of vagal disturbance after SCI is extensive (for review) 45 and suggests the factors necessary to produce pica are present after injury.
Ingestion of bedding is common in rodents and because this phenomenon was not directly measured or manipulated, no solid conclusions can be drawn at this time. The presence of bedding in the feces could be the result of increased ingestion (from pica) or impaired processing of the material in the gastrointestinal tract. Because this behavioral response has not been well characterized after SCI, it represents an important avenue of investigation in GI function after injury.
The data for the wet weight of the fecal pellets indicates a change in fecal composition over the recovery period from lighter pellets (acute period) to heavier pellets (after day 21; Fig. 4B) that occurred in animals with a spinal cord contusion. Later in the recovery period, the lower number of pellets combined with the heavier mass indicates a larger fecal pellet. Larger fecal pellets are indicative of slower motility and constipation, which is consistent with deficits in gastrointestinal motility seen in humans after SCI. 17 –22
Anorectal activity has been shown to be inhibited by descending projections from the nucleus raphe obscurus (nRO) as electrical stimulation of this area reduces spontaneous activity in the colon. 56 After injury, plastic changes that occur in the serotonergic systems cause EAS hyperreflexivity. 57,58 Therefore, a similar upregulation of serotonergic input on colonic contractions may cause pathological activation of the outflow system and subsequent reduction in motility and constipation.
The EAS EMG results indicate that the muscle is compromised after injury. The duration of activity was reduced, the latency to contraction was increased, the integral was reduced, and the maximum amplitude was reduced (Fig. 5). Taken together, the EAS is slower to respond and has a weaker response after injury. These EMG results are somewhat surprising due to the previous studies showing EAS hyperreflexia after SCI, 57,58 in addition to the known spasticity in humans after injury. Previous data from Holmes shows that the EAS muscle has excessive activity after SCI and will contract during defecation of a pellet. 57 Importantly, the previous data came from animals with a T4 complete transection that were tested while awake. The current data is from an anesthetized preparation after a T9 incomplete contusion. Methodological differences and/or severity/location of the injury could account for the difference in responsiveness of the EAS muscle during testing.
Baseline differences in rectal function (increased pressure and contraction frequency; Fig. 6) are consistent with the UMN phenotype of a spastic rectum found in humans. 12 –14 This baseline change in function could represent a functional blockade to the passage of fecal pellets through the rectum. Further, these ARM data are consistent with the finding in humans that after SCI there is a reduction in propulsive and an increase in segmental contractions. 12 As these baseline differences show, the increased colonic activity suggests that this is the case after rodent SCI as well.
After SCI the colon shows changes in histological measures (thickness, collagen deposition, fibrosis), as well as a reduction in myenteric neuron density. 34 Because neuronal nitric oxide synthase (nNOS)-positive enteric neurons provide inhibitory balance to cholinergic excitatory input, 59 it could be the loss of these neurons that is allowing the increased pressure and contraction frequency to occur. Indeed, these neurons seem to be more sensitive to disruption and loss after SCI than other enteric cell types, 60,61 and therefore are a likely mechanism for this elevated activity.
The marked reduction in the occurrence of GCs found in this study is consistent with reduced propulsive contractions found after chronic SCI in humans. A difference in the number of GCs may be more difficult to detect in human studies that are not measured over a long period of time (only ∼2GC/day), 25 but is markedly absent from the rat model which has a much higher number of these events (∼40/h). 25,34 A similar reduction in the number and amplitude of GCs was found after a T3 contusion (300 kDyne 15 sec dwell), 34 which suggests a loss of GC responsiveness is present at multiple injury levels and is a fundamental change in the function of the organ after SCI in rats.
A study by Frias and colleagues found decreased cholinergic responses in ex vivo colonic muscle strips after SCI. 37 Although using different methods, a T3 complete transection with ex vivo testing, the reduction in cholinergic responsiveness may underlie the reduction in GCs that occurs after injury. Investigation of the neurotransmitters involved in GCs showed that a balance of excitatory cholinergic and inhibitory nitrergic inputs is required for GC function. 46 Therefore, changes that occur after SCI in cholinergic responsiveness and nNOS cell populations may be changing this balance and causing the functional changes observed.
Additionally, there is evidence of impaired cholinergic responses in humans with severe chronic idiopathic constipation, 62 indicating this mechanism could be related to the deficits found in this experiment as they have similar phenotypic deficits. A disruption of GCs would create a disruption to the typical function of the organ, in addition to the increased baseline activity, and is likely to contribute to the withholding phenotype seen after chronic SCI (constipation, fecal impaction).
Study limitations include the pica response, which was not systematically measured or manipulated in this study. Therefore, even though there is circumstantial evidence for this being a component of the acute injury period in rats, further studies are needed to confirm this finding using established models of pica (kaolin clay ingestion). Indeed, measurement of all materials being ingested, including food, would help to provide a clearer picture of what is occurring in these animals after SCI. Further, because the wet weight of fecal pellets can be variable due to environmental factors (e.g., desiccation due to bedding, colony humidity), conclusions about this outcome measure should be taken with caution. Subsequent investigations should utilize dry weight as it avoids this variability and allows a better functional comparison for the output of the system in terms of solid material passed by the organ. In addition, because of the depth of probe insertion (11 cm), some tissues may have received supra-physiological stimulation from the probe itself or from probe movement of fecal pellets/obstructions. Therefore, pressure readings at deeper levels (8–11 cm) may not be as reliable as the more distal measurements. Also, it should be noted that meloxicam, the non-steroidal anti-inflammatory drug given to both sham and SCI groups for post-operative pain may confer some neuroprotection. 63
In conclusion, based on the current results, it is evident that the ARM procedure can detect changes in colonic function after SCI in an anesthetized rat, whereas EAS EMG may be more susceptible in the same preparation (different tissue types and sources of innervation). The manometry data are consistent with previous literature in humans (spastic rectum) and therefore is a powerful tool to assess mechanisms and test possible treatments. Additionally, the ARM method is relatively inexpensive, simple to perform, and sensitive enough to detect subtle differences. Further, the addition of metabolic cage (or home cage) measurements allows for comparison between ARM results and functional output as well as a more comprehensive picture of the functioning of the system.
Footnotes
Acknowledgments
We thank James Armstrong and Jason Fell for technical assistance, Christine Yarberry for surgical assistance, Darlene Burke for behavioral assessment assistance, and Dr. Gregory Holmes for recommending the manometry probe.
Funding Information
Funding support for this work was provided by the Craig H Neilsen Foundation (Award #546841), the Leona M. and Harry B. Helmsley Charitable Trust, and the Kentucky Spinal Cord Head Injury Research Trust (KSCHIRT, Grant #17-5).
Author Disclosure Statement
No competing financial interests exist.