
Abstract
Spinal cord injury (SCI) is one of the leading causes of neurological disability and death. So far, there is no satisfactory treatment for SCI, because of its complex and ill-defined pathophysiology. Recently, autophagy has been implicated as protective in acute SCI rat models. Here, we investigated the therapeutic value of a dietary intervention, namely, intermittent fasting (IF), on neuronal survival after acute SCI in rats, and its underlying mechanism related to autophagy regulation. We found remarkable improvement in both behavioral performance and neuronal survival at the injured segment of the spinal cord of animals previously subjected to IF. Western blotting revealed a marked decrease in apoptosis-related markers such as cleaved caspase 3 levels and the bax/bcl-2 ratio in the IF group, which suggested an inhibition of the intrinsic apoptosis pathway. In addition, the expression of the autophagy markers LC3-II and beclin 1 was also increased in the IF group compared with ad libitum fed animals. In parallel, IF decreased the levels of the substrate protein of autophagy, p62, indicative of an upregulation of the autophagic processes. Treatment with 3-methyladenine (3-MA), a selective inhibitor of autophagy, reversed the downregulated apoptosis-related markers by IF. Finally, IF could activate the adenosine monophosphate (AMP)-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) pathway and enhance lysosome function by upregulating transcription factor (TF)EB expression. Altogether, the present findings suggest that IF exerts a neuroprotective effect after acute SCI via the upregulation of autophagy, and further points to dietary interventions as a promising combinatorial treatment for SCI.
Introduction
Spinal cord injury (SCI) is one of the leading causes of neurological disability worldwide. Although the mechanical damage caused by this traumatic event itself cannot be reversed, the secondary injuries that develop in response to factors such as free radical liberation, oxidative stress, and inflammation, and which contribute to worsen SCI outcomes, can be modulated. 1,2 So far however, there is no satisfactory and ubiquitous treatment for SCI, 3 so developing any new strategy that reduces the development of secondary injuries post-SCI would have potential clinical relevance.
Autophagy is a highly conserved homeostatic process involved in cell survival, especially during periods of cell stress. It allows damaged organelles and cytoplasmic agents, such as proteins, to be degraded in lysosomes and then recycled. Autophagy has been implicated in various diseases, including cancer, neurodegenerative diseases, and autoimmune diseases 4,5 An increasing body of evidence suggests that autophagy could also play a pivotal role in controlling the neuronal death process following SCI. In particular, the activation of autophagy could contribute to reducing neuronal loss and apoptosis post-SCI. 6 –8
Dietary restriction regimes, such as calorie restriction and intermittent fasting (IF), have been shown to increase life span and to remarkably reduce the incidence of cardiovascular diseases, diabetes, and cancers. 9,10 Calorie restriction typically involves a reduced calorie intake ranging from 20% to 40%, whereas IF generally involves fasting for 24 h followed by ad libitum (AL) access to food for 24 h. Both approaches have been reported to have positive outcomes in animal models of traumatic brain injury, incomplete cervical SCI, and thoracic contusion injury, 11 –13 but the underlying mechanisms remain to be established. Starving is a common stimulator of autophagy and food restriction has been demonstrated to induce a dramatic upregulation in neuronal autophagy. 14 Moreover, a long-term calorie-restricted diet can induce a marked upregulation of autophagy-related indicators, such as beclin-1 and microtubule-associated protein light chain 3 (LC3). 15 In a preliminary study, we found that IF could modulate autophagy after SCI, 16 but the mechanistic basis for IF's neuroprotective action remains unknown.
In the present work, therefore, we directly addressed the question of whether IF stimulates neuronal autophagy in a rat model of experimental SCI. Our data provide evidence that IF can upregulate the overall autophagic flux both by inducing autophagy activation via an adenosine monophosphate (AMP)-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR) /p70s6k pathway and by improving lysosome function by promoting transcription factor (TF)EB expression, which plays a pivotal role in mediating its neuroprotective action post-SCI.
Methods
Animals
Male Sprague–Dawleyrats (200–220 g, n = 105), obtained from Beijing HFK Bioscience Cooperation, were housed in a standard environment with two to three animals per cage at a room temperature of 23 ± 3°C, humidity of 55 ± 3%, on a 12 h light/dark cycle. All animal procedures were approved by the Medical Ethics Committee of the China Medical University in accordance with National Institutes of Health (NIH) guidelines for the care and use of laboratory animals (NIH Publication No.85/23). Every effort was made to minimize the number and suffering of animals.
Feeding regime
The rats were randomly divided into two groups, an AL feeding group and an IF group. The animals in the AL group had free access to food and water during the entire period of the experimental procedures until euthanasia, whereas the animals in the IF group got continuous free access to water and had free access to food for the first 2 weeks and then were deprived of food for an entire day every other day until euthanasia. Surgical interventions were performed in the morning subsequent to an overnight period of feeding (Fig. 1).

Feeding and experiment protocols in ad libitum and intermittent fasting groups. Color image is available online.
Weight drop contusive SCI
The procedure of contusive SCI rat model was performed as described by Jeong and coworkers. 13 Briefly, rats were deeply anaesthetized with sodium pentobarbital (65 mg/kg, i.p.) until no reflex could be evoked by pinching the tail. The skin was incised above the T9-T11 vertebrae and the muscles were gently retracted on both sides to expose the T10 vertebra. A laminectomy was performed with rongeur forceps to remove T10 lamina and expose the dorsal part of the spinal cord. Rats then received a contusive injury with a 10 g impactor device (diameter: 2 mm) which was dropped from a height of 2.5 cm onto the surface of the exposed spinal cord. At the impact, ectopic nerve discharges briefly evoked hindlimb twitch and tail flap but then paralysis occurred. Rats in the sham group were subjected to the same experimental protocol up until laminectomy, but did not receive the contusion injury. Muscles and skin were sutured in layers, and rats were allowed to fully recover from the surgery in an incubator maintained at 37°C. Animals were intraperitoneally administered with penicillin G (10,000 U/day) for 3 days post-surgery. The SCI rats received manual bladder emptying twice daily until bladder function recovered.
Chemicals and kits
Anti-lysosomal-associated membrane protein 1 (Lamp 1) monoclonal antibody (#ab24170) was purchased from Abcam (Cambridge, MA, USA). Antibodies against LC3B (#2775S), cleaved caspase 3 (#9661) and Atg7 (#8558) were obtained from Cell Signaling Technology (Beverly, MA, USA). The antibody against TFEB (A303-673A) was obtained from Bethyl Laboratories (Montgomery, TX, USA). The antibody against NeuN (#MAB377X) was purchased from Sigma (St. Louis, MO, USA). Antibodies against p62 (#18420-1-AP), Bax (#50599-2-Ig) and Bcl-2 (#12789-1-AP) were obtained from Proteintech Group, Inc. (Chicago, IL, USA). Anti-GAPDH (#Ab103-01) antibody was purchased from Vazyme Biotech Co. (Nanjing, China). The horseradish peroxidase (HRP)-conjugated anti-mouse (#ZB-2305) and anti-rabbit (#ZB-2301) antibodies were obtained from ZSGB-BIO (Beijing, China). Cathepsin B and cathepsin D fluorometric assay kits (#K140-100, K143-100) were obtained from Biovision (Milpitas, CA, USA).
3-Methyladenine (3-MA) and chloroquine preparation and treatment
3-MA is an inhibitor of phosphatidylinositol 3-kinases (PI3K) that could block autophagosome formation by inhibiting class III PI3K. 17 3-MA was dissolved in dimethylsulfoxide (DMSO) (10 mg/mL), and the rats received 3-MA (10 mg/kg, i.p.) daily after SCI until euthanasia. Chloroquine inhibits autophagic flux by inhibiting autophagosome and lysosome fusion. Chloroquine was dissolved in phosphate buffered saline (PBS) (10 mg/mL) and rats received chloroquine (10 mg/kg, i.p.) 2 h before euthanasia, as described by Yao and coworkers. 18
Behavioral testing
The Basso, Beattie and Bresnahan (BBB) locomotion rating scale and the inclined plane test were applied to assess the recovery of locomotion function at 0, 1, 3, 7, 21, and 28 days after SCI. 19 Scores on the BBB scale range from 0 to 21, with 0 representing complete paralysis and 21 representing normal locomotion. Two trained researchers made BBB score evaluations in a blinding manner. The inclined plane test was conducted according to previous studies. 20,21 The maximum angle that rats could withstand for at least 5 sec was taken as the inclined plane score. Mean scores were then used for further analysis.
Hematoxylin-Eosin (HE) staining
Rats were euthanized 7 days after surgery with an overdose of sodium pentobarbital, and were transcardially perfused with 0.9% NaCl followed by 4% paraformaldehyde. The spinal segments containing the lesion (1 cm, centered at the injury epicenter) were excised, post-fixed in 4% paraformaldehyde overnight, and later dehydrated in 30% sucrose until the samples sank to the bottom. Transverse sections (15 μm) cut with a cryostat (Leica, Germany) were used for HE staining. The sections were stained with hematoxylin solution for 10 min followed by 1% acid alcohol for 30 sec, soaked in running water, and stained with eosin solution for 1 min. After rinsing in running water for 5 min, the sections were dehydrated in a grade alcohol series and cleared in xylene. The sections were observed under a light microscope and the images were captured on a Nikon 300 microscope.
Immunohistochemistry
The frozen sections 5 mm rostral to the injury epicenter with a thickness of 15 μm were obtained 7 days after SCI and used for immunohistochemical staining. After the sections were dried at room temperature, they were incubated with blocking buffer (5% normal goat serum) for 1 h at room temperature and then incubated with primary antibodies at 4°C overnight. To assess apoptosis-related changes, double labeling of anti-cleaved caspase 3 (1:100) and anti-NeuN (1:500) was performed. The sections were incubated with Alexa Fluor 488 conjugated goat anti-mouse (1:200) and Alexa Fluor 594 conjugated goat anti-rabbit (1:200) secondary antibodies for 2 h at room temperature. The nucleus was visualized with a 4,6-diami-dino-2-phenylindole (DAPI) solution. All images were captured under a fluorescence microscope. The number of double-labeled neurons of cleaved-caspase 3 was counted at six randomly chosen fields restricted to the gray matter area per section. Five random sections in the same range with 1 mm interval from each rat were analyzed, and three rats in each group were included. The total number of positive neurons was determined in a blinding manner.
Western blot analysis
Spinal cord tissues of ∼1 cm centered on the injury site or corresponding site in the sham group were incubated on ice with radioimmunoprecipitation assay (RIPA) buffer containing protease inhibitor and phenylmethylsulfonyl fluoride (PMSF) for 30 min and centrifuged at 12,000g (4°C, 30 min). The final protein concentration was determined from the supernatant with a BCA kit (Beyotime Biotechnology, Shanghai, China). Similar amounts of protein samples (20–40 μg) were subjected to electrophoresis and transferred onto a polyvinylidene difluoride (PVDF) membrane (Millipore, CA, USA). The membranes were blocked with 5% non-fat milk or 5% bovine serum albumin (BSA) in tris-buffered saline (TBS) with 0.1% Tween 20 for 2 h and then incubated with primary antibodies at 4°C overnight. The membranes were then incubated with corresponding HRP-conjugated secondary antibodies for 2 h at room temperature. Immunoreactivity was detected with an enhanced chemiluminescence detection kit (ECL kit, Millipore). Densities of bands were measured with ImageJ Software (NIH) and were normalized to a loading control n-Glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Protease activity assay
The catalytic activities of cathepsin B and cathepsin D were determined according to the instructions of the assaying kits. Briefly, 10 mg of spinal cord tissue was homogenized in 200 μL lysis buffer on ice. Fifty microliters of lysate was transferred onto 96-well plates, mixed with reaction buffer and substrate, and incubated at 37°C for 1 h. The activities were detected under a microplate reader with 400 nm excitation/505 nm emission and 328 nm excitation/460 nm emission filters, respectively, and further normalized to the protein concentration.
Statistical analysis
Data were expressed as means ± standard deviation (SD) with at least three independent experiments. Unpaired student t test or one-way analysis of variance (ANOVA) followed by Bonferroni's multiple group comparison were applied for group comparisons. P values <0.05 were considered statistically significant.
Results
IF slows weight increase and promotes motor function recovery after SCI
Before the onset of the IF regime, there was no difference in the weight of the rats in both groups. However, after 8 weeks of the IF regime, despite the weight gain, the weight of the rats in the IF group (n = 10) was notably lower than that in the AL group (n = 10). The recovery of locomotor function was evaluated using the BBB scoring method applied before (day 0) and 1, 3, 7, 14, 21, and 28 days after SCI. As is shown in Figure 2A, within 3 days after contusion, there was no significant difference in the BBB score between the AL+SCI group (n = 7) and the IF+SCI group (n = 7). However, the BBB score was significantly higher (p < 0.05) in the IF+SCI group than in the AL+SCI group by day 7 after contusion, and remained higher until the end-point of the study. Similarly, we observed that the inclined plane test scores in the IF+SCI group were significantly higher (p < 0.05) than those obtained in the AL+SCI group at 7, 14, 21, and 28 days after AL+SCI (Fig. 2B). The above findings therefore indicated that IF can improve the recovery of locomotor and postural function after SCI.
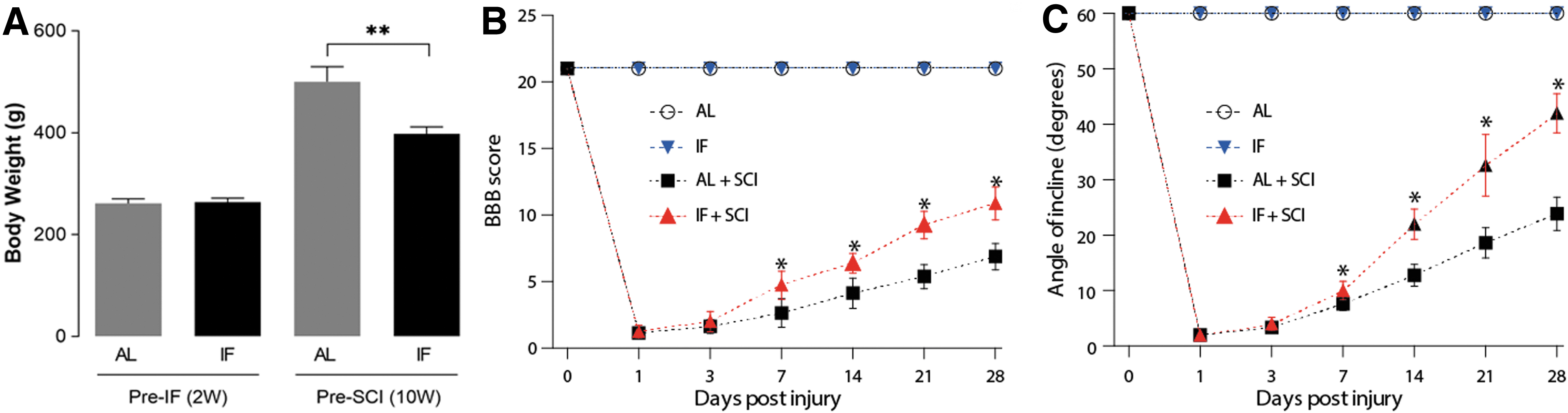
Intermittent fasting (IF) slows weight increase and promotes motor function recovery after spinal cord injury (SCI). The body weight of rats before the IF regime and after 8 weeks of IF regime in both groups, n = 10, **p < 0.01
IF attenuates apoptotic cell death caused by SCI
To determine whether IF affects cellular apoptotic mechanisms following SCI, Western blots were employed to quantify the levels of apoptosis-related markers in the different experimental animal groups. As is shown in Figure 3, both the AL+SCI and IF+SCI groups showed a significant increase (p < 0.05) in the expression of the pro-apoptotic proteins bax (Fig. 3A) and cleaved-caspase 3 (Fig. 3B), and a significant decrease (p < 0.05) in the anti-apoptotic protein bcl-2 (Fig. 3A) level, whereas there was no significant difference between the AL (n = 3) and IF (n = 3) groups in terms of the expression of these markers. Altogether, these changes suggested the occurrence of an activation of apoptosis induced by SCI at day 7 after injury. Moreover, we found that the bax/bcl-2 ratio (Fig. 3A), as well as the cleaved-caspase 3 (Fig. 3B) level were significantly lower (p < 0.05) in the IF+SCI group (n = 3) than in the AL+SCI group (n = 3), suggesting that IF attenuates the degree of neuronal apoptosis induced by SCI.
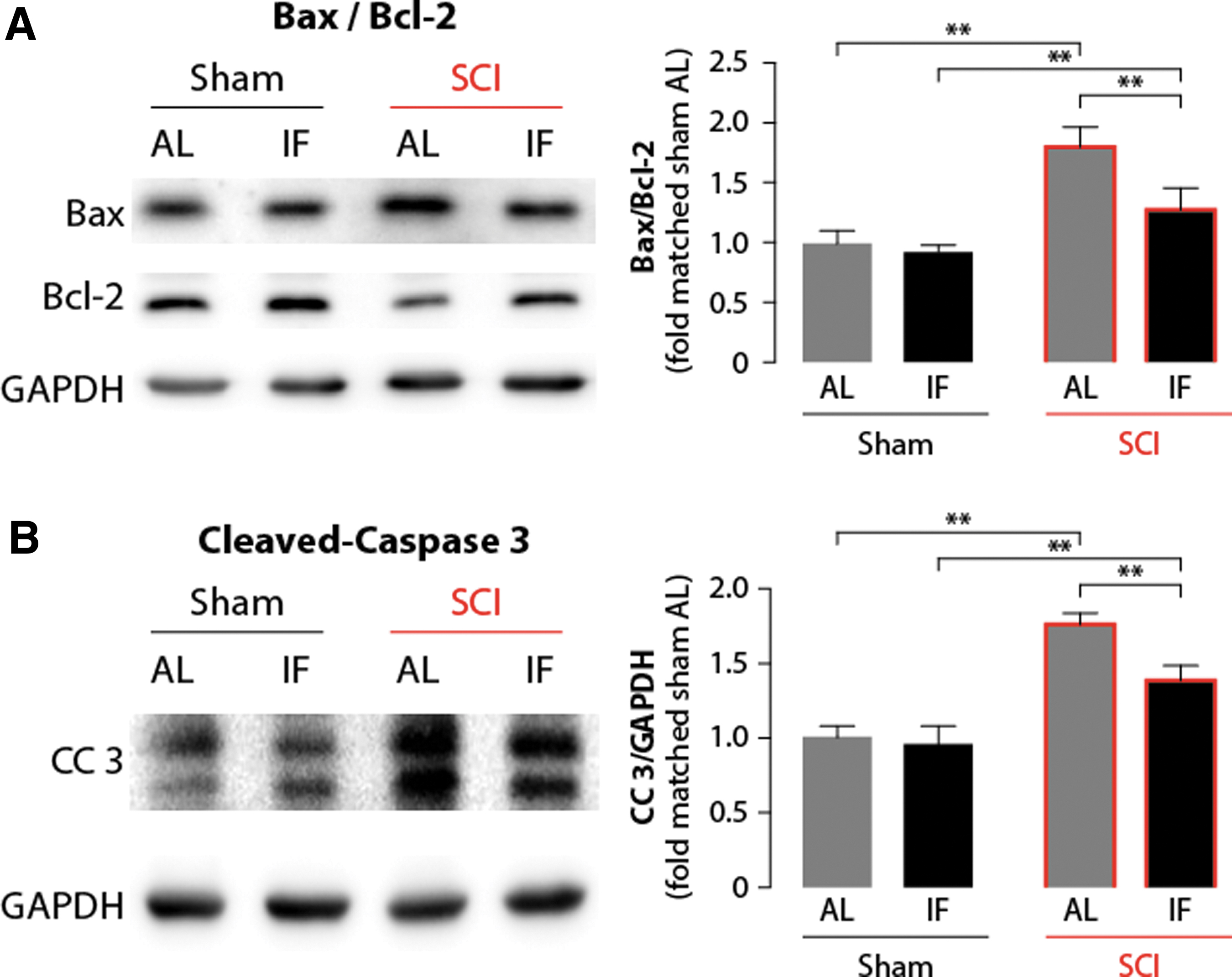
Intermittent fasting (IF) attenuates spinal cord injury (SCI)-induced cell apoptosis. Western blot analysis and quantification of apoptosis-related markers of bax, bcl-2
IF promotes autophagy activation after SCI
To evaluate the effect of IF on autophagy, rats were euthanized 1 week after SCI, and Western blotting was performed to evaluate the expression of autophagy-related proteins, including LC3-II, p62 and Atg7. First, the expression level of Atg7, a key factor in the biogenesis of autophagosomes, was found to be significantly increased in the IF, AL+SCI, and IF+SCI groups compared with the AL group (n = 3) (p < 0.05; Fig. 4A). In addition, when compared to the AL+SCI group, the IF+SCI group, showed a higher level of Atg7 that attained statistical significance (p < 0.05). Because the conversion of LC3-I to LC3-II is critical in the formation of autophagosomes, LC3-II is also considered to be a marker of autophagosome formation. As is shown in Figure 4B, the expression levels of LC3-II among the experimental groups were similar to the Atg7 patterns, further suggesting that IF can indeed upregulate autophagy.
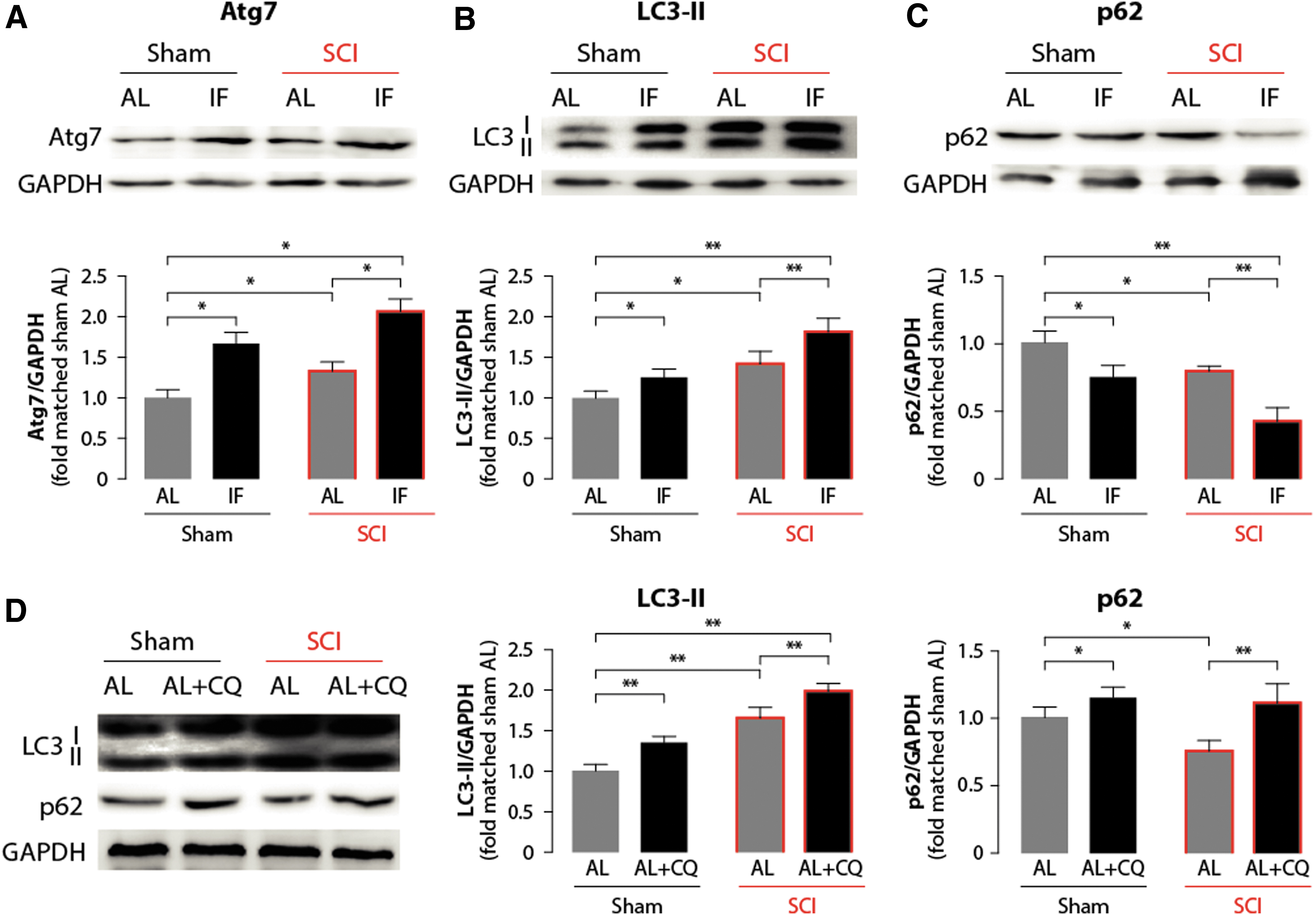
Intermittent fasting (IF) promotes autophagy activation after spinal cord injury (SCI). Autophagy-related markers, Atg7
Because it is a protein mediating the delivery of ubiquitinated proteins to autophagosomes for degradation, which functions as a substrate for the autophagic process, the expression level of the p62 protein is considered to be an indicator of the state of autophagic flux. 22 Compared with the AL group, the expression level of p62 decreased in the IF, AL+SCI, and IF+SCI groups. Here again, we found a significant difference between the IF+SCI and AL+SCI groups, with the lowest p62 level being observed in the former group (p < 0.05; Fig. 4C). Impairment of the autophagic flux post-SCI was further explored by applying chloroquine in vivo as described previously. 23 As is shown in Figure 4D, a single chloroquine treatment in the AL group (n = 3) increased levels of LC3-II and p62, suggesting an intact state of autophagic flux. Chloroquine treatment after SCI in the AL group (n = 3) also induced a significant increase in LC3-II expression and additionally reversed SCI-induced downregulation of p62 (p < 0.05; Fig. 4D). This finding indicates, first, that the autophagic flux state was not impaired after SCI, and second, that IF could indeed further enhance the autophagic flux.
IF can improve lysosome function via an upregulation of TFEB expression
We further assessed whether IF impacts downstream lysosome expression and function. Western blot analysis revealed that both IF and SCI increase the expression of the lysosome-membrane marker Lamp 1 which, in addition, was more important in the IF+SCI group (n = 3) than in the AL+SCI group (n = 3) (Fig. 5A). As lysosome-related protein biogenesis is mainly controlled by the transcriptional factor TFEB, 24 we also quantified its expression (Fig. 5B). The results obtained with Western blot showed changes that parallel the Lamp 1 expression across the different groups. Finally, we examined the lysosome function by assessing the activity of two main proteases acting within the lysosome; namely, cathepsin B (Fig. 5C) and cathepsin D (Fig. 5D). The results revealed that IF could increase the activity of both proteases compared with AL, but there was no statistical difference between the AL group (n = 3) and AL+SCI group (n = 3), suggesting that SCI itself had little impact on the lysosomal protease. In addition, compared with the AL+SCI group, the IF+SCI group showed a significantly enhanced activity (p < 0.05) of both cathepsin B and cathepsin D. Collectively, these results suggest that IF could increase lysosome function, possibly via an upregulation of TFEB.
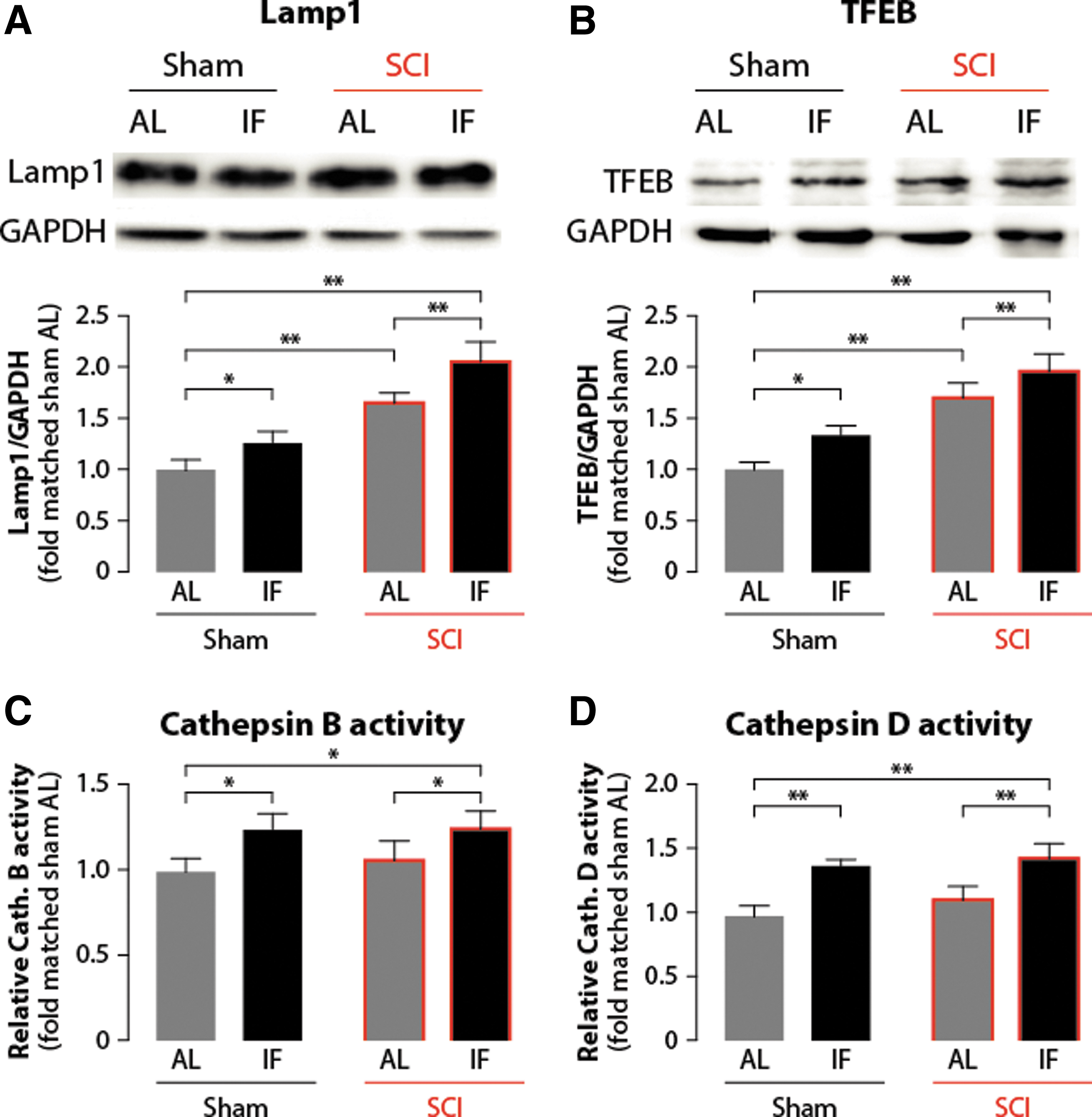
Intermittent fasting (IF) improves lysosome function by upregulating transcription factor (TF)EB expression. Western blot was applied to show the effect of IF on lysosome-related proteins, including lysosomal-associated membrane protein 1 (Lamp 1)
The neuroprotective effect of IF after SCI is related to autophagy modulation
3-MA inhibits the early formation of autophagosomes by inhibiting the production of phosphatidylinositol-3-phosphate. 17 Therefore, we next administrated 3-MA to inhibit autophagy at an early post-injury stage to assess whether IF exerts an anti-apoptotic effect by regulating autophagy. As is shown in Figure 6A, 3-MA downregulated the expression of LC3-II and reversed the drop in p62 expression after SCI (p < 0.05), suggesting that it successfully inhibited autophagosome formation. Western blot analysis of apoptosis-related proteins showed that 3-MA led to an increased expression of cleaved-caspase 3 compared with the IF+SCI group (n = 3). In addition, we found that the ratio of bax/bcl-2 was also increased in the IF+SCI +3-MA group (n = 3) (p < 0.05; Fig. 6B). Immunofluorecence double staining targeting NeuN and cleaved-caspase 3 also revealed a significant increase in the presence of double-stained cells in the IF+SCI +3-MA group (n = 3) compared with the IF+SCI group (n = 3) (p < 0.05; Fig. 7A, B). Further, HE staining revealed a larger lesion size in the IF+SCI +3-MA group (n = 3) than in the IF+SCI group (n = 3) (Fig. 7C). Altogether, the above results suggest that inhibiting autophagy counteracts the neuroprotective effect of IF after SCI.

Autophagy inhibition impairs the anti-apoptotic effect of intermittent fasting (IF) on spinal cord injury (SCI). Western blot and quantification of LC3 and p62 to assess the autophagy inhibitory effect of 3-methyladenine (3-MA) after SCI

Autophagy inhibition impairs the neuroprotective effect of intermittent fasting (IF) on spinal cord injury (SCI). Immunofluorescent double staining of NeuN and cleaved-caspase 3 was applied to study the effect of autophagy inhibition on the neuroprotective effect of IF in SCI
The AMPK/mTOR/p70s6k pathway is involved in IF-induced autophagy
In a final step, to further explore how IF regulates autophagy, upstream regulators of autophagy, including AMPK, p-AMPK, mTOR, p-mTOR, p70s6k and p-p70s6k, were assessed using Western blot analysis. AMPK is a serine/threonine protein kinase that is reported to mediate the neuroprotective effect of calorie restriction in Parkinson's disease. 25 AMPK can activate autophagy by negatively regulating mTOR, a key regulator of autophagy, and its phosphorylation (p-AMPK) shows a reciprocal relationship with autophagy. 26 p70s6k is a downstream target of mTOR that in turn reflects the activity of mTOR. The results showed that IF significantly upregulates the ratio of p-AMPK/AMPK, and induces a significant decrease in the ratio of p-mTOR/mTOR as well as that of p-P70S6K/P70S6K (n = 3) (p < 0.05; Fig. 8), suggesting that IF could activate AMPK and lead to mTOR inhibition, and in turn to autophagy activation.

Involvement of adenosine monophosphate (AMP)-activated protein kinase (AMPK)/mammalian target of rapamycin (mTOR)/p70s6k pathway in intermittent fasting (IF-induced autophagy. Western blot and quantification of AMPK, p-AMPK, mTOR, p-mTOR, p70s6k, and p-p70s6k was applied to show the effect of IF on AMPK/mTOR/p70s6k signaling in spinal cord injury (SCI) rat models, with n-Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) serving as an internal control. Data are presented as means ± standard deviation (SD), n = 3, *p < 0.05, **p < 0.01. Color image is available online.
Discussion
The current study has revealed that IF prior to an SCI in rats can effectively alleviate resultant neuronal death and improve functional recovery. Altogether, our data indicate that the increased neuroprotective action mediated by IF is closely related to a modulation of autophagy activation.
Dietary interventions such as IF or calorie restriction entail a reduction in total daily calorie intake and have been demonstrated to improve the average lifespan in an invertebrate (nematodes) and in vertebrates including rodents, primates, and humans. 27 –29 In addition to the beneficial effects of IF in metabolic diseases, cardiovascular diseases, and cancer, it can also exert a protective effect following acute insults to the central nervous system, such as stroke or traumatic brain injury. 30 –32 It has also been reported that IF initiated after cervical and thoracic SCI results in preserved neuronal survival, reduced lesion volume, increased sprouting, and improved functional recovery in Sprague–Dawley rats. 12,13,33 However, in contrast to Sprague–Dawley rats, IF in C57BL/6 mice following a moderate to severe thoracic SCI does not improve recovery. The authors suggested that mice and rats could have a different whole-body adaptive responses to fasting, in particular related to the genes involved in ketone synthesis. 34 With respect to the body weight change by IF intervention, Plunet and coworkers 12 reported that the IF group did not lose weight on the restricted food regimen throughout the experiment and continued to gain weight, albeit at a lower rate than the AL group. And according to the study by Jeong and coworkers, 13 the animal body weight did not differ between the pre-IF and the AL group at the time of the injury, and the pre-IF and post-IF groups gained weight over time, but less than the AL group did. In the current study, we observed that IF could slow the weight increase and lead to a notable difference in weight between the two groups as well as an improvement in hindlimb motor performance, which was indicated by higher BBB and inclined plane test scores. The protein bax is involved in the induction of apoptosis, whereas bcl-2 exerts an anti-apoptotic effect, and cleaved-caspase-3 is a hallmark of apoptotic cell death acting as the final executor of apoptosis. 35 Here, using Western blot analyses, we found that levels of cleaved-caspase 3 and the ratio of Bax/Bcl-2 were remarkably low in the IF group of SCI animals. In this latter group, intrinsic apoptosis activation was particularly inhibited, further confirming the neuroprotective effect of IF after SCI.
Previous studies investigating the underlying mechanisms of IF have mainly focused on microglial and apoptosis inhibition, 36 the anti-inflammatory process involving the regulation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) transcriptional activity, 37 or anti-oxidative stress through an increase in mitochondrial uncoupling levels and activity. 38 Thus far, whether IF also exerts its neuroprotective effect via the regulation of autophagy has remained to be elucidated.
Autophagy is an essential lysosome-dependent intracellular catabolic mechanism that maintains cellular homeostasis by clearing damaged organelles and proteins. 39 It has been implicated in various pathological conditions including cancer, 40 heart diseases, 41 and neurodegenerative diseases. 42 The conversion of the protein LC3-I to LC3-II is a hallmark of autophagy induction during autophagosome formation, and Atg7 is a key factor in the biogenesis of autophagosomes. 43 In the present study, we observed a marked upregulation of autophagy markers such as LC3-II and Atg7 in the IF+SCI group compared with the AL, IF, and AL+SCI groups, suggesting that pre-lesion IF and SCI synergistically potentiate the activation of autophagy. In contrast, decreasing autophagy activation (as in our 3-MA experiments) resulted in an increase in the level of apoptosis and more tissue damage. Therefore, it is very likely that the neuroprotective action of IF is at least partially mediated by the upregulation of autophagy.
Currently, the role of autophagy in SCI remains controversial. Both pharmacological activation of autophagy and inhibition of autophagy have been reported to be neuroprotective in animal models of traumatic SCI. A number of studies have reported that upregulating autophagy with specific activators such as rapamycin could promote functional recovery after SCI. 44 –46 On the other hand, autophagy inhibition has also been revealed to yield protective effects on neuronal survival. 47,48 It is possible that these apparently contradictory roles of autophagy reported after SCI could be not only because of the different injury models and severities studied, but also, the steady state of autophagy at the time of injury may partly account for such discrepancy. In the current study, we applied the acute weight drop contusive SCI rat model to study the activation of autophagy and the state of autophagy though non-pharmacological means – by IF – which could improve the lysosome function and the overall autophagic flux quality.
In a previous study, we showed that autophagic flux plays a pivotal role in neuronal survival. 23 Here, we used chloroquine to detect the in vivo state of autophagic flux, which can raise the lysosomal pH and hence inhibit the fusion between autophagosomes and lysosomes, thus preventing the maturation of autophagosomes into autolysosomes and causing a blocking of the late step of autophagy. 49 As p62 directs ubiquitinated cargo to autophagosomes for degradation, and is degraded with its cargo, 50 the level of p62 reflects the degree of degradation of autophagosomes and cell clearance. We found that under chloroquine treatment, the level of LC3-II is increased because of a reduced turnover. Moreover, the level of p62 is also increased, indicating that autophagic flux in our SCI model was not impaired. In this situation, we did not additionally block the autophagic flux in IF groups to explore whether the neuroprotective effect was associated with autophagic flux modulation. However, it should be noted that with the application of chloroquine in vivo to assess autophagic flux, we could not completely rule out the possibility that a partial blockade of autophagic flux was occurring. It is also possible that IF could achieve its neuroprotective effect by reversing a partial blockage of autophagic flux. Further investigation based on the use of real-time monitoring of autophagic flux in vivo will be necessary to further clarify this issue.
TFEB is a member of the microphthalmia-transcription E subfamily of basic helix-loop-helix leucine zipper transcription factors. TFEB contributes to the regulation of lysosomal biogenesis by positively regulating genes of the coordinated lysosomal expression and regulation (CLEAR) network that includes genes associated with lysosomal hydrolases, lysosomal membrane proteins, and components of the vacuolar-type H+-ATPase (V-ATPase) complex. 51,52 In addition, TFEB regulates the expression of autophagy genes, and TFEB downregulation with RNAi results in a decrease of LC3-II levels. 24 Lysosomal digestion is the final stage of autophagy, and evaluating its efficacy is a crucial part of the evaluation of autophagic flux, or completion of an autophagy process. 49 In other words, autophagy relies strictly on lysosomal function, which is regulated by TFEB. In a previous study, we showed that promoting lysosome biogenesis through TFEB overexpression could protect SH-SY5Y cells against 6-hydroxydopamine (6-OHDA)-induced neurotoxicity. 23,53 Interestingly, fasting-induced TFEB signaling was shown to be linked with lysosome homeostasis in mouse livers. 54 In addition, TFEB-mediated transcriptional priming of the autophagy-lysosome machinery mediates the beneficial effects of fasting-induced autophagy in myocardial ischemia-reperfusion injury. 9 IF was also shown to preserve organelle quality via the autophagy-lysosome pathway to enhance beta cell survival, and stimulates markers of regeneration in obesity-induced diabetes. 55 Here, the upregulation of TFEB is consistent with the expression of autophagy markers such as LC3II and Atg7 after SCI. In addition, we also found that IF could enhance lysosome function as suggested by increased cathepsin activities and Lamp 1 expression. Collectively, these findings suggest that IF could enhance the overall autophagic flux process, from autophagy activation to lysosome function.
Finally, we also explored how IF could upregulate TFEB to enhance autophagy after SCI. Recent studies suggest that TFEB is regulated by mTOR, and mTOR can directly phosphorylate TFEB on the lysosomal surface and avoid its nuclear translocation under normal conditions. 56 Under nutrient starvation or energy stress, activated AMPK (p-AMPK) leads to phosphorylation and activation of the TSC1/2 complex, which inhibits mTOR activity. 57 AMPK is a serine/threonine protein kinase that not only is an important metabolic sensor of energy balance at the cellular and organism levels, but also plays an important role in neuronal development by modulating neurite outgrowth. 58 Previous studies showed that AMPK/mTOR pathway activation can stimulate autophagy and exert neuroprotective effects following SCI. 20,59 Here we found that IF significantly upregulates the ratio of p-AMPK/AMPK and the level of TFEB, while decreasing the ratios of p-mTOR/mTOR and p-p70s6k/p70s6k, suggesting that IF could promote autophagy possibly through the AMPK/mTOR/TFEB pathway. Further work will be needed to elucidate the upstream regulatory mechanisms.
There are several limitations in the current study. As we focused on the effect of IF on autophagy modulation, we mainly studied the histological and biochemical changes at 7 days after SCI, as previous studies reported most notable changes in autophagy and apoptosis-related markers at day 7. A prolongation of the study to ≥28 days could better elucidate the dynamic changes of behavior, autophagy, and apoptosis as well as the long-term effect of autophagy inhibition on tissue injury after SCI. Moreover, we did not test the blood glucose, ketones, or neuropeptides that are related to metabolism in different groups, which might have helped elucidate the underlying mechanism behind the beneficial effect of IF, and which requires further research. 33,60
Conclusion
In conclusion, our results show that IF, by modulating autophagy activation, exerts a neuroprotective action after SCI and improves the recovery of posturolocomotor function. Moreover, IF appears to enhance lysosome expression and function by upregulating TFEB and improving the overall autophagic flux state (schematic representation shown in Fig. 9). Future studies will be required to trace autophagic and apoptosis processes over time to decipher the dynamic of the IF-mediated regulation of cellular responses post-SCI. Additionally, further work will be needed to understand whether and how IF could also regulate other forms of autophagy (for example, chaperon-mediated autophagy) after SCI.

Schematic representation of the mechanisms underlying the neuroprotective effect of intermittent fasting (IF) on spinal cord injury (SCI). IF can activate adenosine monophosphate (AMP)-activated protein kinase (AMPK), which weakens the inhibitory effect of mammalian target of rapamycin (mTOR) on autophagy activation. In addition, IF upregulates the level of transcription factor (TF)EB, which contributes to lysosomal biogenesis and function. IF improves the overall autophagic flux by upregulating autophagy induction and promoting lysosome function, which thereby ameliorates neuronal apoptosis after SCI.
Footnotes
Acknowledgment
We thank Dr. John Simmers for his helpful comments on the manuscript and its editing.
Authors' Contributions
W.Y designed and performed experiments, analyzed data, and drafted the manuscript; W.Y., X.H., and X.L. performed experiments; D.M. and J.L. analyzed data; G.B. analyzed data and drafted the manuscript; Y.Z. designed the study and drafted the manuscript.
Data Accessibility
All data presented in the current manuscript can be obtained from the corresponding author.
Funding Information
This work was funded by the National Natural Science Foundation of China (grant number 81902191), the Natural Science Foundation of Liaoning Province, China (grant number 20180550926), and the Scientific Research Fund of Educational Department of Liaoning Province, China (grant number QN2019001).
Author Disclosure Statement
No competing financial interests exist.